
Abstract
Satoyama is one of the semi-natural ecosystems in Japan. Traditional agricultural practices in satoyama provide preferable habitats for many species, thus, enhancing biodiversity. However, many satoyama ecosystems have been altered by agricultural intensification and rapid urbanization. Meanwhile, ageing and shrinking population has led to the abandonment of remaining satoyama ecosystems, resulting in their potential degradation. We argue that counteracting the abandonment of satoyama is particularly essential in urbanized landscapes because conserving its biodiversity would have a payoff for urban communities. We focused on nature-oriented park use of satoyama ecosystems, utilizing their original scenery, topography, water features, and vegetation. To evaluate the potential of nature-oriented park use for biodiversity conservation, we examined the differences in diversity and species composition of vascular plants and butterflies among three land-use types (agricultural-use, park-use, and abandoned sites) of satoyama ecosystems. Diversity of native plants and butterflies did not significantly differ between agricultural-use and park-use sites and that of native plants was significantly lower at abandoned sites than at agricultural-use sites. Although species composition of native plants at agricultural-use sites and park-use sites significantly differ, that of native butterflies did not. The efficacy of park use for biodiversity conservation, thus, depended on the taxa. Nonetheless, for striking a balance between the difficulty in maintaining agricultural use and accelerating satoyama abandonment, our study highlights the utility of park use as an alternative usage, contributing to biodiversity conservation. Balancing the wise use of satoyama in urbanized landscapes is a fundamental step toward the conservation of its biodiversity and human–nature interactions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Human activities have adversely affected biodiversity in many ecosystems (Foley et al. 2005; Grimm et al. 2008; Sala 2000). In semi-natural ecosystems, such as agricultural fields and pastures, however, many studies have suggested that the effects of human activities on biodiversity are often positive rather than negative (Queiroz et al. 2014; Uchida and Ushimaru 2014; Bonari et al. 2017; Uchida et al. 2018b). In Japan, satoyama is broadly defined as a mosaic of different habitats including secondary woodlands and grasslands, paddy fields, and crop fields (Fukamachi et al. 2001; Koyanagi et al. 2012; Ministry of the Environment 2010; Takeuchi et al. 2003). Historically, traditional agroforestry practices in satoyama, such as mowing and tree thinning, has been implemented to obtain fodder and fuels (Berglund et al. 2014; Takeuchi et al. 2003). These practices in satoyama provide suitable habitats for many species across taxa; periodic and intermediate-level disturbances create environmental heterogeneity and mosaics, which in turn enhance biodiversity (Katoh et al. 2009; Koyanagi et al. 2009; Uchida and Ushimaru 2014; Ohwaki 2015).
Over the past 100 years, however, many satoyama ecosystems have been altered due to agricultural intensification (Uematsu et al. 2010) and rapid urbanization (Koyanagi et al. 2012; Soga and Koike 2013; Uchida et al. 2018a). Meanwhile, the ageing and shrinking population has led to the abandonment of remaining satoyama (Fig. 1). Given that urban shrinkage and population decline are expected to continue in Japan during the next few decades, the emerging issue for satoyama would be to figure out a way to counteract the abandonment of remaining satoyama (Normile 2016). Uchida and Ushimaru (2014) demonstrated that abandonment of formerly managed satoyama ecosystems decreases the species richness of plants, butterflies, and grasshoppers. They suggested that a low level or absence of anthropogenic disturbance following land abandonment causes a decline in habitat quality, resulting in biodiversity loss. Here, we argue that counteracting the abandonment of satoyama is particularly essential in urbanized landscapes because conserving the biodiversity would have a payoff to the urban communities (Dallimer et al. 2012; Aoshima et al. 2018). Biodiversity in satoyama ecosystems potentially provides a range of opportunities, such as appreciation of various flora and fauna (Lindemann-Matthies et al. 2010), perception of the human–nature relationship in preserving biodiversity (Fuller et al. 2007), and environmental education for children (Soga et al. 2016). The Japanese National Biodiversity Strategy 2012–2020 has been highly concerned with the degradation of satoyama ecosystems due to their abandonment in urbanized areas, recognized as one of the national biodiversity crises (Ministry of the Environment 2012). Nevertheless, in the context of Japanese demographic shifts even in urbanized areas (United Nations 2014), the reintroduction of traditional agricultural use of satoyama would not be realistic because the number of farmers is predicted to decrease rapidly (Ministry of Agriculture, Forestry and Fisheries 2016). Consequently, we know little about an appropriate alternative usage, if any, of satoyama to ensure its biodiversity conservation, and the efficiency of such usage (Berglund et al. 2014; Katoh et al. 2009).

