
Abstract
Bladder cancer (BC) is the most common cancer of the urinary tract. Bozepinib (BZP), a purine-derived molecule, is a potential compound for the treatment of cancer. Purinergic signaling consists of the activity of nucleosides and nucleotides present in the extracellular environment, modulating a variety of biological actions. In cancer, this signaling is mainly controlled by the enzymatic cascade involving the NTPDase/E-NPP family and ecto-5’-nucleotidase/CD73, which hydrolyze extracellular adenosine triphosphate (ATP) to adenosine (ADO). The aim of this work is to evaluate the activity of BZP in the purinergic system in BC cell lines and to compare its in vitro antitumor activity with cisplatin, a chemotherapeutic drug widely used in the treatment of BC. In this study, two different BC cell lines, grade 1 RT4 and the more aggressive grade 3 T24, were used along with a human fibroblast cell line MRC-5, a cell used to predict the selectivity index (SI). BZP shows strong antitumor activity, with notable IC50 values (8.7 ± 0.9 µM for RT4; 6.7 ± 0.7 µM for T24), far from the SI for cisplatin (SI for BZP: 19.7 and 25.7 for RT4 and T24, respectively; SI for cisplatin: 1.7 for T24). BZP arrests T24 cells in the G2/M phase of the cell cycle, inducing early apoptosis. Moreover, BZP increases ATP and ADP hydrolysis and gene/protein expression of the NPP1 enzyme in the T24 cell line. In conclusion, BZP shows superior activity compared to cisplatin against BC cell lines in vitro.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bladder cancer (BC) is one of the 10 most common cancers worldwide, accounting for 3% of all diagnoses [1]. The likelihood of developing BC is higher in smokers, which is the main risk factor for the disease [2]. Around 20% of BC cases are caused by occupational exposure to raw materials for paints, diesel, metallurgical industries, and car factories, among others [2]. As a result, men are diagnosed with BC four times more often than women, making it the sixth most common neoplasm and the ninth most fatal cancer in men [1, 3]. In addition, approximately 80% of people with the disease are adults over the age of 65 [4]. The treatment of BC depends on the pathological stage of the disease at the time of diagnosis. Around 70% of BC diagnoses are non-muscle-invasive bladder cancers (NMIBC), which are usually treated with transurethral resection of the bladder tumor (TURBT) accompanied by intravesical immunotherapy with Bacillus Calmette-Guérin (BCG) or intravesical chemotherapy for tumors with a greater chance of progression or recurrence [5, 6]. NMIBC is one of the neoplasms with the highest cost of treatment per patient since it requires constant observation and resections [7]. Muscle-invasive bladder cancer (MIBC) is often treated with radical cystectomy with bilateral pelvic lymphadenectomy and neoadjuvant cisplatin-based chemotherapy [8]. Metastatic BC or unresectable BC is treated with cisplatin-based chemotherapy combined with other available chemotherapeutic agents, resulting in a median survival of 14 months [5]. However, the prognosis of patients with BC is poor, with a 5-year recurrence rate of up to 70% and a 10–30% risk of progression [9].
The compound [(RS)-2,6-dichloro-9-[1(p-nitrobenzenesulfonyl)-1,2,3,5-tetrahydro-4,1-benzoxazepine-3yl]-9 H-purine] is called bozepinib (BZP) and has been studied in the treatment of cancer. We and others have already shown its antitumor activity and selectivity for cancer cells in glioblastoma (GBM), breast and colon cancer [10,11,12].
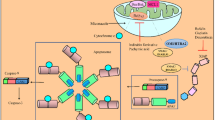
Extracellular purines can interact with purinergic receptor molecules, which can be P1 (adenosine receptor) or P2 (extracellular nucleotide receptor) [13]. Extracellular adenosine (ADO) production can be obtained through an enzymatic cascade involving the E-NTPDase and E-NPP family, which is responsible for hydrolyzing extracellular adenosine triphosphate (ATP) to adenosine diphosphate (ADP) and then to adenosine monophosphate (AMP) [14] Finally, AMP is hydrolyzed to ADO by the enzyme ecto-5′-nucleotidase/CD73 [15]. It is well known that this system is involved in several types of cancer [16,17,18]. Previous work from our group showed an unequal pattern of ectonucleotidase expression and activity in malignant BC cells (T24) compared to early stage cells (RT4) [19]. In addition, we have shown an altered expression of NTPDase3 and CD73 enzymes during BC progression in an in vivo model of bladder cancer in mice [20].
Purine-derived compounds, such as BZP, have a potent antitumor activity and could be a new bet for cancer treatment [21]. Interestingly, BZP showed activity on the purinergic system, inhibiting the activity of ecto-5’-nucleotidase/CD73 along with the formation of adenosine, without altering the expression of the CD73 enzyme. On the other hand, an increase in the expression and activity of the enzyme ENTPDase1/CD39 was observed [10]. Taking all this information together, the aim of the present work was to evaluate the cytotoxic effect of BZP in BC cell lines and the potential effect on the purinergic system.
Materials and methods
Cell culture
The human bladder cancer (BC) cell lines RT4 (grade 1) and T24 (grade 4) and the normal lung fibroblast cell line MRC-5 were obtained from the American Type Culture Collection (ATCC, USA). The MRC-5 and RT4 cell lines were cultured in Dulbecco’s modified Eagle medium (DMEM) and the T24 cell line was cultured in Roswell Park Memorial Institute’s (RPMI) 1640 medium (pH 7.4), supplemented with 10% fetal bovine serum (FBS) containing 0.5 U/mL of the antibiotics penicillin/streptomycin and kept in an incubator at 37 °C with 95% relative humidity and 5% CO2.
