
Abstract
Environmental and energy security has now become a serious global problem, requiring a lot of research to find and implement its cost-effective and environmentally friendly alternatives. The development and use of renewable energy sources is necessary and important in order to avoid the emergence of a global economic crisis. One of the solution to prevent a future crisis caused by energy shortages is to introduce biofuels into the fuel market. Despite the fact that various forms of renewable energy are currently used, the prospects for the production of biofuels from cyanobacteria are quite high due to their unique properties, such as a high lipid content and a suitable fatty acid (FA) composition for the production of biofuels, their suitability for growing open water and the ability to grow on wastewater. The purpose of this article is to provide a comprehensive overview of the potential of cyanobacteria in the conversion of wastewater into biofuels. The article covers comparative data on the accumulation of lipids and the content of fatty acids in various representatives of cyanobacteria and their possibilities in the remediation of wastewater. Various approaches to the extraction of lipids from phototrophic microorganisms that are currently available, their advantages and disadvantages, and the results of the monitoring of the main key points of the development of the technology for converting cyanobacterial biomass into biofuels, with an emphasis on the existing barriers, effects and solutions, are also considered. Further research in this field is required for the successful implementation of this technology on an industrial scale.
Graphic abstract

Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
At present, world oil consumption is about 11.6 million tons per day, in connection with this, underground resources of fossil oil will only last for several decades (Kumar 2017). Many researchers assume that a decrease in oil production will be observed starting from the 2030s, which will entail a crisis caused by energy shortages (Kumar and Rao 2015). Therefore, the search is currently underway for an economically viable, inexhaustible source of energy that can replace crude oil, in order to avoid the emergence of a global economic crisis. In addition, burning fossil fuels leads to large emissions of greenhouse gases (GHGs), which in turn leads to irreversible climate change. According to forecasts of climatologists, greenhouse gases will grow to about 43 billion metric tons by 2040 year. Thus, additional energy resources are needed that are renewable and readily available. In this regard, exploring the possibilities of replacing fossil fuel resources with alternative energy is an important field of scientific research. Among other renewable energy sources, biofuels can be considered a substitute for oil due to their non-toxic, S-free, biodegradable nature, originating from renewable sources (Liew et al. 2014; Ghasemi et al. 2012; Sukenik et al. 1993; Torres et al. 2013; Chen et al. 2013; Sharma et al. 2012; Sajjadi et al. 2016).
Nevertheless, biofuel obtained from the biomass of phototrophic microorganisms is a relatively new area of research. Over the past 30 years, cyanobacteria have been intensively studied in biofuel production, especially the ability of their cells to produce lipids and rapid biomass accumulation as a result of photosynthesis (Khan et al. 2018). It is to be noted that the lipids of cyanobacteria are rather diverse in comparison with lipids of algae and plants, since they consist of diacylglycerols located in thylakoid and photosynthetic membranes. Considering the possibilities of photosynthesis on the planet, their lipids, being the most abundant, attract wide attention of researchers and have great potential in the production of biofuels.
In addition, it is important to note about the high photosynthetic activity of cyanobacteria, due to which their mass cultivation can effectively reduce carbon dioxide emissions into the atmosphere and, thus, reduce the impact of global warming, making an important contribution to the protection of the natural environment and maintaining the ecological balance. Also, the ability of cyanobacteria to grow on waste waters of various origins and carry out their bioremediation makes it possible to consider the possibility of using these effluents as a medium for their cultivation, reducing the pressure on fresh water resources in the future.
Thus, the multipurpose use of cyanobacteria in the treatment of wastewater, in the purification of air from CO2 can be integrated by the associated production of their biomass in order to produce environmentally friendly, renewable biofuel (Maheshwari et al. 2018). The production of such biofuels from the biomass of phototrophic microorganisms is not only cost effective, but also environmentally sustainable technology with minimization of waste generation during use (Sheng et al. 2011).
Considering the above, it is necessary to note the high relevance of studies of the capabilities of cyanobacteria in converting waste (wastewater, CO2) into biofuel. In general, there are many different reviews and experimental articles in the literature on the production of biofuels based on phototrophic microorganisms, where the attention of researchers is paid more to microalgae (Abdelhamid et al. 2019; Kalavathi et al. 2001; Shashirekha et al. 2008; Uma et al. 2009; Balaji et al. 2014; Dellamatrice et al. 2017; Kumar et al. 2018), the study of their lipid accumulation, their fatty acid composition (Kumar et al. 2018; Singh 2015a, 2015b; Aratboni et al. 2019).
Besides, the rapid development of science in the field of bioenergy requires an analysis of the current state of scientific data in this area and their systematization.
This review analyzes the capabilities of cyanobacteria in the production of biodiesel, presents comparative data on the accumulation of lipids and the content of fatty acids in their various representatives. And since cyanobacteria have the ability to decompose various environmental pollutants, and are also promising tools for wastewater treatment, in addition to studying the possibility of obtaining biodiesel from cyanobacterial biomass, this review discusses the potential of such cyanobacteria in low- and waste-free technologies for bioremediation of contaminated water with associated production their useful biomass. Furthermore, in view of the fact that the production of high-quality products, including biofuels from the biomass of cyanobacteria, is also determined by the use of effective methods for biomass pretreatment and lipid extraction, this article analyzes the influence of various approaches on the total lipid yield and the possibility of their use when establishing this technology. In addition, the article monitors the key points in the development of technology for the conversion of cyanobacterial biomass into biofuel, considers the main barriers, effects and solutions.
Potential of cyanobacteria in bioenergy and wastewater treatment
A key role in the productivity of any biological product, including biofuels, undoubtedly plays the right choice of a suitable object. As a rule, microorganisms of interest as raw materials for biofuels are selected on the basis of a number of properties, which include the natural ability to produce the fuel of interest or the main raw material for its production, high productivity, active growth, the ability to grow on cheap media, the ability to succumb to various metabolic and genetic manipulations to improve their properties. All this determines the possibility of their industrial application.
Thus, the ability to naturally intensively accumulate the necessary fatty acids and use sunlight as a a source of energy makes cyanobacteria, along with algae, promising objects for the production of biodiesel.
As an environmentally friendly and high-performance raw material for the production of biodiesel, cyanobacteria are recognized as one of the most potentially suitable objects used for the production of third-generation biofuels. When compared with some plant objects that are first-generation fuel sources, phototrophic microorganisms are able to generate almost 30 times more energy per acre. And given their main advantage over plants, such as the lack of competition as a food source, they can be one of the potential substrates for bioenergy production today. The use of food crops as feedstocks for biofuel production is already unprofitable due to the use of agricultural land for their cultivation and competition in the food industry. Therefore, cyanobacteria, as the most ancient, fast-growing and high-energy crops, can become the basis for large-scale production of biofuels, thereby creating the basis for sustainable energy development of the future. Modern catalytic methods open up wide opportunities for processing cyanobacteria biomass into various products. Thus, various types of biofuels can be obtained from their biomass: liquid (biodiesel, biobutanol, bioethanol, bio-oil) and gaseous (methane, biohydrogen). In recent years, the attention of researchers has been increasingly focused on the use of the biomass of cyanobacteria, which actively accumulate lipids in the production of biodiesel.
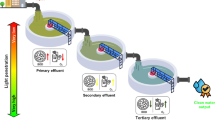
As pointed out before, the waste of cyanobacterial biomass after the extraction of lipids for the production of biodiesel could be used to produce other types of biofuels. For example, carbohydrate-rich biomass waste can be a good raw material for the fermentation and production of alcohol-containing fuels, such as bioethanol and biobutanol (Fig. 1). This also highlights the great potential of cyanobacteria and the feasibility of using their biomass in bioenergy, taking into account the global trends in the development of various low-and non-waste technologies, which are currently one of the important trends.

Scheme of obtaining biofuels from cyanobacteria cultivated in wastewater
Lipid accumulation and fatty acid profile of cyanobacteria
Natural lipid profile of cyanobacteria and influence on fuel quality
Cyanobacteria, being sources of lipids, are considered as important objects in biotechnology and bioenergy. Their lipid components mainly consist of neutral galactolipids (monohalactosyldiacylglycerol and digalactosyldiacylglycerol) and two types of anionic lipids (sulfoquinovosyldiacylglycerol and phosphatidylglycerol). Within a certain class of lipids, cyanobacteria lipids may differ in the number of carbon atoms in the fat acyl substituent and the level of saturation (Hewelt-Belka et al. 2020). Currently, the species Synechocystis, Anabaena, Synechococcus, Oscillatoria and Spirulina are well studied with respect to lipid accumulation (Rajeshwari and Rajashekhar 2011). They have the ability to accumulate significant amounts of diacylglyceride as a reserve lipid (20 to 50% of the dry mass of the cell) under photo-oxidative stress or other stressful environmental conditions.
