
Abstract
Arsenic (As) is the cause for concern worldwide due to its high toxicity. Its presence in agricultural soils and groundwater adversely affects soybean (Glycine max L.) growth and yield and also endangers food safety. Plant growth-promoting rhizobacteria (PGPR) could be used as part of cost-effective and eco-friendly strategies to mitigate As phytotoxicity. However, simple inoculation of soybean with PGPR Bradyrhizobium japonicum E109 (E109), a common practice in Argentina, is not effective in counteracting the effects of As exposure. Our aim was to assess whether the response of soybean to arsenate (AsV) and arsenite (AsIII) could be helpfully modulated by co-inoculating E109 with the free-living PGPRs Azospirillum brasilense Cd (Cd) or Bacillus pumilus SF5 (SF5). Co-inoculation with E109 + SF5 alleviated As-induced depletion of chlorophyll a and b, and carotenoid content, reaching an increase of 26, 28 y 31%, respectively. It also enhanced nodulation (15–19%) under As exposure. E109 + Cd and E109 + SF5 induced changes in the antioxidant system, which could be related to the maintenance of redox homeostasis. Moreover, As accumulation was reduced by 53% in aerial parts of plants inoculated with E109 + Cd, and by 16% in the roots of those inoculated with E109 + SF5. The strains selected show interesting potential for the development of biotechnological schemes to improve soybean yield while guaranteeing safer food production.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Arsenic (As) is a toxic metalloid that occurs naturally in certain soils and in groundwater. It is found in both organic and inorganic forms, although the latter are prevalent (Singh and Srivastava 2020; Jia et al. 2021). The relative presence of different As species depends on the pH and redox potential of the surrounding environment. Microorganisms are also known to play a crucial role in the As geocycle (Ben Fekih et al. 2018).
The most common inorganic forms of As that adversely affect plants are arsenate (AsV) and arsenite (AsIII). Given that both can become accumulated in plant tissues, their presence in cultivable lands is cause for concern. Aquaporins mediate the uptake of AsIII and phosphate transporters that of AsV. The metalloid is then transferred to the shoots by xylem loading (Bali and Sidhu 2021). Once inside the plant, it negatively impacts morphological, physiological, biochemical, and metabolic processes. More precisely, it can disturb root-shoot biomass, chlorophyll content, photosynthetic rate, gas exchange, water content, and sugar metabolism as well as trigger the production of reactive oxygen species (ROS) (Abbas et al. 2018; Bali and Sidhu 2021). Since ROS are able to engage in reactions with proteins, lipids, carbohydrates, and DNA, they can cause irreparable damage unless scavenged through activation of the antioxidant system (Banerjee and Roychoudhury 2019). This is why enzymatic and non-enzymatic antioxidants constitute one of the main defense mechanisms that determine As tolerance (Zulfiqar and Ashraf 2022). Furthermore, As interferes with nodulation and nitrogen fixation in legume-Rhizobium interactions (Armendariz et al. 2019; Pajuelo et al. 2019) and can thus reduce crop yield and affect food safety (Bustingorri and Lavado 2014). Several plant food sources, in fact, have been found to contain harmful levels of As (Upadhyay et al. 2019).
Soybean (Glycine max L.), a major crop worldwide, is mainly produced in the USA, Brazil, and Argentina (Goonde and Ayana 2021). Because of its ability to grow under a wide range of environmental conditions, it has been introduced into marginal lands where it is frequently exposed to As. Evidence of the adverse effects of As has therefore been found in this plant species, both in studies conducted by our lab (Talano et al. 2013; Armendariz et al. 2016; Vezza et al. 2018, 2021) and by other authors (Bustingorri and Lavado 2014; Chandrakar et al. 2017; Fatima et al. 2020; Gálusová et al. 2020).
Several physicochemical methods have been developed to remediate As, but most of them have practical limitations. In addition to being costly, they may generate toxic sludge, be inefficient under natural conditions, and pose operational problems. By contrast, bioremediation is sustainable, cost-effective, and reliable (Irshad et al. 2021). The potential of As-tolerant plant growth-promoting rhizobacteria (PGPRs) as biofertilizers, phytostimulants, biopesticides, and bioremediators may be harnessed to improve plant performance and contribute to the mitigation of As (Molina et al. 2021; Kumar et al. 2022). Well-chosen strains could induce the antioxidant defense system, repair photosynthetic activity, improve nutritional status, and help to re-establish symbiotic interaction (Mondal et al. 2021). Microbial activity can also strongly influence the bioavailability, adsorption/absorption, and translocation of As within the soil–plant system (Srivastava and Shukla 2019; Khan et al. 2021). In short, appropriately selected PGPRs could potentially be used alone or as part of consortia to regulate stress caused by As in host plants.
In Argentina, soybean is frequently inoculated with formulations based on Bradyrhizobium japonicum E109 (E109), designed to boost yield. However, previous studies by our laboratory have shown that E109 is not a sufficient strategy to assist plant growth in environments with moderate-to-high As concentrations. Certain benefits have been reported for the co-inoculation of rhizobium with other strains in leguminous plants subjected to regular and stressful conditions. For instance, the application of Azospirillum brasilense strains together with rhizobium was observed to increase biomass, nitrogen content, flavonoid signals, nod-gene transcription, and grain yield (Dardanelli et al. 2008; Galindo et al. 2018; Rondina et al. 2020; Filipini et al. 2021). Similar results were obtained after co-inoculating rhizobium and Bacillus pumilus strains (Stefan et al. 2010; Hasibuan et al. 2021). Specifically, Azospirillum brasilense Cd (Cd) is a collection strain used for commercial inoculant formulation, while Bacillus pumilus SF5 is an endophytic strain isolated from the rhizosphere under drought stress and exhibits interesting PGPR properties and tolerance to a wide range of As concentrations (Forchetti et al. 2007; Vezza et al. 2020). However, nothing is known about the mechanisms underlying these properties or the strains’ ability to contribute to plant success in actual As-contaminated fields.
