
Abstract
Integrated aerobic granules were first cultivated in two sequencing batch reactors (SBRs) (A1 and A2). Then, A1’s influent organic loading rate (OLR) was changed from alternating to constant (cycling time was still 6 h), while A2’s cycling time varied from 6 to 4 h (influent OLR strategy remained alternating). After 30-day operation since the manipulative alternations, granule breakage happened in two reactors at different operational stages, along with the decrease of granule intensity. Granule diameter in A1 declined from the original 0.84 to 0.32 cm during the whole operation, while granules in A2 dwindled to 0.31 cm on day 22 with similar size to A1. Both the amount of total extracellular polymeric substances (EPSs) and the protein were declining throughout the operation, and the large molecular weight of protein was considered closely related to the stability of aerobic granules. The relative AI-2 level decreased at the same time, and influent OLR strategy might had more evident impact on quorum sensing (QS) ability of sludge compared with starvation period. Combined with microbial results, the decline of total EPS amount in two reactors could be concluded as follows: During the reactor operation, some functional bacteria gradually lost their dominance and were eliminated from the reactors, which finally caused granule disintegration. In summary, the results further confirmed that alternating OLR and proper starvation period were two major factors in effective cultivation and stability of aerobic granules from the perspective of QS.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Aerobic granulation technology has attracted world attention for its various advantages, including excellent settleability, high biomass concentration, and strong resistance to organic loading rate (OLR) (Adav et al. 2008). However, the mechanism of aerobic granulation still remains unclear, and the loss of granule stability is one of the major obstacles against its practical application in wastewater treatment. To achieve granule stability, operational conditions should be coordinated to optimization. Both influent OLR strategy and starvation period play an essential part in granule stability, for they are closely related with cell adhesiveness, extracellular polymeric substance (EPS) secretion, and microbial development (Lee et al. 2010). Alternating OLR and proper length of starvation period could accelerate aerobic granulation by promoting cell adhesiveness and total EPS amount, as well as the level of large molecular weight (MW) EPS (Liu et al. 2016; Sun et al. 2016).
Quorum sensing (QS) existed generally in microbial world where most microbes could secrete autoinducers (AIs) (Bandara et al. 2012). AI concentration was positively related with population density. Once the level of AI reaches to a certain threshold, the receptor protein would be activated and trigger the expression of specific genes such as bioluminescence, mobility, virulence, pathogenicity, and so on (Lesprit et al. 2003). Gram-negative and gram-positive bacteria communicate with each other with autoinducer-2 (AI-2), which is an analogy of “universal language” between microbes. Researchers have discovered that aerobic granulation process was strongly related to QS system. Liu et al. (2016) cultivated aerobic granules using different starvation period and detected a rising tendency of AI-2 in all reactors. Xiong and Liu (2010) also observed that AI-2 level ascended evidently after aerobic granules reached maturity. On the other hand, the drop of AI-2 concentration and granule breakage happened soon after AI-2 synthesis was inhibited (Jiang and Liu 2013).
This paper was aimed to investigate granule stability during the variation of influent OLR strategy and starvation period from the perspective of granule stableness maintenance. Chemical oxygen demand (COD) removal ability, granule intensity, relative AI-2 content, EPS characteristics, and microbial communities were combined together in the perspective of quorum sensing. Moreover, a theoretical explanation was proposed for granule destabilization caused by the variation of operational conditions.