Thus, recent studies have shown that the production of lipids by the Synechocystis PCC6803 strain was 12.5% (Maheshwari et al. 2018). The data on a sufficiently large number of lipids in the cells of cyanobacteria Oscillatoria sp. (31.9%), Synechococcus sp. (30.6%), Croococcidiopsis sp. (22.7%), Leptolyngbya sp. (21.15%), Limnothrix sp. (20.73%) are presented (Hossain et al. 2020). They also reported that a 10.8% lipid accumulation for the strain Oscillatoria sp. isolated from a rice field. Two species of filamentous cyanobacteria were isolated, their fatty acid composition was studied, and lipid productivity was determined in Spirulina subsalsa BDU 141201 and Oscillatoria willei BDU 130791 (16.2 and 15.9%), respectively. Palanisamy et al. (2021) established a high lipid accumulation of up to 34.5% for Spirulina phlatensis, which was increased to 35.8% after light optimization. Han et al. (2016) optimized the Anabaena variabilis strain for lipid production, achieving its productivity of up to 46.9%. Yalcin and Hoffmann (2020) cultivated the freshwater cyanobacteria Dolichospermum affine strain under starvation and limitation conditions, using six different concentrations of nitrate and phosphate in order to study fatty acid profile. Lipid content of the strain was 10.67% dry weight. Another related work was hold with Nagappan et al. (2020) research group. They isolated 11 nitrogen fixing cyanobacterial strains from natural sources and identified their fatty acids compositions of FAME. According to obtained results, between investigated strains the high about 16.1% DW (dry weight) lipid yield was extracted from dry weight biomass of Anabaena sp. (g24). As previously mentioned, Bolatkhan et al. (2020) investigated several cyanobacterial strains for lipid production that grown in handmade photobioreactor and the result showed unicellular cyanobacteria Cyanobacterium sp. IPPAS B-1200 contains 16.1% lipid from dry weight biomass. As reported by Blažina et al. (2019) Synechococcus sp. MK568070 strain investigated as a high productive for biodiesel production and lipid content was higher from previously mentioned strains - 21.4% dry weight. Cyanobacterium Limnothrix sp. ASM-1 was found between investigated strains as a high lipid producer (9.12% of dry weight) in Oliveira et al. (2018) work after cultivation in microelement-limited medium. In Cordeiro et al. (2017) research work, the maximum lipid yield was obtained after weekly cultivation of Microcystis protocystis in stress conditions, and the lipid parentage was near to 41.5%, while in same work Microcystis panniformis contains 35.8% lipid of dry weight. Among the N2-fixing strains, the Calothrix crustacea cell was found to aacumulate 5% of lipid, while non-heterocystous filamentous cyanobacteria Limicolaria martensiana contains 5% lipid of dry weight as shown in Nagarkar et al. (2004) work.
Table 1 below shows some data on cyanobacteria strains characterized by intensive lipid accumulation.
Biodiesel is composed of fatty acid methyl ester (FAME), obtained by transesterification of biologically derived lipids. Thus, the lipid composition has a significant impact on the biodiesel production technology and product quality.
An important point when choosing the types of cyanobacteria for the production of biodiesel is not only high lipid productivity, but also the correct profile of fatty acids. An important parameter that determines the quality of biodiesel is the composition of fatty acid esters, the structure of which affects the key properties of biofuels, especially its cetane number, ignition temperature, viscosity, oxidative stability, fluidity at low temperatures (Deshmukh et al. 2019). The high content of esters of saturated fatty acids leads to an increase in the cetane number, while the delay time of the fuel combustion is shortened, that is, the fuel mixture will burn more calmly and smoothly. Thereby, a higher resistance to oxidation characterizes biofuels containing compounds with a large number of saturated bonds, and glycerol polymerization does not occur during its combustion, which significantly increases the reliability of engines (Gopinath et al. 2015). Therefore, the fatty acid composition of cyanobacteria lipids, as potential producers of biodiesel, is subject to such requirements as a high content of saturated fatty acids and a low amount of polyunsaturated fatty acids. Among the fatty acids, the chemical compositions C10: 0, C16: 0, C18: 1 and C18: 2 are important for the production of biodiesel.
The study of the carbon chain length of fatty acids provides a mechanism for the production of biofuels with different properties, since C10-14 fatty acids are used as components of reactive biofuels. In addition, C16-18 fatty acids are the main objects of biodiesel production. Studies conducted so far have shown that cyanobacteria contain a large amount of C10-12 fatty acids (Trichodesmium grown in the laboratory accounted for 50% of the total amount of fatty acids). In addition, C14 fatty acid has been found in many cyanobacteria strains—14: 0 Trichodesmium erythraeum (Lu 2010; Chi et al. 2008) or Phormidium J and Prochlorothrix hollandica in some cases contains 30% of cyanobacteria 14: 1 fatty acids (Sarsekeyeva et al. 2014).
All cyanobacteria are conditionally united into four groups according to the fatty acid profile. Group 1 includes cyanobacteria species, in particular representatives of mesophilic and thermophilic Synechococcus species, characterized by only one double bond at the position Δ9 of fatty acids in the SN-l region (C16 or C18) (Sarsekeyeva et al. 2014); in cyanobacteria of group 2, fatty acid C18 is saturated at positions Δ9, Δ12 and Δ15, and acid C16 contains saturated fatty acids at positions Δ9 and Δ12, which is typical for Anabaena, Nostoc, etc. C18 fatty acid in Group 3 cyanobacteria contains saturated fatty acids at positions Δ6, Δ9, and Δ12, this is typical for Spirulina and Arthrospira species (Maslova et al. 2004). Four group of cyanobacteria is characterized by saturation of positions Δ6, Δ9, Δ12 and Δ15 of the C18 fatty acid (model strain Synechocystis sp. PCC 6803). Palmitic acid C16 is not saturated in groups 3 and 4 and allow the whole genomes of cyanobacteria strains to be detected during sequencing. In addition, the genes of the main fatty acid desaturases directly control the quality of fatty acids, the study of these genes opens up opportunities for the classification and differentiation of cyanobacteria based on fatty acids. As already studied, in most cyanobacteria, the fatty acids C18 and C16 are saturated at positions—Δ9 and Δ12 (Table 2). These are mainly representatives of the species Synechococcus and Prochlorococcus (Chi et al. 2008) and they are all adapted to the production of diene fatty acids (Sarsekeyeva et al. 2014).
Temina et al. (2007) investigated the content of saturated and unsaturated fatty acids of Nostoc and other species by growing them in various wastewater and recording the release of lipids. It was shown that the occurrence of fatty acids in the Nostoc family is different. A statistical analysis of the variability of the fatty acids of the Nostoc family was carried out. Behnam et al. (2020) performed the direct transesterification process of the Fremyella diplosiphon strain and identified the fatty acid methyl esters (FAME). Alkanes, which are the main component of conventional fuels, were contained in transesterified lipids of the Fremyella diplosiphon strain, which confirmed that the natural synthesis of these hydrocarbons is not inhibited in the production of biodiesel (Behnam et al. 2020).
Sarsekeyeva et al. (2014) made a phylogenetic analysis of the Cyanobacterium sp. IPPAS B-1200 strain isolated from the salt Balkhash lake and studied its morphological and ultrastructural characteristics. The strain was characterized by a temperature-independent Δ9-desaturation system and a relative amount of myristic (14: 0-30%) and myristolic (14: 1δ9-10%) acids. It is also reported that the total content of C14 fatty acids reaches 40%. It is reported that the remaining fatty acids are mainly composed of palmitic (16: 0) and palmitoleic (16: 1δ9) acids (the sum reaches about 60%). This content of fatty acids, combined with a relatively high growth rate, makes this strain a fairly potential producer for the production of biodiesel. Meanwhile, Sheng (2011) studied the lipid composition of the wild species strain Synechocystis PCC 6803. According to the research profile of fatty acids is as follows - C16: 0 (∼ 60%), C16: 1 (∼ 9.5%), C18: 0 (∼ 1.2%), C18: 1 (∼ 2%), C18: 2 (∼ 9.8%), and C18: 3 (∼ 16.5%).
Genetic engineering of cyanobacteria to improve lipid composition
Since most of the research conducted in the twenty-first century was related to genetic engineering, there is not a little information in the literature about improving the productivity of phototrophic microorganisms by genetic manipulation. Thus, Eungrasamee et al. (2020) was able to find the fatty acid genes of the Synechocystis sp. PCC 6803 strain and increase the amount of lipids by modifying them. They studied the absorption of carbon dioxide by the Synechocystis sp. PCC 6803 strain and also performed a metabolic engineering process. Rosgaard et al. (2012) developed biosynthetic genes or regulatory genes for local cyanobacteria alkanes and used engineering methods to obtain strains that produce active lipids. One such strategy is given in Ruffing and Jones (2012) work, where they increased the production and release of free fatty acids in cyanobacteria S. elongatus PCC 7942 by knocking out the acyl-ASP synthetase gene and thioesterase expression. Later, Santos-Merino et al. (2018) also in her work with S. elongatus PCC 7942, studied the content of fatty acids and conducted experiments of the expression of genes encoding enzymes involved in reactions using genetic engineering. They obtained a mutant of the Synechococcus elongatus strain PCC 7942 and made changes in the composition of fatty acids. Some of these modifications are used to improve the production of omega-3 fatty acids. In this paper, the authors presented new data on the pathway of fatty acid synthesis and new strategies that can be used to manipulate cyanobacteria with saturated fatty acid (Radakovits et al. 2010). The genetic modifications of the FFA synthesis pathways in cyanobacteria Synechocystis described in Liu et al. (2011), included the deletion of the acyl-ACP synthetase gene (slr1609), the inclusion and expression of acetyl-CoA carboxylase genes (fabH and fadI), and the expression of modified genes for acyl-ACP thioesterase (tesA, fatB1, or fatB2) that control the FAS chain length. In addition, Ruffing et al. (2013) conducted experiments for cultivation and testing of S. elongatus PCC7942 strains engineered for FFA production. The authors used several type of genetically modified Synechococcus species in order to increase their FAME yields. Using RNA-seq and targeted genetic engineering, they identified several candidate genes, which can improve cell growth parameters during FFA production. These genes include hypothetical proteins (Synpcc7942_1655) ROS-degrading enzymes (Synpcc7942_1214) and a porin protein (Synpcc7942_1464). The authors also concluded that temperature plays an important role in cyanobacterial FFA tolerance. In Synechococcus sp. PCC7002, the physiological effects of FFA production were abated by decreasing the growth temperature from 38 to 30 °C. Eungrasamee et al. (2019) employed a metabolic engineering design with target genes involved in selected processes, including fatty acid synthesis, phospholipid hydrolysis, alkane synthesis, and free fatty acid recirculation in unicellular cyanobacteria Synechocystis sp. PCC 6803. All engineered strains grew slower than WT and had reduced levels of the intracellular pigment chlorophyll a and carotenoids. A higher lipid content was noted in all engineered strains compared to WT cells, with a maximum content and productivity of 34.5% of DW and 41.4 mg/L/day, during the growth phase of 4th day, respectively.