Here, we explored the effects of co-inoculating soybean with E109 + Cd and E109 + SF5 on the plant’s response to As during the early vegetative stage. Exposure to AsV and AsIII was considered, since they are the most frequent chemical forms in the environment and they differ in phytotoxicity. Parameters associated with As stress were analyzed, such as those related to growth, nodulation, photosynthetic pigments, the antioxidant system, and As accumulation in plant tissues. Our working hypothesis was that co-inoculation with E109 + Cd and E109 + SF5 could be a better strategy than E109 simple inoculation on soybean early development, nodulation, and limitation of As uptake in metalloid-contaminated environments. The findings are encouraging enough to consider the selected strains for future research on a larger and deeper scale.
Materials and methods
Biological material
Soybean (Glycine max L.) cv. Don Mario 4670 seeds were surface disinfected as described by Vezza et al. (2018). The bacterial strains used for seed inoculation were Bradyrhizobium japonicum E109 (E109), Azospirillum brasilense Cd (Cd), and Bacillus pumilus SF5 (SF5). E109 and Cd are collection strains used to produce commercial inoculant formulations worldwide. SF5 is an endophytic bacteria isolated from sunflower (Helianthus annuus L.) roots grown under drought (Forchetti et al. 2007), which was gently provided by Dr. S. Alemano.
Experimental design and growth conditions
Experiments were carried out in a completely randomized factorial design with three seed inoculation conditions: E109 simple inoculation and co-inoculations with E109 + Cd or E109 + SF5, and three As treatments: without As (control), and exposure to AsV or AsIII as described below.
Soybean inoculation with E109 is a very widespread practice in Argentina, and this strain remains in the soil from one season to the next (Narożna et al. 2015), so it constituted the control condition for the comparison to co-inoculation strategies. Non-inoculated condition compared with E109 simple inoculation has already been evaluated in our previous works (Talano et al. 2013; Armendariz et al. 2019).
E109 was grown in yeast extract mannitol (YEM) medium (Vincent 1970), while Luria–Bertani (LB) (Bertani 1951) medium was used for the growth of Cd and SF5, at 28 °C and 150 rpm. Cultures in the exponential growth phase were adjusted to 1 × 10−9, 1 × 10−6, and 1 × 10−8 CFU mL−1 (colony forming units per milliliter), respectively. Inoculation doses were adjusted to a final volume of 250 μL for each 10 g of seeds, using a mixture of equal volumes of each strain for co-inoculations. Inoculated seeds were placed into pots containing sterile perlite as an inert substrate to avoid interferences with As treatment. They were irrigated by capillarity with 1/4 Hoagland solution without nitrogen and kept in a growth chamber (model E15, Conviron, Winnepeg, Canada) under controlled conditions: 14-h light/10-h dark cycle, 200 μmol m−2 s−1 light intensity, 28 °C, and 80% relative humidity. Seven days after inoculation, plants were irrigated with water without As (control), supplemented with 25-μM sodium arsenate (Na2HAsO4.7H2O) (SIGMA) (AsV) or 25-μM sodium arsenite (NaAsO2) (SIGMA) (AsIII). The As concentration used was selected because it is found in many rural areas around the world (Mariño et al. 2020; Singh and Srivastava 2020) and causes alterations in soybean growth, according to our first report (Talano et al. 2013). The plants were evaluated at 35 days of growth. Harvested plant tissues were homogenized with liquid N2 and immediately kept at − 80 °C until their further use.
Plant growth and nodulation
Dry (DW) and fresh weight (FW) of total aerial part, leaves, and roots were registered. In addition, the number and FW of total nodules and those located in the main root were evaluated.
Photosynthetic pigments
The determination of photosynthetic pigments was performed according to Dere et al. (1998). For this, 100 mg of fresh leaf tissues were homogenized with 1 mL of 96% methanol. After centrifugation at 5000 rpm for 7 min, supernatant absorbance was measured at 662 nm, 645 nm, and 470 nm. The content of chlorophyll a (Ca), chlorophyll b (Cb), and carotenoids (Cx+c) was calculated using the following formulas: Ca = 15.65 × A666 – 7.34 × A653; Cb = 27.05 × A653 – 11.21 × A666; Cx+c = 1000 × A470 – 2.86 × Ca – 129.2 × Cb/245. The results were expressed as mg g−1 FW.
Antioxidant system
Antioxidant enzyme activity
Enzyme extraction was performed by homogenizing plant tissues (leaves and roots separately) with buffer solution (1:10 w/v) [50 mM KH2PO4/K2HPO4 buffer pH 7.8, 0.5 mM EDTA, and polyvinylpolypyrrolidone (PVPP)] and subsequent centrifugation at 10,000 rpm and 4 °C for 30 min. After that, the supernatant was used for assays of total peroxidase (Px), ascorbate peroxidase (APx), and superoxide dismutase (SOD) activities. Total Px activity was determined spectrophotometrically with o-dianisidine and H2O2 as substrates, following the appearance of a colored product [bis (3,3-dimethoxy-4amino) azo-biphenyl] (ε 11.3 mM−1 cm−1) at 470 nm (Agostini et al. 1997). One Px unit was defined as the amount of enzyme that generates 1 μmol of the colored product after 1 min of reaction. APx activity was determined following the oxidation of L-ascorbic acid (ε 2.8 M−1 cm−1) at 290 nm (Hossain and Asada 1984). One APx unit was defined as the amount of enzyme that produced the oxidation of 1 μmol of ascorbic acid after 1 min of reaction. SOD activity was determined according to Beauchamp and Fridovich (1973) with some modifications. An SOD unit was defined as the amount of enzyme required to cause 50% inhibition of the rate of photochemical reduction of nitro-blue tetrazolium (NBT) at 560 nm (Armendariz et al. 2016).