Materials and methods
Reactor setup
Two sequencing batch reactors (SBRs) (A1 and A2) were set up using alternating influent OLR to cultivate mature aerobic granules. The operation cycle was 6 h which consisted of 4 min feeding, 15 min settling, 335 min aeration, and 6 min effluent discharge, giving a volumetric exchange ratio of 60%. The influent COD content switched among three levels which are 1250, 2500, and 5000 mg/L, while the change of influent COD content was synchronous in A1 and A2. During the cultivation period, the SBR cycles of two reactors remained at 6 h until mature aerobic granules took shape. After mature granules were successfully cultivated in two reactors, the influent COD of A1 remained constant at 2500 mg/L while cycling time was still 6 h, meaning that the starvation period remained at a normal length. When A2’s influent COD was kept alternating between 2500 and 5000 mg/L, its cycling time was shortened to 4 h which meant a shortened starvation period. Both reactors were operated for 30 days, and the sludge samples were collected synchronously, which were also at the end of one SBR cycle. Both A1 and A2 could reduce influent COD content, whatever the content was, to less than 100 mg/L in an hour. Therefore, the starvation period was defined as an hour shorter than the cycling period in this study (Liu et al. 2016).
The inoculated sludge was from a wastewater treatment plant in Shanghai with the content of 7.0–8.0 g/L. The influent was synthetic wastewater containing sodium acetate and sodium propionate as carbon source. The COD ratio of sodium acetate to sodium propionate was 3:1. Other nutrient elements included nitrogen, phosphate, calcium, magnesium, etc. Table S1 gives the details of influent composition. The influent pH value remained at 6.8 ± 0.2. The basic parameters are listed in Table S2.
Granule intensity
Ultrasonication was adopted to measure granule intensity in this study (Wan et al. 2013). Several aerobic granules were chosen randomly and washed gently in advance. Then, they were placed in a beaker (20 mL), and 15 mL deionized water was added. The optical density at the wavelength of 600 nm (OD600) value of the supernatant was measured using a spectrophotometer (DR5000, HACH). After that, the supernatant was poured back into the beaker and the ultrasonication started. The supersonic apparatus (JY92-IID Broken Series, Xinzhi) was set at 60 W, while the process was 3 s on and 2 s off. The OD600 supernatant was measured every 5 s, recorded as OD600′. This step was repeated until the OD600′ value was constant. The final OD600′ was recorded as OD600 0. The granule breakage curve was worked out using OD600′/OD600 0 as the ordinate scale and ultrasonication time as horizontal axis. The granule intensity was negatively correlated with the curve slope.
EPS extraction and measurement
All EPS samples were extracted using heat method (Lv et al. 2014). About 15 mL crushed aerobic granules were placed in a 50-mL centrifugal tube and centrifuged at 5000 rpm for 5 min. Then, the supernatant was discarded, and deionized water was added till the initial volume. The mixture was homogenized and bathed in hot water (80 °C) for 30 min. After that, the samples were centrifuged at 8000 rpm for 10 min and filtered through 0.22-μm membrane. The filtrate was the extracted EPS and kept at −20 °C.
The measurement of protein (PN) content was conducted according to the modified Lowry method (Lowry et al. 1951). Polysaccharide (PS) concentration was measured by phenol-sulfuric acid method (Herbert et al. 1971).
The MW distribution of EPS measurement
The MW distribution was measured using HPLC (Agilent 1260, USA) with SEC chromatographic column (PL1149-6801). The mobile phase was the mixture of 0.001 M NaH2PO4, 0.001 M Na2HPO4, and 0.03 M NaCl while the flow speed was 0.5 mL/min. The EPS samples were detected using DAD at the wavelength of 254 nm. The correlation between MW and retention time was established in advance. Therefore, the MW distribution of EPS could be gained (Yan et al. 2012).
To ensure whether the individual EPS peak belongs to PN or PS, protease K solution (20 mg/mL) was added into EPS samples. Then, the samples were bathed in hot water at 60 °C for 2 h, and PN content was measured. After PN was removed completely, the MW distribution of EPS was measured again. The protease K was purchased from Sangon Biotech (Shanghai) Co., Ltd.
The extraction and detection of AI-2 molecules
The extraction of AI-2 molecules was performed according to Xiong and Liu (2012) with some modifications. Before extracting AI-2, 4–5 granules were placed in a beaker and eluted several times. Then, the mixture was sonicated in an ultrasonic apparatus (JY92-IID Broken Series, Xinzhi). After the OD600 value of the mixture got to a constant value, 10 mL sludge was bathed in ice for 5 min and centrifuged at 10000 rpm. Finally, the supernatant was filtered through 0.22-μm membrane.