Treatments
Bozepinib (BZP) was dissolved in dimethyl sulfoxide (DMSO) and stored at -20 °C. For cell viability experiments, cells were seeded in 24-well plates (30 × 104, 30 × 104 and 20 × 104, for MRC-5, RT4 and T24, respectively) and grown for 48 h until semi-confluence. BC cells were treated with increasing concentrations of BZP from 1 to 10 µM, whereas the MRC-5 cell line was treated with 80 to 180 µM for 24 h. For the other experiments, BC cell lines were seeded in 24-well or 6-well plates (100 × 104 cells for the T24 cell line) and treated with inhibitory concentrations of 25 and 50 (IC25 and IC50, respectively) (3 and 6 µM for T24; 4 and 8 µM for RT4).
Cisplatin, obtained from Sigma-Aldrich, was dissolved in type I water and stored at -20 °C. For cell viability experiments, T24 and MRC-5 cells were seeded in 24-well plates and grown to semi-confluence for 48 h and treated with increasing concentrations of cisplatin from 80 to 150 µM for the T24 cells and 150 to 300 µM for the MRC-5 cells for 24 h. For the other experiments, the T24 cell line was seeded in 24-well plates and treated with concentrations of IC25 and IC50 (49 and 98 µM).
Cell viability assay
After the described treatments, the cells were washed with phosphate buffer saline (PBS) (pH 7.4), trypsinized, and immediately counted by the trypan blue exclusion assay (0.1%). The membranes of live cells remained intact without dye penetration, whereas the membranes of dead cells were ruptured and stained [22]. Results were expressed as mean percentage ± standard deviation of cell viability.
Cell cycle assay
At the end of the treatment, the supernatant was collected, and the cells were trypsinized and centrifuged at 400 × g for 6 min. They were immediately suspended in a staining solution containing 3.5 mM sodium citrate; 0.5 mM TRIS.HCl (pH 7.6); Nonidet P-40 0.1% (v/v); 100 µg/mL RNAse; 50 µg/mL propidium iodide (PI) and incubated for 15 min at room temperature, protected from light. Data were obtained using a flow cytometer (Accuri C6, BD Biosciences, USA) and the results were analyzed using FlowJo® software (USA).
Annexin V-FITC/PI assay
After 24 h of treatment, the supernatant was collected and the cells were trypsinized and centrifuged at 400 × g for 6 min. They were suspended using the Annexin V-FITC/PI (BD Biosciences, USA) labeling kit and incubated at room temperature for 15 min, protected from light. Data and results were obtained using a flow cytometer (Accuri C6, BD Biosciences, USA) and analyzed using FlowJo® software (USA).
Acridine orange staining
At the end of the treatments described above, we measured the formation of acidic vesicular organelles (AVOs) by flow cytometry. Briefly, the T24 cell line was washed with PBS, trypsinized, and centrifuged twice at 400 × g for 6 min. The cells were then incubated with acridine orange (1 µg/mL) for 15 min in the dark and analyzed by flow cytometry (Accuri, BD Biosciences, USA). The results were analyzed using FlowJo® software (USA).
CD133 and PD-L1 immunocontent
After completion of the treatment described above, the culture medium was discarded, the wells were washed with blocking buffer containing PBS with 2% FBS, and the T24 cells were trypsinized and centrifuged at 400 × g for 6 min and incubated with anti-PDL1 (cat. 558,017, BD Biosciences, USA) and anti-CD133 (cat. 566,594, BD Biosciences, USA) antibodies for 30 min in the dark. The data were obtained using a flow cytometer (FACSCalibur, BD Biosciences, USA) and analyzed using FlowJo® software (USA).
Treatment cycles
At the end of the 3 µM and 6 µM BZP treatments, the T24 cells were washed with PBS, trypsinized, trypan blue dye solution (0.1%) was added, and the cells were counted using a Neubauer chamber. Cells that remained alive were reseeded according to the number of viable cells in the 6 µM BZP treatment. The cells were allowed to grow to semi-confluence and were retreated with BZP, and the above protocol was repeated until the cells exposed to the 6 µM BZP treatment were completely eliminated.
ATP, ADP and AMP hydrolysis
After treatment, the cells were washed three times with phosphate-free incubation medium. The enzymatic reaction was initiated by adding incubation medium (2 mM MgCl2/CaCl2, 120 mM NaCl, 5 mM KCl, 10 mM glucose, and 20 mM Hepes, pH 7.4) containing 2.5 mM ATP or ADP or 2 mM AMP at 37 °C [19]. After 30 min of incubation, 150 µL of the incubation medium was collected and transferred to Eppendorf tubes containing 150 µL of trichloroacetic acid (5% w/v) on ice. Inorganic phosphate release was measured using the malachite green method [23]. Protein concentration was quantified using the Coomassie blue method [24]. Specific activity was expressed as nanomoles of Pi released per minute per milligram of protein.
CD39, CD73 and NPP1 immunocontent
After the treatment described above, the cell culture medium was removed, the wells were washed with blocking buffer, and the T24 cells were trypsinized and centrifuged at 400 × g for 6 min and incubated with PE anti-CD73 (cat. 550,257, BD Biosciences, USA) or APC anti-CD39 (cat. 560,239, BD Biosciences, USA) monoclonal antibodies for 30 min, while for the NPP1 enzyme expression the T24 cells were incubated with guinea pig anti-NPP1 polyclonal antibody (ectonucleotidases-ab.com, CA) for 30 min. The cells were then washed again with blocking buffer and incubated with anti-guinea pig secondary antibody (cat. SAB4600040, Sigma, USA) for 30 min, followed by washing with blocking buffer and resuspension in PBS. Data were obtained using a flow cytometer (Accuri C6, BD Biosciences, USA) and the results were analyzed using FlowJo® software (USA).