Total antioxidant activity
Total antioxidant activity (TAA) was determined as the capacity of radical scavenging against stable DPPH (2,2-diphenyl-1-picrylhydrazyl) according to Brand-Williams et al. (1995) with some modifications. Leaf and root tissues (200 mg) were homogenized separately with 1 mL of 50% (v/v) methanol, incubated in ice for 2 h (vortexed every 30 min) and centrifuged at 10,000 rpm and 4 °C for 15 min. The supernatants (15 μL and 80 μL from leaf and root extracts, respectively) were mixed with a necessary amount of 60 μM DPPH/methanol solution to complete a final volume of 1 mL and then incubated in the dark for 30 min at room temperature. The absorbance was measured at 515 nm before and after incubation. Finally, TAA was calculated and expressed as follows: DPPH inhibition (%) = [(IA-FA)/IA] × 100, where IA is the initial absorbance (before incubation) and FA is the final absorbance (after incubation).
Total As concentration
Total As concentration in soybean leaves and roots was measured using homogenized oven-dried material. For this, an optimized method comprising flow injection hydride generation atomic absorption spectroscopy (FI-HGAAS) was used, according to Sigrist et al. (2016). Data were expressed as μg As per g−1 DW, and translocation factor (TF) was calculated as follows: TF (%) = (As concentration in leaves/As concentration in roots) × 100.
Statistical analysis
The data were submitted to normality and homogeneity of variance assumptions using Shapiro–Wilk and Levene tests, respectively. When these assumptions were confirmed, the comparisons between the different treatments were carried out using an analysis of variance (ANOVA), followed by a comparison of multiple treatment levels using Tuckey’s test. Differences were considered to be significant for p < 0.05. InfoStat software (v. 2012e; InfoStat, National University of Cordoba, Argentina) was used for this.
Results
Plant growth and nodulation
Growth-related parameters were evaluated in soybean plants during the early vegetative stage (V5). As a result of exposure to As, biomass production was around 25–39% lower in aerial parts and 16–20% lower in roots compared to the control (without As) (Table 1). Co-inoculation with E109 + Cd and E109 + SF5 did not disrupt plant growth: The values obtained were the same as those for simple inoculation with E109 (Table 1).
Nodules on the main root (Fig. 1a) and total nodules (Fig. 1b) were also registered. The amount of nodules on the lateral roots was very few, and therefore, it was not reported here. The adverse effects of As were more pronounced on the nodules’ FW than on their number. In plants inoculated only with E109, the FW of the main root nodules was reduced by 20 and 15% in the presence of AsV and AsIII, respectively (Fig. 1a). Co-inoculation led instead to increased nodule FW (17% for E109 + Cd and 14% for E109 + SF5), in comparison to simple inoculation under AsV treatment. Still, co-inoculation with E109 + SF5 in plants exposed to AsIII, specifically, caused a more relevant increase of 20% with respect to simple inoculation (Fig. 1a).

Nodule FW and number from the main root (a) and all the roots (total) (b) in inoculated soybean plants exposed to AsV and AsIII. Results represent the mean ± SE (n = 15–20). Different capital letters indicate significant differences between inoculation conditions for the same As treatment, while different lowercase letters indicate significant differences between As treatment for the same inoculation condition (Tukey test, p ≤ 0.05)
Photosynthetic pigments
The presence of As considerably reduced the content of chlorophyll a (Fig. 2a) and b (Fig. 2b) in plants inoculated only with E109. The effects were more severe with AsV (28% reduction in type a and 25% reduction in type b) than with AsIII (19 and 16% less, respectively). There was also a decrease in carotenoid content (of around 25%) under AsV treatment (Fig. 2c). In plants exposed to AsV, small increases in the photosynthetic pigment content were obtained after co-inoculation with E109 + Cd, compared to E109 on its own. More significant advantages were achieved with E109 + SF5, since these plants had a higher content of chlorophyll a (26 and 13%), b (28 and 14%), and carotenoids (31 and 9%) compared to those inoculated with E109 under AsV and AsIII stress, respectively (Fig. 2).

Photosynthetic pigments in soybean plants exposed to AsV and AsIII and under different inoculation conditions. Results represent the mean ± SE (n = 6). Different letters indicate significant differences (Tukey test, p ≤ 0.05)
Antioxidant system
The activity of antioxidant enzymes (APx, Px, and SOD) and TAA was estimated in leaves (Fig. 3a, c, e and g) and roots (Fig. 3b, d, f and h). Differences were found between As-stressed and control plants, and in turn, the responses exhibited variations depending on the inoculation strategy applied.

Antioxidant response in leaves (a, c, e, g) and roots (b, d, f, h) in soybean plants exposed to AsV and AsIII and under different inoculation conditions. Results represent the mean ± SE (n = 6). Different letters indicate significant differences (Tukey test, p ≤ 0.05)
In leaves, APx increased significantly in E109-inoculated plants exposed to either AsV or AsIII. When the combination E109 + Cd was used, APx rose only under AsV treatment, and this increment was greater in magnitude than that observed after simple inoculation with E109 (Fig. 3a). Both chemical forms of As led to an increase in Px activity with respect to the control under all inoculation conditions, but similar to what happened with APx (Fig. 3a), the enhancement was greater with E109 + Cd under AsV treatment (Fig. 3c). In general, SOD did not vary in any relevant way from one condition to the other (Fig. 3e). Regarding TAA, higher values were obtained through inoculation with E109 in plants exposed to AsV, and with E109 + Cd in plants exposed to AsIII. The combination E109 + SF5, for its part, rendered higher TAA levels under both As treatments (Fig. 3g).