Three distinct land use types for satoyama ecosystems. a Agricultural use is managed by thinning to obtain timber and wood fuel at woodlands, and mowing practices eliminate pests constraining crop growth at grasslands. b Playground equipment (slide) was set in small valleys (called yato). This environment is managed by thinning to avoid tree falling and modulate brightness for park users at woodlands and mowing to prevent the appearance of harmful insects and control succession from grassland to woodlands. c Cessation of farmland activities from agricultural use land leads to abandoned land
In the search for alternative usage of satoyama ecosystems that potentially enables the maintenance of biodiversity, we focused on nature-oriented park use of satoyama ecosystems, utilizing their original scenery, topography, water features, and vegetation. Periodic human disturbance can occur under the park use: woodlands need to be managed to avoid tree falling and modulate brightness for aesthetic landscape design, and grasslands need to be managed to prevent the appearance of harmful insects and control succession from grasslands to woodlands (Fig. 1). Although motivations for management practices (i.e., periodic disturbances on biodiversity) would be different between agricultural use and park use, human disturbances may contribute to maintaining biodiversity. At the same time, in nature-oriented park, there are many artificial constructions, such as paved walking paths, playgrounds, and rest areas associated with the establishment and improvement of park environments would possibly facilitate the invasion of exotic species by increasing their invasion opportunities.
Here, we investigated the differences in diversity and species composition of plant and butterfly assemblages among three land-use types of satoyama ecosystems in the eastern part of Kanagawa prefecture, Japan: agricultural use, park use, and abandoned land (Fig. 1). We surveyed all vascular plant assemblages in the herb and shrub layer for flora and butterfly assemblages, as a representative indicator of fauna responses across three habitats, including woodland, woodland–grassland edge, and grassland in each land-use type. Butterfly assemblages are one of the significant components of satoyama ecosystems because both larvae and adults of butterflies depend on particular food plant species and flowering plant species such as Rorippa indica (L.) Hiern, Smilax china L., and Pleioblastus chino (Franch. et Sav.) Makino widely distributed across satoyama ecosystems. Focusing on both plant and butterfly assemblages is intended to perform a more holistic evaluation of biodiversity under different land use of satoyama ecosystems. Most of our study sites are located within Yokohama City, where the green coverage ratio of the area in Yokohama City has rapidly decreased from 50.0% in 1970 to 29.8% in 2009 (Yokohama Environment Planning Bureau 2013), following a high economic growth and associated urbanization. Nonetheless, relatively large greeneries currently or formerly used as satoyama have remained in highly urbanized landscapes. Specifically, we tested the following hypothesis: (i) diversity and species composition of native plants and butterflies at park-use site are similar to agricultural-use site, (ii) diversity of native plants and butterflies at abandoned site is lower than agricultural-use site, and their species composition at abandoned sites is different from agricultural-use site, and (iii) diversity of exotic plants and butterflies is higher at park-use sites than at agricultural-use sites.
Methods
Study area
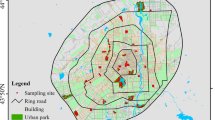
The study was conducted in the eastern part of Kanagawa prefecture, mainly in Yokohama City, and partly in Kamakura City (35°31′–57′N, 139°50′–57′E) (Fig. 2). Yokohama is the second largest city in Japan, which covers 435.290 km2 and has a population of 3.7 million (City of Yokohama 2017). After a period of rapid economic development in the 1970–1980s, Yokohama had experienced remarkable urbanization processes and population growth, with concomitant declines in the extent of natural areas (Yokohama Environment Planning Bureau 2013). The mean annual temperature was 15.8 °C, with a minimum monthly average temperature of 5.9 °C in January and a maximum of 26.7 °C in August, during the period of 1981–2010. The mean annual rainfall during the same period was 1688.6 mm. All these climatic descriptions are based on the data from the Japan Meteorological Agency.

Location of the 14 study sites in the eastern part of Kanagawa prefecture, Japan. Agricultural-use site is indicated with black circles, nature-oriented park-use site with grey triangles and abandoned site with grey squares
Study site and land-use description
We selected 14 study sites with three different land-use types (four sites for agricultural use, four sites for park use, and six sites for abandoned land) (Fig. 2). The study sites except for abandoned sites have more than 5 ha of area and are composed of the typical landforms of satoyama landscape, i.e., hills and water-eroded small valleys (Katoh et al. 2009; Berglund et al. 2014). We designated the areas of abandoned sites by our on-site survey, and the estimated areas are generally less than 1.5 ha. The small valleys have woodland and grassland, and the grassland has been historically used as paddy and crop fields. We defined the land-use types based on several criteria, including the percentage area of farmland and artificially built areas within each site, determined using ArcGIS 10.3 (ESRI, Red-lands, CA, US), the number of playground equipment, and the purpose of visit to each site by users, via questionnaires (Matsumura et al. unpublished data) (Table 1). We confirmed the trajectories of land use at the study sites over the past 100 years, using multi-temporal aerial photographs (1947–1948, 1977–1979, and 2007–2008) and Zinsoku-Sokuzu (the oldest Japanese maps that cover our study sites and were based on a plane-table survey during the 1880s). We summarized the attributes of three land-use types in Table 1 and their brief descriptions are provided below.
