
Abstract
Rhizoctonia solani is a destructive soil-borne plant pathogen having a vast host range that infects various agricultural and horticultural crops. A total of 25 R. solani isolates were collected from a field of rice-potato rotation to determine their morphological variability, mycelial compatibility, cross-infectivity and fungicide sensitivity. Considerable variability in cultural and morphological features was observed among R. solani isolates collected from rice and potato crops. Rice isolates grew faster than potato isolates and hyphal width of both the host isolates varied from 6.67 to 9.37 μm. Significant variability was observed in sclerotial characteristics such as sclerotia colour, size, intensity and sclerotial distribution pattern on the Petri plates. Most of the isolates from both hosts produced micro (≤ 1.25 mm) size sclerotia and only few isolates produced macro (> 1.25 mm) size sclerotia. The sclerotial colour of most of the rice isolates was brown and some isolates exhibited black sclerotial colour. All the potato isolates produced brown-coloured sclerotia. Hyphal interaction studies revealed that potato isolates belonged to AG-3 and rice isolates belonged to AG1-IA group. Mycelial compatibility studies showed that 80.7% of the pairings were non-compatible, while 19.3% of the pairings exhibited a compatible interaction. Cross-infectivity revealed that R. solani potato isolates did not infect rice plants whereas rice isolates showed a varied degree of pathogenicity on potatoes and Rhizoctonia-like atypical symptoms were observed on the tuber surface. The EC50 and EC90 values of representative isolates from both hosts exhibited considerable variation with regard to sensitivity level against fungicides thifluzamide 24% SC and mancozeb 50% + carbendazim 25% WS. The outcomes of this study will help to develop management strategies and breeding programmes on varietal evaluation for effective management of Rhizoctonia diseases of potatoes and rice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rhizoctonia solani Kühn (teleomorph: Thanatephorus cucumeris) is a soil and tuber-borne fungus having a broad host range. Rhizoctonia is considered as an economically important phytopathogen of rice and potato that reduces the yield and quality of both crops worldwide, including India. R. solani is a complex of genetically diverse group of fungi that exhibits distinct cultural, morphological, physiological and pathological characteristics (Ou 1985; Chaudhary et al. 2019). Based on the average number of nuclei per vegetative hyphal cell, isolates of Rhizoctonia can be divided into the groups of multinucleated Rhizoctonia (MNR), binucleated Rhizoctonia (BNR) and uninucleated Rhizoctonia (UNR). Isolates of R. solani can be recognized and grouped into numerous anastomosis groups (AGs) based on hyphal interaction among the isolates having different phenotypic and genotypic properties (Ogoshi 1987). Presently, 13 AGs designated as AG1–AG13 and one bridging group (AG-BI) have been assigned within MNR, whereas is BNR composed of 19 AGs designated as AG A–AG W, based on hyphal fusion (Sharon et al. 2008; Dong et al. 2017; Zhao et al. 2019; Chaudhary et al. 2023). R. solani isolates of some multinucleate AGs are further classified into different subgroups (Carling et al. 2002; Ogoshi 1987). R. solani can infect crop plants as well as weed plants (Chahal et al. 2003). R. solani infects rice and potato crops causing sheath blight and stem canker/black scurf disease in rice and potato, respectively. Kaur and Kaur (2013) collected 22 isolates of R. solani from potato and rice from Punjab, India, and reported that R. solani isolates infecting potato belonged to AG-3, while rice infecting isolates belonged to AG1-IA.
Under North Indian conditions, in the past few years sheath blight and black scurf diseases have posed serious threats to rice and potato production. Most of the cultivated varieties of rice and potato growing in the states are susceptible to R. solani. In major areas, farmers cultivated potato crop after rice crop in the same field and it is assumed that following this crop rotation over the years increases the inoculum level of pathogen in the soil. However, several studies of cross-infectivity of R. solani isolates attacking various host crops like cereals and pulses have been reported (Simay 1993; Singh et al. 2002); very few reports are available on intra-field variability and cross-pathogenicity of R. solani isolates from rice and potato. The complexity and variability in R. solani have been documented from various regions of the globe that can affect management strategies of the diseases. Rhizoctonia disease of rice and potato can be controlled through agronomic practices such as crop rotation, use of biocontrol agents and use of certified pathogen-free seeds. However, the application of synthetic fungicides remains the widely used strategy for the management of Rhizoctonia diseases. A wide range of site-specific fungicides have been extensively used for controlling Rhizoctonia diseases in India. It is reported that fungicide sensitivity to various anastomosis groups (AGs) varied. Olaya et al. (2013) reported resistance (insensitivity) in isolates of R. solani AG1-1A to fungicide azoxystrobin (QOI) in Louisiana and against thifluzamide in Henan Province, China, by Chen et al. (2012). Additionally, Rhizoctonia AGs have been reported to show variable sensitivity to different fungicides in both in vitro and in vivo conditions (Amaradasa et al. 2014; Calleros et al. 2000; Campion et al. 2003). Therefore, this study aimed to characterize intra-field R. solani isolates collected from rice-potato crop rotation based on cultural and morphological features, mycelial compatibility, cross-infectivity potential and sensitivity to fungicides from both hosts.
Material and Methods
Sampling and Isolation of R. solani from Rice-Potato Rotation
Generally, in potato research experimental fields at ICAR-Central Potato Research Institute, Regional Station, Meerut (29.1° N, 77.92° E, 300 masl), India, dhaincha-potato rotation is followed, but in some fields rice-potato and maize-potato systems are also applied. In rice-potato rotation, typical symptoms of rice sheath blight, stem canker and potato black scurf were regularly observed in the fields. The field soil was sandy loam, organic carbon 0.38–0.40% and pH was 6.5–7.5. The sheath blight-infected rice plants were collected during the Kharif season and stem canker-infected potato plants were collected after 45 days of crop growth at a depth of 20 cm during the Rabi season. The black scurf-infected potato tubers were collected at the time of tuber harvesting. Collectively, a total of 35 symptomatic rice plants, stem canker and potato tuber samples were collected from rice-potato rotation and brought to the laboratory for pathogen isolation. Isolation of R. solani was done by following the standard protocol (hyphal tip/single sclerotial method) on water agar (WA) or potato dextrose agar (PDA) media (Rangaswami and Mahadevam 2004). Small pieces (5-mm cube) of the infected rice sheath and sclerotia from potato tubers were surface sterilized with 0.25% sodium hypochlorite solution, washed twice in sterilized distilled water and air dried under aseptic conditions. The pieces and sclerotia (four per plate) were placed on 90-mm diameter Petri plates on PDA supplemented with 100 µg/ml of streptomycin sulphate and incubated at 25 ± 2 °C for 3 days in the dark. As the mycelial growth appeared, colonies possessing the typical characteristics of R. solani hyphae (Ogoshi 1987) under a microscope were hyphal tipped and purified on PDA medium, incubated at 25 ± 2 °C for 5 to 6 days and observed periodically.
