
Abstract
Soil salinity reduces growth of Glycyrrhiza uralensis in arid and semi-arid areas of north-west in China. Silicon (Si) nutrition may alleviate salt stress in many crops including grain crop, fruit crop, and vegetable crop. In this study, the alleviating effects of Si on growth characteristics, antioxidant enzyme activity (SOD and POD) and MDA concentration, and K+ and Na+ concentrations in G. uralensis seedlings subjected to 50 mM NaCl stress were investigated. The results showed that NaCl stress imposed significant reduction in root length, secondary root number, leaf number, and stem and total dry weight of G. uralensis. NaCl stress also significantly reduced the activities of SOD and POD, and ration of K+/Na+, but significantly increased MDA concentration in leaves of G. uralensis seedling. The addition of Si increased SOD and POD activities, and reduced MDA concentration, which resulting in greater reactive oxygen species detoxification and lower lipid peroxidation. Si also significantly increased the ratio of K+/Na+ in stem and leaves of G. uralensis. In conclusion, Si could alleviate adverse effects of salt stress probably by decreasing Na+ concentration and improving antioxidant enzyme activity of G. uralensis, and these alleviating effects were dependent on Si concentration and on Si processing time.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Salt stress is one of the major abiotic factors limiting crop growth and production worldwide (Hashemi et al. 2010). It has harmful effect on approximately 7 % of the land on earth and 20 % of the total arable area (Rasool et al. 2013). And resulting in the reduction of crops production and desertification, which is attributed to two factors. First, high salinity usually causes ions imbalance, resulting in accumulation increase of Na+ and Cl− and accumulation decrease of others such as K+ and Ca2+ (Khan et al. 2000; Wang and Han 2007). High Na+ concentration causes a reduction of plant growth and over-production of reactive oxygen species (ROS) (Mahajan and Tuteja 2005). Second, plants generally subject to a water stress that also causes over-production of ROS under salinity condition (Liang 1999), which lead to metabolism disturbance (Zushi et al. 2009) and membrane lipid peroxidation intensification (Gill and Tuteja 2010). Plants have established several protective enzymatic and non-enzymatic mechanisms to scavenge ROS and mitigate its harmful effects. ROS scavenging enzymes including superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), etc. (Tiryakioglu et al. 2006; Gill and Tuteja 2010). In which SOD is one of the most important antioxidants and its enzymatic action causes the dismutation of O2 − into H2O2 and O2 (Deeba et al. 2012), in which H2O2 is further detoxified into H2O and O2 by catalase (CAT) and/or peroxidase (POD) (Seckin et al. 2010). Previous studies have shown that salt stress increases the CAT activity in maize plants (Rios-Gonzalez et al. 2002), and does not affect the SOD and POD activities in roots of salt-tolerant cultivar, while decreases the all enzyme activities in roots of salt-sensitive cultivar (de Azevedo Neto et al. 2006). SOD and CAT activities in roots of Aeluropus littoralis increased, but decreased in shoot under higher salt stress, also POD activity almost increased in all stress conditions (Modarresi et al. 2014). CAT and SOD activities in roots of Kandelia candel decreased with the increase of salt concentrations and processing time (Wang et al. 2014). These results indicated that salt stress affected the antioxidant enzyme activities, and this effect was salt level-, species- and processing time specific (Parihar et al. 2015).
Silicon (Si) makes up 31 % of the crust of the Earth and is the second mineral element in the soil (Gong et al. 2006). Si is generally considered to be a beneficial element for plant growth and development, and its uptake has a positive effect on plant growth under salt stress condition (Guntzer et al. 2012). A number of studies have shown that Si can alleviate the adverse effects of salt stress on wheat (Ahmad et al. 1992; Liang et al. 2003), barley (Liang 1999), maize (Shu and Liu 2001), rice (Yeo et al. 1999), tomato (Romero-Aranda et al. 2006), cucumber (Zhu et al. 2004) and alfalfa (Wang and Han 2007). These studies show that the application of Si increases salt tolerance of plants including grain crop, fruit crop, and vegetable crop (Zhu and Gong 2014). However, a few was known about the possible useful effects of the application of Si on medicinal plants, such as Glycyrrhiza which is classified as a non-silicon accumulator, under saline conditions (Zhang et al. 2015).
