
Abstract
We developed a quantum-like model describing the gene regulation of glucose/lactose metabolism in a bacterium, Escherichia coli. Our quantum-like model can be considered as a kind of the operational formalism for microbiology and genetics. Instead of trying to describe processes in a cell in the very detail, we propose a formal operator description. Such a description may be very useful in situation in which the detailed description of processes is impossible or extremely complicated. We analyze statistical data obtained from experiments, and we compute the degree of E. coli’s preference within adaptive dynamics. It is known that there are several types of E. coli characterized by the metabolic system. We demonstrate that the same type of E. coli can be described by the well determined operators; we find invariant operator quantities characterizing each type. Such invariant quantities can be calculated from the obtained statistical data.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Recently the mathematical formalism of quantum mechanics and the ideology of operational description of observed data (elaborated by N. Bohr and W. Heisenberg) were successfully applied outside of physics; in psychology, cognitive science, economics (e.g. Khrennikov 2004; Busemeyer et al. 2006; Cheon and Takahashi 2010; Asano et al. 2010, 2011; Basieva et al. 2010), see the monograph (Khrennikov 2010) for the extended list of references. In 2010 the quantum-like behavior and information processing were found in microbiology and genetics (Basieva et al. 2010). We found experimental evidences, see Inada et al. (1996), demonstrating that the genome processes information nonclassically exhibiting behavior similar to the quantum interference. Hence, the powerful methods developed for quantum physics and quantum information, see e.g. Khrennikov (2010), can be applied even to the description of the mechanism of the genetic regulation in cells.
This paper contains “Appendix” with simple introduction to the quantum formalism and quantum information. We want to make this paper readable for mathematically oriented researchers in microbiology and genetics.
Our quantum-like model can be considered as a kind of the operational formalism for microbiology and genetics. Instead of trying to describe processes in a cell in the very detail (e.g., Wanke and Kilian 2009), we propose a formal operator description. Such a description may be useful in situation in which the detailed description of processes is impossible or extremely complicated. The usage of the operational formalism provides a possibility to obtain the main probabilistic predictions without constructing a detailed model.
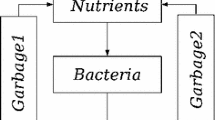
Escherichia coli (E. coli) is a bacterium which has been studied well in biology. One of interesting phenomena in E. coli’s growth is the gene regulation of glucose/lactose metabolism. The energy for E. coli’s activity is produced by metabolizing sugar: glucose, lactose, etc. When E. coli is incubated in a test tube containing both glucose and lactose, E. coli will digest glucose first and lactose second. Many microbiological studies demonstrated that E. coli has a preference for carbon resources, that is, E. coli likes glucose better than lactose. Such preference is caused by the difference of the metabolism efficiency (or cost) between lactose and glucose (Inada et al. 1996) (since the operon theory was proposed in 1956–1961 (Jacob and Monod 1961), the regulatory system of gene expression of lactose operon has been extensively studied and the molecular mechanism of it has been mostly elucidated including the catabolite repression).
Recently, it was pointed out (Basieva et al. 2010) that the complex microscopic biological dynamics of E. coli’s metabolism violates one of basic law of classical probability theory—the law of total probability (playing a crucial role in Bayesian analysis). It was argued that such complex behavior can be described by models based on non-Kolmogorovian probability theory inspired by the quantum phenomena (Khrennikov 2010) (such models are called quantum-like models). In the present paper we shall show that starting with experimental statistical data it is possible to reconstruct basic operators of the quantum-like operational formalism for glucose/lactose regulation in genome.
An important element of our model is the usage of adaptive dynamics (Ohya 2008; Asano et al. 2007)—dynamics which depend nontrivially on biochemical context (Khrennikov 2010). The quantum formalism provides a possibility of the mathematical description of adaptive (contextual) dynamics. This formalism was very well developed in quantum physics and especially recently in quantum information theory (Ingarden et al. 1997; Khrennikov 2010; Ohya et al. 2011). Therefore, we describe the adaptive dynamics of the gene regulation in cells by theory of operators in complex Hilbert spaces. The notions from quantum information are adapted to the microbiological context. The most important for us are the notions of the information channel and lifting. The second one belongs to the advanced level of quantum information theory and its successful usage in the description of the regulatory gene activity is an important step in the invention of the methods of quantum information theory to microbiology.