Information is given where heterologous genes encoding glycerol-3-phosphate dehydrogenase (GPD) and diacylglycerol-acyltransferase (DGAT) were engineered in Synechocystis sp. PCC 6803 to increase the synthesis of G3P (glycerol-3-phosphate dehydrogenase). The resulting recombinant Synechocystis strain produced high levels of lipids without significantly reducing cell growth. Compared to the wild-type strain, the lipid content and productivity of the engineered cyanobacteria increased to 31–36%. They found that enhanced G3P synthesis is an important factor in photosynthetic lipid production and that the introduction of heterologous GPD and DGAT genes is an effective strategy for increasing lipid production in Synechocystis sp. PCC 6803 (Wang et al. 2016).
In general, the methods of genetic modification are especially relevant for fast-growing productive strains of cyanobacteria, but with a low content of lipids and are thus aimed at increasing their lipid productivity. Another key point that the developed genetic engineering methods are aimed at is the development of the ability of cyanobacteria to secrete free fatty acids, the precursors of biodiesel, into the extracellular environment, thus simplifying the release of the product at the output (Radakovits et al. 2010). All these possibilities of genetic engineering stimulate the research of cyanobacteria as a potential source of biodiesel raw materials.
Nutrient deficiency for lipid production in cyanobacteria
An important factor affecting the accumulation of lipids by cells and its biochemical composition is the dependence on the conditions of their cultivation, where nutrient deficiency plays a key role. It is known that the performance of biomass can often have an inverse relationship with the overall performance of lipids. High levels of lipids are usually produced under stress, especially when nutrients are restricted, which in turn reduces the growth of producer cells. Thus, Yalcin and Hoffmann (2020) conducted a study of the biochemical composition of lipids of a single-celled, filamentous strain of cyanobacteria Dolichospermum affine, where the total lipid content in the studied form was 10.67% higher compared to the control. 10 different fatty acid profiles were identified in the biomass grown on 6 different modified nutrient media. In all six different cases studied, it was reported that the main saturated fatty acid of the D. affine strain was palmitic acid (C16: 0), while the fatty acid release was 40.61% higher when N and P were removed from the nutrient media. The release of oleic acid (C18: 1) was detected in all cases, but the highest rate was recorded in the control group up to 33.72%. The results of this study showed that under conditions of nitrogen and phosphorus starvation, affecting valuable secondary metabolites, they also positively affected the accumulation of lipids by the D. affine strain.
Similar data were obtained by Cordeiro et al. (2017) who studied the lipid productivity of 4 species of the genus Microcystis. They found that the greatest accumulation of lipids was characteristic of the Microcystis protocystis strain (41.5%) when cultured on a control medium than under conditions of nitrogen enrichment of the nutrient medium. It was noted that the nutrient medium with a low content of nutrients contributed to optimal cell growth and stimulation of lipid accumulation, which can positively affect the cost of the cultivation process by reducing the consumption of nutrients.
In the work of Loura et al. (1987), data on the effect of nitrogen deficiency on the lipid and fatty acid composition of filamentous cyanobacteria Pseudanabaena sp. M2 and Oscillatoria splendida L3. They noted a twofold increase in dry weight under conditions of nitrogen deficiency, which is apparently associated with the production of cellular polyphosphates and glycogen in response to nitrogen deficiency. The total amount of lipids in Osc. splendida under nitrogen deficiency conditions increased from 46 to 50 mg/g dry wt. In addition, in the case of Pseudanabaena sp. the amount of total lipid was the same as the control and was 40 mg/g dry wt, but the fatty acid composition showed a high level of fatty acids with C18: 3 and with C18: 1.
In addition to macronutrients, the deficiency of certain trace elements can also positively affect the accumulation of lipids in cyanobacteria cells. Thus, according to Rathore et al. (1993) Anabaena doliolum, grown under conditions of molybdenum deficiency, produced less biomass, but there was an increase in total lipid by 1.5-fold (from 88 to 122 mg/g dry wt). Molybdenum deficiency caused an increase in the content of digalactosyldiacylglycerol and phosphatidylglycerol lipids in the cells.
Thus, determining the potential of cyanobacteria in the production of biodiesel, it should be noted that they are quite productive, produce biomass in a short time, and are able to accumulate a lipid product 50–300 times more than some other sources (Mondal et al. 2017). However, despite this, they are still not used commercially, since the production and processing of cyanobacteria biomass into biodiesel today remains quite expensive and unprofitable process. The cultivation of cyanobacteria requires considerable expenses for macro- and microelements, certain costs are associated with the separation of biomass from water, extraction and purification of lipids.
But despite the existing problems, the production of biodiesel from cyanobacteria biomass is considered very promising from an environmental and social point of view, this serves as an incentive to find and develop technologies that can reduce its cost. In this context, the use of waste water of various origins as a nutrient medium for their cultivation can be a solution in reducing the cost of the resulting fuel.
Simplification of cyanobacteria cultivation technology using wastewater
Considering the dramatic increase in the world's population and the rising standard of living, it is necessary to note the increase in wastewater, against the background of a shortage of fresh water around the world. So, in the world, under the influence of anthropogenic activities, at least 3 billion tons of waste water are released into the environment annually. waste, only some of which are re-treated. For example, in India alone, the amount of waste thrown out per year is 300 million tons, and over time it only increases. In the capital of India alone, the amount of wastewater discharged into the external environment is about 40 billion liters, and only less than half of it is subject to treatment and recycling. In the vast majority of countries, especially in China, most of the wastewater generated from domestic, municipal, agricultural and industrial sources is discharged into the environment without sufficient complete treatment (Boretti and Rosa 2019).
The ability of cyanobacteria to grow on a medium rich in organic carbon and inorganic nitrogen and phosphorus allows us to consider the possibility of using industrial and domestic wastewater as a nutrient medium in order to reduce the cost of their cultivation technology. Considering the potential of these facilities in the treatment of polluted wastewater, it is necessary to note the possible great prospects of low-and non-waste technologies for bioremediation of polluted water with the associated production of their useful biomass. They can be grown in special natural and artificial ponds for the treatment of industrial and domestic wastewater and used as a part of a treatment system consisting of several stages. Thus, cyanobacteria are characterized by the ability to oxidize petroleum components, complex organic compounds, and accumulate metal ions, such as Zn, Co, and Cu (Gupta et al. 2000). Their great potential in the purification of various metals is due to the content in their cells of various functional groups, such as carboxyl, hydroxyl, sulfate and other charged groups important for the binding of metals. Thus, information is provided on the biosorption of zinc and nickel by Spirulina indica, Spirulina maxima, and Spirulina platensis strains (Dmytryk et al. 2014). In addition, cyanobacteria are known to have great potential in removing nutrients from wastewater rich in nitrogenous and phosphoric compounds. The ability to remove nitrogen and phosphate ions from wastewater has been observed in cyanobacteria such as Oscillatoria, Phormidium, Aphanocapsa and Westiellopsis (Lau et al. 2015).
Cyanobacteria have also been reported to accumulate and detoxify various pesticides. Previous studies have shown that Synechocystis sp. PUPCC 64, Westiellopsis prolifica, Nostoc hatei and Anabaena sphaerica are capable of decomposing organophosphorus or organochlorine insecticides in the aquatic environment (Debnath et al. 2012).
In the natural environment, many cyanobacteria form symbiotic associations with other aerobic or anaerobic microorganisms. Cyanobacterial mats, including members of the genera Oscillatoria, Synechocystis, and Pleurocapsa have been shown to promote the degradation of hydrocarbons present in oil. Despite the fact, cyanobacteria are not directly responsible for the degradation of hydrocarbons, they facilitate the degradation process by providing oxygen and nutrients to the associated oil-destructive bacteria (Ichor et al. 2016). A consortium consisting of Phormidium, Oscillatoria, Chroococcus, and an oil-decomposing bacterium Burkholderia cepacia has been successfully developed and used to efficiently decompose petroleum compounds.
The ability of cyanobacteria to grow on polluted wastewater allows them to be used in low-and non-waste technologies that provide for simultaneous water purification with parallel production of crop biomass, which have a great potential in the production of biodiesel. According to the global trends in the development of biotechnology, such low-and no-waste strategies are currently one of the important trends, since the absence of waste reduces the environmental burden on the environment and increases the profitability of any technology.
Wastewater is chemically/physically different due to its release from different environments (urban, industrial and agriculture streams). Depending on the concentration of macro-and micronutrients, biologically active compounds and toxins found in them, the growth rates of cyanobacteria grown in them also vary. Depending on the substances found in wastewater, they are divided into three groups: municipal, agricultural wastewater, industrial wastewater.
Municipal wastewater
Municipal wastewater is mainly generated from domestic wastewater and small businesses, as well as from some wastewater such as melted snow or storm water (Becker 1994). Municipal wastewater is the preferred wastewater source for algae cultivation due to its high volume and content of phosphorus and nitrogen. However, municipal wastewater contains significant amounts of heavy metals (zinc, lead and copper) which can interfere with cyanobacterial cultivation (Cai et al. 2013).