RNA extraction and quantitative real-time polymerase chain reaction
Total RNA was isolated from human BC cell line T24 using TRIzol reagent (Invitrogen) according to the manufacturer’s instructions. The quality and concentration of total RNA were determined by spectrophotometry. Reverse transcription was performed with 2 µg RNA plus M-MLV RT (Promega Corporation, Madison, WI, USA) and 0.5 µg random hexamer according to the manufacturer’s protocol. Real-time PCR was performed in triplicate on a StepOne Plus™ real-time PCR system (Applied Biosystems) using GoTaq® qPCR Master Mix (Promega Corporation, Madison, WI, USA), also following the manufacturer’s instructions. All primer sequences used in this study are described in Table 1. Quantitative analysis of data was performed using the ΔΔCt method [25]. Values were normalized with endogenous control GNB2L1 and expressed as relative expression levels.
Statistical analysis
The data obtained were analyzed using GraphPad Prism® software with one-way ANOVA and Tukey’s post-test. Results are expressed as mean ± SD of three independent experiments. Differences were considered significant at p < 0.05.
Results
BZP reduces BC cell viability with a significant in vitro selectivity index
First, we evaluated the in vitro potential of BZP against two BC cell lines (T24 and RT4). For this purpose, the half maximal inhibitory concentration (IC50) was determined and compared with MRC-5 cells. BZP reduced the percentage of viable BC tumor cells at lower IC50 concentrations compared to MRC-5 cells (Table 2).
The in vitro selectivity index (SI) was determined by the ratio of the IC50 of the MRC-5 lineage to the IC50 of the BC tumor cell lines. As shown in Table 2, the IC50 of MRC-5 cells is approximately 19-fold and 25-fold greater than the IC50 of the RT4 and T24 cell lines, respectively.
Cisplatin reduces the viability of T24 cells with a lower selectivity index
Next, we decided to compare the effects of BZP with cisplatin, one of the most commonly used drugs in the treatment of bladder cancer. For this, we used the most aggressive BC cell line, T24. We observed that the IC50 of BZP (6.71 ± 0.75, Table 2) was significantly lower than the IC50 of cisplatin for the T24 cell line (97.98 ± 5.87, Table 2). Most importantly, BZP has a higher in vitro SI (25.77) than cisplatin (1.70).
BZP and cisplatin induce cell cycle arrest and apoptosis in BC cell lines
After observing a significant reduction in cell viability in BC cell lines, we decided to evaluate the effect of BZP on cell cycle progression. In the T24 cell line, we observed an increase in the percentage of cells in the G2/M phase (18.2 ± 1.29%) at 3 µM BZP (Fig. 1a), and in the S (33.0 ± 1.36%) and G2/M (20.3 ± 1.1%) phases at 6 µM BZP (Fig. 1a), indicating a gradual arrest in cell cycle progression depending on the concentration. Consequently, there was a decrease in the percentage of cells in the G0/G1 phase at 3 µM and 6 µM BZP (43.0 ± 1.05% and 39.2 ± 1.17%, respectively) compared to the DMSO control, G0/G1 (50.9 ± 1.11%), S (28.5 ± 1.37%), and G2/M (13.8 ± 0.47%) (Fig. 1a). We did not observe any interference in cell cycle progression after BZP treatment for the RT4 lineage compared to the DMSO control (Fig. 1b).

BZP and cisplatin alter cell cycle progression and cause autophagy and apoptosis in BC cell lines, The BC cell lines were treated with BZP (3 µM/6 µM for the T24 line and 4 µM/8 µM for the RT4 line) for 24 h. The T24 cell line was treated with cisplatin (49 µM/98 µM) for 24 h. (a) Histograms and cell cycle quantification of the T24 cell line. (b) Histograms and cell cycle quantification of the RT4 cell line. (c) Histograms and cell cycle quantification of the T24 cell line. (d) Dot-plots (Q1 = early apoptosis, Q2 = late apoptosis, Q3 = necrosis and Q4 = viable cells) and quantification of apoptosis by % of T24 cell line. (e) Dot-plots and quantification of apoptosis by % of RT4 cell line. (f) Dot-plots and quantification of apoptosis by % of T24 cell line. (g) Quantification of AVO by % of cells. Data are presented as mean ± SD of three independent experiments (*p < 0.05, **p < 0.01, ***p < 0.001 vs. DMSO). CIS = cisplatin
We also evaluated the effects of cisplatin on the cell cycle of the T24 cell line. We observed an increase in the percentage of cells in the G0/G1 phase with 49 µM and in the sub-G1 phase with 98 µM cisplatin (62.6 ± 0.93% and 7.1 ± 1.42%, respectively) (Fig. 1c). Arrest in the G0/G1 phase leads to a reduction in the percentage of cells in the S and G2/M phases (15.5 ± 1.63% and 17.8 ± 1.09%, respectively) at 49 µM, and in the S phase (17.7 ± 5.83%) at 98 µM cisplatin compared to the control, G0/G1 (52.3 ± 1.45%), S (22.5 ± 1.18%), G2/M (23.3 ± 0.96%), and sub-G1 (1.8 ± 0.48%) (Fig. 1c).
To assess the type of cell death caused by BZP in BC tumor cell lines, the Annexin V-FITC/PI assay was performed. Flow cytometry analysis showed a low percentage of PI or PI + Annexin-labeled cells, indicating no necrosis or late apoptosis (Q2 and Q3, Fig. 1d). On the other hand, we observed a significant increase in Annexin V-labeled cells after 6 µM of BZP treatment (11.3 ± 0.94%) in the T24 cell line (Q1, Fig. 1d) and after 8 µM of BZP treatment (49.7 ± 14.57%) in the RT4 cell line (Q1, Fig. 1e), indicating early apoptotic cell death in the RT4 cells. Unlabeled cells were considered viable (Q4, Fig. 1e).