We analyze statistical data obtained from experiments (Inada et al. 1996), and we compute the degree of E. coli’s preference within adaptive dynamics. It is known that there are several types of E. coli characterized by the metabolization system. We demonstrate that the same type of E. coli can be described by the well determined operators; we find invariant operator quantities characterizing each type. Such invariant quantities can be calculated from the obtained statistical data. We do not need difficult simulation of complex biochemical systems. We use only simple calculation based on adaptive dynamics and its operational representation. Our quantum-like model can explain E. coli’s adaptive behavior.
Violation of total probability law
In this section, we show that statistical data obtained from experiments on the gene regulation of E. coli metabolism violate the total probability law, which is one of basic laws in classical probability theory; see Basieva et al. (2010) for detail. We modify the probabilistic representation (Basieva et al. 2010) of statistical data from Inada et al. (1996) to operate solely with conditional probabilities (as it is done in the standard applications of the formula of total probability in, e.g., Bayesian analysis).
In order to digest lactose, E. coli must produce the enzyme of β-galactosidase. The amount of its production is regulated by a series of genes, called lactose operon. The activity of E. coli’s lactose operon is estimated by the value of Miller’s Units (MU), which is measured in the experiment of β-galactosidase assay (Inada et al. 1996).
Let us consider the measurement of MU in the different situations: E. coli grows in the test tube containing (1) only lactose; (2) only glucose; (3) both glucose and lactose. Table 1 shows the measured value of MU for the cases (1), (2) and (3). We use the wild type of E. coli (W3110) in data-(I) and -(II). The lactose/glucose concentration in data-(I) is different from that in data-(II). The MU values in the case of (3) are 43 and 64, respectively. It is clear from this result that the activities of β-galactosidase in case of (3) are as week as E. coli has strong preference for glucose.
Such property of sugar metabolism in E. coli has been explained in biology as follows. E. coli cell has various transport systems including lactose permease (LacY) and glucose transporter (PtsG). They recognize their substrates (glucose or lactose). If there is either sugar in the medium, the sugar enters into cells with its concentration dependent manner. Therefore lactose outside the cell can induce lactose operon expression in concentration dependent manner, if there is no glucose outside: Gene expression apparatus recognizes the lactose concentration inside as high enough to induce mRNA synthesis.
On the other hand, if there is glucose outside at the same time with lactose, the cell can recognize glucose and lactose concentrations outside but, glucose transport process itself inhibits the lactose permease activity so the lactose concentration inside decreases. In this situation, the gene expression apparatus can not recognize the lactose concentration inside as high enough to induce mRNA synthesis.
This process can be quantitatively discussed with metabolic networks. However, the metabolic networks can be very large, so that, the construction of a mathematical model of the network and its analysis are a daunting task; most of the kinetic parameters are unavailable. Instead of such detailed model, we here explain an approach of coarse-grained model which include the process of the gene expression in the cell.
We focus on the behavior of one cell. There are many molecules of lactose or glucose around the cell. These molecules sometimes arrive at the surface of the cell, and they enter into the cell through the gate of a transport system. We call this event “E. coli’s detection”. In this experiment, we use two kinds of sugars, so that we consider an event system {L, G}: L means the event that E. coli detects a lactose molecule around its cell; G means the E. coli detects a glucose molecule around its cell. We use only two kinds of sugars in this experiment, so that, we assume that P(L ∪ G) is equal to one.
Let P(L) and P(G) be probabilities of event L and G, respectively. It is clear that, in the case of (1) where there is only lactose in medium, these probabilities are given by P(L) = 1 and P(G) = 0. In the case of (2), P(L) = 0 and P(G) = 1 are given. In the case of (3), these probabilities are proportional to the concentrations of lactose/glucose. The probabilities for data-I are
The probabilities for data-II are
Let us consider another event system {+, −}: + means the event that E. coli activates its lactose operon, that is, the event that β-galactosidase is produced through the transcription of mRNA from a gene in lactose operon; − means the event that E. coli does not activates its lactose operon.