Agricultural wastewater
Agricultural wastewater, frequently derived from agricultural operations and livestock production (e.g. beef cattle, dairy cattle, pigs and poultry), manured wastes, and it is another source of wastewater that contains very high levels of nitrogen and phosphorus, making their suitable substrate for cyanobacterial cultivation and sustainable production of cyanobacterial biodiesel (Abou-Shanab et al. 2013).
Industrial wastewater
Industrial wastewater is one of the most serious sources of water pollution. Significant quantities of polluted water have been discharged into lakes, coastal areas and rivers over the 1–2 decades, leading to serious problems for both human health and ecosystems (Edokpayi et al. 2017). The composition of this wastewater varies greatly from industry to industry, depending on end products. Each sector has its own specific combination of pollutants, which may require specific treatment processes (Cai et al. 2013). Although, wastewater is source dependent to some extent, most industrial wastewater contains less phosphorus and nitrogen but more heavy metals (Fe, Cu, Zn and W) than agricultural wastewater and municipal waswater.
In research works, wastewater is used to grow cyanobacteria by partial or direct replacement of nutrient media. Bolatkhan et al. (2020) conducted research work in the application of municipal wastewater for the cultivation of cyanobacteria strains, enabling the development of bioenergy industry and environmentally friendly environmental remediation technologies. They studied the effect of different concentrations of carbon dioxide and municipal wastewater on the growth of cyanobacteria and determined their capabilities in wastewater treatment processes. It was found that the optimal concentration of CO2 for growing Cyanobacterium sp. IPPAS B-1200 and Desertifilum sp. IPPAS B-1220 is 10%. It is shown that the cultivation of Cyanobacterium sp. IPPASB-1200 on sewage wastewater reduces the concentration of organic pollutants and, accordingly, improves the physical and chemical properties of water. The percentage of cleaning of the selected pollutants was 68–100%. The resulting cyanobacteria biomass was characterized by a significantly high content of lipids (22%). The qualitative and quantitative composition of the fatty acids of the studied strains was the most suitable for the production of biodiesel. Of course, the development of such technologies will allow the development of waste-free technologies, the processing of liquid and air waste into energy resources, which will save raw materials and energy, in turn, reduce waste generation.
In the Yalcin and Hoffmann (2020) study, a single-celled, filamentous strain of Dolichospermum affini was grown on a BG-11 medium modified with municipal wastewater containing six different concentrations of nitrate (NaNO3) and phosphate (K2HPO4). The molecular characteristics of D. affin strains were determined and the lipid yield rate was 18.10%. Cells grown in wastewater had a higher lipid accumulation activity compared to other media. A study by Soydemir et al. (2016) examined mixed cultures of cyanobacteria grown in treated municipal wastewater. The total lipid yield of the mixed cultures was 26.2% by dry biomass weight. Gas chromatography analysis showed that the FAME composition mainly consists of methyl esters of palmitic, palmitoleic, stearic, oleic, linoleic and linolenic acids. This study suggests that treated domestic wastewater can be a favorable and sustainable environment for cyanobacteria to grow as a biodiesel feedstock.
Blazina et al. (2019) studied the possibility of cyanobacteria Synechococcus sp. MK568070 in the bioremediation of oil refinery wastewater and determined its potential in the production of biomass and lipids. According to their data, despite the existing dependence of biomass productivity on the source of nutrients, their concentration, salinity and the addition of CO2, in general, there is a positive correlation between the increase in biomass yield (767 mg dry wt/l) and the accumulation of lipids (21.4%) against the background of wastewater treatment. The possibility of accumulation of lipids due to functional photosynthesis, even at a minimum concentration of nutrients, in the conditions of waste water and an excess of industrial CO2, undoubtedly determines the prospects of the Synechococcus sp. strain (MK568070) for industrial production of biodiesel and wastewater treatment.
Kamilya et al. (2006) evaluated the potential for removing nitrogen and phosphorus sources from wastewater by cultures of cyanobacteria Spirulina platensis and Nostoc muscorum. Thus, they found that during 7 days of cultivation, the removal of NH4, NO2, NO3, and PO4 by N. muscorum biomass was 83.6, 44.2, 14.17 and 41.79%, respectively, while these indicators for S. platensis were significantly higher and amounted to 92.4, 48.7, 50.39 and 47.76%, respectively. The high growth potential of these crops on wastewater and their potential for purification determine the prospects for their further use as a source of biofuels.
Mostafa (2012) show a significant increase in the productivity of cyanobacteria in wastewater. According to their data, the yield of the biomass of Nostoc humifusum reached the maximum values (572.80 mg dry weight/l), when cultivated on wastewater without any treatment and additives, compared with the control (BG-11 medium) value (400 mg dry weight/l), and for Phormedium fragile these indicators were 156.16 mg dry weight/l in the experiment. An increase in the productivity of cyanobacteria in wastewater was also noted in the work Ashokkumar et al. (2019) work for cyanobacteria of the genus Synechocystis and El-Sheekh et al. (2014) for Nostoc muscorum and Anabaena subcylindrica. The prospects of the cyanobacteria Nostoc linkkia in the purification of wastewater from nitrogen-containing, phosphorus-containing compounds and surfactants with the simultaneous production of lipid-containing biomass were noted in Bolatkhan et al. (2020).
Thus, cyanobacteria have a great potential in the bioremediation of industrial, domestic and agricultural wastewater. In addition, some of their species can grow at high concentrations of carbon dioxide, can convert carbon dioxide into organic compounds and simultaneously have a cleansing effect on the atmosphere and hydrosphere.
As previously mentioned, the combination of water purification processes with the production of microalgae biomass will significantly increase the environmental and economic efficiency of the production of biomass of biodiesel producers. Therefore, the use of wastewater is considered as a promising alternative to the use of synthetic media with a significant reduction in cultivation costs due to their high availability and availability of nutrients necessary for growth (Molazadeh et al. 2019). This is especially important when scaling the biomass production process of cyanobacteria. Therefore, the development of effective approaches to the cultivation of cyanobacteria, along with the selection of highly effective strains and the study of their metabolic capabilities, is an important step towards the development of an economically sound technology for the production of biodiesel from cyanobacteria biomass.
Efficiency of the approaches used in obtaining lipids from the biomass of phototrophic microorganisms
However, in addition to the development of effective cost-effective approaches to the cultivation of cyanobacteria, there is another important point that plays a significant role in the commercial implementation of this technology. This is a methodological support for the technology of separation of cyanobacteria biomass and isolation of lipids from it. In particular, one of the current problems of the widespread use of cyanobacterial biomass for the production of biofuels is the separation of biomass and the optimization of the methodology for the separation of lipids from the liquid. In addition, cyanobacteria cells are surrounded by cell walls that prevent the release of internal lipid. In this regard, studies on the destruction of the cell envelope to improve the extraction of lipids, the choice of optimal solvents still requires full-fledged research. The dependence of the lipid yield on the methods used for biomass pretreatment and lipid isolation determines the importance of analyzing the methods currently used. Therefore, the optimization and modification of existing methods and the constant search for new catalysts for processing microalgae biomass into biodiesel can significantly reduce the cost of this process (Aratboni et al. 2019).
The methodological support of the technology of industrial production of biodiesel based on cyanobacteria includes all the methods of the main stages of this process, starting from the concentration of the grown cyanobacteria biomass to the methods of lipid extraction (Fig. 2). It is necessary to note the wide variety of existing methods, to which quite a few review and experimental articles have been devoted to date. In this section of the article, we will not dwell in detail on the description of all known methods for obtaining lipids, but only conduct a brief comparative analysis of some of them, with the definition of advantages and disadvantages and the determination of their effect on the total lipid yield in phototrophic microorganisms.

Methods for separating the biomass of phototrophic microorganisms from the culture fluid
Methods for separating biomass of phototrophic microorganisms from the culture liquid
Thus, the known methods of separating the obtained biomass of phototrophic microorganisms, including cyanobacteria, from the culture medium, such as centrifugation, sedimentation deposition, filtration, flocculation, flotation and ultrasonic processing, have their advantages and disadvantages that affect the cost of this process. The main difficulties of this stage are associated with the small size of cyanobacteria cells and their low number in the suspension, which makes it difficult to concentrate them. In addition, the method used must preserve the viability of the cells in order to prevent changes in the biochemical composition of the cells obtained as a result of cultivation. It is known that separation by means of centrifugation is characterized by high-energy consumption and the filtration method requires frequent replacement of filters, membranes and a long process time (Lal et al. 2018). Sedimentation deposition of biomass, despite its low energy consumption, also requires a long time of its implementation and is additionally characterized by a low degree of cell separation from the supernatant, which certainly limits its industrial use. The method of separating biomass by electro flotation requires frequent replacement of electrodes and high-energy costs (Jeevanandam et al. 2020).
The efficiency of the separation of microbial biomass by electrocoagulation was successfully demonstrated in the studies of Uduman et al. (2011), which studied the electrocoagulation of microalgae using various types of electrodes. They determined the high efficiency of the use of aluminum, apparently associated with its charge, which provides a greater probability of the formation of agglomerates due to the neutralization of the surface charge of the cells.
Of all the known methods, the most widely used is the method of concentrating biomass-using flocculants. Flocculation of microorganisms, including cyanobacteria, occurs due to the neutralization of the charge and a decrease in the electrostatic repulsive forces between the charged cells in the suspension, so their individual cells aggregate to form larger particles that precipitate faster. Despite the fact that this method is characterized by high efficiency and low cost, the dependence of the effectiveness of salts of various metals (Zn, Cu, Al, Fe, etc.) used as flocculants on external factors requires additional research and possible modifications of this method (Vandamme et al. 2013).