In roots, APx decreased significantly in plants inoculated with E109 and E109 + Cd under AsV treatment, while activity values in those inoculated with E109 + SF5 were similar to the control (Fig. 3b). No significant variations under the different conditions were observed for Px (Fig. 3d). Co-inoculation with E109 + SF5 improved SOD activity upon exposure to AsV and AsIII. The use of E109 + Cd also enhanced the activity of this enzyme under AsIII treatment (Fig. 3f). A similar trend was found for TAA, which was higher in plants co-inoculated with E109 + SF5 and exposed to AsV (Fig. 3h).
Total As concentration
Total As content was determined in aerial parts (Fig. 4a) and roots (Fig. 4b). The metalloid was more accumulated in the latter than in the former. In plants inoculated with E109 alone, the TF was 5.4% upon AsV exposure and 5.7% upon AsIII exposure (Fig. 4c). Co-inoculation with E109 + Cd reduced the As content by 53% in aerial parts with respect to E109-inoculated plants under AsV treatment (Fig. 4a). Accordingly, TF was also lower (2.7%) under these conditions (Fig. 4c). Moreover, the roots of plants inoculated with E109 + SF5 and treated with AsIII featured around 16% less total As than those otherwise inoculated (Fig. 4b).

As concentration in leaves (a) and roots (b) and TF (c) in inoculated soybean plants exposed to AsV and AsIII. Results represent the mean ± SE (n = 3). Different letters indicate significant differences (Tukey test, p ≤ 0.05)
Discussion
The presence of As in the environment poses a serious risk to the growth of crops such as soybean, and also in terms of food-chain contamination. In agricultural soils with high levels of As, solutions to this issue could take the form of inoculation strategies based on the use of PGPRs, such as those tested here: E109, E109 + Cd, and E109 + SF5.
In agreement with previous reports, we observed that soybean plants exposed to stress by AsV and AsIII were impaired in their growth in comparison to the respective controls (Table 1) (Talano et al. 2013; Armendariz et al. 2016; Vezza et al. 2019). In addition, a significant reduction in nodulation was detected after inoculation with E109 (Fig. 1). Arsenic causes alterations in photosynthesis, nutrient uptake, phytohormone balance, and oxidative status, all of which contribute to deficient plant development (Bali and Sidhu 2021; Khan et al. 2021). The application of strains Cd and SF5 in combination with E109 did not affect plant growth (Table 1), but did slightly improve nodulation, mainly in plants exposed to AsIII and inoculated with E109 + SF5 (Fig. 1). The impact of PGPRs on biometric parameters could largely depend on crop management practices, environmental conditions, and the plant’s phenological stage. The scope of the present study was limited to the early vegetative stage, since the assay was conducted in a growth chamber. When it comes to other stages, Stefan et al. (2010) found the inoculation of soybean with B. pumilus Rs3 to have more marked effects during flowering and fruiting. Studies on the inoculation of several crops with Azospirillum, on the other hand, have come up with multi-causal variable results, as in the case of the soybean-A. brasilense association analyzed by Cassán et al. (2020).
Assessing photosynthetic pigment content is a good method to indirectly estimate plant productivity. Chlorophyll metabolism is usually sensitive to changes in the environment and in the plant’s physiological state, so pigment content could be an indicator of how the plant responds to stress (Sharma et al. 2020). The effects of metal(loid)s depend on the chemical species in question. For instance, soybean plants exposed to AsV experienced more noticeable photosynthetic pigment depletion than those under AsIII treatment (Fig. 2). This becomes more evident after prolonged periods of exposure and intensifies when As concentrations increase (Armendariz et al. 2016). Shahzad et al. (2021) did not detect symptoms of chlorosis or necrosis in maize plants exposed to cadmium and inoculated with Bacilus pumilus, which suggests that appropriate inoculants can alleviate the damage wrought by exposure to metals. Likewise, soybean plants treated with AsV had markedly higher levels of chlorophyll (Fig. 2a and b) and carotenoids (Fig. 2c) when co-inoculated with E109 + SF5 than when receiving only E109. This co-inoculation strategy contributes to counteracting the harmful influence of As, since pigment content did not significantly differ between the control (without As) and the plants treated with either form of As (Fig. 2). The E109 + Cd co-inoculation also led to improvements in pigment content over inoculation with E109, although the differences were not statistically significant (Fig. 2). The dependence of photosynthetic pigment content on the presence and ratio of mineral elements was established for different crops (Bojović and Stojanović, 2005), and PGPRs could be responsible for increased mineral bioavailability. Phosphate solubilization and siderophore production have been reported for SF5 and Cd, respectively, and both strains have shown nitrogen fixation capacity (Forchetti et al. 2007; Vezza et al. 2020). These properties may work synergistically with the biofertilizing abilities of E109, which have proven to be very sensitive to As (Talano et al. 2013; Armendariz et al. 2019). Furthermore, a decrease in photosynthetic pigments has been posited as one of the injuries caused by metal(loid)-induced oxidative stress, so redox state regulation may play a vital role (Awan et al. 2020).