In the case of (1), the only event L occurs since there are only lactose in the medium. Therefore, we can give the conditional probability P(+|L) with MU values in (1), for example, we can give the probability P(+|L) for data-I as
and that for data-II as
Similarly, we can give the probability P(+|G) for each data with MU value in (2); P(+|G) for data-I is
and P(+ |G) for data-II is
Here, let us discuss the relation among the experiments of (1), (2) and (3). If we assume that the experiment (3) is a simple sum of (1) and (2), then the value of MU (1) × P(L) + MU (2) × P(G) should be equal to MU (3) where MU (n) denotes MU values in the case of experiment (n). However we can easily see that such relation does not hold for each data. We remark that the experiment (3) is not the simple sum of (1) and (2).
In classical probability theory, the following well-known formula of probabilities, called “total probability law”, should hold, Khrennikov (1999).
Here P(+|L ∪ G) denotes the probability of the event + if either L or G occurs in the case of (3). However, it is important to note that the probabilities calculated from obtained data do not satisfy the above formula of total probability law; we can calculate the value given by right-hand side of Eq. (1);
for data-(I) and
for data-(II).
Also we can calculate the value given by left-hand side of Eq. (1) in the case of (3). So that, we can give P(+|L ∪ G) for data-(I) as
and that for data-(II) as
These calculated probabilities are shown in Table 2.
The value of left-hand side of Eq. (1) is much smaller than that of right-hand side of Eq. (1) for both data, that is, total probability law of the Eq. (1) is violated.
This violation of total probability law is well over the experimental errors. We can give the real values of MU (1), MU (2) and MU (3) with an error rate δ as MU (1)(1 ± δ), MU (2)(1 ± δ) and MU (3)(1 ± δ), respectively. If we assume that the total probability law is hold, then the following equation should be hold.
From this equation, we obtain the error rate δ ≈ 0.959 for data-(I), and error rate δ ≈ 0.889 for data-(II). In general, we may have 20 % or sometimes 30 % standard deviations of the real value. However this calculation shows that over 85 % error rate is needed to satisfy the total probability law.
Here let us consider the opposite situation that E. coli has “no preference for any sugar”. Although such egalitarian E. coli is ideal and unrealistic, he must never distinguish lactose from glucose. Therefore this ideal and unrealistic E. coli must probabilistically produce β-galactosidase in proportion to the concentration of lactose. In this case, the experiment (3) is just simple sum of experiment (1) and (2), and total probability law is satisfied. We remark that the strength of E. coli’s preference is expressed as the difference between left-hand side and right-hand side in total probability law.
New mathematical law computing the probability in adaptive dynamics for glucose effect of E. coli
In this section, we briefly explain how to compute these probabilities with adaptive dynamics and lifting theory, see Ingarden et al. (1997), Ohya et al. (2011), Asano et al. (2007) for the details of lifting theory).
First, let us introduce the initial state \(\rho_0 = \vert x_0 \rangle \langle x_0 \vert \), the density operator in Hilbert space \(\mathcal{H}=\mathbf{C}^{2}\) (the qubit space). The state vector x 0 is written as
The basis {e 1 = (0, 1)T, e 2 = (1, 0)T} describes the detection of lactose or glucose by E. coli, that is, the events L and G. The initial state ρ0 is the state of E. coli before the detection of the molecules. When the E. coli recognizes these molecules, the following state change occurs;
where D is an operator given by a diagonal matrix
with |α|2 + |β|2 = 1. The operator D represents E. coli’s adaptive effect for surroundings (concentration of lactose or glucose), and it is called detection operator. Note that |α|2 and |β|2 are nothing else than the probabilities for the events L and G, that is, P(L) and P(G). The state σ D ≡ DD * means the distribution of P(L) and P(G). In this sense, the state σ D is derived from the solution concentrations of lactose and glucose. The state determining the activation of the operon in E. coli depends on the detection state ρ D . In our adaptive dynamical model such state is produced by the following state-change
where Q is an operator given with complex numbers a, b, c and d as
This operator Q represents the state-change of lactose operon and we call it activation operator. The correlation between the activity of lactose operon and concentrations of lactose/glucose is described as the lifting
with a map (channel) \(\Uplambda_{A}\rho=A\rho A^{\ast}/{\rm tr}\left\vert A\right\vert ^{2}\rho.\) One can define the joint probabilities P(+ ∩ L) and P(+ ∩ G) as
where E 1 and E 2 are projection operator given by
Also one can define conditional probabilities as
and
The above definition of the conditional probability is different from classical one. We remark that the following property;
Invariant operator quantities in E. coli’s preference obtained from a statistical data
In this section, we show how to find invariant operator quantities in E. coli’s preference.