The use of chemical coagulants for the concentration of biomass from suspension, including titanium tetrachloride and manganese sulfate (Politaeva et al. 2018), and slaked lime, has been widely studied. However, the disadvantages of using chemical coagulants are associated with the transition of part of the cell contents of phototrophic microorganisms into the culture liquid (Bonnet et al. 2019).
Thus, summing up the methods for separating the biomass of phototrophic microorganisms, including cyanobacteria, from the culture liquid, we would like to note the complexity, energy intensity and duration of such methods as centrifugation, separation, filtration and reducing the quality of the resulting products when using chemical precipitation methods. Therefore, the search for an effective way to separate the biomass from the culture suspension is still an urgent problem (Branyikova et al. 2018).
Methods of cell disintegration of phototrophic microorganisms and lipid extraction
Analyzing the methodology of the next stage of this technology, it would be better to note the equally important methods used in the destruction of cyanobacteria cells. As is known, the procedure of lipid extraction itself involves the destruction of cells and the actual extraction of lipids using various solvents (Ren et al. 2017). Unfortunately, there are only limited works in the literature devoted to the study of the total lipid yield. In addition, the complexity of comparative study is associated with the use of experimenters of combinations of different methods of cell destruction with different methods of extraction, as well as the use of solvents of different nature. But despite all this, the correct choice of the method of cell destruction undoubtedly determines the success of the final result. Despite the more simplified structure of the cell wall of cyanobacteria compared to microalgaeand, the overall lipid productivity of the strain directly depends on the correctly selected method of disintegration of their cells. Of course, preference is given to methods characterized by low energy consumption and high productivity, which exclude destruction or changes in the target product.
Cell destruction methods can be physical, chemical, or biological. Physical methods—shaking, ultrasound, autoclaving, microwave oven and others. These methods cause mechanical failure due to contact, pressure or denaturation. One of the advantages of these methods is that they are efficient and do not generate waste, but the disadvantage is that they require energy. On the other hand, chemical methods are characterized by the use of acidic or alkaline chemicals to destroy cells. The advantage of this process is that it simplifies two processes, since cell destruction and hydrolysis of polysaccharides can be achieved using chemical reagents (mainly sulfuric acid). The biological method, in turn, is characterized by the use of enzymes both for the destruction of cells and for the hydrolysis of polysaccharides. The advantage of enzymes is that they do not harm the environment. However, today in the world the method of mechanical/physical destruction of cell membranes is widely used.
Mechanical methods of destruction of the cell envelope, carried out mainly with the use of solid (for example, high-speed homogenization) and liquid mechanical forces (high-pressure homogenization, micro-fluidization), require the transfer of energy to the cells, either on the basis of waves (laser, ultrasonic and microwave processing), or conventional heat (autoclave and water bath) or using a pulsed electric field.
Among physical methods, one of the most frequently used methods of destroying the cells of phototrophic microorganisms is Expeller Pressing, which does not require expensive equipment and involves the use of simple chemicals. Basically, the production of lipids using an expeller is used for filamentous cyanobacteria, and currently up to 75% of the lipids produced by phototrophic microorganisms are obtained by destroying cells by this method. Depending on the type of biomass, different configurations of expellers can be used: screw, piston, etc. (Topare et al. 2011). However, this method is relatively slow and requires a lot of cyanobacterial biomass. In the studies of Milledge and Heaven (2014) this method of mechanical processing of cells is shown as a method that requires a large amount of biomass.
One of the most effective methods most often used in the work on the crushing of microbial biomass is Bead Milling. This method has many advantages, such as high cell destruction, easy biomass loading, optimal operating temperature, and application for various types of biomass from laboratory to industrial scale. The successful destruction of cells is ensured by the correct position and speed of the mixer, the optimal rotation of the suspension flow, regardless of the biomass concentration. Cheng et al. (2011) in his studies showed high lipid productivity of the Bead Milling method modified using a mixture of SC-CO2 for cyanobacteria Pavlova sp cells. Cyanobacteria cells were dried using a 3 mm diameter spray gun, and the extraction process through glass beads was carried out under high pressure. The Bead Milling method, performed using a mixture of SC-CO2, showed a high lipid yield (15.7%) rather than using N-hexane (10.4%).
The method of pulsed electric field in which an external electric field creates a critical electric potential through the cell membrane increases its porosity and the release of lipids from the cells. In this way, treating Synechocystis PCC 6803 with a pulsed electric field (intensity > 35 kWh/m3) and isopropanol as a solvent significantly increased the total lipid yield (Sheng et al. 2011).
Recently, a widely used method of destroying microbial cells of microalgae and cyanobacteria, bacteria and yeast is High-Pressure Homogenization. High-pressure homogenization is a simple system for breaking down microbial biomass, and it can often be used for blasting wet biomass, where the processing liquid is subjected to pressure in the amplifiers and passes through the homogenization chamber. Samarasinghe et al. (2012) in their research work with microalgae Nannochloris oculata using this method recorded a lipid yield of up to 60%. Kim et al. (2010) in their work showed that the use of high pressure is the simplest and easiest method in cell destruction. The authors used osmotic pressure to destroy microalgae Chlamydomonas reinhardtii, Botryococcus sp., Chlorella vulgaris and Scenedesmus sp. when studying the accumulation of lipids by their cells.
One of the most frequently used effective methods of destruction of cyanobacteria cells is the method of Ultrasonication. When cells are treated with ultrasound, acoustic flow and cavitation are created, affecting the cells in the form of microtubules, creating pressure that leads to the destruction of cell walls and intracellular membranes. Many scientific papers have been devoted to the production of lipids in combination with ultrasound (Kumar and Banerjee 2019). Wu et al. (2011) used low-frequency (20 kHz) high-intensity (0.0403 W/cm3) ultrasound treatment to destroy Microcystis aeruginosa cells and found that ultrasonic water degradation generates free radicals that weaken the cyanobacteria cell wall. Studies by Keris-Sen et al. (2014) showed that disruption of the cell structure by ultrasound significantly increases lipid productivity, while significantly reducing the processing time of cells. In Kaiwan-arporn et al. (2012), a lipid extraction procedure involving ultrasound treatment and chloroform: methanol extraction resulted in up to 18.58% lipid yield in cyanobacteria Synechocystis aquatilis TISTR8612. Despite the high efficiency of this method, there are methods that can further increase the total lipid yield. So Qv et al. (2014) found that when extracting lipids of the microalgae Dunaliella tertiolecta using ultrasound and microwave rays, the extraction rate during ultrasound treatment was lower and amounted to 45.94% with an extraction time of 5 min than when exposed to Microwave radiation, where this indicator was equal to 57% with a reduction in the exposure time to 160 s. The results obtained by them indicate a much greater efficiency of microwave rays than ultrasound. The Microwave Irradiation method is a fairly widely used method for extracting lipids from microorganisms by the action of electromagnetic waves on cell suspensions in an organic solvent. A comparison of lipid extraction in Nannochloropsis sp. using the method of cell destruction by microwave irradiation and heating in a water bath is also presented, the results obtained were also in favor of microwave irradiation - 38.31% and 23.01% (Wahidin et al. 2013).
Of course, a combination of several methods simultaneously in order to optimize the conditions for their implementation (reducing processing time, power and, accordingly, energy consumption, increasing the extraction rate) can be successfully used and contribute to an increase in overall lipid productivity (Qv et al. 2014).
Along with mechanical methods of cell disintegration, which have a number of disadvantages, such as insufficient extraction performance, the use of harmful solvents and the duration of the procedure, non-mechanical methods of destruction, such as chemical and enzymatic cell lysis, have great prospects. If the use of chemical reagents (alkalis, acids, salts) involves their effect on the cell wall, causing denaturation and coagulation of proteins, oxidation of cell wall components, accompanied by the effect of reagents on other cell structures, then the use of various enzymes - xylanase, cellulase, amylase, papain, pectinase and hemicellulase ensures the destruction of bonds between the substances of the cell wall without destroying the target product. Han et al. (2016) used this method for the Scenedesmus sp.strain to obtain a 96.4% lipid yield. Maffei et al. (2018) when working with the marine microalgae Nannochloropsis sp., treating the strain with cellulase and mannanase, improved the yield of lipid extraction from 40.8 to 73%.
Data on the use of hydrophilic ionic liquid, 1-butyl-3-methylimidazolium for the lysis of microalgae cell walls in the isolation of cellular lipids are presented (Salvo et al. 2010). The hydrophilic ionic liquid breaks down the cell walls and forms two immiscible phases, one of which consists of lipids. After mixing the hydrophilic ionic liquid with a suspension of microalgae cells, the hydrophobic phase of lipids, due to the action of gravity, is placed on top, where it is removed from the mixture and purified.
Despite the variety of existing methods of cell disintegration, it should be noted that cyanobacteria, unlike microalgae, are easily subjected to genetic manipulation, which in turn makes it possible to carry out genetic engineering work with them in order to simplify some stages of this technology. Genetically modified strains, which are of great interest to researchers as a resource for obtaining fourth-generation fuel, allow them to avoid the expensive stages of biomass collection and cell disintegration. The ability of such genetically engineered strains of cyanobacteria to release the end products of metabolism including free fatty acids into the nutrient medium undoubtedly greatly simplifies the technology of processing biomass in the production of biodiesel (Ashokkumar et al. 2019). Thus, it is reported that the removal of the sll1951 gene for the surface protein in the genetically modified Synechocystis strain significantly weakened the peptidoglycan layer of the cell wall and thus contributes to increased FFA secretion into the culture medium (Liu et al. 2011). Information is given that such free fatty acids are deposited in the form of a ring on the walls of the flasks and accumulate in the nutrient medium above the aqueous phase (Haedrich et al. 2020). Of course, this ability of cyanobacteria strains to secrete hydrophobic fatty acids into the nutrient medium greatly simplifies the process of their separation and, most importantly, eliminates the stages of periodic biomass build-up and cell destruction.