ROS are intermediates in several cellular cascades and play an important part in cell signaling. However, their production is exacerbated by the uptake of As, and this can cause oxidative damage to biologically relevant macromolecules (Chandrakar et al. 2017). The damage can be offset by non-enzymatic and enzymatic antioxidants (Armendariz et al. 2016; Vezza et al. 2019). Interestingly, our results suggest that the bacterial strains chosen for inoculation are able to modulate antioxidant defense in As-stressed soybean plants (Fig. 3). Co-inoculation with Cd and SF5 induced noticeable differences in the plant’s response compared to that observed with E109 alone. For example, APx and Px were up-regulated in the leaves of plants inoculated with E109 + Cd (Fig. 3a and c), while APx and SOD activity was enhanced in the roots of those inoculated with E109 + SF5 under exposure to AsV (Fig. 3b and f). Besides, the redox potential determined from the non-enzymatic components and estimated through TAA was improved in leaves and roots under treatment with AsIII and AsV, respectively, due to the application of E109 + Cd and E109 + SF5 (Fig. 3g and h). This could be the result of transcriptional and translational regulation, since PGPRs have affected the expression of genes encoding different antioxidant enzymes, as was described for tomato plants exposed to cadmium (Khanna et al. 2019a). The effects on antioxidant defense appear to be dependent on the strain, plant organ, chemical As form, and antioxidant components. The activity of catalase (CAT), SOD, and Px isoforms followed different trends when rice under stress by AsV was inoculated with B. thuringiensis and P. glucanolytus (Banerjee et al. 2020). In soybean exposed to cadmium, B. cereus induced an increase in SOD and CAT but down-regulated the activity of other enzymes such as Px (Sahile et al. 2021). Other examples of fluctuations in the antioxidant defense machinery modulated by PGPRs were described by Qadir et al. (2020) and Saleem et al. (2018) for sunflower plants treated with chromium and lead, respectively. What all the reports have in common, however, is the fact that they found a beneficial correlation between changes in the antioxidant system mediated by PGPRs, free radical scavenging, and, hence, the mitigation of oxidative stress brought about by metal(loid)s. Therefore, the inoculation of soybean with E109 + Cd and E109 + SF5 may very well contribute to creating a state of adequate antioxidant activity that would maintain cellular redox homeostasis in the presence of As.
Another key issue is the accumulation of As in plant tissues, not only due to its direct impact on metabolism but also because of the likelihood of edible parts becoming contaminated (Corguinha et al. 2015). Cd and SF5 could also be beneficial in this respect, since their application in combination with E109 noticeably impacted on the content and distribution of As in soybean plants. E109 + Cd markedly reduced the amount of As in aboveground tissues and allowed for improved rhizostabilization under AsV treatment (Fig. 4a). A significant decrease in As accumulation was also observed in roots exposed to AsIII when they were inoculated with E109 + SF5 (Fig. 4b).
The plant responses described so far may be mediated by PGPRs modulating phytohormone levels. Indole acetic acid (IAA) produced by Cd could induce As translocation changes in soybean. Accordingly, inoculation of wheat with A. brasilense Sp 245 was found to decrease As accumulation (Lyubun et al. 2006), while A. lipoferum 137 modified cadmium content in barley roots (Belimov and Dietz 2000) and A. brasilense Az39 reduced As translocation to soybean plant shoots (Armendariz et al. 2019). Similar effects were obtained with IAA-producing bacteria belonging to other genera (Wang et al. 2017; Das and Sarkar 2018), as well as with the exogenous application of IAA (Zhu et al. 2013). Several authors have suggested that IAA could improve the synthesis of phytochelatins (PCs) and, by this means, increase the capacity for As chelation in roots (Pál et al. 2018; Singh et al. 2021). PCs are known to chelate As and mediate its compartmentalization within the cell vacuole, thus immobilizing it. The mobility of As in apoplastic compartments and/or the surrounding soil and thus its entry into root cells could also be slowed down by other bacteria-assisted mechanisms, such as As bioaccumulation, precipitation, oxidation–reduction, and biosorption to surface components like biofilm (Mondal et al. 2021). Microbial ability to convert AsIII into dimethyl AsV [DMA(V)], trimethylarsine [TMA(III)], and then TMA(III) oxide (TMAO), which is volatile, could also be harnessed to decrease As bioavailability in soils (Guo et al. 2020). Events such as these might be mediated by SF5 and explain the decrease in AsIII uptake by soybean roots when inoculated with this strain (Fig. 4b). In addition, PGPR-supplemented plants are thought to feature a down-regulation of metal transporter gene expression (Khanna et al. 2019b).
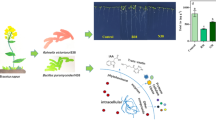
Our main findings are summarized in the diagram in Fig. 5. We believe that several mechanisms acting synergistically could be responsible for the attenuation of the negative effects by As that was observed after co-inoculating soybean with E109 + Cd and E109 + SF5. Nevertheless, the abilities of Cd and SF5 to bioremediate and respond to As are still poorly understood. The present work was designed as an exploratory, laboratory-scale study to observe the benefits of Cd and SF5 on soybean stressed by AsV and AsIII. The results should encourage further studies aimed at elucidating the underlying mechanisms and testing the strains’ potential under field and scaled-up conditions.

Representative scheme of the main effects of co-inoculation strategy using E109 + Cd and E109 + SF5 on soybean plants growing in the presence of As
Conclusions
Our findings evidence that the response of soybean to As can be modulated by bacterial inoculants, in a manner dependent on the combination of strains used and the chemical As form to which the plants are exposed. In general terms, seed co-inoculation with E109 + Cd and E109 + SF5 was advantageous compared to simple inoculation with E109. The positive impact on nodulation, photosynthetic pigment content, and the activity of the antioxidant system, as well as the significant reduction in As accumulation in plant tissues exposed to AsV and AsIII, constitute novel findings attributable to the co-inoculation strategies tested here. Therefore, new research lines are opened to study the potential use of E109 + Cd and E109 + SF5 for the formulation of inoculants to improve soybean performance under stress by As and reduce the metalloid’s entry into the food chain. A better understanding of the modes of action that restrict As uptake and mobility, among others, will be necessary to exploit the strains’ beneficial properties in polluted fields.