- i.
Agricultural-use type (4 sites; 14 survey transects): These sites mainly consist of woodlands dominated by broadleaf deciduous trees managed through thinning to obtain timber and wood fuel, and grasslands dominated by forbs managed through mowing to eliminate pests which constrain crop growth. More than 1.0% of the total area was farmlands and the purpose of the visit by users was to seek natural experience and events. Although two of these sites are named as a park (i.e., Maioka Park and Kamakura Central Park), these sites included relatively high percentage of agricultural land use and are open to public mainly for agricultural experience and enjoying agricultural landscapes (this is reflected in the result from the on-site questionnaire about the purpose of visit by users; Table 1). We, therefore, designated these sites as agricultural-use sites. These sites have traditionally been used for agricultural production at least for a century (we confirmed this evidence from Zinsoku-Sokuzu) by landowners and located within the areas including a number of important satoyama for biodiversity conservation (Ministry of the Environment 2016). We thus assumed that the agricultural-use sites are essentially similar to traditional satoyama ecosystems managed purely for agricultural production. These sites were dominated by P. chino, Oplismenus undulatifolius (Ard.) Roem. et Schult., Houttuynia cordata Thunb.
- ii.
Park-use type (4 sites; 15 survey transects): These sites mainly consist of woodlands dominated by broadleaf deciduous trees managed through thinning to avoid tree falling and modulate the brightness for park users, and grasslands dominated by forbs managed through mowing to prevent the appearance of harmful insects and control succession from grassland to woodlands. More than 1.0% of the total area was covered by artificially built surfaces, more than three playground equipment was installed, and the purpose of the visit by users was mainly to play with children and appreciate various flora and fauna. Dominant species at the ground layer were P. chino, H. cordata, and O. undulatifolius.
- iii.
Abandoned type (6 sites; 6 survey transects): These sites had been formerly used for agriculture, but were eventually abandoned for at least 3 years, as revealed by historical aerial photographs. Dominant species at the ground layer were P. chino and Persicaria thunbergii (Siebold et Zucc.) H.Gross.
Landscape variables
We estimated the area of farmland, grassland, paddy field, artificial woodland, secondary woodland and urban settlements within 1 km radius from the centre of each transect, based on aerial photographs from Google maps (Map data©2018 Google). To do this, we used Spatial Analyst 10.3 in ArcGIS (ESRI). To reduce the dimensionality of six landscape variables, we used principal component analysis (PCA). We found that two primary axes explained 50.9% of the total variance. PCA axis 1 value represented an increasing area of paddy field and a decreasing area of urban settlements. Negative PCA1 value thus indicates relatively high urbanization around the study transect (Fig. S1). PCA axis 2 value represented an increasing area of secondary woodland and a decreasing area of artificial woodland. Negative PCA2 value thus indicates increasing human interventions (Fig. S1).
Field survey design
We surveyed plant and butterfly assemblages at each site in summer (between late May and late June) and autumn (between early September and early October) 2017. At every site, each of the survey transects was established in a different valley to account for differences in environmental and topographical conditions among valleys, and a 30–40 m transect (10 or 20 m on the grassland side and 20 m on the woodland side) was established perpendicular to the woodland and grassland edge (Fig. S2). The number of transects at each site was generally three, but we increased the number of transects at the sites with relatively extensive areas and set only one transect at the abandoned sites, as we could not sample different valleys for practical reasons (in our study area, there are few extensively abandoned sites that include two or more valleys). We defined the woodland edge as the start of area where no trees with > 3 cm diameter at breast height could be found. At each transect, 1 m × 1 m quadrats were established at different distances: 20, 10, 3, and 0 m from the edge to the woodland interior, and 20, 10, and 3 m from the edge to the grassland (woodland exterior). However, we could not set quadrats at 20 m from the edge in the grassland side of 12 transects because of the limited extent of the grassland side. Consequently, our data consisted of 232 quadrats (six or seven quadrats per transect), along 35 transects, at 14 sites. We divided the quadrat into 16 grids to estimate species abundance. We recorded the vascular plant species present in each quadrat and the abundance of each species as the number of grids (ranging from 0 to 16) where the species occurred. Plant nomenclature was based on the YList (http://ylist.info).