The relative AI-2 content was measured using Vibrio harveyi BB170 (ATCC BAA-1117), which would give out fluorescence (Xiong and Liu 2012) in the presence of AI-2. The AI-2 concentration was positively related with fluorescence intensity of V. harveyi BB170, with V. harveyi BB120 as positive control (Han and Lu 2009). These two reporter strains were first cultured in autoinducer bioassay (AB) medium at 30 °C with a shaker (Blue Pard HZQ-X300C, China). The cultivation lasted for 14–16 h until the liquid became turbid (OD600 = 0.7–1.2). Next, the bacterial suspension was diluted in fresh AB medium, mixed with the extracted AI-2 at 9:1 ratio, and added into a 96-well plate. Then, the 96-well plate was shaken with a lid at 150 rpm at 30 °C for 6 h. Finally, the bioluminescence intensity from each well was measured at 490 nm wavelength by a microplate reader (BioTek synergy H1, USA). The obtained optical density at the wavelength of 490 nm (OD490) divided by volatile suspended solid (VSS) was calculated to characterize the relative AI-2 content of different samples. Each sample was analyzed in triplicate. The composition of AB medium AB is listed in Table S3, while KOH was added to regulate pH at 7.5. AB medium was filtered through 0.22-μm membrane before use.
Microbial analysis
Before microbial analysis, aerobic granules were first crushed and dewatered. The UltraClean™ Soil DNA Isolation Kit was used to extract DNA. The polymerase chain reaction (PCR) amplification was conducted according to Sun et al. (2016). The PCR primers were synthesized by Takara Co., Ltd. The PCR products were detected by electrophoresis in 1% agarose gels, and the positive PCR samples were performed with denaturing gradient gel electrophoresis (DGGE). After the targeted bands were excised and recycled, the purified amplicons were linked to pMD-18T and inoculated into Escherichia coli DH5α cells. Then, three positive clones were picked randomly of each sample and sent to Sangon Co., Ltd. (Shanghai, China) for sequencing. Finally, the obtained sequences were compared on GenBank to identify their closest genes.
Other analytical methods
Mixed liquor suspended solids (MLSS), VSS, and COD concentration were quantified according to APHA (1998).
Results
Granule morphology
Mature granules of 0.7–0.9 cm in diameter with regular shape and yellowish color were first cultivated in two identical SBRs (A1 and A2). After the alteration of operational conditions, sludge in A1 and A2 was photographed every day to record its morphological changes. The mean granule diameter was calculated using ImageJ (version 1.45). Since the OLR of A1 was changed from alternative to constant, the original aerobic granules with integral round shape began to lose stability on the 11th day. Granules with large size almost disappeared on the 18th day and only few integral aerobic granules were left on the 30th day (Fig. S1). Combining Fig. S1 and Fig. 1, the mean diameter of granules in A1 descended obviously, reaching 0.84 cm on the 5th day. Then, the granule size dropped steadily to 0.32 cm on 30th day with weakened intensity and crushed structure.

Granule diameter variation of A1 and A2
Granule breakage in A2 was first observed on the 15th day since the cycling time was shortened from 6 to 4 h. According to Fig. S2, integral granules barely existed on the 22nd day. The granule diameter topped 0.74 cm on the 9th day. After that, the granule size also shrank, especially between the 13th day and the 18th day. The destabilization speed of granules in A2 accelerated since the 20th day, while the granule diameter fell drastically to 0.31 cm until the 22nd day. The results demonstrated that mature aerobic granules in both SBRs suffered from destabilization after long-term operation with alternating OLR or shortened starvation period.