Thus, since the stage of dehydration of biomass with subsequent drying is a rather energy-intensive process, due to which the energy contribution during the production of lipids exceeds the energy output from the resulting lipid, the development of methods for extracting lipids without these auxiliary operations is very relevant today. In addition to the effect on cells by genetically engineered methods, avoiding the stage of dehydration, drying and disintegration of cells can be achieved by using special solvents. Thus, information is provided on the use of dimethyl ether (DME), which has a high affinity for both water and organic compounds and is able to penetrate through cell walls. It was possible to extract 26.4% using DME of the total amount of crude lipids and 54.4% of the total amount of fatty acid methyl esters (Wang et al. 2021). Of course, such approaches are more economical, since they allow simultaneous dehydration with the extraction of lipids and do not require cell destruction.
In general, the lipid extraction stage is a crucial step in the process and represents an important bottleneck for the production of biodiesel on an industrial scale.
Despite the success in the cultivation of cyanobacteria, obtaining their active strains, which are of great interest in the field of bioenergy, there are still no commercially successful industrial methods for extracting lipids from their biomass. The currently existing set of methods for extracting lipids from the biomass of phototrophic microorganisms are represented by varieties of such basic methods as: Soxhlet method, Superior Solvent Extraction Methods, Bligh and Dyer method and Folch method, which differ in the use of various solvents or a combination of diverse solvents and their different ratios. Traditional extraction methods have a number of limitations for large-scale biodiesel production, such as the toxicity of the solvent used in the extraction, the specificity of the solvents used, the environmental pollution of the spent solvents, and the high-energy intensity of the solvent-lipid separation process, which involves high costs for their subsequent purification and distillation (Ren et al. 2017). The main problem in the extraction of lipids from biomass is the presence of residual water, which prevents complete phase separation and reduces the efficiency of the process. In this regard, technologies that ensure the extraction of lipids and do not require pre-drying of biomass are of great importance (Khan et al. 2018). To eliminate these shortcomings, researchers using many modifications are constantly looking for improved methods. Known methods of in situ lipid hydrolysis and supercritical in situ transesterification (SC-ST/E) for the extraction of lipids from wet algae biomass. In this method, wet algae biomass is processed to extract lipids, followed by transesterification to produce biodiesel.
Thus, there are many ways to destroy microbial cells and isolate lipids from them. Each method is effective and specific, and there are still modification processes going on. However, according to the review, the most commonly used and more productive methods for destroying cell structure are Microwave Irradiation and Ultrasonication. By the method of Microwave irradiation from the type Trichosporon sp. during production, the amount of lipids was 43.1% (Kumar and Banerjee 2019), while 57.1% of lipids were obtained by the method of Microwave Irradiation (Qv et al. 2014).
Regarding the methods of the actual extraction of lipids, the existing methods are based on the Folch method, which uses organic solvents, the use of which causes the separation and settling of lipids in the upper phase of the solution. The Bligh and Dyer method, a gravimetric method characterized by the ratio of organic solvents, is still widely used for the evaluation of lipids by algae technologists, and is used for both pilot and large-scale extraction processes. There are quite a few modifications of this method aimed at increasing productivity and reducing its toxicity. The most common modification is the addition of 1 M NaCl instead of water, 0.2 M phosphoric acid, HCl, acetic acid, and methyl tert-butyl ether (MTBE) (Matyash et al. 2008). However, all these listed modifications of the method involve the use of chloroform, which, despite the fact that it provides effective extraction, is still not suitable for large-scale use due to toxicity. Therefore, less toxic but less effective substitutes such as ethanol, isopropanol, butanol, MTBE, acetic acid esters, hexane, and various solvent combinations have been and are being investigated by many researchers for the extraction of microalgae lipids (Sheng et al. 2011). Thus, the above-mentioned methods should be tested for its commercial applicability and further research in this area is required for the successful implementation of this technology on an industrial scale.
Prospects and problems of production of biodiesel based on cyanobacteria
Summing up the results of the previous sections of our article, we note that cyanobacteria have the ability to produce a large number of various necessary products, including lipids. And along with microalgae, they can be considered as promising objects for the production of biodiesel due to their correct fatty acid composition and the ability to be easily subjected to various genetic manipulations.
However, our monitoring of the main key points in the development of the technology for converting cyanobacterial biomass into biofuels indicates that, despite the prospects for their use as a renewable source of biomass, at present, the industrial production of such biofuels is not economically justified and has significant barriers (Fig. 3). In addition to the problem of increasing the production of lipids by individual strains of cyanobacteria, an important problem is the scaling of the process of their cultivation. The cultivation of cyanobacteria is one of the most expensive parts of any production process, regardless of whether it is in the conditions of photobioreactors or open ponds.

Monitoring of key points in the development of the technology of conversion of cyanobacterial biomass into biofuels
Therefore, despite the great energy potential of cyanobacteria, today the production of biodiesel based on them is limited by the high costs of its production due to the high cost of the resources used, including water, components of the nutrient medium and electricity, etc. Given that the economic efficiency of producing biofuels from phototrophic microorganisms depends on their large-scale productivity, the greatest obstacle to the commercialization of cyanobacteria biofuels is the difficulty of repeating the productive potential for large-scale production. Therefore, despite their potential for high-speed biomass production relative to land-based raw materials, carrying out this process on a large scale may require significant effort.
A number of researchers conducted a technical and economic analysis of the feasibility of different ways of producing biofuels based on phototrophic microorganisms indicate a large variability in the estimated cost of fuel. Thus, Benemann and Oswald (1996) report a low production cost of $1.65 gal−1, while Richardson et al. (2012) report a high production cost of $33.16 gal−1. The differences between the presented results can be explained by differences in the processing paths and modeling of the current systems. A meta-analysis performed by Sun et al. (2011) to align the modeling efforts of the studies conducted with the published results, which initially ranged from $11.40 to $27.10 gal led to much more closely matched ranges of $11.68–$14.31 galon, illustrating a significant reduction in variability due to the use of similar growth conditions and production trajectory. Significant differences in cost are also shown between competing cultivation types, open air and closed photobioreactors. Studies comparing the two costs have concluded that ORP is more cost-effective than PBR. In the work of Pienkos and Darzins (2009), the economic costs of biodiesel for trucks with a capacity of 30 g/m2/d and 15 g/m2/d were studied. The economic cost per 1 t/km of fuel for trucks with biodiesel with a capacity of 30 g/m2/d from cyanobacteria at 100% CO2 (ammonia plant) was $2.8, and for biodiesel at 15% CO2 (flue gases) was $2.2. The cost of biodiesel from rapeseed was $4.2, according to their calculations. They also conducted a technical and economic analysis of this production process in order to estimate the cost of biofuels based on phototrophic microorganisms. They estimated the cost to range from $25 per gallon for low performance (10 g/m2/day at 15% TAG) to $2.50/gallon for high performance (50 g/m2/d at 50% TAG). They determined that only with high productivity forecasts, the price of fuel based on microalgae can be competitive with the price of soy (Comar and Nelson 1975). Obviously, the challenge facing the commercialization of the lipid fuel of phototrophic microorganisms is to achieve high productivity while further reducing capital and operating costs. Thus, according to the results of the analysis of technical and economic calculations, it is assumed that the cost of biofuels from phototrophic microorganisms will be approximately 6–10 times higher than traditional diesel fuel.
Of course, this price comparison is approximate, since it is necessary to take into account the existence of a number of «hidden costs» for fossil fuels that are not reflected in the price of ULS diesel fuel. These include the fact that the initial high capital costs have been largely written off over time, in addition, most governments provide subsidies and tax incentives for oil production, and thus taxpayers have indirect costs that do not directly affect the price of fuel. It should also be noted that the costs incurred by governments and individuals due to the health effects of fossil fuels are also not reflected in fuel prices (Balan 2014).
For example, at lower production rates, the final product may be more expensive than fossil diesel, and potentially even more expensive than biodiesel from rapeseed. Profitability requires a source of carbon dioxide and a high rate of cyanobacteria production. But it should be taken into account that despite the low cost of ULS fuel, with the obvious increase in the price of fossil oil, due to its reduction, technological advances in the production of biodiesel from phototrophic microorganisms, including cyanobacteria, can be quite profitable.
For the successful production of biodiesel by cyanobacteria, additional aspects of improving producers through genetic and metabolic approaches need to be explored. The development of genetic engineering, high-performance technologies and advances in systems biology using genome-wide tools make it possible to create strains of cyanobacteria with a given genotype and the required fatty acid composition.
Another key to the successful commercialization of cyanobacteria biodiesel is the improvement of ways to efficiently process biomass. The improvement of biomass separation methods, the selection of optimal solvents and their ratios are important key points that affect the outcome and success of this technology. In addition, the methods used today involve the use of various alkaline and acidic catalysts, which have such disadvantages as the difficulty of separating them from the final products and high requirements for the quality of the feedstock. Innovative technologies promising in this direction are the use of biocatalysts based on immobilized enzymes, which have a number of advantages over transesterification catalysts. This is a low process temperature, usable and easy to separate from the final products. All this will contribute to a significant reduction in the cost of the technological process of obtaining biodiesel.