For butterfly surveys, the transects were divided into woodland and grassland (Fig. S2). We recorded the number of species (hereafter richness) and number of individuals (hereafter abundance) of butterflies within 5 m width along the transects for 15 min on woodland and grassland sides, respectively (i.e., Pollard Walks). Butterfly sampling was conducted on sunny days between 9:30 a.m. and 15:00 p.m. in summer and autumn 2017. When we could not visually identify butterfly individuals, the individuals were caught by an insect net and were released after species identification. Butterfly nomenclature was based on the Japan Butterfly Conservation Society (2012).
Data analysis
To avoid the complexity in interpretation of results, we classified the data from quadrats at 10 and 20 m from the woodland–grassland edge in the woodland side as woodland habitat, those at the edge and at 3 m from the edge either in the woodland and grassland sides as edge habitat, and those at 10 and 20 m from the edge in the grassland side as grassland habitat (Fig. S1). We examined the differences in richness and Shannon diversity index (hereafter diversity) (Shannon and Weaver 1949) of native and exotic plant species in each habitat (woodland, edge, and grassland) among the land-use types. We also examined the differences in richness, abundance, and diversity of butterfly species in each habitat (woodland and grassland) among the land-use types. Similar to the plant species, we intended to categorize butterfly species into native and exotic ones, but we found only one exotic species (ratio of abundance of exotic butterfly species in total butterfly abundance: 0.04) (Hestina assimilis (Linnaeus, 1758)), so we examined only the native butterfly species. We also examined how landscape variables (summarized by PCA axis 1 and 2 values; Fig. S1, Tables S1, 2) affect the diversity of plant and butterfly assemblages, by including landscape variables as explanatory variables. Throughout these analyses, we used generalized linear mixed models (GLMM) with a Gaussian (for Shannon diversity) or Poisson (for plant and butterfly species richness, and butterfly abundance) error distribution. In these models, we included transect and site identity nested within season as random factor for plant analyses, and site identity nested within season as a random factor for butterfly analyses, to account for pseudo-replication.
To examine the differences in species composition of native plants and butterflies in each habitat among the land-use types, we performed an ordination based on the abundance matrix of species per quadrat for plants, and an ordination based on the abundance matrix of species per transect for butterflies, through permutational MANOVA (PERMANOVA) and visualization with non-metric multidimensional scaling (NMDS) using Bray–Curtis dissimilarity. We did not perform ordination analyses of species composition of exotic plants and butterflies, due to their scarce presence (average exotic species richness of plants per plot: 0.05). We summarized species composition of plant and butterfly assemblages with the first two NMDS axes and compared convex hulls that outline native plant species composition in each habitat under each land-use type. All statistical analyses were performed using the R software (version 3.4.2; R Development Core Team). The GLMM, PERMANOVA and NMDS analyses were performed using the glmer and lmer functions of the “lme4” package (Bates et al. 2015) and the adonis and meta-MDS function of the “vegan” package (Oksanen et al. 2011) of R, respectively.
Results
In total, we found 340 plant species (310 native and 30 exotic species), and 47 butterfly species (46 native and 1 exotic species, for a total of 670 individuals). The number of total plant species found under each of the land-use types was 262 species across agricultural-use sites, 251 species across park-use sites, and 108 species across abandoned sites. The number of total butterfly species found was 37 across agricultural-use sites, 36 across park-use sites, and 23 across abandoned sites.
Species richness and diversity of native and exotic plants did not significantly differ between agricultural-use and park-use sites across all habitats (Figs. 3, S3, Tables S1, 2). Native plant species richness and diversity were significantly lower at abandoned sites than at agricultural-use sites among all habitat types. Exotic plant species richness and diversity at park-use and abandoned sites generally did not differ significantly from agricultural-use sites. Native plant species richness and diversity had no relationships with landscape variables, regardless of the habitats, but exotic plant species richness increased with PCA axis 1 value in woodland habitats (Tables S1, 2).

Comparison of species richness of native and exotic plants among land-use types. Agricultural use was used as a reference category. Box plots represent medians (bold black horizontal line), and first and third quartiles (box perimeters), the whiskers are 90th percentiles, and the small circles are outliers. ***P < 0.001, **P < 0.01, *P < 0.05, n.s. (not significant)
Species richness, abundance, and diversity of butterflies at park-use did not significantly differ from those at agricultural-use sites across all habitats (Figs. 4, S4, Tables S1, S2). However, the diversity of butterflies at park-use sites was lower than at agricultural-use sites in grassland habitats (Fig. S4, Table S2). In addition, species richness, abundance, and diversity of butterflies were marginally lower at abandoned sites than at agricultural-use sites across all habitats (Figs. 4, S4). Furthermore, species richness and diversity of butterflies had no relationship with landscape variables, regardless of the habitats, but the abundance of butterflies increased with PCA axis 1 value in woodland habitats (Tables S1, S2).