We give the activation operator Q by the following decomposed form:
with real numbers k L , k G and θ. Note that the above Q satisfy Eqs. (2)–(5) for any k L , k G and θ. From the Eqs. (6) and (7), we can express the conditional probability P D (+ | L ∪ G) as
with
If k L = k G and θ = π/2, then the total probability law holds;
However, for general k L , k G and θ, the total probability law is violated;
Let us remind that the violation of total probability law means E. coli’s preference. Therefore we can say the values of k L /k G and θ describe E. coli’s preference, which is given as an invariant quantity for the strain of W3110. It is natural that we assume that each strain of E. coli has own invariant quantity which describes E. coli’s character in metabolization.
From data-I and -II of Table 2, we can determine the operators A (I) and A (II), and we can easily confirm that A (I) is approximately same as A (II). The operator of E. coli W3110 is given by
Further we can obtain the values of k L /k G and θ by solving two equations which are given by assigning the probabilities for data-(I) and data(II) to the Eq. (8);
With \(\left\{A,\sqrt{k_{L}/k_{G}},\cos \theta\right\}\) which are shown the above, we can compute the probability P D (+|L ∪ G) for any D. Figure 1 shows the value of P D (+|L ∪ G) with respect to P(L). As seen in Fig. 1, lactose operon is active only when P(L) is very close to one. We can quantitatively estimate E. coli’s glucose preference for any concentration of lactose/glucose.

Value of P D (+|L ∪ G) with respect to P(L)
Conclusion
We found the decomposed form of the activation operator and the new invariant operator quantities A and B for the type W3110 of E. coli. We determine these operators from experimental data with the aid of a simple calculation based on the concept of adaptive dynamics.
References
Asano M, Ohya M, Togawa Y (2007) Entropic chaos degree of rotations and log-linear dynamics. In: Accardi L, Freudenberg W, Ohya M (eds) Quantum bio-informatics: from quantum information to bio-informatics, WSP, Singapore, pp 36–52
Asano M, Ohya M, Khrennikov A (2010) Quantum-like model for decision making process in two players game. Found Phys 41(3):538–548
Asano M, Ohya M, Togawa Y, Khrennikov A, Basieva I (2011) Dynamics of entropy in quantum-like model of decision making. J Theor Biol 281:56–64
Basieva I, Khrennikov A, Ohya M, Yamato I (2010) Quantum-like interference effect in gene expression; glucose-lactose destructive interference. Syst Synth Biol 5(1–2):59–68
Busemeyer JR, Wang Z, Townsend JT (2006) Quantum dynamics of human decision making. J Math Psychol 50:220–241
Cheon T, Takahashi T (2010) Interference and inequality in quantum decision theory. Phys Lett A 375:100–104
Inada T, Kimata K, Aiba H (1996) Mechanism responsible for glucose-lactose diauxie in Escherichia coli challenge to the cAMP model. Genes Cells 1:293–301
Ingarden RS, Kossakowski A, Ohya M (1997) Information dynamics and open systems: classical and quantum approach. Kluwer, Dordrecht
Jacob F, Monod J (1961) Genetic regulatory mechanism in the synthesis of proteins. J Mol Biol 3:318–356
Khrennikov A (1999) Interpretations of probability. VSP Int. Sc. Publishers, Utrecht/Tokyo; (2009) De Gruyter, Berlin, 2nd edn (completed)
Khrennikov A (2004) On quantum-like probabilistic structure of mental information. Open Syst Inf Dyn 11(3):267–275
Khrennikov A (2010) Ubiquitous quantum structure: from psychology to finance. Springer, Heidelberg
Ohya M (2008) Adaptive dynamics and its applications to chaos and NPC problem. QP-PQ:Quantum Prob White Noise Anal 21:186–216
Ohya M, Volovich I (2011) Mathematical foundations of quantum information and computation and its applications to nano- and bio-systems. Springer, Heidelberg
Wanke D, Kilian J (2009) A basic introduction to gene expression studies using microarray expression data analysis. In: Accardi L, Freudenberg W, Ohya M (eds) Quantum bio-informatics II: from quantum information to bio-informatics, WSP, Singapore, pp 314–326
Acknowledgments
Two authors (Irina Basieva and Andrei Khrennikov) were supported through visiting fellowships at Quantum BioInformation Center, Tokyo University of Science, March 2011.