Cultural Characteristics and Microscopic Studies of R. solani
Initially, the isolates were identified as R. solani using cultural features on PDA (Sneh et al. 1991). All the isolates were grown on 2% water agar (WA) at 25 ± 2 °C for 3 days and stained with 0.05% lactophenol blue and hyphal morphology, viz., angle of hyphal branching, constrictions at the point of origin and hyphal width were examined under a microscope. For cultural and sclerotial characterization of R. solani isolates from both crops, mycelial plugs (5-mm diameter) were cut from the margin of actively growing colonies of 3-day-old cultures and transferred at the centre of 90-mm PDA Petri plates. The plates were incubated at 25 ± 2 °C for 7 days in the dark. The colony’s radial growth was measured at every 24-h intervals. Sclerotial characteristics like size, colour, intensity and formation patterns were recorded. The morphological data were converted into numerical values (0, 1). The generated data matrix using combined cultural and morphological characteristics was statistically analysed by Multi-Variate Statistical Package Plus (MVSP version 3.2) using Gower’s coefficient (Chaudhary et al. 2020a).
Anastomosis Grouping
R. solani isolates from both crops were assigned to anastomosis groups (AGs) by pairing with a tester strain following the clean-slide technique described by Kroland and Stanghellini (1988). The tester strain of rice R. solani AG1-IA (ITCC 7650) was procured from the Indian Type Culture Collection (ITCC), ICAR-Indian Agricultural Research Institute (IARI), Pusa, New Delhi, and AG3 (RS-20) was procured from ICAR-Central Potato Research Institute (CPRI), Shimla, Himachal Pradesh, India. Mycelial plugs (5 mm2 dia.) were excised from the margin of actively growing colonies of unknown isolate and tester isolate and placed on 1.5% WA-coated clean microscope glass slide 2–3 cm away. After 48–72 h when hyphae of both the isolates overlapped, slides were mounted with 0.05% lactophenol cotton blue (HiMedia) and examined under an optical microscope for potential hyphal fusion events under different magnifications. For all isolates, the pairings were repeated twice and at least five fusion events per pairing were analysed. Anastomosis reactions were assigned to one of the four categories, i.e. C0: no reaction, C1: contact fusion, C2: somatic fusion or perfect anastomosis and C3: auto-anastomosis, as described by Carling (1996). Anastomosis reaction was considered positive when hyphae of both crop isolates and AG tester strain made contact with each other and their walls fused, with subsequent lysis of adjacent cells.
Ten random microscopic locations on each glass slide were considered to observe hyphal interactions between the unknown isolate and the tester isolate of respective AG and fusion frequency percentage (FF%) was calculated by the following formula (Carling and Sumner 1992):
where:
A = sum of fusion locations (in C1, C2, C3) in 10 microscopic fields
B = sum of contact points in 10 microscopic fields
Isolates showed pairing at more than 80% of locations belonging to the respective anastomosis group (AG).
Mycelial Compatibility among R. solani Isolates from Rice-Potato Crop Rotation
To determine mycelial compatibility, R. solani isolates from both crops were paired in all possible combinations on PDA medium according to the procedure of MacNish et al. (1997). Each isolate was grown on PDA for 2–3 days and paired with one another in all possible combinations by placing 5-mm agar plugs taken from the edge of the growing mycelia onto PDA in a 90-mm Petri dish, spaced 5 cm apart. Plates were incubated at 25 ± 2 °C until the mycelia from each isolate met or formed sclerotia (1 week). A modified rating based on Ceresini et al. (2002) was used. Mycelial interaction was scored as compatible when mycelia merged or incompatible when a visible clear zone or a sclerotial barrier was formed in the contact zone between paired colonies. By using binary matrices, the pairing reactions were scored as 0 (incompatible) or 1 (compatible) for each isolate pair.
Cross-Pathogenicity of R. solani Isolates from Rice-Potato Crop Rotation
Pathogenicity of Potato Isolates on Rice
Cross-pathogenicity of selected R. solani potato isolates were evaluated on rice plants under laboratory and greenhouse conditions.
Under Laboratory Conditions
A detached leaf method was used for inoculation under laboratory conditions. After 45 days of growth, plant leaves were cut, washed with distilled water three times and placed in the tray. A moist chamber was prepared by placing a wet blotter sheet in a plastic tray (45 × 35 × 7.5 cm) and maintaining humidity with distilled sterilized water. Three leaves per isolate were used and inoculated by placing a bit of 5.0-mm size of mycelium culture (5 days old) in the centre of the leaf (Chaudhary et al. 2020b). Control was maintained in a similar manner in which leaves were inoculated with sclerotia of a rice isolate. Un-inoculated leaves were also kept similarly. The trays with leaves were incubated in a growth chamber (25 ± 2 °C, 12 h light). The observations were recorded after 3 days of inoculation for the appearance of disease lesions development.
Under Greenhouse Conditions
Surface sterilized rice seeds were sown in earthen pots (22.9 cm in diameter) filled with autoclaved field soil in the first week of June 2019. The field soil was sandy loam and had 6.5–7.5 pH. A stem inoculation technique was used for inoculation. At 45 days of growth, leaf sheaths of rice plants were inoculated with each of the selected potato R. solani isolates by a bit of 5.0mm size of mycelium culture (5 days old) and covered with aluminum foil along with a drop of sterilized water. Three plants per isolate were maintained. The plants were also inoculated with a rice isolate as a control. Un-inoculated plants were also kept as another control. The pots were observed daily for the appearance of disease symptoms.