Comparison of species richness and abundance of native butterflies among land-use types. Agricultural use was used as a reference category. Box plots represent medians (bold black horizontal line), and first and third quartiles (box perimeters), the whiskers are 90th percentiles, and the small circles are outliers. n.s. (not significant)
Species composition of native plants at park-use sites significantly differed from that at agricultural-use site across all habitats, whereas species composition of native butterflies at park-use did not (Fig. 5, Table S3). Species composition of native plants and butterflies at abandoned sites significantly differed from that at agricultural-use sites across all habitats.

Non-metric multidimensional scaling (NMDS) ordination diagram based on Bray–Curtis dissimilarities of native plant and butterfly species composition. Convex hulls outline native plant and butterfly species composition under each land-use type. Each point represents the sample quadrat with a shape according to the land-use type. The inset table represents the result of PERMANOVA comparing the species composition of native plants and butterflies among land-use types. Agricultural use was used as a reference category in performing PERMANOVA. ***P < 0.001, n.s. (not significant)
Discussion
We provide the first evidence with regard to the potential of nature-oriented park use of satoyama ecosystem for securing biodiversity conservation, by comparing diversity and species composition of plant and butterfly assemblages between park-use and agricultural-use sites (Figs. 3, 4, 5). Nonetheless, lower diversity of plant and butterfly assemblages in abandoned sites than in agricultural-use sites suggests that an abandonment of satoyama ecosystems degrades the original biodiversity, in accordance with the results of previous studies in Japan (Figs. 3, 4) (Osawa et al. 2013; Uchida et al. 2016; Uchida and Ushimaru 2014; Uematsu et al. 2010). However, our results suggest that species composition at park-use sites does not necessarily resemble that at agricultural-use sites (Fig. 5); under the park use, species composition of native butterflies was similar to that under agricultural use, whereas species composition of native plants at park-use sites differed from that at agricultural-use sites.
Although limited stakeholders (i.e., farmer) benefit from the agricultural use of satoyama ecosystems, it is likely that biodiversity conservation under the park use can directly provide various benefits for a range of stakeholders (Dallimer et al. 2012). Therefore, for striking a balance between the difficulty of maintaining agricultural use and accelerating abandonment in satoyama ecosystems in the face of ageing and shrinking population, our study highlights the utility of park use as one of the alternative usages contributing to biodiversity conservation. Yet, species composition at park use and agricultural use were different depending on taxa. This highlights the need for a more careful evaluation of biodiversity across other taxa in future studies.
We found no discernible differences in species richness and diversity of native plants and butterflies between agricultural-use and park-use sites, regardless of the habitats (Figs. 3, 4, S3, S4, Tables S1, S2). We generally observed species that are known to be specific to satoyama ecosystems such as Cephalanthera falcata (Thunb.) Blume and Chloranthus serratus (Thunb.) Roem. et Schult (Kanagawa prefecture 2016) at both park-use and agricultural-use sites. This suggests that, although the management purpose is different, park management could mimic environmental modification by traditional agroforestry practices, thereby resulting in the maintenance of biodiversity. In addition, generally, we did not find significant effects of landscape variables on species richness and diversity of native plants and butterflies (Tables S1, S2). We thus argue that, even if the management purpose is different among land-use types, maintaining periodic and intermediate-level disturbances is essential for biodiversity conservation regardless of surrounding landscape contexts (Aguilera et al. 2018). However, species richness of exotic plants and abundance of native butterflies were significantly increased with area of paddy, grassland and secondary woodland (PCA axis 1) in woodland habitats (Table S1). Potential interactive effects between land use types and surrounding landscapes need to be further explored in future studies.
However, we observed that species composition of native plants at park-use sites significantly differed from those at agricultural-use sites (Fig. 5, Table S3). Aggemyr et al. (2018) suggested that species diversity and composition differ in their response to landscape properties, and species composition strongly depended on local habitat environmental heterogeneity. Although species composition of native butterflies at park-use sites did not significantly differ from those at agricultural-use sites (Fig. 5, Table S3), diversity of native butterflies was significantly lower at park-use sites than at agricultural-use sites in grassland habitats (Fig. S4, Table S2). This might be reflected that some butterfly species (e.g., Limenitis glorifica (Fruhstorfer, 1909)) were rarely observed in grassland habitats at park-use sites. Previous studies reported that species replacement would occur due to land-use changes through the extinction of specialist species (e.g., Bagaria et al. 2019). We thus argue that species composition of native butterflies may probably shift in the future at park-use sites.