Glycyrrhiza usually grow in arid and semi-arid regions of China, Mongolia, Russia, the Middle East, and along the shores of the Mediterranean, in which Glycyrrhiza uralensis Fischer or Glycyrrhiza glabra Linne is very important for agricultural production because its root and stolon has long been used as a flavoring and natural sweetener worldwide, including Asia, Europe, and North America (Henriet et al. 2006), and crude drugs is the most frequently used in China and Japan (Ozaki and Shibano 2014). Currently, the licorice used in traditional medicines and food additives harvest from wild resources and its annually export volume in China equals to 3500 tons (Yin et al. 2014). Wild G. uralensis is well adapted to growing under unfavorable environment such as salinity and drought, which are common in the deserts and semi-deserts of northwest China (Zhang and Peng 1960). G. uralensis is widely distributed in lands with soil where salt concentrations range from 0.1 to 0.2 % (w/w), up to 0.3–0.6 % (w/w). It can also grow poorly at 1–2 % (w/w) salt environment (Tang et al. 2008). However, in recent years, irresponsible over-collection of wild G. uralensis has lead to rapidly reduction of wild resource, and even extinction in some places. What is more, the Chinese Government has, therefore, prohibited the collection of wild G. uralensis plants since 2000. As a result that cultivars have become the primary resource for supply market. Unfortunately, many studies have reported that salt stress inhibited the growth of cultivated G. uralensis above 50 mM NaCl in hydroponic experiment or 0.3 % (w/w) NaCl in soil of pot experiment. Moreover, This inhibitory effect increased at high salt concentrations compared to low salt concentrations. It was of interest, therefore, how to improve the salt tolerance of commercial cultivars of G. uralensis has become a key problem for sustainable development (Yang et al. 2006; Tang et al. 2008; Wan et al. 2011).
Accordingly, the present study aimed to evaluate the ability of Si in ameliorating salt stress of G. uralensis plant during the early seedling growth stage. In this regard, the changes of plant growth, Na+ and K+ concentration, malondialdehyde (MDA) concentration, and antioxidant enzyme activity were studied in G. uralensis seedlings under 50 mMNaCl stress supplied without or with different concentration Si.
Materials and methods
Plant material, NaCl and Si application
Seeds of G. uralensis were surface sterilized with 0.1 % H2O2 for 10 min after screening for uniform size and color, and completely washed with autoclaved deionized water. Then they were incubated in deionized water for 12 h at 25 °C. Eighty-one seeds of uniform size were sown in each of ninety germinating boxes (12 × 12 × 6 cm) padded with autoclaved sand medium and further grown till 30 days in growth chamber. The growth chamber (RXZ-430D, Ningbo Southeast Instrument Inc., P. R. China) conditions were programmed for a 12 h photoperiod at 37.5 µmol m−2 s−1 PAR (08:00–20:00; 28 °C; relative humidity 50 %) and 12 h dark (20:00–08:00; 28 °C; relative humidity 50 %) cycle. Hoagland’s nutrient solution was used as a growth medium. Treatments were arranged factorially in a randomized experimental design with 15 boxes per treatment. The treatments include (1) control (growth medium), (2) 50 mM NaCl, (3) NaCl plus 1 mM Si, (4) NaCl plus 2 mM Si, (5) NaCl plus 4 mM Si, (6) NaCl plus 6 mM Si.
Treatments were initiated at 30 days after seedling emergence. Levels of salt stress were achieved by adding NaCl in the Hoagland solution up to the level of 50 mM. Si treatments (0, 1, 2, 4 or 6 mM) were added as K2SiO3. An corresponding amount of K (as KCl) was also added to the sole NaCl and lower Si treatment of 1, 2 or 4 mM to compensate for the K concentration of K2SiO3 6 mM.
Five boxes plants of every treatment were harvested, respectively, at 10, 20 and 30 days, respectively, after starting treatment and used for the assessment of growth parameter and biochemical analyses.