Data Availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Abbas G, Murtaza B, Bibi I, Shahid M, Niazi NK, Khan MI, Amjad M, Hussain M (2018) Arsenic uptake, toxicity, detoxification, and speciation in plants: physiological, biochemical, and molecular aspects. Int J Environ Res Public Health 15(1):59
Agostini E, de Forchetti SM, Tigier HA (1997) Production of peroxidases by hairy roots of Brassica napus. Plant Cell Tissue Organ Cult 47(2):177–182
Armendariz AL, Talano MA, Olmos Nicotra MF, Escudero L, Breser ML, Porporatto C, Agostini E (2019) Impact of double inoculation with Bradyrhizobium japonicum E109 and Azospirillum brasilense Az39 on soybean plants grown under arsenic stress. Plant Physiol Biochem 138:26–35
Armendariz AL, Talano MA, Travaglia C, Reinoso H, Wevar Oller AL, Agostini E (2016) Arsenic toxicity in soybean seedlings and their attenuation mechanisms. Plant Physiol Biochem 98:119–127
Awan SA, Ilyas N, Khan I, Raza MA, Rehman AU, Rizwan M, Rastogi A, Tariq R, Brestic M (2020) Bacillus siamensis reduces cadmium accumulation and improves growth and antioxidant defense system in two wheat (Triticum aestivum L.) varieties. Plants 9(7):878
Bali AS, Sidhu GPS (2021) Arsenic acquisition, toxicity and tolerance in plants – from physiology to remediation: a review. Chemosphere 283:131050
Banerjee A, Roychoudhury A (2019) Genetic engineering in plants for enhancing arsenic tolerance. In: Prasad MNV (ed) Transgenic plant technology for remediation of toxic metals and metalloids. Academic Press, pp 463–475
Banerjee A, Hazra A, Das S, Sengupta C (2020) Groundwater inhabited Bacillus and Paenibacillus strains alleviate arsenic-induced phytotoxicity of rice plant. Int J Phytoremediat 22(10):1048–1058
Beauchamp CO, Fridovich I (1973) Isozymes of SOD from wheat germ. Biochem Biophys Acta 317:50e54
Belimov AA, Dietz KJ (2000) Effect of associative bacteria on element composition of barley seedlings grown in solution culture at toxic cadmium concentrations. Microbiol Res 155(2):113–121
Ben Fekih I, Zhang C, Li YP, Zhao Y, Alwathnani HA, Saquib Q, Rensing C, Cervantes C (2018) Distribution of arsenic resistance genes in prokaryotes. Front Microbiol 9:2473
Bertani G (1951) Studies on lysogenesis I.: the mode of phage liberation by lysogenic Escherichia coli. J Bacteriol 62(3):293–300
Bojović BM, Stojanović J (2005) Chlorophyll and carotenoid content in wheat cultivars as a function of mineral nutrition. Arch Biol Sci 57(4):283–290
Brand-Williams W, Cuvelier ME, Berset C (1995) Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci Technol 28(1):25–30
Bustingorri C, Lavado RS (2014) Soybean as affected by high concentrations of arsenic and fluoride in irrigation water in controlled conditions. Agric Water Manage 144:134–139
Cassán F, Coniglio A, López G, Molina R, Nievas S, de Carlan CLN, Donadio F, Torres D, Rosas S, Olivera Pedrosa F, de Souza E, Díaz Zorita M, de Bashan L, Mora V (2020) Everything you must know about Azospirillum and its impact on agriculture and beyond. Biol Fertil Soils 56:461–479
Chandrakar V, Yadu B, Meena RK, Dubey A, Keshavkant S (2017) Arsenic-induced genotoxic responses and their amelioration by diphenylene iodonium, 24-epibrassinolide and proline in Glycine max L. Plant Physiol Biochem 112:74–86
Corguinha APB, de Souza GA, Gonçalves VC, de Andrade CC, de Lima WEA, Martins FAD, Yamanaka CH, Francisco EAB, Guilherme LRG (2015) Assessing arsenic, cadmium, and lead contents in major crops in Brazil for food safety purposes. J Food Compos Anal 37:143–150
Dardanelli MS, de Cordoba FJF, Espuny MR, Carvajal MAR, Díaz MES, Serrano AMG, Okon Y, Megías M (2008) Effect of Azospirillum brasilense coinoculated with Rhizobium on Phaseolus vulgaris flavonoids and Nod factor production under salt stress. Soil Biol Biochem 40(11):2713–2721
Das J, Sarkar P (2018) Remediation of arsenic in mung bean (Vigna radiata) with growth enhancement by unique arsenic-resistant bacterium Acinetobacter lwoffii. Sci Total Environ 624:1106–1118
Dere S, Günes T, Sivaci R (1998) Spectrophotometric determination of chlorophyll A, B and total carotenoid contents of some algae species using different solvents. Turk J Bot 22:13e18
Fatima A, Kataria S, Prajapati R, Jain M, Agrawal AK, Singh B, Kashyap Y, Tripathi DK, Singh VP, Gadre R (2020) Magnetopriming effects on arsenic stress-induced morphological and physiological variations in soybean involving synchrotron imaging. Physiol Plant 173(1):88–99