In contrast to our hypothesis, invasion of exotic species due to an increasing number of artificially built areas was not observed under the park use (Fig. 3, Table S1). Previous studies have also reported that the invasion of exotic species is limited to the naturalized areas in urban landscapes, where the disturbance intensity is relatively low, in contrast with highly urbanized areas (Uddin et al. 2013), and that occupation of complementary niches by native plant species prevents biological invasion (Elton 1958; Hector et al. 2001).
We observed a decrease in native plant diversity due to the abandonment of satoyama (as suggested by Fig. 3). This would be attributable to the absence of disturbance, enhancing competitively superior, taller species such as P. chino and Pueraria lobata (Willd.) Ohwi, which restrict the amount of sunlight available to shorter species (Kitazawa and Ohsawa 2002). Furthermore, species richness, abundance and diversity of native butterflies at abandoned sites were marginally lower than at agricultural-use sites across habitats (Figs. 4, S4, Tables S1, S2). Abandonment would lead to a decrease in trophic resources formerly supported by farmland areas (Stefanescu et al. 2009) and concomitant changes in plant community structure by the dominance of tall, competitive superior plant species (Kitazawa and Ohsawa 2002).
Implications
Our finding on the potential of nature-oriented park use of satoyama for securing biodiversity conservation can also be applicable to other areas of urbanized landscapes in Japan facing accelerated satoyama abandonment. We argue that, although management regimes under park use would not conserve biodiversity of all taxa, they would maintain aesthetic sceneries such as landscapes surrounded by hilly woodlands and bright habitats inheriting the legacy of satoyama. Moreover, instead of provisioning services such as timber and crop under the agricultural use, the park use will provide a range of cultural services for city dwellers, such as aesthetic and recreational value (Bolund and Hunhammar 1999). These multiple benefits created by park use should be evaluated in further studies.
Current opinions on the future usage of semi-natural ecosystems have been extensively debated worldwide, as to whether we need to re-wilding them through abandonment or to conserve the “nature” of semi-natural ecosystems by continuing human involvement (Navarro and Pereira 2012; Queiroz et al. 2014). This controversy stems from the fact that the impact of abandonment of semi-natural ecosystems on biodiversity is not consistent across geographic regions and environmental contexts. However, previous studies in Japan and our results consistently indicate that biodiversity in satoyama ecosystems would decrease if they are abandoned. Hence, counteracting the abandonment of satoyama would be essentially important for the biota as well as for human well-being. Nature-oriented park use is one of such counteracting options. We do not suggest that continuing the agricultural use of satoyama is not in line with the current socio-demographic contexts and needs to be replaced by park use. As the agricultural use of satoyama itself includes a variety of traditional knowledge and cultural heritages (Takeuchi 2010), the National Spatial Strategy such as agriculture enhancement and agricultural landscape conservation (Ministry of Land, Infrastructure, Transport and Tourism 2015) that secures its sustainability should be simultaneously implemented. Balancing the wise use of satoyama in urbanized landscapes is a fundamental step toward conservation of its biodiversity and human–nature interactions for future generations in this urban era.
References
Aggemyr E, Auffret AG, Jädergård L, Cousins SAO (2018) Species richness and composition differ in response to landscape and biogeography. Landsc Ecol 33:2273–2284. https://doi.org/10.1007/s10980-018-0742-9
Aguilera G, Ekroos J, Perssoon AS, Pettersson LB, Öckinger E (2018) Intensive management reduces butterfly diversity over time in urban green spaces. Urban Ecosyst 22:335–344. https://doi.org/10.1007/s11252-018-0818-y
Aoshima I, Uchida K, Ushimaru A, Sato M (2018) The influence of subjective perceptions on the valuation of green spaces in Japanese urban areas. Urban For Urban Green 34:166–174. https://doi.org/10.1016/j.ufug.2018.06.018
Bagaria G, Rodà F, Pino J (2019) Extinction and colonisation of habitat specialists drive plant species replacement along a Mediterranean grassland-forest succession. J Veg Sci 30:331–340. https://doi.org/10.1111/jvs.12722