We also analyzed cisplatin treatment followed by Annexin V-FITC/PI assay. Cisplatin treatments show a greater number of cells labeled with Annexin V and PI (11.3 ± 0.76% and 27.2 ± 0.64% for 49 µM and 98 µM, respectively), indicating late apoptosis or necrosis in the T24 cell line (Q2, Fig. 1f).
We decided to evaluate AVO since previous studies have already shown the ability of cisplatin to induce autophagy [26]. We observed that cisplatin was able to increase AVO with 98 µM of treatment as expected (Fig. 1g). BZP was also able to induce AVO formation after 6 µM of treatment (Fig. 1g).
BZP enhances a cellular resistance phenotype after one cycle of treatment, but can kill them after three cycles of treatment
Our next step was to evaluate the effects of BZP on markers of cellular resistance. Thus, we targeted an important immune checkpoint ligand, PD-L1 [27]. We found that one cycle of treatment with BZP at a concentration of 6 µM (Fig. 2a) was able to increase the number of cells expressing PD-L1. We also evaluated the possibility of BZP selecting CD133 + cells. We did not observe any significant results from the BZP treatments compared to the DMSO control (Fig. 2b).

BZP induces an increase in PD-L1 expression and disables T24 cells after three cycles of treatment. BC of the T24 cell line was treated with BZP (3 µM/6 µM) for 24 h. (a) Quantification of PD-L1 by % of cells and expression of PD-L1, represented by MFI (mean fluorescence intensity). (b) Quantification of CD133 by % of cells and expression of CD133, represented by MFI. (c) Cell viability was determined by the trypan blue exclusion test. (d) Quantification of CD133 by % of cells and CD133 expression, represented by MFI, after four treatment cycles. Data are presented as mean ± SD of three independent experiments (*p < 0.05, **p < 0.01, ***p < 0.001 vs. DMSO)
We then decided to perform treatment cycles with BZP on T24 cells, simulating chemotherapy treatment, to analyze tumor resistance. Three cycles of BZP treatment were sufficient to virtually eliminate the T24 cells at a concentration of 6 µM (Fig. 2c). The remaining cells (0.96 ± 0.41%) did not replicate for a new treatment cycle. Cells treated with 3 µM showed a resistance profile with constant cell viability (69.4 ± 6.66%, 70.7 ± 5.11% and 56.8 ± 8.39% for the first, second and third treatment cycles, respectively) (Fig. 2c). Considering this resistance profile, we decided to analyze the frequency of cells expressing CD133 after 3 µM of BZP treatment. We did not observe any significant results in CD133 expression from the BZP treatment compared to the DMSO control (Fig. 2d).
BZP increases ATP and ADP hydrolysis and gene/protein expression of the NPP1 enzyme in the T24 cell line
Since BZP is a purine-derived compound, we decided to analyze its effects on the purinergic system. We observed a significant increase in ATP and ADP hydrolysis at 6 µM of BZP treatment in T24 cells, whereas no changes in AMP hydrolysis were observed compared to the DMSO control (Fig. 3a). In the RT4 cell line, we observed an increase in ADP hydrolysis at 8 µM with no changes in ATP and AMP hydrolysis compared to the DMSO control (Fig. 3b).

BZP induces an increase in the hydrolysis of ATP and ADP, altering the expression of the NPP1 enzyme, The BC cell lines were treated with BZP (3 µM/6 µM for the T24 line and 4 µM/8 µM for the RT4 line) for 24 h, (a) ATP, ADP and AMP hydrolysis in the T24 cell line. (b) ATP, ADP and AMP hydrolysis in the RT4 cell line. (c) Relative gene expression of ENPP1, ENTPD3 and ENTPD5 was evaluated in the T24 cell line. (d) Quantification of NPP1 by % of cells, and NPP1 expression represented by MFI in the T24 cell line. (e) Quantification of CD39 and CD73 by % of cells, and CD39/CD73 expression represented by MFI in the T24 cell line. Data are presented as mean ± SD of three independent experiments (#p < 0.05, ##p < 0.01 vs. control, *p < 0.05, **p < 0.01 vs. DMSO)
To try to explain the increase in ATP and ADP hydrolysis observed in the T24 cell line, we measured the gene expression of three possible enzymes involved in this substrate hydrolysis, NPP1, NTPDase3 and NTPDase5. We found that BZP was able to increase the gene expression of the ENPP1 enzyme after 3 µM of treatment (Fig. 3c). In addition, we also observed a significant decrease in the expression of the ENTPD3 enzyme at 6 µM (Fig. 3c) and ENTPD5 at both concentrations (3 and 6 µM) (Fig. 3c). Considering the effect of BZP on increasing the mRNA expression of the NPP1 enzyme in the T24 cell line, we decided to evaluate its effect on protein expression. Accordingly, we observed an increase in protein expression of the NPP1 enzyme after 6 µM of BZP treatment (Fig. 3d). Finally, we also evaluated the expression of the two main enzymes of the purinergic system, CD39 and CD73, but we did not find any significant results after BZP treatment compared to the DMSO control (Fig. 3e).
Discussion
Cisplatin is the chemotherapeutic agent of choice for the treatment of BC. It can be used as neoadjuvant therapy or as a single agent for metastatic BC [28]; however, the benefits of treatment are limited due to cisplatin resistance [29, 30]. In addition, individuals with comorbidities such as neuropathy, a single kidney, advanced age and cardiovascular complications should not use cisplatin [31]. Therefore, there is a lack of chemotherapy for the treatment of BC.