Granule intensity
Figure 2 shows the variation of granule intensity in two reactors, both of which declined step by step. The initial granules did not break until the 100th second, while granules on the 12th day remained integrity for 30 s during ultrasonication. In the latter operational period of A1, the value of OD600′/OD600 0 reached 100% in only 20 s, which implied that granule structure broke up in less time compared with initial granules. Similarly, the initial granules of A2 disintegrated in 180 and 150 s on the 12th day. It took 30 s for granules to break completely on the 18th day. Although the granule intensity of A2 on d1-d12 was close, it performed better on d12 than on d1, which could be attributed to the sudden increase of A2’s OLR from 1250 to 2500 mg/L. Elevated OLR might be beneficial to granule stability which implied stronger granule strength (Sun et al. 2016). The results indicated that long-term constant OLR and short starvation period would harm granule stability.

Granule intensity variation of a A1 and b A2
COD degradation ability
Judging from Fig. 3, COD removal efficiencies of A1 and A2 were both excellent. When the influent OLR was kept constant at 2500 mg/L, the COD removal rate remained over 95% in A1 during the whole operation. Different from A1, the influent OLR of A2 alternated between 2500 and 5000 mg/L. The COD removal ability of A2 was no weaker than A1, over 90% on the whole. However, the COD removal rate of both reactors decreased a little along with granule destabilization. It turned out that the operational conditions variation would have some negative influence on COD removal ability of granules, while this kind of influence was slender during the reactor operation.

COD removal efficiencies of a A1 and b A2
EPS content of granules
Figure 4 shows EPS content variation after the operational conditions changed in A1 and A2. During the loss of granule stability, the protein content was predominant in the EPS, much higher than PS. The protein content of A1 dropped from 22.33 mg/g MLSS on the 1st day to 13.98 mg/g MLSS on the 12th day. Then, the decreasing trend accelerated, reaching 5.02 mg/g MLSS on the 18th day. The protein content fell to 3.62 mg/g until the end of A1 operation. In comparison, the trend of PS content in A1 declined at first and increased afterwards, which fell to 1.42 mg/g MLSS on the 12th day and climbed to 1.93 mg/g MLSS on the 18th day. The fluctuation was mild during the whole operation.

Extracellular protein (PN) and polysaccharides (PS) content variation of granules in a A1 and b A2
The protein content in A2 climbed a little after the cycling time was shortened to 4 h. However, the protein level started to decline since the 4th day, from 19.53 to 8.11 mg/g MLSS on the 12th day. In the late period of A2, the protein content remained at 6.0 mg/g MLSS. On the contrary, PS content in A2 remained at a low level, fluctuating between 2.77 and 3.67 mg/g MLSS in the first 20 days. Since then, the value slightly descended and touched 1.32 mg/g MLSS on the last day of operation. Comparatively, the protein value of A1 with alternating OLR dropped more severely than that of A2, which had shorter starvation period.
MW distribution of EPS
The MW distribution of EPS during A1 and A2’s operation experienced distinct variation as well. According to Fig. 5a, the MW of dominant EPS in A1 was below 10,000 Da, among which the proportion of EPS with medium MW (1000–10,000 Da) remained over 70% during the whole operation. The proportion of EPS with small MW (<1000 Da) ascended steadily since the middle stage of the operation, reaching 26.62% on the 26th day from 11.03% on the 12th day. During the 4th day to the 18th day, large MW EPS (10,000–1,000,000 Da) appeared in A1, while EPSs with extremely large MW (>1,000,000 Da) were only detected in the middle period. Since the 24th day, the MW of EPS in A1 was mostly below 10,000 Da. Therefore, it was inferred that constant influent OLR was not suitable for secretion of large MW EPS, which had positive impact on cell adhesion, total EPS amount, and granulation process (Liu et al. 2016).