Pathogenicity of Rice and Potato R. solani Isolates on Potato
Preparation of Inocula
Wheat kernels were used to prepare mass inocula of R. solani isolates from both crops. Mass inocula of selected 20 isolates (10 from each crop) were prepared separately in Erlenmeyer flasks of 100 ml capacity. The wheat grains were soaked in water overnight. The imbibed grains thus obtained were put in the flasks @ 20 g/100 ml flask and autoclaved at 20 psi for 60 min. After cooling, the grains were shaken by jerking the flask against the palm of the hand gently to avoid clumping. The autoclaved grains were then inoculated with 4–5 mycelial discs (5 mm diameter) of 7-day-old culture of each isolate separately and incubated at 25 ± 2 °C for 15 days. Flasks were shaken daily to prevent mycelium clumps. After 15 days, the wheat grains were fully covered with running mycelium and dark brown to black sclerotial bodies. The wheat grains with hyphae and sclerotia were collected, dried naturally and used for inoculation.
Preparation of Soil
Soil for filling the pots was prepared by mixing potato field soil, sand and well-decomposed farm yard manure (FYM) in the proportion of 2:1:1. The mixture was sterilized with formalin (40%) diluted with water. The soil was then heaped in a square block. The soil heap was covered by a polythene sheet for 48 h. After the 7th day of treatment, sterilized pots (33 × 25 cm2) were filled up with the sterilized soil.
Pathogen Inoculation
R. solani inocula multiplied on wheat grains were used to study the pathogenic variability of selected 20 isolates (10 from rice and 10 from potato) under greenhouse conditions. The wheat grains colonized by R. solani isolates were mixed with pot soil @ 5 g/kg soil. After inoculation, pot soils were moistened regularly for 10 days to create congenial conditions for R. solani growth. Healthy-looking potato tubers of cv. Kufri Bahar surface sterilized for 20 min with 0.5% KMnO4 were used for planting. The experiment was designed with three replications per treatment in randomized block design (RBD). Three pots planted with un-inoculated healthy tubers served as the control. Black scurf infested seed tubers of cv. Kufri Bahar were also maintained in three replications as another control.
Harvesting and Data Collection
At 120 days after planting, tubers were harvested and kept in a dry place at room temperature for 2 days to dry the sclerotia and then washed carefully to remove soil. The tubers infected by other pathogens and damaged by pests were removed. Each replication of each treatment all the tubers were selected to evaluate the disease severity of black scurf. The ratio of tuber surface area covered with sclerotia was used as a general method to evaluate the pathogenicity of R. solani isolates. The tuber surface area covered with sclerotia was estimated using the following scale (Lal et al. 2022):
-
0: no sclerotia
-
1: 1–25% of tuber surface covered
-
2: 26–50% of tuber surface covered
-
3: 51–75% of tuber surface covered
-
4: > 75% of tuber surface covered
Fungicide Sensitivity Assay
The fungicide sensitivity level of selected R. solani isolates from both crops was tested using the poisoned food technique described by Grower and Moore (1962). Poisoned food technique is a primary assay used for the evaluation of the efficacy of chemicals against pathogens under in vitro conditions. Two commercial fungicides, namely, thifluzamide 24% SC and mancozeb 50% + carbendazim 25% WS, were used. Concentrated fungicide suspensions of each fungicide were pipetted into flasks containing autoclaved ¼ strength PDA medium to obtain the following concentrations: 1, 5 and 10 ppm. The fungicide-amended PDA media with each concentration was poured at 20 ml per 90-mm diameter Petri dish. The Petri dishes containing PDA media without fungicides were used as control. Mycelial discs of 5-mm diameter were cut from the margin of the newly growing 4-day-old culture of each selected rice and potato R. solani isolates were placed separately on PDA plates containing different concentrations of test fungicides. Four replicates for each fungicide concentration and control Petri plates per isolate were used and repeated twice in a randomized complete block design. The plates were incubated at 25 ± 2 °C in darkness for 7 days. The diameter of the mycelial growth for each fungicide-amended plate was measured along two perpendicular directions when the mycelial growth of the control reached the edge of the Petri plate. The diameter of the mycelial plug was subtracted and the average growth of each isolate was determined using the two growth measurements. The percent growth inhibition for each isolate by fungicide combination was calculated using the following formula.
For each isolate, a linear regression of the probit of the percent of inhibition related to the control of mycelia growth versus the log10 transformation of the three concentrations of each fungicide was obtained. The 50% effective concentration (EC50) and 90% effective concentration (EC90), which were the fungicide concentrations that resulted in 50% and 90% mycelial growth inhibition, respectively, were calculated using regression equation for all the selected R. solani isolates (Zhang et al. 2007).
Statistical Analysis
Data of pathogenicity testing was statistically analysed using OP Stat (http://14.139.232.166/opstat/). Analysis of variance was used to test differences between variables and means were separated using Fisher’s protected least significant differences (LSD).
Results
Sampling and Isolation of R. solani from Rice-Potato Crop Rotation
Typical greenish-grey and water-soaked lesions on the leaf sheath of rice plants and stem canker/black scurf symptoms on potato progeny tubers were regularly observed in experimental fields at the ICAR-Central Potato Research Institute, Regional Station, Meerut, Uttar Pradesh, India, where rice-potato crop rotation is practised. Initially, usually lesions on infected rice sheath were elliptical or irregular, and water-soaked, while greenish-grey spots were noticed. As the infection progressed, these spots enlarged and coalesced forming larger lesions with greyish-white centres surrounded by irregular tan to dark brown borders (Fig. 1a), sometimes whitish-brown coloured sclerotia were also observed on mature plants. On the other hand, dark brown to black coloured usually irregularly shaped and small, flat to large sclerotial structures adhered tightly to the potato progeny tuber skin (Fig. 1b). Furthermore, typical symptoms of stem canker (Fig. 1c) and aerial tuber formation (Fig. 1d) were also observed; however, the severity of stem canker was low. In total, 25 isolates of R. solani (14 from rice and 11 from potato) were isolated on WA medium. The hyphal tips of actively growing mycelium were sub-cultured on PDA medium and the purified isolates were cultured on PDA slants and maintained at 4 °C.

Typical symptoms/signs of Rhizoctonia diseases: a rice sheath blight, b potato black scurf, c potato stem canker and d aerial tuber formation on potato stem due to Rhizoctonia infection
Cultural Variability and Microscopic Studies among the R. solani Isolates
Isolates collected from rice and potato crops were morphologically characterized as per the descriptions of R. solani given by Ogoshi (1987) and Sneh et al. (1991). All isolates showed typical cultural and morphological features of R. solani under visually and microscopically observations. Microscopic studies revealed that the hyphae of all the R. solani isolates were branched, at right angles and a septum was always present in the hyphae branch near the origin point, with a slight constriction at the branch (Fig. 2a, b).