Author information
Authors and Affiliations
Corresponding author
Appendix: Essentials of the mathematical formalism of quantum mechanics
Appendix: Essentials of the mathematical formalism of quantum mechanics
The mathematical formalism of quantum mechanics describes states of systems and observables; see book (Khrennikov 2010) for a simple representation of quantum mathematics for biologists and psychologists (the quantum-like approach is based on the observation that “quantum mathematics” can be applied outside quantum physics).
The basic mathematical structure of quantum mechanics is a complex Hilbert space H: a linear space over complex numbers (i.e., it is possible to form linear combinations of vectors with complex coefficients) endowed with a Hermitian bilinear form mapping a pair of vectors \(\psi_{1}, \psi_{2} \in H\) into a complex number denoted \(\langle\psi_{1} \vert\psi_{2}\rangle\) (we use Dirac’s notation which is typical for quantum information theory). We shall be interested in complex vectors normalized by one, i.e., \(\psi\in H\) such that \(\Vert\psi\Vert^{2}= \langle\psi\vert\psi\rangle=1.\) Such vectors encode so called pure states of quantum systems. Normalization by one is crucial for the probabilistic interpretation of pure states. Observables (e.g., the energy-observable or the position observable) are encoded by Hermitian operators.
The theory is especially simple in the finite dimensional case (which is typically considered in quantum information theory). Here \(H=\mathbf{C}^{n}\) is the Cartesian product of n-copies of the set of complex numbers \(\mathbf{C}\). Hence, a pure state \(\phi= (z_{1},\ldots,z_{n}),\) where \(\Vert \psi\Vert^{2}= \vert z_{1}\vert^{2}+\cdots+\vert z_{n}\vert^{2}=1\). Observables are given (in an orthonormal basis) by Hermitian matrices A = (a ij ); here \(\bar{a}_{ij}= a_{ji}\). In the Dirac notation the matrix elements are written as \(\langle\phi_{1}\vert A\vert\phi_{2}\rangle\). Coordinates of a state vector are interpreted as probabilities.
As was mentioned, in quantum mechanics the special Dirac symbolic notations are in use. A pure vector ψ is denoted as \(|\psi\rangle.\) This notation is especially useful for eigenvectors of quantum observables. Let A be a quantum observable, a Hermitian operator. By one of the basic theorems of linear algebra A always has the basis consisting of its eigenvectors, Ae j = α j e j (in the last equation we have not yet use the Dirac notations), where \(\alpha_{i}\in\mathbf{R}\) are real numbers and eigenvectors can be selected orthonormal, \(\langle e_{i}|e_{j}\rangle=\delta_{ij}(=1,i=j;=0,i\not =j)\). Suppose now that all eigenvalues are different, \(\alpha_{j}\not =\alpha_{i}\). Such operators are called operators with nondegenerate spectra. Then in the Dirac notations eigenvectors are written as \(|\alpha_{i}\rangle;\) typically the index is omitted: \(|\alpha\rangle\). Hence, each state-vector can be expanded in the form: \(|\psi\rangle=\sum_{\alpha}c_{\alpha}|\alpha\rangle,\;\sum_{\alpha}|c_{\alpha}|^{2}=1\). Consider the two dimensional case \(H=\mathbf{C}^{2}.\) Take some Hermitian operator with eigenvalues α = 0,1; its eigenvectors are denoted as \(|0\rangle ,|1\rangle\). The aforementioned expansion has the form:
In quantum information the vectors \(|0\rangle,|1\rangle\) encode 0, 1. The vector (9) encodes superposition of 0 and 1. It is considered as the quantum generalization of the classical bit of information; it is called quantum bit or simply qubit.