We observed that BZP induces cell death in BC cell lines with low IC50 values (Table 2). This confirms previous results with this compound showing low IC50 values for GBM, colon and breast cancer cell lines [10,11,12]. Based on these results, we calculated the SI in vitro, which must be considered when searching for new drugs for cancer therapy, as it serves as a preliminary safety parameter [32]. Our data show that BZP manifests a significant SI in vitro, with values of 19.74 and 25.77 for the RT4 and T24 cell lines, respectively (Table 2). Studies suggest that high SI values indicate selectivity for cancer cells, while values ≤ 2 show low selectivity [33, 34]. Compounds with an SI ≥ 10 are highly selective [35, 36]. In contrast, cisplatin has a higher IC50 value compared to BZP for the T24 cell line (Table 2). Previous work has also shown that cisplatin has a high IC50 value for the BC HT1197 cell line [37], with a considerably low selectivity index [35, 36]. It should be noted that MRC-5, although non-tumoral, is a proliferative cell, which makes the low toxicity of BZP compared to cisplatin appreciable.
The reduction in cell viability in the T24 cell line was accompanied by cell cycle arrest. We observed an increase in the number of cells in the G2/M phase due to cell cycle arrest compared to the DMSO control. As a reflection of this increase, there was a decrease in the percentage of cells in the G0/G1 phase in the T24 cell line with both treatments (Fig. 1a). The arrest of cell cycle progression in the G2/M phase caused by BZP treatment in the T24 cell line, prevents cells from progressing to mitosis and may complete action by inducing cell death by apoptosis. Other works have already observed the accumulation of cells in the G2/M phase followed by cell death by apoptosis [38,39,40]. The present work is the first report in the literature showing a perturbation in cell progression caused exclusively by BZP treatment. A previous study showed that combined treatment with BZP and the cytokine interferon-alpha (IFNα) caused an accumulation of S-phase cells in a breast cancer cell line [12]. Cisplatin also caused cell cycle arrest in the T24 cell line with both treatments (Fig. 1c), confirming what has been shown in the hepatocellular carcinoma cell line [41]. Furthermore, in a lung adenocarcinoma cell line, the cell cycle was blocked in the S and G2/M phases by the action of cisplatin [42].
As a possible consequence of the arrest in the cell cycle progression, our results show that BZP induces early apoptosis in BC cell lines, at 6 µM of treatment for the T24 cell line (Fig. 1d) and 8 µM for the RT4 cell line (Fig. 1e), in agreement with previous studies in other cancer types such as breast, colon and GBM cancer cells [10,11,12]. On the other hand, cisplatin induced T24 cell death by late apoptosis in both treatments (Fig. 1f). Studies show that cisplatin induces cell death by apoptosis in various cancers [43,44,45]. New studies in the treatment of cancer are directing the development of new drugs that make it difficult for cancer cells to escape through apoptosis, and new apoptosis inducers are beginning to be matched with currently used drugs [46]. Furthermore, cell death by early apoptosis induced by BZP is considered better than late apoptosis or necrosis induction.
To conclude our comparison, we decided to investigate the effect of BZP on AVO formation, an indication of autophagy. To confirm the induction of autophagy we need complementary experiments. In cancer, the role of autophagy is still unclear. Studies show that autophagy protects cancer cells from apoptosis and provides resistance to chemotherapy treatment [47, 48]. On the other hand, when cells are not optimally nourished or are malformed, such as in the tumor microenvironment, autophagy can cause cell death [49, 50]. Our results show that BZP at a concentration of 6 µM causes the formation of AVO (Fig. 1g). Previous work shows that BZP can induce autophagy in other cancer cell lines [10, 12]. We also observed that cisplatin induced AVO formation in the T24 cell line at 98 µM of treatment (Fig. 1g). The induction of autophagy by cisplatin treatment is already known [26]. Studies show that the increase in autophagy induction in cancer cells precedes cell death by apoptosis, so our results confirm this finding [51, 52]. Thus, AVO formation after one cycle of treatment could be associated with cell resistance or even with a cross-pathway to apoptosis induction.
In recent years, a new approach to cancer treatment has emerged: inhibition of the PD-1 or PD-L1 immune checkpoint. PD-L1 binds to its receptor, PD-1, which is expressed on the surface of immune-related lymphocytes such as T cells. This binding is able to inhibit T cell cytotoxicity and cancer cells benefit from this mechanism to protect themselves from the immune system [53, 54]. We observed that treatment with 6 µM BZP induced an increase in cells expressing PD-L1 after one cycle of treatment (Fig. 2a). This result shows a possible mechanism of cellular resistance and two more cycles of BZP treatment could kill these resistant cells (Fig. 3c). PD-1 or PD-L1 inhibitors are already being used in the treatment of BC, in combination with chemotherapy or radiotherapy [55].
Cancer stem cells (CSCs) are characterized by rapid proliferation and resistance to conventional treatments. In addition, studies indicate that these cells initiate cancer recurrence and metastasis [56, 57]. BZP has already been shown to select CD133+ cells in GBM cell lines [10], but we did not observe the same pattern in the BC T24 cell line (Fig. 2b). Next, we decided to treat the T24 cells with different treatment cycles to analyze the cell resistance. Three treatment cycles with 6 µM BZP were required for cell viability to fall below 1% (Fig. 2c), where the cells failed to replicate for a new treatment cycle, showing the efficiency of treatment in cycles with BZP. This result confirms the efficiency of BZP in combatting cancer cells, including those that may acquire resistance phenotypes. In agreement, a previous study from our group showed that two cycles of BZP treatment were necessary to eliminate GBM cells [10]. On the other hand, treatment with 3 µM BZP failed to eliminate the cells, showing constant cell viability, indicating resistance as a result of using an inappropriate concentration to induce bladder cancer cell death (Fig. 2c). Considering CD133 as a marker of cancer stem cells, we decided to reassess CD133 + cells after treatment cycles, but we did not find any significant results (Fig. 2d).