Antioxidant enzyme activity and lipid peroxidation determination
Antioxidant enzyme extraction was according to the method reported by Zhang et al. (2013). In brief, each fresh 0.5 g leaf of G. uralensis seedlings was homogenized with 10 mL phosphate buffer (100 mM, pH 6.8), then the homogenate was centrifuged for 20 min at 12,000g and the liquid supernatant next was stored at 4 °C for assay of SOD and POD activity, and MDA concentration.
The activities of SOD and POD were determined and expressed as Zhang et al. (2013). Moreover, the lipid peroxidation was determined by TBA test and expressed as the amount of MDA per gram fresh weight, which was an end product of lipid peroxidation.
K+ and Na+ determination
Extraction and determination for K+ and Na+ was according to Yu et al. (2010). Generally, 0.25 g of dried powder sample was digested with sulfuric acid at 200 °C on a hot stove in glass tube, and 2 mL of H2O2 was dropwise added during the digestion. It has been considered as complete finish for digestion when only white residue was left in the glass tube. The white residue then was dissolved with distilled water to 100 mL. K+ and Na+ concentration was measured using a flame photometer (FP 640).
Statistics analysis
All experimental data were analyzed by ANOVA using SPSS 17.0 software (SPSS Inc., USA) and significant differences were tested using the least significant differences (LSD) test at P ≤ 0.05. Mean values and standard errors (SE) were presented.
Results
Plant growth parameters
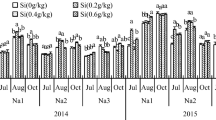
As shown in Fig. 1, the application of Si ≤2 mM increased root length of G. uralensis seedlings over the experiment period, and only at the 2 mM concentration at 10 days after treatment was there no significant difference from the 50 mM NaCl treatment sole (Fig. 1a). The application of Si at 1 mM significantly increased secondary root numbers of G. uralensis seedlings at 10 days after treatment, while Si at 6 mM significantly reduced secondary root numbers at 20 days after treatment, compared to the 50 mM NaCl treatment sole (Fig. 1b). The application of Si at 2 mM at 10 days and ≥2 mM at 20 days after treatment significantly decreased plant height of G. uralensis seedlings, while all concentrations of Si had no significant effect on plant height at 30 days after treatment, compared to the 50 mM NaCl treatment sole (Fig. 1c). The application of Si at 1 mM significantly increased leaf numbers of G. uralensis seedling at 20 or 30 days after treatment, compared to the 50 mM NaCl treatment sole (Fig. 1d).

Effect of exogenous silicon (K2SiO3) on the root length (a), secondary root number (b), plant height (c) and leaf number (d) in G. uralensis seedlings grown under salt (NaCl) stress. For each growth stage (10, 20, or 30 days), histogram bars with the same lower-case letters are not significantly different at P ≤ 0.05 by the LSD test
Biomass dry weight production and allocation
As shown in Fig. 2, at 10 days after treatment, the application of Si at 1 mM significantly increased the dry weight of root, leaf and total, compared to the 50 mM NaCl treatment sole. At 20 days after treatment, the application of Si ≤2 mM significantly increased the dry weight of root, stem, leaf, and total, compared to the 50 mM NaCl treatment sole. At 30 days after treatment, the application of Si ≤4 mM significantly increased the dry weight of root, stem, leaf, and total, compared to the 50 mM NaCl treatment sole.

Effect of exogenous silicon (K2SiO3) on the root dry weight (a), stem dry weight (b), leaf dry weight (c) and total dry weight (d) of G. uralensis seedlings grown under salt (NaCl) stress. For each growth stage (10, 20, or 30 days), histogram bars with the same lower-case letters are not significantly different at P ≤ 0.05 by the LSD test
Antioxidant enzymes and lipid peroxidation
Activities of SOD and POD in G. uralensis seedlings were significantly decreased after NaCl exposure, compared to plants grown without NaCl. However, SOD activity was significantly intensified with Si ≤4 mM over the experiment period, compared to plants grown without Si (Fig. 3a). POD activity was significantly intensified only with Si at 2 mM at 10 days after treatment, and with Si at 6 mM at 30 days after treatment, compared to plants grown without Si. At 20 days after treatment, however, the POD activity was significantly increased with Si at all treated concentrations (Fig. 3b).