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using ‘lme4’. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Berglund BE, Kitagawa J, Lagerås P et al (2014) Traditional farming landscapes for sustainable living in Scandinavia and Japan: global revival through the Satoyama initiative. Ambio 43:559–578. https://doi.org/10.1007/s13280-014-0499-6
Bolund P, Hunhammar S (1999) Ecosystem services in urban areas. Ecol Econ 29:293–301. https://doi.org/10.1016/S0921-8009(99)00013-0
Bonari G, Fajmon K, Malenovský I et al (2017) Management of semi-natural grasslands benefiting both plant and insect diversity: the importance of heterogeneity and tradition. Agric Ecosyst Environ 246:243–252. https://doi.org/10.1016/j.agee.2017.06.010
City of Yokohama (2017) Population News No. 1095 (Nov. 1, 2017). Statistical look at Yokohama. http://www.city.yokohama.lg.jp/ex/stat/jinko/news-j.html. Accessed 27 Dec 2018
Dallimer M, Irvine KN, Skinner AMJ et al (2012) Biodiversity and the feel-good factor: understanding associations between self-reported human well-being and species richness. Bioscience 62:47–55. https://doi.org/10.1525/bio.2012.62.1.9
Elton C (1958) The ecology of invasions by animals and plants. Methuen, London
Foley JA, Defries R, Asner GP et al (2005) Global consequences of land use. Science 309:570–574. https://doi.org/10.1126/science.1111772
Fukamachi K, Oku H, Nakashizuka T (2001) The change of a satoyama landscape and its causality in Kamiseya, Kyoto Prefecture, Japan between 1970 and 1995. Landsc Ecol 16:703–717. https://doi.org/10.1023/A:1014464909698
Fuller RA, Irvine KN, Devine-Wright P et al (2007) Psychological benefits of greenspace increase with biodiversity. Biol Lett 3:390–394. https://doi.org/10.1098/rsbl.2007.0149
Grimm NB, Faeth SH, Golubiewski NE et al (2008) Global change and the ecology of cities. Science 319:756–760. https://doi.org/10.1126/science.1150195
Hector A, Dobson K, Minns A et al (2001) Community diversity and invasion resistance: an experimental test in a grassland ecosystem and a review of comparable studies. Ecol Res 16:819–831. https://doi.org/10.1046/j.1440-1703.2001.00443.x
Japan Butterfly Conservation Society (2012) Field guide to the butterflies of Japan. Japan Butterfly Conservation Society, Tokyo (in Japanese)
Kanagawa prefecture (2016) Kanagawa biodiversity plan. http://www.pref.kanagawa.jp/docs/t4i/cnt/f12655/p1042709.html. Accessed 22 Dec 2018
Katoh K, Sakai S, Takahashi T (2009) Factors maintaining species diversity in satoyama, a traditional agricultural landscape of Japan. Biol Conserv 142:1930–1936. https://doi.org/10.1016/j.biocon.2009.02.030
Kitazawa T, Ohsawa M (2002) Patterns of species diversity in rural herbaceous communities under different management regimes, Chiba, central Japan. Biol Conserv 104:239–249. https://doi.org/10.1016/S0006-3207(01)00170-7
Koyanagi T, Kusumoto Y, Yamamoto S et al (2009) Historical impacts on linear habitats: the present distribution of grassland species in forest-edge vegetation. Biol Conserv 142:1674–1684. https://doi.org/10.1016/j.biocon.2009.03.002
Koyanagi T, Kusumoto Y, Yamamoto S, Takeuchi K (2012) Potential roles of small and linear habitat fragments in satoyama landscapes for conservation of grassland plant species. Urban Ecosyst 15:893–909. https://doi.org/10.1007/s11252-012-0253-4
Lindemann-Matthies P, Junge X, Matthies D (2010) The influence of plant diversity on people’s perception and aesthetic appreciation of grassland vegetation. Biol Conserv 143:195–202. https://doi.org/10.1016/j.biocon.2009.10.003
Ministry of Agriculture, Forestry and Fisheries (2016) Annual Report on Food, Agriculture and Rural Areas in Japan. http://www.maff.go.jp/e/data/publish/index.html#Annual. Accessed 15 July 2018
Ministry of Land Infrastructure Transport and Tourism (2015) National Spatial Strategy (National Plan). http://www.mlit.go.jp/common/001127196.pdf. Accessed 14 July 2018
Ministry of the Environment (2010) Action plan for the conservation and sustainable use of socio-ecological production landscapes (Satochi-satoyama)
Ministry of the Environment (2012) The National Biodiversity Strategy of Japan 2012–2020
Ministry of the Environment (2016) Important Satochi Satoyama 500. (in Japanese)
Navarro LM, Pereira HM (2012) Rewilding abandoned landscapes in Europe. Ecosystems 15:900–912. https://doi.org/10.1007/978-3-319-12039-3_1