Molecular weight (MW) distribution of extracellular polymeric substances (EPS) of granules in a A1 and b A2
On the other hand, the proportion of extremely large MW EPS in A2 decreased at first and rose afterwards. This part of EPS occupied over 20% in the initial operation stage, falling to less than 5% on the 12th day. Then, the EPS proportion climbed to 43.29% till the 30th day. The trend of EPS with medium MW declined slowly since the 12th day, from 85.02 to 16.65% on the 30th day. In addition, the proportion of small MW EPS increased during the late stage of A2 operation, which implied that this part of EPS might ascend with granule breakage process. The results showed that the proportion of EPS with medium MW declined along with granule breakage, which might be closely related with granule stability. On the other hand, the level of EPS with large MW remained steady which might be relevant with the alternating influent OLR of A2 (Sun et al. 2016). Besides, granules might not have enough time to consume large MW EPS under short starvation period, which might also contribute to their accumulation.
Figure 6 compares the MW distribution of EPS before and after PN was removed from EPS samples, which could determine whether individual peak represented for PN or not. Judging from Fig. 6a–d, respectively, the removal of PN caused the peaks to dwindle or even disappear before 10 min, between 15 and 22 min and after 25 min. The peaks existing before 10 min as well as after 25 min dwindled obviously, while some even vanished. These peaks stood for EPS with MW over 4,000,000 Da and below 100 Da, which demonstrated that the MW of PN produced by the sludge in this experiment was mainly within the range. In addition, the peaks existing between 15 and 22 min were shortened, which represented for removed PN with MW between 600 and 100,000 Da.

EPS MW distribution of A1 and A2. a Day 18 EPS before PN removal of A1. b Day 18 EPS after PN removal of A1. c Day 30 EPS before PN removal of A2. d Day 30 EPS after PN removal of A2
QS ability of granules
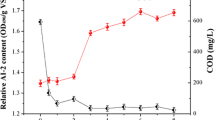
Figure 7 displays the relative content variation of AI-2 in two reactors, both of which declined evidently along with granule disintegration. In the first 9 days of A1 operation, AI-2 level did not change much. The influent OLR of A1 was just altered to constant, so the influence of OLR on granule QS ability had not been revealed. However, since the 9th day the AI-2 level of A1 fell drastically from 2.21 OD490/g VSS to 1.34 OD490/g VSS and stabilized at 1.3 OD490/g VSS in the end.

Quorum sensing ability of granules in A1 and A2
After A2’s starvation period was shortened, the relative AI-2 content first climbed to 2.18 OD490/g VSS on the 4th day. Then, the value descended steadily to 1.63 OD490/g VSS on the 18th day and finally stopped at around 1.60 OD490/g VSS. The results suggested that AIs might be strongly related with granule stability. Therefore, it was speculated that the ability of microbes to produce AIs would be imperiled by the variation of operational conditions, leading to granule destabilization even disintegration at last.
Microbial communities
Twenty representative bands were detected in the DGGE pattern of two reactors (Fig. S3), and sequence analysis results are listed in Table 1. Some bacteria remained high abundance throughout the operation, such as bands 2, 5, 9, 10, and 11. Band 2 belonged to Acinetobacter, which had the ability to self-aggregate and coaggregate. This kind of microbes would secrete a great deal of EPS to enhance cell adhesion and hydrophobicity of epicyte, which could strengthen granule structure (Adav et al. 2010). Some bacteria only existed in the early and middle stage of the operation like bands 3, 4, 8, 12, and 15, whose abundance dwindled along with the change of operational conditions. Bands 3 and 4 belonged to Xanthomonadaceae, which was reported to produce plenty of flavochrome (Wan et al. 2014). Flavochrome was favorable to link and flocculation between cells (Ashassi-Sorkhabi et al. 2012; Wan et al. 2013), remaining structure stability during granule cultivation. Other bands were not present in the initial sludge, but their abundance increased steadily such as bands 19 and 20. Band 19 stood for Zoogloea ramigera strain, which was an essential obligate aerobic and chemoheterotrophic bacteria in wastewater treatment. Band 20 was Thiobacillus denitrificans strain, specializing in denitrification and desulfidation.