Microscopic observations of hyphae of R. solani isolates. A potato and B rice showing right-angle branched hyphae and arrow indicates constriction at the point of branching
Considerable variation was observed in the sclerotial and cultural characteristics of the rice and potato R. solani isolates. Based on mycelial growth rate, isolates were classified as slow (up to 10.0 mm/day), medium (10.1–20.0 mm/day) and fast (> 20.0 mm/day) growing. Mycelial growth rate (mm/day) in both the crop isolates ranged from 9.5 to 23.8 mm/day. The results revealed that most of the rice isolates (92.8%) exhibited fast growth rate (20.2–23.8 mm/day). In the case of potato isolates, most of the isolates (81.8%) exhibited medium (10.7–14.0 mm/day) growth rate, except isolates PR-1 (9.7 mm/day) and PR-6 (9.5 mm/day) which showed slow growth rate (Table 1). It was observed that most of the rice isolates almost filled the 90-mm PDA plate within 48 h, whereas potato isolates attained the full radial mycelial growth in 96 h. Most of the rice isolates exhibited whitish brown mycelial colour, while few isolates showed white and one isolate RR-23 exhibited light brown mycelial colour. In case of potato isolates mycelial colour varied from whitish brown to dark brown. Two isolates (PR-3 and PR-5) had dark brown mycelial colour and isolates (PR-4 and PR-7) exhibited brown colour; remaining isolates showed whitish brown mycelial colour. Based on the type of mycelial growth, isolates were categorized as appressed, raised and fluffy. The hyphal width of R. solani isolates from both the crops varied from 6.7 to 9.7 μm. The hyphal width of rice isolates varied from 6.7 to 8.4 μm, while potato isolates varied from 8.2 to 9.4 μm (Table 1). The hyphal width among the isolates was statistically non-significant.
Observations on sclerotial features of R. solani isolates from both crops revealed significant variation in sclerotial size, colour, intensity, sclerotial position, honeydew secretion and sclerotial distribution (Table 2). It was observed that in the case of rice isolates, sclerotia formation initiated after 72–96 h of incubation and matured after 96 to 120 h, while in the case of potatoes, most of the isolates initiated sclerotia formation after 96 h of incubation and matured after 144 to 168 h. No sclerotia formation was observed in isolate PR-5. Based on the sclerotial size, isolates were categorized in two groups; the first group was micro (≤ 1.25 mm), and the second group was macro (> 1.25 mm) sclerotia. All the rice and potato isolates produced micro size sclerotia, except isolates RR-12 and RR-14 of rice and PR-4 and PR-11 of potato, which formed macro size sclerotia, respectively. The sclerotial colour of most of the rice isolates was brown and some isolates exhibited black sclerotial colour. All the potato isolates produced brown-coloured sclerotia, except isolate PR-2, which had black sclerotial colour.
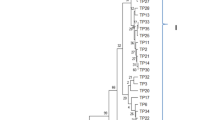
In most of the rice isolates, sclerotia were distributed in a scattered manner, except in isolates RR-13 and RR-16, which formed sclerotial ring towards the periphery, and isolates RR-12 and RR-25 produced a ring towards the centre; in RR-19 centrally concentrated pattern was observed (Fig. 3a). Likewise, most of the potato isolates also produced sclerotia in a scattered manner, except isolates PR-9 and PR-11 generated a centrally concentrated pattern and isolate PR-8 created sclerotial ring towards the centre (Fig. 3b). Based on sclerotial intensity, isolates were categorized as scanty, moderate and abundant. Most of the potato isolates produced abundant or moderate sclerotia, except isolates PR-4 and PR-9 had scanty sclerotia. In the case of rice isolates, moderate or abundant type sclerotia were produced by most of the isolates, except isolates RR-21 and RR-23 formed scanty type sclerotia. No honey dew formation was observed in potato isolates, except in five isolates (PR-1, PR-4, PR-8, PR-9 and PR-11), and in the case of rice isolates only three isolates, namely, RR-11, RR-20 and RR-25, produced honey dew (Table 2). A dendrogram was constructed with a combined cultural and morphological data set of 11 characters by UPGMA using Gower’s coefficient (Fig. 4). The resulting dendrogram (Gower’s coefficient) divided all the isolates into five clusters with similarity between 52 and 100% among isolates and linked to each other at 52% similarity. The clades I and II had isolates from rice and clade III grouped two potato isolates. Clade IV contained potato isolates with one isolate from rice (RR-25) and clade V also contained potato isolates with one rice isolate (RR-22).


a Cultural and morphological variations among the R. solani potato isolates from intra-field of rice-potato crop rotation on PDA. b Cultural and morphological variations among the R. solani rice isolates from intra-field of rice-potato crop rotation on PDA

Un-weighted pair group method with arithmetic mean dendrogram of R. solani isolates from intra-field of rice-potato rotation based on morphological characters
Anastomosis Reaction of Rhizoctonia solani Isolates
R. solani isolates from rice and potato paired with respective tester AG strains and different reaction types (i.e. C0, C1, C2 and C3) were recorded.
Hyphal Anastomosis Interaction of Potato Isolates
All potato isolates formed C2 type: somatic fusion of perfect anastomosis interactions with the tester isolate AG-3, except one isolate PR9, which failed to show perfect interaction. C3 type or self-anastomosis interactions were not taken into consideration as they represented the interactions between hyphae of the same isolates. The C2 type hyphal fusion frequency of the tested isolates was more than 80%. The selected isolates were identified as members of the group AG-3. Potato isolates were C0 type: no reaction with the tester strain of R. solani AG1-IA from rice.
Hyphal Anastomosis Interaction of Rice Isolates
Most of the rice isolates showed C2: somatic fusions of perfect anastomosis interactions with the tester isolate AG1-IA. The selected isolates were identified as members of the group AG1-IA. All the rice isolates showed C0: no reaction with the tester strain of R. solani AG-3.