Effect of exogenous silicon (K2SiO3) on the activities of superoxide dismutase (SOD) and peroxidase (POD), and the content of malondialdehyde (MDA) in seedlings of G. uralensis grown under salt (NaCl) stress. For each growth stage (10, 20, or 30 days), histogram bars with the same lower-case letters are not significantly different at P ≤ 0.05 by the LSD test
MDA concentration in seedlings significantly increased under NaCl stress at 20 or 30 days after treatment, compared to the control. However, the MDA concentration significantly decreased with Si application at all treated concentrations at 20 or 30 days after treatment, compared to the 50 mM NaCl treatment sole (Fig. 3c).
Mineral contents (K+, Na+ and K+/Na+)
K+ concentration in stem of G. uralensis seedling was significantly increased when Si ≥2 mM concentration at 10 days after treatment, compared to the 50 mM NaCl treatment sole. K+ concentration in stem was significantly increased when Si at all treated concentrations at 20 days after treatment (Fig. 4a). As leaf tissue, K+ concentration was significantly increased when Si at 6 mM concentration at 10 days after treatment, and at 2 or 6 mM concentrations at 20 days after treatment, compared to the 50 mM NaCl treatment sole (Fig. 4b).

Effect of exogenous silicon (K2SiO3) on the content of K+ in stem and leaf (a, b), Na+ in stem and leaf (c, d), and the K+/Na+ ratio in stem and leaf (e, f) of G. uralensis seedlings grown under salt (NaCl) stress. For each growth stage (10, 20, or 30 days), histogram bars with the same lower-case letters are not significantly different at P ≤ 0.05 by the LSD test
Na+ concentration in stem of G. uralensis seedling was pronounced decreased when Si ≤4 mM concentrations at 10 days after treatment, and ≥2 mM concentrations at 30 days after treatment, compared to the 50 mM NaCl treatment (Fig. 4c). As leaf tissue, Na+ concentration was pronounced decreased when Si at 2 mM concentration at 10 days after treatment, and ≥2 mM concentrations at 30 days after treatment, compared to the 50 mM NaCl treatment sole (Fig. 4d).
K+/Na+ ratio in stem of G. uralensis seedling was significantly increased when Si at all treated concentrations over the experiment period, compared to the 50 mM NaCl treatment sole (Fig. 4e). As leaf tissue, K+/Na+ ratio was significantly increased when Si at 2 mM or higher concentrations over the experiment period, compared to the 50 mM NaCl treatment sole (Fig. 4f).
Discussion
Harmful effects of salt stress on plant manifest through alter the growth of roots and shoots, which may invariably result in a reduced plant growth. Chen et al. (2008) report that as the concentrations of NaCl increased, the plant height, leaf area and biomass accumulation of two Epimedium species decreased. At present study, root length, leaf number of G. uralensis was significantly decreased, which possibly lead to the low biomass of plant under NaCl stress (Figs. 1, 2). However, this study suggested that Si supplied may be beneficial to offset this harmful effect of NaCl stress on G. uralensis growth as reported previously in Prosopis juliflora (Bradbury and Ahmad 1990), barley (Liang 1999), rice (Gong et al. 2006), tomato (Romero-Aranda et al. 2006), wheat (Tuna et al. 2008), Glycine max L. (Lee et al. 2010) and Saccharum officinarum L. (Ashraf et al. 2010a, b).