Filipini LD, Pilatti FK, Meyer E, Ventura BS, Lourenzi CR, Lovato PE (2021) Application of Azospirillum on seeds and leaves, associated with Rhizobium inoculation, increases growth and yield of common bean. Arch Microbiol 203(3):1033–1038
Forchetti G, Masciarelli O, Alemano S, Alvarez D, Abdala G (2007) Endophytic bacteria in sunflower (Helianthus annuus L.): isolation, characterization, and production of jasmonates and abscisic acid in culture medium. Appl Microbiol Biotechnol 76(5):1145–1152
Galindo FS, Teixeira M, Buzetti S, Ludkiewicz MG, Rosa PA, Tritapepe CA (2018) Technical and economic viability of co-inoculation with Azospirillum brasilense in soybean cultivars in the Cerrado. Rev Bras Engenharia Agríc Ambient 22:51–56
Gálusová T, Piršelová B, Rybanský Ľ, Krasylenko Y, Mészáros P, Blehová A, Bardáčová M, Moravčíková J, Matušíková I (2020) Plasticity of soybean stomatal responses to arsenic and cadmium at the whole plant level. Pol J Environ Stud 29:3569–3580
Goonde DB, Ayana NG (2021) Genetic diversity and character association for yield and yield related traits in soybean (Glycine max L.) genotypes. J Agri Sci Food Res 12:280
Guo J, Muhammad H, Lv X, Wei T, Ren X, Jia H, Atif S, Hua L (2020) Prospects and applications of plant growth promoting rhizobacteria to mitigate soil metal contamination: a review. Chemosphere 246:125823
Hasibuan RFM, Miyatake M, Sugiura H, Agake SI, Yokoyama T, Bellingrath-Kimura SD, Katsura K, Ohkama-Ohtsu N (2021) Application of biofertilizer containing Bacillus pumillus TUAT1 on soybean without inhibiting infection by Bradyrhizobium diazoefficiens USDA110. Soil Sci Plant Nutr 67(5):535–539
Hossain MA, Asada K (1984) Inactivation of ascorbate peroxidase in spinach chloroplasts on dark addition of hydrogen peroxide: its protection by ascorbate. Plant Cell Physiol 25:1285–1295
Irshad S, Xie Z, Mehmood S, Nawaz A, Ditta A, Mahmood Q (2021) Insights into conventional and recent technologies for arsenic bioremediation: a systematic review. Environ Sci Pollut Res 28:18870–18892
Jia X, Cao Y, O’Connor D, Zhu J, Tsang DC, Zou B, Hou D (2021) Mapping soil pollution by using drone image recognition and machine learning at an arsenic-contaminated agricultural field. Environ Pollut 270:116281
Khan I, Awan SA, Rizwan M, Ali S, Zhang X, Huang L (2021) Arsenic behavior in soil-plant system and its detoxification mechanisms in plants: a review. Environ Pollut 286:117389
Khanna K, Jamwal VL, Kohli SK, Gandhi SG, Ohri P, Bhardwaj R, Adb Allah EF, Hashem A, Ahmad P (2019a) Plant growth promoting rhizobacteria induced Cd tolerance in Lycopersicon esculentum through altered antioxidative defense expression. Chemosphere 217:463–474
Khanna K, Jamwal VL, Gandhi SG, Ohri P, Bhardwaj R (2019b) Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci Rep 9(1):1–14
Kumar S, Choudhary AK, Suyal DC, Makarana G, Goel R (2022) Leveraging arsenic resistant plant growth-promoting rhizobacteria for arsenic abatement in crops. J Hazard Mater 425:127965
Lyubun YV, Fritzsche A, Chernyshova MP, Dudel EG, Fedorov EE (2006) Arsenic transformation by Azospirillum brasilense Sp245 in association with wheat (Triticum aestivum L.) roots. Plant Soil 286(1–2):219–227
Mariño EE, Ávila GT, Bhattacharya P, Schulz CJ (2020) The occurrence of arsenic and other trace elements in groundwater of the southwestern Chaco-Pampean plain, Argentina. J South Am Earth Sci 100:102547
Molina MDC, White JF, García-Salgado S, Quijano M, González-Benítez N (2021) A gnotobiotic model to examine plant and microbiome contributions to survival under arsenic stress. Microorganisms 9(1):45
Mondal S, Pramanik K, Ghosh SK, Pal P, Mondal T, Soren T, Maiti TK (2021) Unraveling the role of plant growth-promoting rhizobacteria in the alleviation of arsenic phytotoxicity: a review. Microbiol Res 250:126809
Narożna D, Pudełko K, Króliczak J, Golińska B, Sugawara M, Mądrzak CJ, Sadowsky MJ (2015) Survival and competitiveness of Bradyrhizobium japonicum strains 20 years after introduction into field locations in Poland. Appl Environ Microbiol 81(16):5552–5559
Pajuelo E, Rodríguez-Llorente ID, Caviedes MA (2019) Effect of arsenic on legumes: analysis in the model Medicago truncatula-Ensifer interaction. In: de Bruijn F (ed) The model legume Medicago truncatula. Jonh Wiley & Sons, Inc. Pub., pp 268–280.