Mycelial Compatibility among R. solani Isolates from the Intra-field of Rice-Potato Crop Rotation
The mycelial compatibility assay for R. solani isolates from rice and potato was conducted with a total of 300 combinations (25 paired isolates in all possible combinations and each pairing repeated two times). The observations revealed that in compatible interaction, two paired isolates coalesced freely and fused to form a single colony; further, no demarcation line was observed at contact. In incompatible interaction, clear demarcation line as abundant, tuft and brown patches of aerial growth or distinct zone was visible (Fig. 5a, b). Among all pairing combinations, only 58 pairings showed compatibility reaction and 242 pairings were incompatible (Fig. 6). All of the isolates were self-compatible since there was continuous mycelial growth observed between them. For combinations that showed antagonistic reactions to each other, a thin band of living or dead mycelia was formed. Based on mycelial compatibility, 80.7% non-compatible and 19.3% compatible reactions were observed among the tested isolates. Based on the compatible and incompatible interaction scoring matrix, a dendrogram was generated with UPGMA using the Multi-Variate Statistical Package ver. 3.1 grouped all the R. solani isolates into 3 main clusters (I, II and III). These three clusters were further separated into seven MCGs, viz., MCG A, MCG B, MCG C, MCG D, MCG E, MCG F and MCG G (Fig. 7). MCG A had six isolates of rice (RR-12, RR-13, RR-16, RR-21, RR-23 and RR-24) and MCG B contained four isolates of potato (PR-2, PR-3, PR-5 and PR-6). MCG C and MCG D grouped with 5 and 3 rice isolates, respectively. MCD E had only two isolates of potato, namely, PR-8 and PR-9. MCG F contained three potato isolates (PR-7, PR-10 and PR-12) and one rice isolate (RR-19). MCG G was grouped with two potato isolates (PR-1 and PR-4).


a Mycelial compatibility among rice R. solani isolates from intra-field of rice-potato rotation. b Mycelial compatibility among potato R. solani isolates from intra-field of rice-potato rotation

Macroscopic somatic interaction between different R. solani isolates from intra-field of rice-potato rotation. The categories of interactions:
 : incompatible, when a tuft of hyphae or barrage zone was observed in the line of contact between paired isolates;
: incompatible, when a tuft of hyphae or barrage zone was observed in the line of contact between paired isolates;
 : compatible, when no contact line was observed between paired isolates
: compatible, when no contact line was observed between paired isolates

Un-weighted pair group method with arithmetic mean dendrogram of R. solani isolates from intra-field of rice-potato rotation based on compatibility interaction
Cross-Pathogenicity of R. solani Isolates from Rice-Potato Crop Rotation
Pathogenicity of Potato Isolates on Rice Plants
The results obtained from detached leaf assay in the laboratory and stem inoculation tests conducted under greenhouse conditions revealed that no isolate of potato was able to infect rice as compared with rice isolate which served as a control. Irrespective of the growth stage at which a sclerotium formed, all the 10 potato isolates were unable to infect rice while prominent lesions developed on plants inoculated with rice isolate RR17.
Pathogenicity of Potato and Rice Isolates on Potato Plants
The results of the pot inoculation experiment revealed varying levels of disease incidence (%) and severity of black scurf caused by different rice and potato isolates (Table 3). These differences in virulence patterns among the isolates highlight their varying impacts on the potato cv. Kufri Bahar. The data revealed that 3 out of 10 rice isolates (RR-16, RR-17 and RR-22) were unable to infect potato tubers. However, potato tubers inoculated with the remaining 7 isolates exhibited significantly varied levels of disease incidence and severity of black scurf. The disease incidence of rice isolates varied from 28.9 to 67.6% and disease severity ranged from 13.3 to 37.7%. Of the 7 rice isolates of R. solani producing black scurf symptoms on potato cv. ‘Kufri Bahar’, RR-12 appears to be the most virulent followed by RR-19. The disease incidence of potato isolates varied from 84.4 to 100% and disease severity ranged from 30.7 to 87.1%. Of the 10 potato isolates of R. solani producing black scurf symptoms on potato cv. ‘Kufri Bahar’, PR-10 appears to be most virulent followed by PR-4 and PR-6. In the case of planted-infested tubers, the disease incidence and disease severity were 88.4% and 63.9%, respectively, whereas potato tubers remain healthy in the untreated control. The sclerotia produced by the rice isolates were less firm and adhered loosely to the tuber surface and easily removed by washing tubers in running tap water. However, Rhizoctonia-like atypical symptoms, i.e. corky spot and black crust on tubers, were also observed. In the case of potato isolates, sclerotia formed on tubers were more firm and not removed easily while washing in water.
Fungicide Sensitivity Assay
The sensitivity levels of selected R. solani isolates from both crops against tested fungicides in terms of inhibition percentage and their EC50 and EC90 values were determined using the poisoned food technique. The data revealed the existence of variation among isolates in their response to fungicides. Additionally, R. solani isolates from potato and rice were inhibited differentially by the tested fungicides. In the case of mancozeb 50% + carbendazim 25% WS the maximum inhibition percentage (94.4%) was at 10 ppm in both rice and potato isolates, except one potato isolate PR-9 which showed 87.2% inhibition. However, thifluzamide 24% SC inhibited potato isolates by 86.1 to 90.6% at 10 ppm, while in the case of rice isolates, the inhibition percentage ranged from 32.2 to 62.2%. Mean EC50 and EC90 values of representative isolates of potato were less (6.9 and 11.0 μg/ml) than the rice isolates (12.5 and 47.9 μg/ml) against thifluzamide 24% SC. Similarly, mean EC50 and EC90 values were also higher with rice isolates than potato isolates against mancozeb 50% + carbendazim 25% WS. The inhibitory values indicated that mancozeb 50% + carbendazim 25% WS was the most effective followed by thifluzamide 24% SC. Variation in EC50 and EC90 values of test fungicides against representatives of potato and rice isolates is also evident in Table 4. The EC90 value of mancozeb 50% + carbendazim 25% WS was lowest against isolate PR-10 (9.2 μg/ml) indicating it to be most sensitive to mancozeb 50% + carbendazim 25% WS followed by isolate PR-8 (9.3 μg/ml), while it was maximum for PR-9 (10.5 μg/ml) among representative potato isolates. While mancozeb 50% + carbendazim 25% WS showed the lowest EC90 value of 9.2 μg/ml for rice isolate RR-19, it was the highest in the case of isolate RR-17 (9.3 μg/ml). Thifluzamide 24% SC showed a maximum EC90 value of 12.9 μg/ml in the case of isolate PR-9 and a minimum (10.1 μg/ml) in isolate PR-10 among both potato and rice isolates.