NaCl treatment significantly impaired G. uralensis growth by increasing the concentration of Na+ and reducing the concentration of K+ in stem and leaf (Fig. 4). These changes are consistent with previous reports (Guerrier 1996; Khan et al. 2000; Wang and Han 2007), some of which indicated that high Na+ concentration in plant cells changes cellular metabolism, resulting in reducing plant growth and overproduction of ROS (Mahajan and Tuteja 2005). Application of Si greatly alleviated biomass and Na+ accumulation in the shoots of G. uralensis due to NaCl-induced reduction in this study. Liang and Ding (2002) reported that Si addition decreases Na+ and Cl− levels while increases K+, which encourage Na+ and K+ to more evenly distribute in root of barley under salt-stressed. Similarly, Si addition decreases Na+ content in roots of alfalfa (Medicago sativa L.) rather than shoots, and notably increases K+ content in the shoots of alfalfa (Wang and Han 2007). What is more, Si addition dramatically decreases the Na+ concentration in shoots of rice plants rather than roots when they grow in salt stress environment, which correlates with improved shoot growth in the presence of Si (Gong et al. 2006). These results suggest that Si addition is beneficial to reduce ion toxicity in plants under salt stress environment by decreasing toxic ion accumulation and this may be one of the mechanisms whereby Si enhanced salt tolerance in these species.
Salinity induced a water deficit as well as an ionic toxicity in the plants resulting in an alteration in the ionic homeostasis. In addition to the osmotic and toxic effects, salt stress is also considered as an oxidative stress, contributing all these factors to the deleterious effects of salinity in plants (Barba-Espín et al. 2011; Acosta-Motos et al. 2015). The oxidative stress induced by salt stress is closely related to production of highly toxic reactive oxygen species (ROS). Generally, oxidative stress increase the production of superoxide radical (O2 −), hydroxyl radical (HO−), hydrogen peroxide (H2O2), and singlet oxygen (1O2) in plants (Abogadallah 2010) by altering the process of photosynthesis, photorespiration, fatty acid oxidation, and senescence, which cause the nonspecific reactions of lipids, proteins, and nucleic acids (Davies 1995). Therefore, imbalance of oxidative stress leads to plant death by significantly damaging the structural integrity of cell membrane. Plant suffering from an oxidative damage could activate the antioxidant enzymes to scavenge the detrimental ROS groups by catalyzing the primary reaction, specifically, superoxide dismutase (SOD) catalyzes the dismutation of O2 − into H2O2 and O2, in which H2O2 is also toxic to plant cells and it is further detoxified into H2O and O2 by catalase (CAT) and/or peroxidase (POD), guaiacol peroxidase (GPX) and ascorbate peroxidase (APX) (Soundararajan et al. 2015). In this study, only NaCl treatment decreases the SOD and POD activities. Addition of Si significantly increases the SOD activity during the experiment when the Si concentration is or below 4 mM in the NaCl medium (Fig. 3a), and these results also are observed in tomato (Al-aghabary et al. 2005), cucumber (Zhu et al. 2004), maize (Moussa 2006) and Dianthus caryophyllus (Soundararajan et al. 2015) under salt stress. The SOD catalyzes the dismutation of O2 − into H2O2 and O2, resulting from chloroplast, mitochondria, and peroxisome (Choudhury et al. 2013). With the increase of SOD activities, more O2 − is effectively converted into H2O2 and further into H2O by POD, APX, and CAT detoxifications. Accordingly, we understood that by increasing SOD activity, the activity of POD also increased and so it means, there are fully correlation between these two enzyme activities against salinity stress. SOD and POD have same pattern changes activity, because peroxidase is involved in scavenging H2O2 resulting from O2 − catalyzed by SOD. Moreover, antioxidant enzymes activity is not significantly correlated with different Si treatments. However, addition of Si does not affect the SOD activity of salt-stressed grapevine, and decreases the POD activity, whereas the APX activity is dependent on cultivars (Soylemezoglu et al. 2009). A number of studies have shown that the plant resistance to salt stress was commonly related to efficient antioxidant system (Kim et al. 2005), in which, ROS is very important for normal biological processes, especially in signaling and transportation. For example, low level of ROS concentrations are necessary for plant signaling process (Rouhier and Jacquot 2008), while high level of ROS concentrations are harmful to the plant cells (Choudhury et al. 2013). Therefore, a conclusion is drawn that excess ROS detoxified by Si is involved in adjusting activities of antioxidant enzymes (Fig. 3a, b). Therefore, the results of this study demonstrated that the addition of Si scavenges the excess ROS in G. uralensis under salt-stressed condition. Accordingly, addition of Si at all concentrations deceased the MDA content. This suggests that addition of Si regulates the activities of antioxidant enzymes, and resulting in decrease of oxidative stress in salt-stressed G. uralensis seedlings.