We remark that each pure state \(|\psi\rangle\) determines a Hermitian operator, the projector onto this state; \(\rho\equiv|\psi\rangle\langle\psi|\). The last symbol is simply the Dirac notation: \(\rho|\phi\rangle=\langle\psi|\phi \rangle|\psi\rangle.\) We recall the basic properties of ρψ: (a) it is positively defined, i.e., \(\langle\phi\vert\rho\vert\phi\rangle\geq0\) for any ϕ; (b) it is Hermitian; (c) its trace (the sum of diagonal elements) equals to one.
Consider now a statistical mixture (in the classical sense) of a few projection operators ρ i corresponding to pure states ψ i with weights p i ≥ 0, ∑ p i = 1,
Each operator of this form satisfies conditions (a)–(c) and vice versa. Denote the class of all operators with properties (a)–(c) by the symbol \(\mathcal{D}(H)\). This is the space of states of quantum systems. Its elements (called density operators) can be interpreted as statistical mixtures of pure states. In general a density operator can be represented in the form (10) in many ways. There is one special expansion corresponding to eigenvectors of ρ. The density operator corresponding to a pure state can be characterized in the following way: in the basis of eigenvectors, its matrix has only one nonzero element (equal to one), i.e., up to a permutation of eigenvectors: \(\rho=\left( \begin{array}{cc}1 & 0\\ 0 & 0\end{array} \right)\) , where the blocks of zeros have the corresponding sizes. However, this takes place only in the basis of eigenvectors. Qubit of given by (9) has the density-matrix representation: \(\rho(t_{0})=\left( \begin{array}{cc}|c_{0}|^{2} & c_{0}\bar{c}_{1}\\ \bar{c}_{0}c_{1} & |c_{1}|^{2}\end{array} \right)\).
A quantum channel is a map from the space of quantum states \(\mathcal{D} (H)\) into itself which can be represented in the form
where V i are operators in H such that ∑ i V i V * i = I (here I denotes the unit operator). The ρ0 is the input-state (encoding information coming into the quantum channel) and ρout is the output-state (encoding information going from the quantum channel). The representation (11) can be rewritten in the form
where
and p i = tr V i ρ0 V * i . We have ∑ i p i = 1, p i ≥ 0. Hence, they can be interpreted as probabilities. The operators \(\rho_{\rm out; i}\in\mathcal{D}(H)\). Thus the expansion (12) can be interpreted in the following way: a quantum channel produces states ρout;i with probabilities p i .
Liftings are a class of channels from \(\mathcal{D}(H)\) to \(\mathcal{D}(H \otimes K);\)
Here K is another Hilbert space, and \(\mathcal{D}(H \otimes K)\) is a set of density operator on the compound system. We introduce the following liftings which are often used in physics.
-
Linear Lifting: A linear lifting is affine and its dual is a completely positive map.
-
Pure Lifting: A pure lifting maps pure states into pure states.
-
Nondemolition Lifting: A lifting is nondemolition for a state \(\rho\in S(H)\) if ρ is invariant for any Hermitian operator A in the sense of
Applications of lifting in adaptive dynamics was introduced in Ohya and Volovich (2011).
Rights and permissions
About this article
Cite this article
Asano, M., Basieva, I., Khrennikov, A. et al. Quantum-like model for the adaptive dynamics of the genetic regulation of E. coli’s metabolism of glucose/lactose. Syst Synth Biol 6, 1–7 (2012). https://doi.org/10.1007/s11693-012-9091-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11693-012-9091-1