Finally, we verified whether BZP exerts some kind of modulation on the purinergic system. First, we examined the hydrolysis of ATP, ADP and AMP substrates and observed a significant increase in ATP and ADP hydrolysis, with no change in AMP hydrolysis, in the T24 cell line compared to the DMSO control (Fig. 3a). Previous data from our laboratory show that the T24 cell line expresses NTPDase5 and pyrophosphatase/phosphodiesterase 1 (NPP1) enzymes [19], therefore we analyzed the effect of BZP on the gene expression of these two enzymes. We observed that treatment with BZP was able to induce an increase in the gene expression of the ENPP1 enzyme (Fig. 3c) with a significant increase expression of NPP1 protein (Fig. 3d), while it does not alter the expression of neither the CD39 enzyme nor CD73 enzyme (Fig. 3e). NPP1 is the most important member of the NPP family, which consists of 7 members, and its main substrate is ATP, which can be hydrolyzed to ADP or AMP [14]. Studies have shown that high levels of ATP in the tumor microenvironment favor local progression and metastasis in several types of cancer [58,59,60]. In addition, ATP can activate some P2 family receptors, which have an auxiliary function in the growth of tumor and healthy cells [61]. In addition, treatment with 8 µM BZP caused a significant increase in ADP hydrolysis in the RT4 cell line (Fig. 3b). This increase in ADP hydrolysis may be due to the effect of BZP on the NPP1 enzyme [19], which can also hydrolyze ADP [14]. In addition, it is possible to suggest that the increase in the expression of NPP1 with a consequent increase in the hydrolysis of ATP and ADP would lead to an increase in the availability of AMP, the CD73 substrate, and a decrease in the availability of the two CD73 endogenous inhibitors (ATP and ADP). Thus, although BZP does not have a direct effect on CD73, under biological conditions the observed effect on NPP1 would cause an indirect effect on increase of extracellular adenosine availability.
Conclusions
Our results show the potent chemotherapeutic effect of BZP against BC cell lines, inducing a reduction in cell viability with a low IC50 and arrest in cell cycle progression followed by early apoptosis. BZP modulates the purinergic system with increased expression and activity of the NPP1 enzyme. In addition, BZP presented higher SI values compared to the reference chemotherapeutic agent, cisplatin. Although BZP induced cell resistance after one cycle of treatment, three cycles were sufficient to virtually eliminate resistant cells. Further studies are needed to better understand the mechanism of action and toxicity of BZP in vivo.
Data availability
All data are available upon reasonable request.
References
Saginala K, Barsouk A, Aluru JS et al (2020) Epidemiology of Bladder Cancer. Med Sci (Basel) 8. https://doi.org/10.3390/medsci8010015
Kogevinas M, Mannetje ’T, Cordier A S, et al (2003) Occupation and Bladder cancer among men in Western Europe. Cancer Causes Control 14:907–914. https://doi.org/10.1023/B:CACO.0000007962.19066.9c
Bray F, Ferlay J, Soerjomataram I et al (2018) Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 68:394–424. https://doi.org/10.3322/caac.21492
Mushtaq J, Thurairaja R, Nair R (2019) Bladder cancer. Surg (United Kingdom) 37:529–537. https://doi.org/10.1016/j.mpsur.2019.07.003
Bellmunt J, Orsola A, Leow JJ et al (2014) Bladder cancer: ESMO practice guidelines for diagnosis, treatment and follow-up. Ann Oncol 25:iii40–iii48. https://doi.org/10.1093/annonc/mdu223
DeGeorge KC, Holt HR, Hodges SC (2017) Bladder Cancer: diagnosis and treatment. Am Fam Physician 96:507–514
Lotan Y, Kamat AM, Porter MP et al (2009) Key concerns about the current state of Bladder cancer: a position paper from the Bladder Cancer think Tank, the Bladder Cancer Advocacy Network, and the Society of Urologic Oncology. Cancer 115:4096–4103. https://doi.org/10.1002/cncr.24463
Chang SS, Bochner BH, Chou R et al (2017) Treatment of non-metastatic muscle-invasive Bladder Cancer: AUA/ASCO/ASTRO/SUO Guideline. J Urol 198:552–559. https://doi.org/10.1016/j.juro.2017.04.086
Martinez Rodriguez RH, Buisan Rueda O, Ibarz L (2017) Bladder cancer: Present and future. Med Clin (Barc) 149:449–455. https://doi.org/10.1016/j.medcli.2017.06.009
Dias A, de Scholl F, Moritz JN CEJ, et al (2021) New insights into cytotoxic mechanisms of bozepinib against glioblastoma. Eur J Pharm Sci 162. https://doi.org/10.1016/j.ejps.2021.105823
Ramírez A, Boulaiz H, Morata-Tarifa C et al (2014) HER2-signaling pathway, JNK and ERKs kinases, and cancer stem-like cells are targets of Bozepinib small compound. Oncotarget 5:3590–3606. https://doi.org/10.18632/oncotarget.1962
Marchal JA, Carrasco E, Ramirez A et al (2013) Bozepinib, a novel small antitumor agent, induces PKR-mediated apoptosis and synergizes with IFNα triggering apoptosis, autophagy and senescence. Drug Des Devel Ther 7:1301–1313. https://doi.org/10.2147/DDDT.S51354
Kim SH, Choi JY (2022) Purinergic signaling in the peripheral vestibular system. Purinergic Signal 18:165–176. https://doi.org/10.1007/s11302-022-09855-5