As a general conclusion, NaCl stress decrease plant growth through the accumulation of Na+ ions and the decline of antioxidant defenses, and further inducing membrane damage, as indicated by the MDA accumulation. However, the addition of Si ≤2 mM concentration significantly reverses these trends. These results suggest that Si application ≤2 mM concentration can improve salt stress tolerance in G. uralensis. Notably, this improve effect was time dependent and Si concentration dependent.
Author contribution statement
Conceived and designed the experiments: Yue-Tong Li, Wen-Jin Zhang, Xin-Hui Zhang. Performed the experiments: Yue-Tong Li, Wen-Jin Zhang, Jia-Jia Cui, Duo-Yong Lang. Analyzed the data: Yue-Tong Li, Wen-Jin Zhang, Jia-Jia Cui, Qi-Peng Zhao, Xin-Hui Zhang Contributed reagents/materials/analysis tools: Ming Li, Xin-Hui Zhang Wrote the paper: Yue-Tong Li, Wen-Jin Zhang, Xin-Hui Zhang.
References
Abogadallah GM (2010) Antioxidative defense under salt stress. Plant Signal Behav 5:369–374
Acosta-Motos JR, Diaz-Vivancos P, Álvarez S, Fernández-García N, Sanchez-Blanco MJ, Hernández JA (2015) Physiological and biochemical mechanisms of the ornamental Eugenia myrtifolia L. plants for coping with NaCl stress and recovery. Planta 242:829–846
Ahmad R, Zaheer SH, Ismail S (1992) Role of silicon in salt tolerance of wheat (Triticum aestivum L.). Plant Sci 85:43–50
Al-aghabary K, Zhu Z, Shi Q (2005) Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J Plant Nutr 27:2101–2115
Ashraf M, Afzal M, Ahmed R, Mujeeb F, Sarwar A, Ali L (2010a) Alleviation of detrimental effects of NaCl by silicon nutrition in salt-sensitive and salt-tolerant genotypes of sugarcane (Saccharum officinarum L.). Plant Soil 326:381–391
Ashraf M, Ahmad R, Bhatti AS, Afzal M, Sarwar A, Maqsood MA, Kanwal S (2010b) Amelioration of salt stress in sugarcane (Saccharum officinarum L.) by supplying potassium and silicon in hydroponics. Pedosphere 20:153–162
Barba-Espín G, Clemente-Moreno MJ, Alvarez S, García-Legaz MF, Hernandez JA, Díaz-Vivancos P (2011) Salicylic acid negatively affects the response to salt stress in pea plants. Plant Biol 13:909–917
Bradbury M, Ahmad R (1990) The effect of silicon on the growth of Prosopis juliflora growing in saline soil. Plant Soil 125:71–74
Chen GD, Li YX, Zhang H, Chen J, Cai LY (2008) Effects of salt stress on growth and flavonoids content in different organs of two species of Epimedium. Acta Bot Boreal-Occident Sin 28:2047–2054
Choudhury S, Panda P, Sahoo L, Panda SK (2013) Reactive oxygen species signaling in plants under abiotic stress. Plant Signal Behav 8:e23681
Davies KJ (1995) Oxidative stress: the paradox of aerobic life. Biochem Soc Symp 61:1–31
de Azevedo Neto AD, Prisco JT, Eneas J, de Abreu CEB, Gomes-Filho E (2006) Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt sensitive maize varieties. Environ Exp Bot 56:87–94
Deeba F, Pandey AK, Ranjan S, Mishra A, Singh R, Sharma YK, Shirke PA, Pandey V (2012) Physiological and proteomic responses of cotton (Gossypium herbaceum L.) to drought stress. Plant Physiol Biochem 53:6–18
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gong HJ, Randall DP, Flowers TJ (2006) Silicon deposition in root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ 29:1970–1979
Guerrier G (1996) Fluxes of Na+, K+ and Cl−, and osmotic adjustment in Lycopersicon pimpinellifolium and L. esculentum during short- and long-term exposures to NaCl. Physiol Plant 97:583–591
Guntzer F, Keller C, Meunier JD (2012) Benefits of plant silicon for crops: a review. Agron Sustain Dev 32:201–213