Discussion
In this investigation, total 25 R. solani isolates (14 from rice and 11 from potato) were recovered from an intra-field of rice-potato rotation. All isolates showed typical cultural and morphological characteristics of R. solani as described by Sneh et al. (1991), viz., hyphal branching at a right angle to the constriction found at the point of branching and a septum was always present near the branching junction. Based on cultural and morphological characteristics, considerable variations were observed among the rice and potato isolates. Rice isolates exhibited faster growth on PDA plates, while potato isolates grew slowly to moderately. It was observed that most of the rice isolates exhibited whitish brown mycelial colour, while the mycelial colour of potatoes varied from whitish brown to dark brown. The hyphal width of rice isolates varied from 6.7 to 8.4 μm, while that of potato isolates varied from 8.2 to 9.4 μm.
Considerable variation was observed in sclerotial characteristics of R. solani isolates from both crops as most of the potato isolates initiated sclerotia formation within 96 h after incubation and matured after 144 h, while rice isolates formed sclerotia after 72 h of incubation and matured after 96 h. One potato isolate, namely, PR-5, did not produce any sclerotia. It was observed that the majority of rice and potato R. solani isolates formed micro (≤ 1.25 mm) size sclerotia, and only a few isolates of both crops had macro (> 1.25 mm) size sclerotia. The sclerotial colour of most rice and potato isolates was brown; however, a few isolates also produced black-coloured sclerotia. The majority of potato isolates produced abundant types of sclerotia, while that of rice isolates had moderate sclerotia. A marked difference in the sclerotial formation pattern was observed between R. solani isolates from both hosts and among the isolates from the same host. In most of the rice isolates, sclerotia were distributed in scattered manner, except isolates RR-13 and RR-16, which formed a sclerotial ring towards the periphery, and isolates RR-12 and RR-25 produced ring towards centre; in RR-19 centrally concentrated pattern was observed. Likewise, most of the potato isolates also produced sclerotia in a scattered manner, except in isolates PR-9 and PR-11, in which a centrally concentrated sclerotial pattern was observed and isolate PR-8 formed a sclerotial ring towards centre. Differences in cultural and morphological characters in R. solani isolates from rice and potato have been previously studied by several workers (Goswami et al. 2010; Abdlla et al. 2017). In a study, Gurav et al. (2018) reported the variability in R. solani isolates from potatoes in terms of colony growth, hyphal width and sclerotial formation pattern. They observed that the majority of isolates exhibited a slow growth rate. Similarly, Thind and Aggarwal (2008) also supported that R. solani isolates from rice grew faster than of potato isolates. Recently, Betancourth-Garcia et al. (2021) studied 494 isolates of R. solani collected from potatoes and found significant differences in cultural and sclerotial characteristics among the isolates. They observed that 96.6% of the isolates had an average of 16.6 mm/day growth rate and the mean hyphal width was 9.7 μm. Sclerotial formation started at 6 days and dispersed as well as peripheral sclerotial arrangement with cream to brown colour was observed. The anastomosis behaviour test indicates that tested isolates showed somatic fusion or perfect anastomosis (C2 reaction) with their respective tester isolates with potato isolates belonging to AG-3 and rice isolates to AG1-IA group. It has been reported that generally, isolates from potatoes belong to AG-3 (Kanetis et al. 2016; Patil et al. 2018) while those from rice belong to AG1-IA (Zheng et al. 2013).
Mycelial interaction in filamentous fungi is a self and non-self-recognition system that is controlled by multiple loci and is a useful and easy way to categorize intraspecific heterogeneity. However, information about its genetic mechanisms is limited (Glass and Kaneko 2003). Isolates that are different at one or more of these loci will not show perfect fusion. In this study, the mycelial interaction of the R. solani isolates from both hosts was tested on PDA medium with optimum growth conditions. The results revealed that most of the isolates showed compatible reactions when paired with other isolates. Based on mycelial compatibility, 80.7% of the pairings showed compatibility and 19.3% of the pairings exhibited non-compatible reactions. Based on mycelial interaction, all isolates grouped into seven different MCGs, viz., MCG A, MCG B, MCG C, MCG D, MCG E, MCG F and MCG G. Hussain et al. (2014) studied the compatibility/incompatibility of 20 R. solani isolates from potato and found that about 72.1% of the pairings were compatible, while 27.9% of the pairings were incompatible. However, in a study, Rosa et al. (2012) found about 91% incompatible pairings and 9% compatible pairings with 433 R. solani isolates.
The selected isolates of rice and potato were subjected to cross-infectivity determination under lab and greenhouse pot experiments. The results showed that potato isolates did not infect rice plants. However, R. solani isolates from rice showed varied degrees of virulence on potatoes with a few isolates unable to infect potato tubers, whereas others developed black scurf symptoms of different severity levels. Rhizoctonia-like atypical symptoms, i.e. corky spot and black crust, were also observed on potato tubers. Distinct variations were also observed in the pathogenicity and virulence of potato isolates, which may be isolate-dependent rather than AG-dependent. It was also observed that isolates collected from stem canker-infected potato plants were more pathogenic than that of black scurf isolates. Similar findings indicating differences in pathogenicity and host range of R. solani isolates have been documented by several workers. In a study, Khandaker et al. (2008) studied the disease reaction of different hosts against R. solani potato isolates and reported that pulse crops were less susceptible to R. solani compared to horticultural crops. Thind and Aggarwal (2008) collected the R. solani isolates from rice-potato rotation and cross-infectivity results revealed that potato isolates were unable to infect rice plants whereas potato isolates showed varied virulence on potatoes. Ganeshamoorthi and Dubey (2015) reported the variable virulence of R. solani isolates from Cicer arientum on Vigna mungo, C. arientum and V. radiata. Recently, Esfahani (2020) reported the variable virulence of stem canker and black scurf R. solani isolates on potatoes.