Technical and economic assessment of the production of biofuels based on phototrophic microorganisms indicates the need to plan and invest in new low-and non-waste technologies that allow, on the one hand, to make the anthropogenic cycle of substances more closed, on the other hand, to significantly reduce the cost of the proposed technology. Thus, the main focus should be on the integration of various innovative technologies. An example of such integration is the production of cyanobacterial biodiesel when they are cultivated in wastewater. Such low-and non-waste technologies are the basis for the complex conversion of solar energy by phototrophic microorganisms into bioenergy, and subsequently into biofuels. The so-called WMC FWC Strategy The Sustainable Development Act provides for the use of wastewater as a nutrient medium, as well as industrial flue gases to replace traditional sources of CO2 in the cultivation of cyanobacteria and microalgae. Various types of wastewater can be used, including domestic wastewater, aquaculture wastewater, fermentation wastewater, wastewater from paper production, etc. Undoubtedly, such technologies reduce the load on environmental objects and significantly save resources and thus reduce the cost of production of necessary products, including in the field of bioenergy.
In addition, subsidies and investments should be directed to the development of infrastructure for storage, transportation, and scaling of biodiesel technologies. A significant part of the investment should be directed to highly efficient processing processes of the resulting biodiesel product, all of which require large investments in equipment and modern advanced technologies. Such political support is necessary, since it is clear that the production of biodiesel and the expansion of the market for its use will not occur until sufficient distribution and marketing infrastructure is established. In addition, there is an urgent need to introduce advanced technologies to improve the efficiency of biodiesel production, reduce their cost, which also requires sustainable investment. In general, the creation of a technological and technical base for the development of this bioenergy sector, the introduction of modern engineering developments and the production of necessary equipment, as well as the financing of regional projects in the field of energy and heat production from biodiesel requires large investments and political support from the state (The BEFS 2010).
All of the above determines the impact of various policy aspects on the profitability of biodiesel production in many industries, including agriculture, energy, transport, the environment and trade. The development of appropriate policies and the quantification of their impact in specific cases is complicated by the variety of policy instruments and their application. However, in many cases, these are subsidies aimed at supporting bioenergy and affecting the financial attractiveness of its production, trade and use. It should also be noted that the development of modern trends in the global market of any biofuel is accompanied by the development of contradictions of subjects at all levels from the state to individual economic structures and consumers, in which the economic, environmental and social consequences of the introduction of biofuels remain the subject of extensive disputes (Babu et al. 2013).
Thus, biofuels from phototrophic microorganisms are a complex subject that includes aspects such as economics, ecology, microbiology, biochemistry, genetics, chemical engineering, and politics. Of course, the complete replacement of fossil fuels with biofuels is not considered achievable, but modern biofuels have a crucial role to play in the long term to help limit the growth of greenhouse gas emissions and lead to the transition of the current oil-based society to a more sustainable fuel. According to experts, by 2030 the global production of biofuels can be increased to 150 million tons of oil equivalent with an annual growth rate of 7–9%. Its share will reach 4–6% of the total fuel consumed by the transport sector. Third and fourth generation biofuels, although still in the early stages of development, can replace more than 70 billion liters of fossil fuels annually, with a likely maximum development period in the period 2025–2035 (Babu et al. 2013).
Conclusion
Unconventional renewable energy sources such as biodiesel, which have been actively developed in recent years as a sustainable response to the coming environmental and food crisis, can be a worthy innovation to meet the basic living needs of the world's population. Cyanobacteria have a huge potential in bioenergy, whose lipids, having the right fatty acid profile, promising in the production of biodiesel, it is a promising renewable raw material for the production of biofuels. Their ability to grow in an environment rich in organic substances and the potential of these objects in the treatment of polluted wastewater, allows us to consider possible great prospects for low- and non-waste technologies for bioremediation of polluted water with the associated production of their cheap useful biomass. However, despite their high productivity and a wide range of produced lipid compounds, the existing number of problems that prevent their industrial use still requires intensive research and development. The improvement of biomass collection methods, cell disintegration approaches, the process of lipid recovery from wet biomass, and the search for and use of non-toxic solvents still require significant research efforts. Considering the unique properties of cyanobacteria and the possibilities of modern bioengineering approaches, great hopes are pinned on genetic engineering technologies that allow both to increase the synthesis of lipids by cells, and to establish ways to isolate them from cells, providing opportunities to bypass the intermediate expensive stages of cell destruction and, accordingly, the periodic increase in biomass. Thus, cyanobacteria with great prospects in the "third generation" fuel technologies, where their biomass is collected and chemically processed into fuel, and in the «fourth generation» fuel technologies, where they directly produce fuel from carbon dioxide and water, are of great interest to researchers, allowing them to minimize the consumption of organic resources of nature in the future and lead humanity to a new productive branch of energy-efficient development.
References
Abdelhamid SA, Hussein AA, Asker MS (2019) Optimization of culture conditions for biodiesel production from Egyptian isolate Penicillium commune NRC2016. Bull Natl Res Centre 43:15. https://doi.org/10.1186/s42269-019-0045-6
Abou-Shanab RA, Ji M-K, Kim H-C, Paeng K-J, Jeon B-H (2013) Microalgal species growing on piggerywastewater as a valuable candidate for nutrient removaland biodiesel production. J Environ Manag 115:257–264. https://doi.org/10.1016/j.jenvman.2012.11.022
Aratboni AH, Rafiei N, Garcia-Granados R, Alemzadeh A, Morones-Ramírez JR (2019) Biomass and lipid induction strategies in microalgae for biofuel production and other applications. Microb Cell Fact 18(1):178. https://doi.org/10.1186/s12934-019-1228-4
Ashokkumar V, Chen W-H, Ngamcharussrivichai C, Agila E, Ani FN (2019) Potential of sustainable bioenergy production from Synechocystis sp. cultivated in wastewater at large scale—a low cost biorefinery approach. Energy Convers Manag 186:188–199. https://doi.org/10.1016/j.enconman.2019.02.056
Babu V, Thapliyal A, Kumar PG (2013) Bio-hydrogen production: current scenarios and future prospects. In: Babu V, Thapliyal A, Patel GK (eds) Biofuels production. Wiley, Hoboken
Balaji S, Kalaivani T, Rajasekaran C, Shalini M, Siva R, Singh RK, Akthar MA (2014) Arthrospira (Spirulina) species as bioadsorbents for lead, chromium, and cadmium–a comparative study. Clean (weinh) 42:1790–1797. https://doi.org/10.1002/clen.201300478
Balan V (2014) Current challenges in commercially producing biofuels from lignocellulosic biomass. ISRN Biotechnol 2014:463074. https://doi.org/10.1155/2014/463074
Becker EW (1994) Microalgae: biotechnology and microbiology. Cambridge University Press, Cambridge, p 230
Behnam T, Afua A, Alok SK, Singh PK, Sitther V (2020) Nitrogen deprivation in Fremyella diplosiphon augments lipid production without affecting growth. Energies 21:5769. https://doi.org/10.3390/en13215769
Benemann JR, Oswald WJ (1996) Systems and economic analysis of microalgae ponds for conversion of CO2 to biomass. Final Rep 97:004328. https://doi.org/10.2172/493389
Blažina M, Haberle I, Hrusti E, Budiša A, Petri I, Konjevi L, Šilovi T, Djakovac T, Gecek S (2019) Growth aspects and biochemical composition of Synechococcus sp. MK568070 cultured in oil refinery wastewater. J Mar Sci Eng 7:164. https://doi.org/10.3390/jmse7060164
Bolatkhan K, Sadvakasova AK, Zayadan BK, Kakimova AB, Sarsekeyeva FK, Kossalbayev BD, Bozieva AM, Alwasel S, Allakhverdiev SI (2020) Prospects for the creation of a waste-free technology for wastewater treatment and utilization of carbon dioxide based on cyanobacteria for biodiesel production. J Biotechnol 324:162–170. https://doi.org/10.1016/j.jbiotec.2020.10.010
Bonnet M, Lagier JC, Raoult D, Khelaifia S (2019) Bacterial culture through selective and non-selective conditions: the evolution of culture media in clinical microbiology. New Microbes New Infect 34:100622. https://doi.org/10.1016/j.nmni.2019.100622
Boretti A, Rosa L (2019) Reassessing the projections of the World Water Development Report. Clean Water 2:15. https://doi.org/10.1038/s41545-019-0039-9