Pál M, Janda T, Szalai G (2018) Interactions between plant hormones and thiol-related heavy metal chelators. Plant Growth Regul 85:173–185
Qadir M, Hussain A, Hamayun M, Shah M, Iqbal A, Murad W (2020) Phytohormones producing rhizobacterium alleviates chromium toxicity in Helianthus annuus L. by reducing chromate uptake and strengthening antioxidant system. Chemosphere 258:127386
Rondina ABL, dos Santos Sanzovo AW, Guimarães GS, Wendling JR, Nogueira MA, Hungria M (2020) Changes in root morphological traits in soybean co-inoculated with Bradyrhizobium spp. and Azospirillum brasilense or treated with A. brasilense exudates. Biol Fertil Soils 56(4):537–549
Sahile AA, Khan MA, Hamayun M, Imran M, Kang SM, Lee IJ (2021) Novel Bacillus cereus strain, ALT1, enhance growth and strengthens the antioxidant system of soybean under cadmium stress. Agronomy 11(2):404
Saleem M, Asghar HN, Zahir ZA, Shahid M (2018) Impact of lead tolerant plant growth promoting rhizobacteria on growth, physiology, antioxidant activities, yield and lead content in sunflower in lead contaminated soil. Chemosphere 195:606–614
Shahzad A, Qin M, Elahie M, Naeem M, Bashir T, Yasmin H, Younas M, Areeb A, Irfan M, Billah M, Shakoor A, Zulfiqar S (2021) Bacillus pumilus induced tolerance of maize (Zea mays L.) against cadmium (Cd) stress. Sci Rep 11(1):1–11
Sharma A, Kumar V, Shahzad B, Ramakrishnan M, Sidhu GPS, Bali AS, Handa N, Kapoor D, Yadav P, Khanna K, Bakshi P, Rehman A, Kohli SK, Khan EA, Parihar RD, Yuan H, Thukral AK, Bhardwaj R, Zheng B (2020) Photosynthetic response of plants under different abiotic stresses: a review. J Plant Growth Regul 39:509–531
Sigrist M, Hilbe N, Brusa L, Campagnoli D, Beldoménico H (2016) Total arsenic in selected food samples from Argentina: estimation of their contribution to inorganic arsenic dietary intake. Food Chem 210:96–101
Singh H, Bhat JA, Singh VP, Corpas FJ, Yadav SR (2021) Auxin metabolic network regulates the plant response to metalloids stress. J Hazard Mater 405(5):124250
Singh SB, Srivastava PK (2020) Bioavailability of arsenic in agricultural soils under the influence of different soil properties. SN Appl Sci 2(2):1–16
Srivastava S, Shukla K (2019) Microbes are essential components of arsenic cycling in the environment: implications for the use of microbes in arsenic remediation. In: Arora PK (ed) Microbial Metabolism of Xenobiotic Compounds. Springer, Singapore, pp 217–227
Stefan M, Dunca S, Olteanu Z, Oprica L, Ungureanu E, Hritcu L, Mihasan M, Cojocaru D (2010) Soybean (Glycine max [L] Merr.) inoculation with Bacillus pumilus Rs3 promotes plant growth and increases seed protein yield: relevance for environmentally-friendly agricultural applications. Carpath J Earth Environ Sci 5(1):131–138
Talano MA, Cejas RB, Gonzalez PS, Agostini E (2013) Effect of sodium arsenate and arsenite on soybean development and in the symbiotic interaction with Bradyrhizobium japonicum. Plant Physiol Biochem 63:8e14
Upadhyay MK, Shukla A, Yadav P, Srivastava S (2019) A review of arsenic in crops, vegetables, animals and food products. Food Chem 276:608–618
Vezza ME, Llanes A, Travaglia C, Agostini E, Talano MA (2018) Arsenic stress effects on root water absorption in soybean plants: physiological and morphological aspects. Plant Physiol Biochem 123:8–17
Vezza ME, Luna DF, Agostini E, Talano MA (2019) Glutathione, a key compound for As accumulation and tolerance in soybean plants treated with AsV and AsIII. Environ Exp Bot 162:272–282
Vezza ME, Nicotra MFO, Agostini E, Talano MA (2020) Biochemical and molecular characterization of arsenic response from Azospirillum brasilense Cd, a bacterial strain used as plant inoculant. Environ Sci Pollut Res 27(2):2287–2300
Vezza ME, Alemano S, Agostini E, Talano MA (2021) Arsenic toxicity in soybean plants: impact on chlorophyll fluorescence, mineral nutrition and phytohormones. J Plant Growth Regul. https://doi.org/10.1007/s00344-021-10469-1
Vincent JM (1970) A manual for the practical study of the root-nodule bacteria. Blackwell Scientific Publications, Oxford
Wang X, Nie Z, He L, Wang Q, Sheng X (2017) Isolation of As-tolerant bacteria and their potentials of reducing As and Cd accumulation of edible tissues of vegetables in metal (loid)-contaminated soils. Sci Total Environ 579:179–189
Zhu XF, Wang ZW, Dong F, Lei GJ, Shi YZ, Li GX, Zheng SJ (2013) Exogenous auxin alleviates cadmium toxicity in Arabidopsis thaliana by stimulating synthesis of hemicellulose 1 and increasing the cadmium fixation capacity of root cell walls. J Hazard Mater 263:398–403
Zulfiqar F, Ashraf M (2022) Antioxidants as modulators of arsenic-induced oxidative stress tolerance in plants: an overview. J Hazard Mater 427:127891
Acknowledgements
The authors wish to express their gratitude to Dr. Sergio Alemano (Departamento de Ciencias Naturales, FCEFQyN, Universidad Nacional de Río Cuarto, INIAB-CONICET) for providing the Bacillus pumilus SF5 strain.
Funding
This research was financially supported by Fondo para la Investigación Científica y Tecnológica (FONCYT) (grant PICT 2143/17) and by Secretaría de Ciencia y Técnica-Universidad Nacional de Río Cuarto (SeCyT-UNRC).
MEV holds a post-doctoral grant from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). RPP holds a doctoral grant from FONCyT. ALWO, EA, and MAT are members of the Research Career of CONICET.
Author information
Authors and Affiliations
Contributions
MEV, EA, and MAT: research design. MEV: methodology, investigation, writing – original draft. RPP: data collection. ALWO: writing – review and editing. EA and MAT: funding acquisition, supervision, writing – review and editing.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Gangrong Shi
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Vezza, M.E., Pramparo, R.d.P., Wevar Oller, A.L. et al. Promising co-inoculation strategies to reduce arsenic toxicity in soybean. Environ Sci Pollut Res 29, 88066–88077 (2022). https://doi.org/10.1007/s11356-022-21443-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-022-21443-z