The application of chemical fungicides is the most widely used and effective method to manage Rhizoctonia diseases. As there is more than one AG associated with Rhizoctonia disease of potato, these AGs displayed varying sensitivity to fungicides; therefore, the identification of the group(s) causing disease in any particular field is crucial to fungicide selection (Kataria et al. 1991; Kataria and Gisi 1999). For example, Rhizoctonia AGs 1, 3 and 5 are moderately affected by aromatic hydrocarbon fungicides, while AGs 2–1, 4, 7 and 8 are least sensitive. R. solani isolates were highly sensitive to flutolanil, iprodione and pencycuron, except AG-5 (Campion et al. 2003). In this study, sensitivity levels of different R. solani isolates from intra-field of rice-potato rotation were evaluated in vitro against two commercially used fungicides, i.e. thifluzamide 24% SC and mancozeb 50% + carbendazim 25% WS. The results showed a considerable level of variability in different R. solani isolates from both crops concerning their sensitivity to different fungicides. The high values of EC50 and EC90 were observed in rice isolates than in potato isolates against both evaluated fungicides. Variations in fungicide sensitivity levels of R. solani isolates against various fungicides have been previously reported by several workers (Kataria and Verma 1989; Lehtonen et al. 2007). Significant variation was observed in R. solani isolates from rice and potato against carbendazim, carboxin, pencycuron, propiconazole and validamycin (Thind and Aggarwal 2008). Ajayi-Oyetunde et al. (2017) demonstrated that R. solani isolates collected from soybean were extremely sensitive to penflufen and sedaxane (SDHI fungicides) while either extremely sensitive or moderately sensitive to ipconazole and prothioconazole (DMI fungicides). Muzhinji et al. (2018) studied the sensitivity level of 131 Rhizoctonia isolates from different AGs to six fungicides and reported that all AGs were sensitive to tolclofos-methyl and fludioxonil, and variation in sensitivity to pencycuron, iprodione and benomyl.
Conclusion
In this study, intra-field variability among R. solani isolates from rice-potato rotation was assessed based on morphological and pathological characterization. Besides, sensitivity profiling of R. solani isolates was also examined against commercially used fungicides. The results revealed that AGI-IA and AG-3 were the most prevalent anastomosis groups in rice and potato, respectively. Cross-infectivity experiments revealed that R. solani isolates from potatoes were not pathogenic to rice. Fungicide sensitivity study revealed that R. solani isolates of potato and rice exhibited variable sensitivity levels to different fungicides. Therefore, under crop rotation conditions, regular surveillance for the occurrence of R. solani AGs on potato crops is needed. The outcomes of this study will be helpful in developing management strategies for effective control of Rhizoctonia diseases in potatoes.
References
Abdlla ME, Tohmy AM, Rashid IA, Ahmed NE (2017) Morphological and molecular characterization of potato black scurf disease (Rhizoctonia solani) in Egypt. J Plant Protect Pathol 8(9):473–478
Ajayi-Oyetunde OO, Butts-Wilmsmeyer CJ, Bradley CA (2017) Sensitivity of Rhizoctonia solani to succinate dehydrogenase inhibitor and demethylation inhibitor fungicides. Plant Dis 101:487–495
Amaradasa BS, Lakshman DK, McCall D, Horvath BJ (2014) In vitro fungicide sensitivity of Rhizoctonia and Waitea isolates collected from turfgrasses. J Environ Hortic 32:126–132
Betancourth-Garcia CA, Castro-Caicedo BL, Quiroz-Ojeda C, Sanudo-Sotelo B, Florez-Casanova C, Salazar-Gonzalez C (2021) Morphology and pathogenicity of Rhizoctonia solani Kuhn associated with potato black scurf in Narino (Colombia). Revista Colombiana De Ciencias Horticolas 15(1):e11821
Calleros GV, Portugal VO, Carling DE (2000) Anastomosis groups of Rhizoctonia solani on potato in Central Mexico and potential for biological and chemical control. Am J Potato Res 77:219–224
Campion C, Chatot C, Peraton B, Andrivon D (2003) Anastomosis groups, pathogenicity and sensitivity to fungicides of Rhizoctonia isolates collected in potato fields in France. Eur J Plant Pathol 109:983–992
Carling DE (1996) In Rhizoctonia species: taxonomy, molecular biology, ecology, pathology and disease control. Springer, The Netherlands, pp 37–47
Carling DE, Kuninaga S, Brainard KA (2002) Hyphal anastomosis reactions, rDNA-internal transcribed spacer sequences, and virulence levels among subsets of Rhizoctonia solani anastomosis group-2 (AG-2) and AG-BI. Phytopathology 92:43–50
Carling DE, Sumner DR (1992) In Methods for research on soilborne phytopathogenic fungi. (eds. Singleton II, Mihail JD, Rush CM). American Phytopathological Society
Ceresini PC, Shew HD, Vilgalys RJ, Cubeta MA (2002) Genetic diversity of Rhizoctonia solani AG-3 from potato and tobacco in North Carolina. Mycologia 94:437–449
Chahal KS, Sokhi SS, Rattan GS (2003) Investigation on sheath blight of rice in Punjab. Indian Phytopathol 56:22–26
Chaudhary S, Sagar S, Lal M (2019) A comprehensive overview on sheath blight disease of rice and its management. Agricultura 3–4:145–174
Chaudhary S, Lal M, Sagar S, Tyagi H, Kumar M, Sharma S, Chakrabarti SK (2020a) Genetic diversity studies based on morpho-pathological and molecular variability of the Sclerotinia sclerotiorum population infecting potato (Solanum tuberosum L.). World J Microbiol Biotechnol 36:177
Chaudhary S, Sagar S, Lal M, Tomar A, Kumar V, Kumar M (2020b) Biocontrol and growth enhancement potential of Trichoderma spp. against Rhizoctonia solani causing sheath blight disease in rice. J Environ Biol 41:1034–1045
Chaudhary S, Lal M, Sagar S, Sharma S, Kumar M (2023) Black scurf of potato: insight into biology, diagnosis, detection, host-pathogen interaction, and management strategies. Trop Plant Pathol. https://doi.org/10.1007/s40858-023-00622-4
Chen Y, Zhang AF, Wang WW, Zhang Y, Gao TC (2012) Baseline sensitivity and efficacy of thifluzamide in Rhizoctonia solani. Ann Appl Biol 161:247–254
Dong W, Li Y, Duan C, Li X, Naito S, Conner RL, Yang G, Li C (2017) Identification of AG-V, a new anastomosis group of binucleate Rhizoctonia spp. from taro and ginger in Yunnan province. Eur J Plant Pathol 148:895–906