Bands 5, 10, 11, and 20 were predominant in A2’s DGGE pattern during the whole operation, which represented for Rhizobium sp., Aquimonas sp., Sphingomonas sp., and T. denitrificans strain. Sphingomonas sp. was excellent at degrading refractory organics like mononuclear aromatics, polycyclic aromatic hydrocarbons, and biphenyls, which might be responsible for COD removal efficiency in the experiment (Shan et al. 2015). Several bacteria first existed in the initial granules but gradually disappeared from the reactor like bands 8, 12, 16, and 18. Bands 16 and 18 belonged to Rhodocyclaceae, which are able to produce huge amounts of PS and consolidate attachment between cells (Besaury et al. 2014). The declining trend of these bacteria might account for granule destabilization of A2 to some extent.
The microbial analysis revealed that the change of operational conditions would influence microbial structure. After granule disintegration, microbial diversity decreased, while some functional bacteria were eliminated from the system along with the variation of operational conditions. The impact of constant OLR and shortened starvation period on granule properties was reflected through cell adhesiveness, EPS composition, and content.
Discussion
Granule stability has been reported relevant with many granule properties, including granule intensity, cell adhesion, hydrophobicity, EPS composition, and MW distribution in this study. Since the influent OLR of A1 was changed from alternating to constant, the aerobic granules began to lose stability on the 11th day and cracked mostly during the late stage of operation. Similar to A1, destabilization happened on granules in A2 since the 15th day, which aggravated rapidly until the 23rd day. Moreover, the granule diameter and intensity of two reactors declined together with the variation of operational conditions (Figs. 1 and 2). Combining EPS results from Fig. 4a, the total amount of EPS decreased as soon as granule destabilization happened. The drop of PN content was especially evident, which fell to less than 25% of the original concentration level. It has been reported that reduced EPS secretion is one of the main reasons for granule breakage (Luo et al. 2014), because PN is the hydrophobic component of EPS and it plays an important part in granule stability (Li et al. 2014a). Therefore, the decrease of EPS and PN content in the experiments might lead to the disintegration of aerobic granules.
In addition, it was found out in previous researches that alternating OLR as well as prolonged starvation period was beneficial to the production of large MW EPS (Liu et al. 2016; Sun et al. 2016). Judging from the MW distribution comparison results of EPS before and after PN removal, the majority of large and extremely large MW EPS were PN (Fig. 6). The disappearance of large MW EPS suggested that neither constant OLR nor shorten starvation period would trigger the secretion of the large MW PN. On the contrary, according to Fig. 4, the EPS accumulated during original granulation process might be consumed by the microorganisms gradually (Liu et al. 2016). Therefore, the reduction of PN induced by these two operational conditions further indicated that PN was one of the critical factors in granule disintegration. It was worth noting that although granules of A2 almost lost stability in the late stage of operation, the proportion of EPS with large MW still ascended, which might be due to the descending proportion of large and middle MW EPS. Furthermore, as more microbes were eliminated during the experiment, the microbial diversity of A1 and A2 declined at the same time. With the disappearance of some functional bacteria (for example, bands 3 and 4), the amount of EPS was secreted less which might finally lead to granule collapse.
Quorum sensing has been testified to have an essential role in aerobic granules. Bandara et al. (2012) reported that AI-2 could regulate various kinds of microbial physiological properties such as EPS secretion, virulence expression, and flagellum vitality, which were closely related with granule stability. Li and Zhu (2014) found that N-acyl homoserine lactone (AHL) could regulate aerobic granulation by controlling EPS content and composition. During the very early stage of the experiment, AI-2 content of granules in both reactors remained at a high level along with the relative high content of EPS. According to Fig. S1 and Fig. 7, granule structure began to collapse since the 11th day while the EPS content and the relative AI-2 content fell markedly at the same time, indicating the close relationship between the EPS and the AI-2 content. Similarly, Tan et al. (2014) reported that the specific AHL level decreased sharply during the loss of granule stability. Therefore, it can be inferred that the declining AI-2 concentration might be caused by the altered operational conditions, and then the AI-2 content affect granule stability and lead to the disintegration directly by regulating the EPS secretion.