Normile D (2016) Nature from nurture. Science 351:908–910. https://doi.org/10.1126/science.351.6276.908
Ohwaki A (2015) Ground arthropod communities in paddy fields during the dry period: comparison between different farming methods. J Asia Pac Entomol 18:413–419. https://doi.org/10.1016/j.aspen.2015.05.001
Oksanen AJ, Blanchet FG, Kindt R et al. (2011) Package “vegan”. R package version 2.4-3. https://cran.r-project.org, https://github.com/vegandevs/vegan. Accessed 14 Apr 2018
Osawa T, Kohyama K, Mitsuhashi H (2013) Areas of increasing agricultural abandonment overlap the distribution of previously common, currently threatened plant species. PLoS ONE 8:1–9. https://doi.org/10.1371/journal.pone.0079978
Queiroz C, Beilin R, Folke C, Lindborg R (2014) Farmland abandonment: threat or opportunity for biodiversity conservation? A global review. Front Ecol Environ 12:288–296. https://doi.org/10.1890/120348
Sala OE (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774. https://doi.org/10.1126/science.287.5459.1770
Shannon CE, Weaver W (1949) The mathematical theory of communication. Math theory Commun 27:117. https://doi.org/10.2307/3611062
Soga M, Koike S (2013) Mapping the potential extinction debt of butterflies in a modern city: implications for conservation priorities in urban landscapes. Anim Conserv 16:1–11. https://doi.org/10.1111/j.1469-1795.2012.00572.x
Soga M, Gaston KJ, Koyanagi TF et al (2016) Urban residents’ perceptions of neighbourhood nature: does the extinction of experience matter? Biol Conserv 203:143–150. https://doi.org/10.1016/j.biocon.2016.09.020
Stefanescu C, Peñuelas J, Filella I (2009) Rapid changes in butterfly communities following the abandonment of grasslands: a case study. Insect Conserv Divers 2:261–269. https://doi.org/10.1111/j.1752-4598.2009.00063.x
Takeuchi K (2010) Rebuilding the relationship between people and nature: the Satoyama Initiative. Ecol Res 25:891–897. https://doi.org/10.1007/s11284-010-0745-8
Takeuchi K, Brown DR, Washitani I et al (2003) Satoyama: the traditional rural landscape of Japan. Springer, Tokyo
Uchida K, Ushimaru A (2014) Biodiversity declines due to abandonment and intensification of agricultural lands: patterns and mechanisms. Ecol Monogr 84:637–658. https://doi.org/10.1890/13-2170.1
Uchida K, Takahashi S, Shinohara T, Ushimaru A (2016) Threatened herbivorous insects maintained by long-term traditional management practices in semi-natural grasslands. Agric Ecosyst Environ 221:156–162. https://doi.org/10.1016/j.agee.2016.01.036
Uchida K, Fujimoto H, Ushimaru A (2018a) Urbanization promotes the loss of seasonal dynamics in the semi-natural grasslands of an East Asian megacity. Basic Appl Ecol 29:1–11. https://doi.org/10.1016/j.baae.2018.03.009
Uchida K, Koyanagi TF, Matsumura T, Koyama A (2018b) Patterns of plant diversity loss and species turnover resulting from land abandonment and intensification in semi-natural grasslands. J Environ Manag 218:622–629. https://doi.org/10.1016/j.jenvman.2018.04.059
Uddin MB, Steinbauer MJ, Jentsch A et al (2013) Do environmental attributes, disturbances and protection regimes determine the distribution of exotic plant species in Bangladesh forest ecosystem? For Ecol Manag 303:72–80. https://doi.org/10.1016/j.foreco.2013.03.052
Uematsu Y, Koga T, Mitsuhashi H, Ushimaru A (2010) Abandonment and intensified use of agricultural land decrease habitats of rare herbs in semi-natural grasslands. Agric Ecosyst Environ 135:304–309. https://doi.org/10.1016/j.agee.2009.10.010
United Nations (2014) World urbanization prospects the 2014 revision. United Nations, New York
Yokohama Environment Planning Bureau (2013) The Yokohama Green-Up plan. http://www.city.yokohama.lg.jp/kankyo/midoriup/english/. Accessed 12 Apr 2018
Acknowledgements
We thank our laboratory members for helping with fieldwork, especially Taiki Tachibana, Kousuke Tachibana, Akihito Goto, Rio Matsumura, Daichi Makishima, Hiroki Koyata, and Rui Suto. This work was supported by Hitachi Global Foundation (Grant Number 1331) and the Environment Research and Technology Development Fund (Grant Number S-15-2) of the Ministry of the Environment, Japan.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Iwachido, Y., Uchida, K., Ushimaru, A. et al. Nature-oriented park use of satoyama ecosystems can enhance biodiversity conservation in urbanized landscapes. Landscape Ecol Eng 16, 163–172 (2020). https://doi.org/10.1007/s11355-020-00413-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-020-00413-y