Hashemi A, Abdolzadeh A, Sadeghipour HR (2010) Beneficial effects of silicon nutrition in alleviating salinity stress in hydroponically grown canola, Brassica napus L. plants. Soil Sci Plant Nutr 56:244–253
Henriet C, Draye X, Oppitz I, Swenen R, Delvaux B (2006) Effects, distribution and uptake of silicon in banana (Musa spp.) under controlled conditions. Plant Soil 287:359–374
Khan MA, Ungar IA, Showalter AM (2000) Effects of sodium chloride treatments on growth and ion accumulation of the halophyte Haloxylon recurvum. Commun Soil Sci Plan 31:2763–2774
Kim SY, Lim JH, Park MR, Kim YJ, Park TI, Seo YW, Choi KG, Yun SJ (2005) Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J Biochem Mol Biol 38:218–224
Lee SK, Sohn EY, Hamayun M, Yoon JY, Lee JJ (2010) Effect of silicon on growth and salinity stress of soybean plant grown under hydroponic system. Agrofor Syst 80:333–340
Liang YC (1999) Effects of silicon on enzyme activity, and sodium, potassium and calcium concentration in barley under salt stress. Plant Soil 209:217–224
Liang YC, Ding RX (2002) Influence of silicon on microdistribution of mineral ions in roots of salt-stressed barley as associated with salt tolerance in plants. Sci China Ser C 45:298–308
Liang YC, Chen Q, Liu Q, Zhang WH, Ding RX (2003) Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J Plant Physiol 160:1157–1164
Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses: an overview. Arch Biochem Biophys 444:139–158
Modarresi M, Moranlan F, Nematzadeh GL (2014) Antioxidant responses of halophyte plant Aeluropus littoralis under long-term salinity stress. Biologia 69:478–483
Moussa HR (2006) Influence of exogenous application of silicon on physiological response of salt-stressed maize (Zea mays L.). Int J Agric Biol 8:293–297
Ozaki K, Shibano M (2014) Aim for production of Glycyrrhizae Radix in Japan (3): development of a new licorice cultivar. J Nat Med 68:358–362
Parihar P, Singh S, Singh R, Singh VP, Prasad SM (2015) Effect of salinity stress on plants and its tolerance strategies: a review. Environ Sci Pollut Res 22:4056–4075
Rasool S, Hameed A, Azooz MM, Muneebu-rehman Siddiqi TO, Ahmad P (2013) Salt stress: causes, types and responses of plants. In: Ahmad P, Azooz MM, Prasad MNV (eds) Ecophysiology and responses of plants under salt stress. Springer, New York, pp 1–24
Rios-Gonzalez K, Erdei L, Lips SH (2002) The activity of antioxidant enzymes in maize and sunflower seedlings as affected by salinity and different nitrogen sources. Plant Sci 162:923–930
Romero-Aranda MR, Jurado O, Cuartero J (2006) Silicon alleviates the deleterious salt effect on tomato plant growth by improving plant water status. J Plant Physiol 163:847–855
Rouhier N, Jacquot JP (2008) Getting sick may help plants overcome abiotic stress. New Phytol 180:738–741
Seckin B, Turkan I, Sekmen AH, Ozfidan C (2010) The role of antioxidant defense systems at differential salt tolerance of Hordeum marinum Huds. (sea barleygrass) and Hordeum vulgare L. (cultivated barley). Environ Exp Bot 69:76–85
Shu LZ, Liu YH (2001) Effects of silicon on growth of maize seedlings under salt stress. Agro-Environ Prot 20:38–40
Soundararajan P, Manivannan A, Park YG, Muneer S, Jeong BR (2015) Silicon alleviates salt stress by modulating antioxidant enzyme activities in Dianthus caryophyllus ‘Tula’. Hortic Environ Biotechnol 56:233–239