In conclusion, constant influent OLR and shortened starvation period were both unfavorable to AI-2 secretion and could cause granule breakage in the end. On one hand, since alternating influent OLR would promote AI-2 production, microbes could be motivated to secrete large MW EPS for enhancing cell adhesiveness after AI-2 content was above a certain threshold. When influent OLR no longer varied, the stimulants from outside disappeared simultaneously along with the decreased AI-2 content. Thus, the sticky EPS with large MW regulated by the QS system would be less produced by granules, which could trigger disintegration. On the other hand, microbes would release more AIs to protect themselves from starvation period (Li et al. 2014b), resulting in higher cell adhesiveness and more EPS production. For this reason, microbes would be less willing to secrete AI-2 and EPS after starvation period was shortened. With lower cell adhesiveness and less EPS with large MW, granules in A2 lost stability gradually and disintegrated in the end.
Conclusions
This paper studied the influence of altered operational conditions on aerobic granules stability. Granule breakage happened soon after the variation of influent OLR (A1) or starvation period (A2). The total amount of EPS and PN in both reactors dropped synchronously. Constant OLR could inhibit the secretion of EPS with large MW, while the MW of granule EPS under shortened starvation period declined as well. Therefore, the descent of EPS total amount and MW might account for granule destabilization. The AI-2 level of two reactors showed an obvious decreasing trend, which suggested that constant OLR and shortened starvation period might be unfavorable to granule stability. During granule destabilization process, microbial diversity decreased obviously with some functional bacteria (including Acinetobacter and Thauera) being eliminated from the systems gradually.
References
Adav SS, Lee DJ, Show KY, Tay JH (2008) Aerobic granular sludge: recent advances. Biotechnol Advan 26:411–423
Adav SS, Lee DJ, Lai JY (2010) Potential cause of aerobic granular sludge breakdown at high organic loading rates. Appl Microbiol and Biotechnol 85:1601–1610
APHA (1998) Standard methods for the examination of water and wastewater, 20th edn. American Public Health Association, Washington DC
Ashassi-Sorkhabi H, Moradi-Haghighi M, Zarrini G (2012) The effect of Pseudoxanthomonas sp. as manganese oxidizing bacterium on the corrosion behavior of carbon steel. Mater Sci and Eng C 32:303–309
Bandara H, Lam O, Jin L, Samaranayake L (2012) Microbial chemical signaling: a current perspective. Crit Rev in Microbiology 38:217–249
Besaury L, Pawlak B, Quillet L (2014) Expression of copper-resistance genes in microbial communities under copper stress and oxic/anoxic conditions. Environ Sci Pollut Res 23(5):4013–4023
Han X, Lu C (2009) Detection of autoinducer-2 and analysis of the profile of luxS and pfs transcription in Streptococcus suis serotype 2. Curr Microbiol 58:146–152
Herbert D, Philipps P, Strange R (1971) Carbohydrate analysis. Meth Enzymol B 5:265–277
Jiang B, Liu Y (2013) Dependence of structure stability and integrity of aerobic granules on ATP and cell communication. Appl Microbiol and Biotechnol 97:5105–5112
Lee DJ, Chen YY, Show KY, Whiteley CG, Tay JH (2010) Advances in aerobic granule formation and granule stability in the course of storage and reactor operation. Biotechnol Advan 28:919–934
Lesprit P, Faurisson F, Join-Lambert O, Roudot-Thoraval F, Foglino M, Vissuzaine C, Carbon C (2003) Role of the quorum-sensing system in experimental pneumonia due to Pseudomonas aeruginosa in rats. Amer J of Respir Crit Care Med 167:1478–1482
