
Abstract
Background
It is a common dogma that gastric bypass (GB) induces early satiety and consequent reductions in food intake and nutrient absorption. The aim of the present study was to analyze feeding behavioral and metabolic changes in rats after GB.
Methods
Male Sprague–Dawley rats at the ages of 23 and 42 weeks were placed in metabolic cages connected with a comprehensive laboratory animal monitoring system. At the age of 48 weeks they were subjected to either GB or sham operation, and then placed in metabolic cages at 51 and 62 weeks (or 3 and 14 weeks postoperatively).
Results
GB rats lost 20% of the body weight within 2–3 weeks and remained at this lower level until the end of the study at 14 weeks postoperatively. Satiety ratio was higher during daytime than nighttime in both sham-operated and GB rats, but was not significantly different between the two groups. Neither daily accumulated food intake nor food intake per 100 g of body weight was different between sham-operated and GB rats. Apparently, GB rats ate more frequently during daytime and had smaller meal size during nighttime at 3 weeks postoperatively. These changes were not present at 14 weeks postoperatively. Energy density in the feces was the same in GB and sham-operated rats postoperatively. Energy expenditure declined with age, but increased in GB rats compared with age-matched sham-operated rats.
Conclusions
GB reduced the body weight without causing early satiety, reducing food intake or inducing malabsorption. It did, however, increase energy expenditure.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Overweight, obesity, and their comorbidities have become a major health concern in developed countries during the last decade. Reducing food intake by various behavioral and pharmacological therapeutic strategies has shown limited success in maintaining significant body weight control. Surgical intervention, e.g., gastric bypass (GB), has become a standard treatment for morbid obesity [1–5]. The efficacy of GB has been established mainly based on the data from case series rather than on prospective randomized trials [6]. It is a common dogma that GB would create a small pouch of the stomach to produce early satiety and a consequent reduction in food intake, and moreover induce malabsorption by creating a short gut syndrome and/or by accomplishing distal mixing of bile and pancreatic juice with ingested nutrients thereby reducing absorption. Although a great number of patients are undergoing GB, little data are available regarding their food intake after the surgery. It is well-known that in most patients, GB often leads to changes in eating behavior to avoid unpleasant sensations/symptoms, such as abdominal discomfort, nausea, or vomiting. However, it is difficult to measure the precise food/caloric intake, meal proportions, and total energy expenditure without keeping patients under 24-h observation.
Studies using various rat models have shown that mechanical factors and changes in gastrointestinal and adipose hormones, as well as hypothalamic neuropeptides may contribute to GB-induced weight loss by promoting catabolic processes [7–11]. As the rats are nocturnal animals, their characteristic pattern of consuming food during the day or night provides a useful model for human ad libitum eating. The aim of the present study was to analyze in detail the feeding behavior in rats after GB by utilizing a comprehensive laboratory animal metabolic monitoring system.
Materials and Methods
Animals and Experimental Design
Twenty male Sprague–Dawley rats were obtained from Taconic M&B, Skensved, Denmark for this study. Male rats were chosen because female rats have been shown to have food intake coinciding with ovarian hormones; low intake when estrogen levels are high (ovulation), and high intake when estrogen levels are low [12]. Moreover, male rats grow faster than females, making it easier to detect changes in body weight. The animals were housed in Makrolon cages at 22°C, 50–60% humidity, and 12 h light/dark cycle. They had free access to commercial standard rat food pellets and powder form (RM1, Scanbur BK AS, Sweden) and tap water ad libitum. At the age of 48 weeks (body weight of ∼ 600 g) the animals were subjected to either GB or sham operation (as controls) under anesthesia with isoflurane. GB was performed in 14 rats by anastomosing the esophagus to the proximal jejunum about 2–3 cm distal to the Treitz ligament in an end-to-side fashion [8]. Laparotomy with gentle manipulation of the viscera was performed as sham operation in six rats. Six out of 14 animals died or were terminated after GB during the experimental period. Out of these six animals, one died because of peroperative complications. Another one was terminated after 2 weeks because of an extensive weight loss and the autopsy showed a constricted anastomosis. Yet another one was put to sleep at the same time point because of salivation caused by an obstructive hair and food bolus in the anastomosis. Three weeks postoperatively, two rats died without prior noticeable symptoms, and autopsies showed signs of gastrointestinal infection with hemorrhagic diarrhea in the perianal area. The sixth animal was put down 5 weeks postoperatively because of respiratory problems, and the autopsy revealed signs of lung inflammation. After an initial drop in body weight because of the surgical trauma, the remaining rats started to increase their body weight about 2–3 weeks postoperatively and kept this lower level until the end of the experiments. Ambulatory activity was similar between the sham-operated and the surviving GB rats; thereby indicating that GB rats recovered well from the surgery.