Zimmermann H, Zebisch M, Sträter N (2012) Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal 8:437–502. https://doi.org/10.1007/s11302-012-9309-4
Colgan SP, Eltzschig HK, Eckle T, Thompson LF (2006) Physiological roles for ecto-5’-nucleotidase (CD73). Purinergic Signal 2:351–360. https://doi.org/10.1007/s11302-005-5302-5
Reyna-Jeldes M, Díaz-Muñoz M, Madariaga JA et al (2021) Autocrine and paracrine purinergic signaling in the most lethal types of cancer. Purinergic Signal 17:345–370. https://doi.org/10.1007/s11302-021-09785-8
Campos-Contreras ADR, Díaz-Muñoz M, Vázquez-Cuevas FG (2020) Purinergic Signaling in the hallmarks of Cancer. Cells 9:1–24. https://doi.org/10.3390/cells9071612
Burnstock G, Di Virgilio F (2013) Purinergic signalling and cancer. Purinergic Signal 9:491–540. https://doi.org/10.1007/s11302-013-9372-5
Stella J, Bavaresco L, Braganhol E et al (2010) Differential ectonucleotidase expression in human Bladder cancer cell lines. Urologic Oncology: Seminars and Original Investigations 28:260–267. https://doi.org/10.1016/j.urolonc.2009.01.035
Rockenbach L, Braganhol E, Dietrich F et al (2014) NTPDase3 and ecto-5’-nucleotidase/CD73 are differentially expressed during mouse Bladder cancer progression. Purinergic Signal 10:421–430. https://doi.org/10.1007/s11302-014-9405-8
Nunez M, Diaz-Gavilan M, Conejo-Garcia A et al (2008) Design, synthesis and anticancer activity against the MCF-7 cell line of Benzo-fused 1,4-Dihetero seven- and six-Membered Tethered pyrimidines and purines. Curr Med Chem 15:2614–2631. https://doi.org/10.2174/092986708785909021
Figueiró F, Bernardi A, Frozza RL et al (2013) Resveratrol-loaded lipid-core nanocapsules treatment reduces in vitro and in vivo glioma growth. J Biomed Nanotechnol 9:516–526. https://doi.org/10.1166/jbn.2013.1547
Chan K-M, Delfert D, Junger KD (1986) A direct colorimetric assay for Ca2+-stimulated ATPase activity. Anal Biochem 157:375–380. https://doi.org/10.1016/0003-2697(86)90640-8
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Schmittgen TD, Livak KJ (2008) Analyzing real-time PCR data by the comparative CT method. Nat Protoc 3:1101–1108. https://doi.org/10.1038/nprot.2008.73
Xu J, Gewirtz DA (2022) Is autophagy always a barrier to Cisplatin. Therapy? Biomolecules 12:1–22. https://doi.org/10.3390/biom12030463
Sun C, Mezzadra R, Schumacher TN (2018) Regulation and function of the PD-L1 checkpoint. Immunity 48:434–452. https://doi.org/10.1016/j.immuni.2018.03.014
Narayan VM (2021) Radiotherapy with or without chemotherapy in muscle-invasive Bladder Cancer. 50 studies every urologist should know. 147–152. https://doi.org/10.1093/med/9780190655341.003.0026
Torgovnick A, Schumacher B (2015) DNA repair mechanisms in cancer development and therapy. Front Genet 6:1–15. https://doi.org/10.3389/fgene.2015.00157
Choi W, Porten S, Kim S et al (2014) Identification of distinct basal and luminal subtypes of muscle-invasive Bladder Cancer with different sensitivities to Frontline Chemotherapy. Cancer Cell 25:152–165. https://doi.org/10.1016/j.ccr.2014.01.009
Jiang DM, Gupta S, Kitchlu A et al (2021) Defining cisplatin eligibility in patients with muscle-invasive Bladder cancer. Nat Rev Urol 18:104–114. https://doi.org/10.1038/s41585-020-00404-6
Muller PY, Milton MN (2012) The determination and interpretation of the therapeutic index in drug development. Nat Rev Drug Discov 11:751–761. https://doi.org/10.1038/nrd3801
Awang N, Aziz ZA, Kamaludin NF, Chan KM (2014) Cytotoxicity and mode of cell death induced by triphenyltin (IV) compounds in vitro. Online J Biol Sci 14:84–93. https://doi.org/10.3844/ojbsci.2014.84.93
Badisa RB, Darling-Reed SF, Joseph P et al (2009) Selective cytotoxic activities of two novel synthetic Drugs on human breast carcinoma MCF-7 cells. Anticancer Res 29:2993–2996
Quispe MA, Zavala CD, Rojas CJ et al (2006) Efecto citotóxico selectivo in vitro de muricin H (acetogenina de Annona muricata) en cultivos celulares de cáncer de pulmón. Rev Peru Med Exp Salud Publica 23:265–269
“Pedro Kourí.” Instituto de Medicina Tropical, del Barrio Alonso S, Gutiérrez Gaitén G (1966) Y, Morier Díaz L Revista cubana de medicina tropical. Editorial Ciencias Médicas
Powles T, Perry J, Shamash J et al (2007) A comparison of the platinum analogues in Bladder cancer cell lines. Urol Int 79:67–72. https://doi.org/10.1159/000102917
Rahman NA, Yazan LS, Wibowo A et al (2016) Induction of apoptosis and G2/M arrest by ampelopsin E from Dryobalanops towards triple negative Breast cancer cells, MDA-MB-231. BMC Complement Altern Med 16:1–9. https://doi.org/10.1186/s12906-016-1328-1
Li B, Zhou P, Xu K et al (2020) Metformin induces cell cycle arrest, apoptosis and autophagy through ROS/JNK signaling pathway in human osteosarcoma. Int J Biol Sci 16:74–84. https://doi.org/10.7150/ijbs.33787
Bhosale PB, Abusaliya A, Kim HH et al (2022) Apigetrin promotes TNFα-Induced apoptosis, Necroptosis, G2/M phase cell cycle arrest, and ROS Generation through Inhibition of NF-κB pathway in Hep3B Liver Cancer cells. https://doi.org/10.3390/cells11172734. Cells 11:
Qu K, Lin T, Wei J et al (2013) Cisplatin induces cell cycle arrest and senescence via upregulating P53 and P21 expression in HepG2 cells. Nan Fang Yi Ke Da Xue Xue Bao 33:1253–1259
Wang S, Li W, Xue Z et al (2013) Molecular imaging of p53 signal pathway in Lung cancer cell cycle arrest induced by cisplatin. Mol Carcinog 52:900–907. https://doi.org/10.1002/mc.21930
Magnano S, Hannon Barroeta P, Duffy R et al (2021) Cisplatin induces autophagy-associated apoptosis in human oral squamous cell carcinoma (OSCC) mediated in part through reactive oxygen species. Toxicol Appl Pharmacol 427:115646. https://doi.org/10.1016/j.taap.2021.115646
Li QQ, Lee RX, Liang H et al (2013) Enhancement of cisplatin-induced apoptosis by β-elemene in resistant human Ovarian cancer cells. Med Oncol 30. https://doi.org/10.1007/s12032-012-0424-4
Park H-J, Kim J-S, Lee R, Song H (2022) Cisplatin induces apoptosis in mouse neonatal testes Organ Culture. Int J Mol Sci 23:13360. https://doi.org/10.3390/ijms232113360
Xu X, Lai Y, Hua Z-C (2019) Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci Rep 39. https://doi.org/10.1042/BSR20180992
Li YJ, Lei YH, Yao N et al (2017) Autophagy and multidrug resistance in cancer. Chin J Cancer 36:52. https://doi.org/10.1186/s40880-017-0219-2
Buchser WJ, Laskow TC, Pavlik PJ et al (2012) Cell-mediated autophagy promotes cancer cell survival. Cancer Res 72:2970–2979. https://doi.org/10.1158/0008-5472.CAN-11-3396
Amaravadi R, Kimmelman AC, White E (2016) Recent insights into the function of autophagy in cancer. Genes Dev 30:1913–1930. https://doi.org/10.1101/gad.287524.116
Panda PK, Mukhopadhyay S, Das DN et al (2015) Mechanism of autophagic regulation in carcinogenesis and cancer therapeutics. Semin Cell Dev Biol 39:43–55. https://doi.org/10.1016/j.semcdb.2015.02.013
González-Polo RA, Boya P, Pauleau AL et al (2005) The apoptosis/autophagy paradox: autophagic vacuolization before apoptotic death. J Cell Sci 118:3091–3102. https://doi.org/10.1242/jcs.02447
Trybus W, Król T, Trybus E, Stachurska A (2021) Physcion induces potential Anticancer effects in Cervical Cancer cells. https://doi.org/10.3390/cells10082029. Cells 10:
Shen X, Zhao B (2018) Efficacy of PD-1 or PD-L1 inhibitors and PD-L1 expression status in cancer: Meta-analysis. BMJ (Online) 362:1–9. https://doi.org/10.1136/bmj.k3529
Wu Y, Chen W, Xu ZP, Gu W (2019) PD-L1 distribution and perspective for cancer immunotherapy— blockade, knockdown, or inhibition. Front Immunol 10. https://doi.org/10.3389/fimmu.2019.02022
de Jong FC, Rutten VC, Zuiverloon TCM, Theodorescu D (2021) Improving anti-pd-1/pd-l1 therapy for localized Bladder cancer. Int J Mol Sci 22:1–15. https://doi.org/10.3390/ijms22062800
Soleimani A, Dadjoo P, Avan A et al (2022) Emerging roles of CD133 in the treatment of gastric cancer, a novel stem cell biomarker and beyond. Life Sci 293:120050. https://doi.org/10.1016/j.lfs.2021.120050
Ren F, Sheng WQ, Du X (2013) CD133: a cancer stem cells marker, is used in colorectal cancers. World J Gastroenterol 19:2603–2611. https://doi.org/10.3748/wjg.v19.i17.2603
Zhang JL, Liu Y, Yang H et al (2017) ATP-P2Y2-β-catenin axis promotes cell invasion in Breast cancer cells. Cancer Sci 108:1318–1327. https://doi.org/10.1111/cas.13273
Xia J, Yu X, Tang L et al (2015) P2X7 receptor stimulates breast cancer cell invasion and migration via the AKT pathway. Oncol Rep 34:103–110. https://doi.org/10.3892/or.2015.3979
Takai E, Tsukimoto M, Harada H, Kojima S (2014) Autocrine signaling via release of ATP and activation of P2X7 receptor influences motile activity of human lung cancer cells. Purinergic Signal 10:487–497. https://doi.org/10.1007/s11302-014-9411-x
Schulien I, Hockenjos B, Van Marck V et al (2020) Extracellular ATP and purinergic P2Y2 receptor signaling promote liver tumorigenesis in mice by exacerbating DNA damage. Cancer Res 80:699–708. https://doi.org/10.1158/0008-5472.CAN-19-1909
Acknowledgements
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq/PQ n°302879/2017-0, Battastini AM and CNPq n°406035/2021-0, CNPq/PQ n° 311580/2021-1, Figueiró F). J.S. received support from the Natural Sciences and Engineering Research Council of Canada (NSERC; RGPIN-2023-05498).
Author information
Authors and Affiliations
Contributions
Álisson C. da Silva: Methodology, investigation, writing-original draft preparation. Juliete N. Scholl, Amanda F. Dias and Augusto F. Weber: Investigation, data curation. Fernanda B. Morrone, Olga Cruz-López, Ana Conejo-García, and Jean Sévigny: Investigation. Joaquín María Campos: Investigation and writing-reviewing. Fabrício Figueiró and Ana M. O. Battastini: Supervision, writing-reviewing and editing, funding acquisition.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
da Silva, Á.C., Scholl, J.N., de Fraga Dias, A. et al. Preclinical evaluation of bozepinib in bladder cancer cell lines: modulation of the NPP1 enzyme . Purinergic Signalling (2023). https://doi.org/10.1007/s11302-023-09975-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11302-023-09975-6