Li YC, Zhu JR (2014) Role of N-acyl homoserine lactone (AHL)-based quorum sensing (QS) in aerobic sludge granulation. Appl Microbiol Biotechnol 98:7623–7632
Li Y, Hao W, Lv J, Wang Y, Zhong C, Zhu J (2014a) The role of N-acyl homoserine lactones in maintaining the stability of aerobic granules. Bioresource Technol 159:305–310
Li Y, Lv J, Zhong C, Hao W, Wang Y, Zhu J (2014b) Performance and role of N-acyl-homoserine lactone (AHL)-based quorum sensing (QS) in aerobic granules. J Environ Sci 26:1615–1621
Liu X, Sun S, Ma B, Zhang C, Wan C, Lee DJ (2016) Understanding of aerobic granulation enhanced by starvation in the perspective of quorum sensing. Appl Microbiol Biotechnol 100:3747–3755
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193:265–275
Luo J, Hao T, Wei L, Mackey HR, Lin Z, Chen GH (2014) Impact of influent COD/N ratio on disintegration of aerobic granular sludge. Water Res 62:127–135
Lv J, Wang Y, Zhong C, Li Y, Hao W, Zhu J (2014) The effect of quorum sensing and extracellular proteins on the microbial attachment of aerobic granular activated sludge. Bioresource Technol 152:53–58
Shan LL, Yu YL, Zhu ZB, Zhao W, Wang HM, Ambuchi JJ, Yujie F (2015) Microbial community analysis in a combined anaerobic and aerobic digestion system for treatment of cellulosic ethanol production wastewater. Environ Sci Pollut Res 22:17789–17798
Sun S, Liu X, Ma B, Wan C, Lee DJ (2016) The role of autoinducer-2 in aerobic granulation using alternating feed loadings strategy. Bioresource Technol 201:58–64
Tan CH, Koh KS, Xie C, Tay M, Zhou Y, Williams R, Ng WJ, Rice SA, Kjelleberg S (2014) The role of quorum sensing signalling in EPS production and the assembly of a sludge community into aerobic granules. The ISME J 8:1186–1197
Wan C, Sun S, Lee DJ, Liu X, Wang L, Yang X, Pan X (2013) Partial nitrification using aerobic granules in continuous-flow reactor: rapid startup. Bioresource Technol 142:517–522
Wan C, Yang X, Lee DJ, Sun S, Liu X, Zhang P (2014) Influence of hydraulic retention time on partial nitrification of continuous-flow aerobic granular-sludge reactor. Environ Technol 35:1760–1765
Xiong Y, Liu Y (2010) Involvement of ATP and autoinducer-2 in aerobic granulation. Biotechnol Bioeng 105:51–58
Xiong Y, Liu Y (2012) Essential roles of eDNA and AI-2 in aerobic granulation in sequencing batch reactors operated at different settling times. Appl Microbiol Biotechnol 93:2645–2651
Yan M, Korshin G, Wang D, Cai Z (2012) Characterization of dissolved organic matter using high-performance liquid chromatography (HPLC)-size exclusion chromatography (SEC) with a multiple wavelength absorbance detector. Chemosphere 87:879–885
Acknowledgements
This work is financially supported by the National Natural Science Foundation of China (Nos. 51278128 and 41301332), Shanghai International Science and Technology Cooperation Fund (No. 14230700400), Outstanding Talent Plan of Fudan University (IBH1829001), and Key Laboratory of Reservoir Aquatic Environment, Chinese Academy of Science (No. RAE2014CB05B).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Responsible editor: Diane Purchase
Electronic supplementary material
ESM 1
(DOCX 935 kb)
Rights and permissions
About this article

Cite this article
Zhang, C., Sun, S., Liu, X. et al. Influence of operational conditions on the stability of aerobic granules from the perspective of quorum sensing. Environ Sci Pollut Res 24, 7640–7649 (2017). https://doi.org/10.1007/s11356-017-8417-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-017-8417-7