The experiment was approved by the Norwegian National Animal Research Authority (Forsøksdyrutvalget, FDU).
Recording of Body Weight, Food and Water Intake, as Well as Eating and Drinking Behavior
The body weight was recorded throughout the observed period. Rats were gradually acclimatized to the metabolic measuring system cages during a period of 2 weeks. From the age of 23 weeks the rats were subjected to indirect calorimetry with The Comprehensive Laboratory Animal Monitoring Systems (CLAMS) (Oxymax; Columbus Instruments International, Columbus, OH, USA). The first measurements were performed 3 weeks postoperatively to allow the rats to recover properly from surgery. This monitoring system is composed of a four-chamber indirect calorimeter designed for the continuous monitoring of up to four rats simultaneously, recording data on food and water intake, eating and drinking behavior, and VO2 and VCO2 from each rat in each chamber. An air sample was withdrawn every 7 min from each cage. Energy expenditure (kcal/h) was calculated from the following equation: (3.815 + 1.232RER) × VO2, where RER was the respiratory exchange ratio (volume of CO2 produced per volume of O2 consumed, both ml kg−1 hr−1) and VO2 was the volume of O2 consumed per hour and kilogram animal mass. Urine production was automatically recorded by balances. The rats were placed in calorimeter chambers for 48 h (data from the last day- and night-time were used in the analysis) with access to their normal diet in ground-up form, and tap water ad libitum. The high-resolution feeding data are generated by monitoring all feed balances every 0.5 s and provides accumulated food intake, meal size, and duration. In our program, the end of a feeding event (meal) is when the balance has been stable for more than 10 s and a minimum of 0.05 g has been eaten. Parameters in both daytime and nighttime (12 h in each) for each rat include the number of meals, the average size, the average duration, accumulated food intake, the average intermeal interval, rate of eating, and the satiety ratio. The intermeal interval was defined as the interval in minutes between two meals. The rates of eating were calculated by dividing the average meal size with the average duration of a meal. Finally, satiety ratio, an index of the no-eating (i.e., satiety) time in relation to grams of food consumed, was calculated as the average intermeal interval divided by the average meal size for food [13]. The drinking behavior was collected from each interval, giving the water intake during the interval and the accumulated water intake throughout the experiment. From this application, the parameters, i.e., water intake, number of intervals with drinking activity, and average water intake within an interval, are provided.
Determination of Energy Content in Feces
Feces were collected while the rats were in CLAMS cages and dried for 72 h at 60°C. The energy density was determined by means of an adiabatic bomb calorimeter (IKA-Calorimeter C 5000, IKA-Werke GmbH & Co. KG, Staufen, Germany).
Statistical Analysis
The results are given as the mean ± SEM. Independent Student’s t test or the Mann-Whitney U/Wilcoxon rank-sum test was performed in SPSS version 13.0 for Windows (SPSS Inc., Chicago, IL, USA). A P value of <0.05 was considered statistically significant.
Results
Body Weight
The rats gained weight continuously until the age of 48 weeks when they were subjected either to sham operation or GB. GB rats lost 20% of their body weight within 2–3 weeks postoperatively, as compared to age-matched sham-operated controls or body weight before surgery. They remained at this lower body weight level until the end of the study at 14 weeks after the surgery (Fig. 1).