Esfahani MN (2020) Genetic variability and virulence of some Iranian Rhizoctonia solani isolates associated with stem canker and black scurf of potato (Solanum tuberosum L.). J Plant Protect Res 60:21–30
Ganeshamoorthi P, Dubey SC (2015) Morphological and pathogenic variability of R. solani isolates associated with wet root rot of chickpea in India. Legume Res 38:389–395
Glass NL, Kaneko I (2003) Fatal attraction: nonself recognition and heterokaryon incompatibility in filamentous fungi. Eukaryot Cell 2:1–8
Goswami BK, Bhuiyan KA, Mian IH (2010) Morphological and pathogenic variations in the isolates of Rhizoctonia solani in Bangladesh. Bangladesh J Agri Res 35:375–380
Grower RK, Moore JD (1962) Toxicometric studies of fungicides against brown rot organisms Sclerotinia fructicola and S. laxa. Phytopathology 52:876–880
Gurav NP, Singh S, Basavaraj K, Mehta N, Madane AJ (2018) Characterization of cultural and morphological variability in Rhizoctonia solani isolates associated with black scurf of potato. Int J Curr Microbiol App Sci 7(7):2438–2445
Hussain A, Awan MS, Iqbal M, Anees M (2014) Variability among the potato sclerotial isolates of Rhizoctonia solani of mountain region, Gilgit-Baltistan, Pakistan. Afr J Microbiol Res 8:2849–2860
Kanetis L, Tsimouris D, Christoforou M (2016) Characterization of Rhizoctonia solani associated with black scurf in Cyprus. Plant Dis 100(8):1591–1598
Kataria HR, Gisi U (1999) Selectivity of fungicides within the genus Rhizoctonia. In: Lyr H, Russell PE, Dehne HW, Sisler HD (eds) Modern fungicides and antifungal compounds. Intercept, Andover, U.K., pp 421–429
Kataria HR, Verma PR (1989) Variations in the sensitivity to fungicides among different anastomosis groups of Rhizoctonia solani. Can J Plant Pathol 11:192
Kataria HR, Verma PR, Gisi U (1991) Variability in the sensitivity of Rhizoctonia solani anastomosis groups to fungicides. J Phytopathol 133:121–133
Kaur J, Kaur A (2013) Morphological and molecular variability in Rhizoctonia solani isolate infecting potato and rice. Plant Dis Res 28(2):163–168
Khandaker M, Khair A, Bhuiyan MKA (2008) Disease reaction of different crops against virulent potato isolates of Rhizoctonia solani Kuhn. Bangladesh J Bot 37:75–80
Kroland WC, Stanghellini ME (1988) Clean slide technique for the observation of the anastomosis and nuclear condition of Rhizoctonia solani. Phytopathology 78:820–822
Lal M, Kumar A, Chaudhary S, Singh RK, Sharma S, Kumar M (2022) Antagonistic and growth enhancement activites of native Pseudomonas spp. against soil and tiuber-borne diseases of potato (Solanum tuberosum L.). Egypt J Biol Pest Control 32:22
Lehtonen MJ, Ahvenniemi P, Wilson PS, German-Kinnari M, Valkonen JPT (2007) Biological diversity of Rhizoctonia solani (AG-3) in a northern potato-cultivation environment in Finland. Plant Pathol 57:141–151
MacNish GC, Carling DE, Brainard KA (1997) Relationship of microscopic and macroscopic vegetative reactions in Rhizoctonia solani and the occurrence of vegetatively compatible populations (VCPs) in AG8. Mycol Res 101:61–68
Muzhinji N, Woodhall JW, Truter M, van der Waals JE (2018) Variation in fungicide sensitivity among Rhizoctonia isolates recovered from potatoes in South Africa. Plant Dis 102:1520–1526
Ogoshi A (1987) Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kuhn. Annu Rev Phytopathol 25:125–143
Olaya G, Sarmiento L, Edlebeck K, Buitrago C, Sierotzki H, Zaunbrecher J, Tally A (2013) Azoxystrobin (QUI) resistance monitoring of Rhizoctonia solani isolates causing rice sheath blight in Louisiana. Abstract, APS-MSA Joint Meeting, Austin, Texas.
Ou SH (1985) Rice diseases, 2nd edn. Kew Surrey, UK, Commonwealth Mycological Institute.
Patil V, Girimalla V, Sagar V, Bhardwaj V, Chakrabarti S (2018) Draft genome sequencing of Rhizoctonia solani anastomosis group 3 (AG3-PT) causing stem canker and black scurf of potato. Am J Potato Res 95:87–91
Rangaswami G, Mahadevam A (2004) Diseases of crop plants in India, 4th edn. Prentice Hall of India Pvt. Ltd., New Delhi, pp 65–66
Rosa LF, Johan MC, Paulo C, Celsa GD (2012) Rhizoctonia solani AG-3PT is the major pathogen associated potato stem canker and black scurf in Colombia. Agro Colombiana 30:204–213
Sharon M, Sneh B, Kuninaga S, Hyakumachi M, Naito S (2008) Classification of Rhizoctonia spp. using rDNA-ITS sequence analysis supports the genetic basis of the classical anastomosis grouping. Mycoscience 49:93–114
Simay EI (1993) Pathogenicity of Rhizoctonia isolates on faba bean. FABIS Newslett 32:31–33
Singh V, Singh US, Singh KP, Singh M, Kumar A (2002) Genetic diversity of Rhizoctonia solani isolates from rice: differentiation by morphological characteristics, pathogenicity, anastomosis behaviour and RAPD fingerprinting. J Mycol Plant Pathol 32:332–344
Sneh B, Burpee L, Ogoshi A (1991) Identification of Rhizoctonia species. APS Press
Thind TS, Aggarwal A (2008) Characterization and pathogenic relationship of Rhizoctonia solani isolates in a potato-rice system and their sensitivity to fungicides. J Phytopathol 156:615–621
Zhang CQ, Yuan SK, Sun HY, Qi ZQ, Zhou MC, Zhu GN (2007) Sensitivity of Botrytis cinerea from vegetable greenhouse to boscalid. Plant Pathol 56:646–653
Zhao C, Li Y, Liu H, Li S, Han C, Wu X (2019) A binucleate Rhizoctonia anastomosis group (AG-W) is the causal agent of sugar beet seedling damping-off disease in China. Eur J Plant Pathol 155:53–69
Zheng A, Lin R, Zhang D, Qin P, Xu L, Ai P et al (2013) The evolution and pathogenic mechanism of the rice sheath blight pathogen. Nature Commun 4:1424
Acknowledgements
The present work is based upon research supported by the research grant (EEQ/2016/000298) of the Science and Engineering Research Board (SERB)-Department of Science and Technology (DST), Government of India.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lal, M., Yadav, S., Chaudhary, S. et al. Morphological and Pathological Variability of Intra-field Rhizoctonia solani Isolates in a Rice-Potato Rotation and their Sensitivity to Fungicides. Potato Res. (2024). https://doi.org/10.1007/s11540-024-09746-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11540-024-09746-9