Soylemezoglu G, Demr K, Inal A, Gunes A (2009) Effect of silicon on antioxidant and stomatal response of two grapevine (Vitis vinifera L.) rootstocks grown in boron toxic, saline and boron toxic-saline soil. Sci Hortic-Amsterdam 123:240–246
Tang XM, Wang WQ, Yang Q, Liu CL (2008) Effect of NaCl treatment on growth, physiological index and content of effective composition of Glycyrrhiza uralensis. J Jilin Agric Univ 30:172–175 (in Chinese)
Tiryakioglu M, Eker S, Ozkutlu F, Husted S, Cakmak I (2006) Antioxidant defense system and cadmium uptake in barley genotypes differing in cadmium tolerance. J Trace Elem Med Biol 20:181–189
Tuna AL, Kaya C, Higgs D, Murillo-amador B, Demir Girgin AR (2008) Silicon improves salinity tolerance in wheat plants. Environ Exp Bot 62:10–16
Wan CY, Wang D, Hou JL, Wang WQ, Peng F (2011) Effects of NaCl stress on growth and antioxidant enzyme activities of Glycyrrhiza uralensis. Prog in Mod Biomed 11:1805–1809 (in Chinese)
Wang XS, Han JG (2007) Effects of NaCl and silicon on ion distribution in the roots, shoots and leaves of two alfalfa cultivars with different salt tolerance. Soil Sci Plant Nutr 53:278–285
Wang HM, Xiao XR, Yang MY, Gao ZL, Zang J, Fu XM, Chen YH (2014) Effects of salt stress on antioxidant defense system in the root of Kandelia candel. Bot Stud 55:57–63
Yang XU, Li JM, Dong XU, Duan LS, Li ZH (2006) Effects of salt stress on growth and some physiological indexes in Glycyrrhiza uralensis Fisch seedlings. Acta Agric Boreali-Occident Sin 21:39–42 (in Chinese)
Yeo AR, Flowers SA, Rao G, Welfare K, Senanayke N, Flowers JF (1999) Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in the transpirational bypass flow. Plant Cell Environ 22:559–565
Yin SS, Zhang Y, Gao WY, Wang J, Man SL, Liu H (2014) Effects of nitrogen source and phosphate concentration on biomass and metabolites in adventitious root culture of Glycyrrhiza uralesis Fisch. Acta Physiol Plant 36:915–921
Yu BJ, Luo QY, Liu YL (2010) Effects of salt stress on growth and ionic distribution of salt-born Glycine soja. Acta Agro Sin 27:776–780 (in Chinese)
Zhang PY, Peng ZX (1960) Liquorice in Northwest of China. J Lanzhou Univ 1:57–87
Zhang XH, Lang DY, Zhang EH, Bai CC, Wang HZ (2013) Diurnal changes in photosynthesis and antioxidants of Angelica sinensis as influenced by cropping systems. Photosynthetica 51:252–258
Zhang XH, Zhou D, Cui JJ, Ma HL, Lang DY, Wu XL, Wang ZS, Qiu HY, Li M (2015) Effect of silicon on seed germination and the physiological characteristics of Glycyrrhiza uralensis under different levels of salinity. J Hortic Sci Biotech 90:439–443
Zhu YX, Gong HJ (2014) Beneficial effects of silicon on salt and drought tolerance in plants. Agron Sustain Dev 4:455–472
Zhu JK, Wei GQ, Li J, Qian QQ, Yu JQ (2004) Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci 167:527–533
Zushi K, Matsuzoe N, Kitano M (2009) Developmental and tissuespecific changes in oxidative parameters and antioxidant systems in tomato fruits grown under salt stress. Sci Hortic 122:362–368
Acknowledgments
This study was supported by the National Natural Science Foundation of China (Project Nos. 31260304 and 31460330).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by LA Kleczkowski.
Y.-T. Li and W.-J. Zhang contributed to the manuscript equally.
Rights and permissions
About this article

Cite this article
Li, YT., Zhang, WJ., Cui, JJ. et al. Silicon nutrition alleviates the lipid peroxidation and ion imbalance of Glycyrrhiza uralensis seedlings under salt stress. Acta Physiol Plant 38, 96 (2016). https://doi.org/10.1007/s11738-016-2108-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-016-2108-8