Body weight gain from age 10 to 62 weeks. The animals were subjected to surgery (i.e., sham operation or gastric bypass, GB) at age 48 weeks. Data are expressed as mean ± SEM. **P < 0.01 between sham-operated and GB rats
Satiety, Food Intake, and Meal Pattern
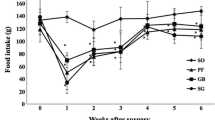
Satiety ratio was higher during daytime than nighttime in both sham-operated and GB rats, but there were no significant differences between these two groups (Fig. 2). Daily accumulated food intake in terms of gram per 24 h per rat or gram per 24 h per 100 g body weight was not different between sham-operated and GB rats (Fig. 3). Both sham-operated and GB rats ate more often during nighttime than daytime, and apparently GB rats ate more frequently and had increased food intake during daytime compared with sham-operated rats at 3 weeks postoperatively (week 51) (Figs. 4, 5). At the same time point, GB rats ate smaller meal size during nighttime (Fig. 6). These changes induced by GB were not present at 14 weeks postoperatively.

Satiety ratio expressed as intermeal interval (min/meal size in gram) during daytime and nighttime in rats before and after sham operation or gastric bypass. The measurements were performed at ages 23, 42, 51, and 62 weeks. The surgeries were done at age 48 weeks, meaning that age 51 or 62 weeks corresponded to 3 or 14 weeks postsurgery, respectively. Data are expressed as mean ± SEM; ns, not significant between the two groups of rats

Accumulated food intake expressed as gram per 24 h per rat (A) and as gram per 24 h per 100 g body weight); (B) in rats before and after sham operation or gastric bypass. The measurements were performed at ages 23, 42, 51, and 62 weeks. The surgeries were done at age 48 weeks, meaning that age 51 or 62 weeks corresponded to 3 or 14 weeks postsurgery, respectively. Data are expressed as mean ± SEM; ns, not significant between the two groups of rats

Feeding frequency expressed as the number of meals during daytime and nighttime in rats before and after sham operation or gastric bypass. The measurements were performed at ages 23, 42, 51, and 62 weeks. The surgeries were done at age 48 weeks, meaning that age 51 or 62 weeks corresponded to 3 or 14 weeks postsurgery, respectively. Data are expressed as mean ± SEM. **P < 0.01 (two-tailed); ns, not significant between the two groups of rats

Food intake expressed as gram/rat during daytime and nighttime in rats before and after sham operation or gastric bypass. The measurements were performed at ages 23, 42, 51, and 62 weeks. The surgeries were done at age 48 weeks, meaning that age 51 or 62 weeks corresponded to 3 or 14 weeks postsurgery, respectively. Data are expressed as mean ± SEM. *P < 0.05 (two-tailed); ns, not significant between the two groups of rats

Meal size in gram during daytime and nighttime in rats before and after sham operation or gastric bypass. The measurements were performed at ages 23, 42, 51, and 62 weeks. The surgeries were done at age 48 weeks, meaning that age 51 or 62 weeks corresponded to 3 or 14 weeks postsurgery, respectively. Data are expressed as mean ± SEM. *P < 0.05 (two-tailed); ns, not significant between the two groups of rats
Drinking pattern was the same as the feeding pattern, except that average water intake during one interval at nighttime was reduced in GB rats (Tables 1, 2). In addition, total urine production was reduced in GB rats at 3 and 14 weeks (Tables 1, 2).
Energy Expenditure and Balance
Energy expenditure in terms of kcal per hour per 100 g body weight declined with age in control animals (before and after sham operation). Energy expenditure significantly increased in GB rats compared with sham-operated rats (Fig. 7). RER was unchanged 3 weeks after GB surgery, but was slightly decreased at 14 weeks postoperatively (Tables 1, 2). Ambulatory activity was unaffected after GB (Tables 1, 2). Energy density in the feces was the same in GB as sham-operated rats 3 weeks after surgery (Table 1) (not measured at 14 weeks postoperatively).

Energy expenditure expressed as kcal per hour per 100 g body weigh in rats before and after sham operation or gastric bypass. The measurements were performed at ages 23, 42, 51, and 62 weeks. The surgeries were done at age 48 weeks, meaning that age 51 or 62 weeks corresponded to 3 or 14 weeks postsurgery, respectively. Data are expressed as mean ± SEM. **P < 0.01 (two-tailed); ns, not significant between the two groups of rats
Discussion
It has been believed for decades that there is a direct link between food intake and body weight. Countless studies have been conducted with an attempt to understand the complex central and peripheral networks that regulate food intake to identify potential targets for effective anti-obesity therapy. One would intuitively expect that restriction of food intake will achieve body weight loss. Unfortunately, it has not been the case in the long-term for overweight people and for most patients suffering from obesity.
Since the first operations were introduced in the 1950s, GB has been demonstrated to efficiently cause the long-term body weight loss, but the mechanisms of its action have been oversimplified [6]. In fact, the capacity of the stomach was normal in obese people before the surgery [14]. After the surgery, food or caloric intake has been shown to vary. It was reported that total caloric intake was reduced after gastric bypass (Roux-en-Y procedure, RYGB), but protein, carbohydrate, and fat intake was not or little reduced [15]. Thus, there is a need for clarifying the relationship between food intake and long-term weight changes in patients after RYGB [16]. Recently, we have shown that the control of food intake and meal size are independent on the food reservoir function of the stomach by studying rat models of gastrectomy and GB [8, 9]. We have also shown that gastric ghrelin is unlikely responsible for the long-term loss of body weight after GB by 1 year of following up the circulating ghrelin levels and the ghrelin-producing cells in the stomach [8, 17]. In the present study, we further show that GB reduced the body weight without causing early satiety, reducing food intake or inducing malabsorption.
It is known that rats gained body weight after 32 weeks of age mainly as a result of expansion of the fat compartment, and that GB impaired the growth of young rats and reduced the fat mass (but not fat-free mass) in adult rats [8]. In the present study, we found that body weight gain was associated with decreased energy expenditure along with aging in normal rats, and interestingly that GB increased the energy expenditure. Thus, we suggest that the mechanism underlying GB-induced body weight loss is unlikely through actions on food intake and/or absorption, and moreover, that the anti-obesity therapeutic strategies should be more focused on increasing energy expenditure rather than reducing food intake. Indeed, drugs that aim to reduce body weight by increasing energy expenditure have been explored. For instance, ß3-adrenergic receptor antagonist (KRP-204) is at phase 2 trials [18].
GB rats did show meal behavioral changes in the short term, but not long-term. They ate and drank more during the daytime than their controls and were unable to keep up with their controls during the nighttime with respect to meal size and eating rate. These feeding habit changes after GB more likely reflect adaptation to the surgery. In general, GB rats had parallel changes in feeding and drinking. For comparison, we found previously that total gastrectomy altered the relationship between feeding and drinking, which was assumed because of depletion of gastric hormones (e.g., ghrelin or obestatin) [9]. In fact, we have shown that GB reduced the circulating levels of ghrelin (by about 40%) only at 3 months postoperatively, but not afterwards, and it reduced moderately the circulating levels of obestatin [8].
Based on our previous [8, 9, 17] and present studies, we challenge the paradigm that GB results in weight loss because of deceased food intake. We also realize that it raises as many questions as it answers, and that the precise relevance of this study to human patients needs to be further studied. In fact, it has been well recognized that observations in animal models make significant contributions to the understanding of the physiological mechanisms in humans. It is obviously important to study the mechanisms by which GB achieves superior body weight loss. As these mechanisms are identified, the basis for understanding the etiology of obesity will be greatly improved and pharmacological interventions that mimic the physiological effects of GB may be developed [6].
References
Bray GA, Tartaglia LA. Medicinal strategies in the treatment of morbid obesity. Nature. 2000;404:672–7.
Mun EC, Blackburn GL, Matthews JB. Current status of medical and surgical therapy for obesity. Gastroenterology. 2001;120:669–81.
Solomon C, Dluhy RG. Bariatric surgery—Quick fix or long-term solution? N Engl J Med. 2004;351:2751–3.
Wolfe BM, Austrheim-Smith IT, Ghaderi N. Surgical treatment of obesity: pyloric electrical stimulation. Gastroenterology. 2005;128:225–8.
O’Brien PE, McPhail T, Chaston TB, et al. Systematic review of medium-term weight loss after bariatric operations. Obes Surg. 2006;16:1032–40.
Elder KA, Wolfe BM. Bariatric surgery: a review of procedures and outcomes. Gastroenterology. 2007;132:2253–71.
Kirchner H, Guijarro A, Meguid MM. Is a model useful in exploring the catabolic mechanisms of weight loss after gastric bypass in humans? Curr Opin Clin Nutr Metab Care. 2007;10:463–74.
Stenström B, Furnes MW, Tømmerås K, et al. Mechanism of gastric bypass-induced body weight loss: one-year follow-up after micro-gastric bypass in rats. J Gastrointest Surg. 2006;10:1384–91.
Furnes MW, Stenström B, Tømmerås K, et al. Feeding behaviour in rats subjected to gastrectomy or gastric bypass surgery. Eur Surg Res. 2007 (in press).
Egecioglu E, Stenström B, Pinnock SB, et al. Hypothalamic gene expression following ghrelin therapy to gastrectomized rodents. Regul Pept. 2007 [Epub ahead of print].
Guijarro A, Suzuki S, Chen C, et al. Characterization of weight loss and weight regain mechanisms after Roux-en-Y gastric bypass in rats. Am J Physiol Regul Integr Comp Physiol. 2007;293:R1474–89.
Varma M, Chai JK, Meguid MM, et al. Effect of estradiol and progesterone on daily rhythm in food intake and feeding patterns in Fischer rats. Physiol Behav. 1999;68:99–107.
Zorrilla EP, Inoue K, Fekete EM, et al. Measuring meals: structure of prandial food and water intake of rats. Am J Physiol Regul Integr Comp Physiol. 2005;288:R1450–67.
Camilleri M, Grudell AB. Appetite and obesity: a gastroenterologist’s perspective. Neurogastroenterol Motil. 2007;19:333–41.
Brolin RL, Robertson LB, Kenler HA, et al. Weight loss and dietary intake after vertical banded gastroplasty and Roux-en-Y gastric bypass. Ann Surg. 1994;220:782–90.
Warde-Kamar J, Rogers M, Flancbaum L, et al. Calorie intake and meal patterns up to 4 years after Roux-en-Y gastric bypass surgery. Obes Surg. 2004;14:1070–9.
Zhao CM, Furnes MW, Stenström B, et al. Characterization of obestatin- and ghrelin-producing cells in the gastrointestinal tract and pancreas of rats: an immunohistochemical and electron microscopic study. Cell Tissue Res. 2007 (in press).
Arch JR. beta (3)-Adrenoceptor agonists: potential, pitfalls and progress. Eur J Pharmacol. 2002;440:99–107.
Acknowledgments
This study was supported by grant from the Cancer Foundation of St. Olav’s Hospital, Trondheim, Norway. MWF received a short-term PhD fellowship from the Department of Cancer Research and Molecular Medicine, Norwegian University of Science and Technology, Trondheim, Norway.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Furnes, M.W., Tømmerås, K., Arum, CJ. et al. Gastric Bypass Surgery Causes Body Weight Loss Without Reducing Food Intake in Rats. OBES SURG 18, 415–422 (2008). https://doi.org/10.1007/s11695-007-9392-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11695-007-9392-8