Branyikova I, Prochazkova G, Potocar T, Jezkova Z, Branyik T (2018) Harvesting of microalgae by flocculation. Fermentation 4(4):93. https://doi.org/10.3390/fermentation4040093
Cai T (2012) Cultivation of Nannochloropsis salina and Synechocystis sp. PCC6803 in anaerobic digestion effluent for nutrient removal and lipid production. Thesis. The Ohio State University. http://rave.ohiolink.edu/etdc/view?acc_num=osu1343748848
Cai T, Park SY, Li Y (2013) Nutrient recovery from wastewater streams by microalgae: status and prospects. Renew Sustain Energy Rev 19:360–369. https://doi.org/10.1016/j.rser.2012.11.030
Chen C-Y, Zhao X-Q, Yen H-W, Ho S-H, Cheng C-L, Lee D-J (2013) Microalgae-based carbohydrates for biofuel production. Biochem Eng J 78:1–10
Cheng CH, Du TB, Pi HC, Jang SM, Lin YH, Lee HT (2011) Comparative study of lipid extraction from microalgae by organic solvent and supercritical CO2. Bioresour Technol 102(21):10151–10153. https://doi.org/10.1016/j.biortech.2011.08.064
Chi X, Yang Q, Zhao F, Qin S, Yang Y, Shen J, Lin H (2008) Comparative analysis of fatty acid desaturases in cyanobacterial genomes. Comp Funct Genomics 284508:25. https://doi.org/10.1155/2008/284508
Comar CL, Nelson N (1975) Health effects of fossil fuel combustion products: report of a workshop. Environ Health Perspect 12:149–170
Cordeiro RS, Vaz ICD, Magalhães SMS, Barbosa FAR (2017) Effects of nutritional conditions on lipid production by cyanobacteria. An Acad Bras Cienc 89:2021–2031. https://doi.org/10.1590/0001-3765201720150707
Debnath M, Mandal NC, Ray S (2012) Effect of fungicides and insecticides on growth and enzyme activity of four cyanobacteria. Indian J Microbiol 52(2):275–280
Dellamatrice PM, Silva-Stenico ME, Moraes Beraldo LA, Fiore MF, Rosim RT (2017) Monteiro degradation of textile dyes by cyanobacteria. Braz J Microbiol 48(1):25–31. https://doi.org/10.1016/j.bjm.2016.09.012
Derakhshandeh M, Atici T, Umran TUN (2019) Lipid extraction from microalgae Chlorella and Synechocystis sp. using glass microparticles as disruption enhancer. Energy Environ 30(8):1341–1355
Deshmukh S, Bala K, Kumar R (2019) Selection of microalgae species based on their lipid content, fatty acid profile and apparent fuel properties for biodiesel production. Environ Sci 26(24):24462–24473. https://doi.org/10.1007/s11356-019-05692-z
Dmytryk A, Saeid A, Chojnacka K (2014) Biosorption of microelements by Spirulina: towards technology of mineral feed supplements. Sci World J 356328:15. https://doi.org/10.1155/2014/356328
Edokpayi JN, Odiyo JO, Durowoju OS (2017) Impact of wastewater on surface water quality in developing countries: a case study of South Africa. Water Qual. https://doi.org/10.5772/66561
El Fatah HMA, El-Baghdady KZ, Zakaria AE, Sadek HN (2020) Improved lipid productivity of Chlamydomonas globosa and Oscillatoria pseudogeminata as a biodiesel feedstock in artificial media and wastewater. Biocatal Agric Biotechnol 25:101588
El-Ardy O, El-Enin SAA, El Semary NA, El Diwani G (2012) Microalgal culture in photo-bioreactor for biodiesel production: case studies from Egypt. Afinidad LXIX, 558, Abril - Junio. https://raco.cat/index.php/afinidad/article/view/268401
El-Sheekh MM, El-Shouny WA, Osman ME (2014) Treatment of sewage and industrial wastewater effluents by the cyanobacteria Nostoc muscorum and Anabaena subcylinderica. J Water Chem Technol 36:190–197. https://doi.org/10.3103/S1063455X14040079
Eungrasamee K, Miao R, Incharoensakdi A, Lindblad P, Jantaro S (2019) Improved lipid production via fatty acid biosynthesis and free fatty acid recycling in engineered Synechocystis sp. PCC 6803. Biotechnol Biofuels 12(8):1–13. https://doi.org/10.1186/s13068-018-1349-8
Eungrasamee K, Incharoensakdi A, Lindblad P (2020) Synechocystis sp. PCC 6803 overexpressing genes involved in CBB cycle and free fatty acid cycling enhances the significant levels of intracellular lipids and secreted free fatty acids. Sci Rep 10:4515. https://doi.org/10.1038/s41598-020-61100-4
Ghasemi Y, Rasoul-Amini S, Naseri AT, Montazeri-Najafabady N, Mobasher MA, Dabbagh F (2012) Microalgae biofuel potentials (review). Appl Biochem Microbiol 48:126–144. https://doi.org/10.1134/S0003683812020068
Gopinath A, Sairam K, Velraj R, Kumaresan G (2015) Effects of the properties and the structural configurations of fatty acid methyl esters on the properties of biodiesel fuel: a review. Proc Inst Mech Eng D 229(3):357–390
Griffiths MJ, Harrison STL (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J Appl Phycol 21:493–507. https://doi.org/10.1007/s10811-008-9392-7
Gupta R, Ahuja P, Khan S, Saxena R, Mohapatra H (2000) Microbial biosorbents: meeting challenges of heavy metal pollution in aqueous solutions. Curr Sci 78(8):967–973
Haedrich J, Stumpf C, Denison MS (2020) Rapid extraction of total lipids and lipophilic POPs from all EU-regulated foods of animal origin: Smedes’ method revisited and enhanced. Environ Sci Eur 32:118. https://doi.org/10.1186/s12302-020-00396-5
Han F, Pei H, Hu W, Jiang L, Cheng J, Zhang L (2016) Beneficial changes in biomass and lipid of microalgae Anabaena variabilis facing the ultrasonic stress environment. Bioresour Technol 209:16–22. https://doi.org/10.1016/j.biortech.2016.02.103
Hewelt-Belka W, Kot-Wasik Á, Tamagnini P, Oliveira P (2020) Untargeted lipidomics analysis of the cyanobacterium Synechocystis sp. PCC 6803: lipid composition variation in response to alternative cultivation setups and to gene deletion. Int J Mol Sci 21:8883. https://doi.org/10.3390/ijms21238883
Hossain MF, Ratnayake RR, Mahbub S, Kumara WKL, Magana-Arachchi DN (2020) Identification and culturing of cyanobacteria isolated from freshwater bodies of Sri Lanka for biodiesel production. Saudi J Biol Sci 27(6):1514–1520. https://doi.org/10.1016/j.sjbs.2020.03.024
Ichor T, Okerentugba PO, Okpokwasili GC (2016) Biodegradation of total petroleum hydrocarbon by a consortium of cyanobacteria isolated from crude oil polluted Brackish Waters of Bodo Creeks in Ogoniland. Rivers State Res J Environ Toxicol 10:16–27. https://doi.org/10.3923/rjet.2016.16.27
Jawaharraj K, Karpagam R, Ashokkumar B, Kathiresan S, Moorthy IMG, Arumugam M, Varalakshmi P (2017) Improved biomass and lipid production in Synechocystis sp. NN using industrial wastes and nano-catalyst coupled transesterification for biodiesel production. Bioresour Technol 242:128–132. https://doi.org/10.1016/j.biortech.2017.03.067
Jeevanandam J, Harun MR, Lau SY, Sewu DD, Danquah MK (2020) Microalgal biomass generation via electroflotation: a cost-effective dewatering technology. Appl Sci 10(24):9053. https://doi.org/10.3390/app10249053
Kaiwan-arporn P, Hai PD, Thu NT, Ajit PA (2012) Cultivation of cyanobacteria for extraction of lipids. Biomass Bioenergy 44:142–149. https://doi.org/10.1016/j.biombioe.2012.04.017
Kalavathi DF, Subramanian Uma G (2001) Degradation and metabolization of the pigment-melanoidin in distillery effluent by the marine cyanobacterium Oscillatoria boryana BDU 92181. Enzym Microb Technol 29:246–251. https://doi.org/10.1016/S0141-0229(01)00383-0
Kamilya D, Sarkar S, Maiti TK, Bandyopadhyay S, Mal BC (2006) Growth and nutrient removal rates of Spirulina platensis and Nostoc muscorum in fish culture effluent: a laboratory-scale study. Aquac Res 37(15):1594–1597. https://doi.org/10.1111/j.1365-2109.2006.01588.x
Keris-Sen UD, Sen U, Soydemir G, Gurol MD (2014) An investigation of ultrasound effect on microalgal cell integrity and lipid extraction efficiency. Bioresour Technol 152:407–413. https://doi.org/10.1016/j.biortech.2013.11.018
Khan MI, Shin JH, Kim JD (2018) The promising future of microalgae: current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb Cell Fact 17(1):36. https://doi.org/10.1186/s12934-018-0879-x
Kim J, Lingaraju BP, Rheaume R, Lee J-Y, Siddiqui KF (2010) Removal of ammonia from wastewater effluent by Chlorella vulgaris. Tsinghua Sci Technol 15(4):391–396. https://doi.org/10.1016/S1007-0214(10)70078-X
Kumar N (2017) Oxidative stability of biodiesel: causes, effects and prevention. Fuel 190:328–350. https://doi.org/10.1016/j.fuel.2016.11.001
Kumar SPJ, Banerjee R (2019) Enhanced lipid extraction from oleaginous yeast biomass using ultrasound assisted extraction: a greener and scalable process. Ultrason Sonochem 52:25–32. https://doi.org/10.1016/j.ultsonch.2018.08.003
Kumar RR, Rao PH, Arumugam M (2015) Lipid extraction methods from microalgae: a comprehensive review. Front Energy Res 6(2):61. https://doi.org/10.3389/fenrg.2014.00061
Kumar A, Kaushal S, Saraf SA, Singh JS (2018) Microbial bio-fuels: a solution to carbon emissions and energy crisis. Front Biosci 23:1789–1802. https://doi.org/10.2741/4673
Lal A, Ghosh S, Das D (2018) Improvement in electrically induced biomass harvesting of Chlorella sp. MJ11/11 for bulk biomass production. J Appl Phycol 30:979–993. https://doi.org/10.1007/s10811-017-1268-2
Lau NS, Matsui M, Abdullah AA (2015) Cyanobacteria: photoautotrophic microbial factories for the sustainable synthesis of industrial products. BioMed Res Int 2015:754934. https://doi.org/10.1155/2015/754934
Liew WH, Hassim MH, Ng DKS (2014) Review of evolution, technology and sustainability assessments of biofuel production. J Clean Prod 71:11–29. https://doi.org/10.1016/j.jclepro.2014.01.006
Liu X, Sheng J, Curtiss R (2011) Fatty acid production in genetically modified cyanobacteria. Proc Natl Acad Sci 108(17):6899–6904. https://doi.org/10.1073/pnas.1103014108
Los DA, Mironov KS (2015) Modes of Fatty Acid desaturation in cyanobacteria: an update. Life 5(1):554–567. https://doi.org/10.3390/life5010554
Loura IC, Dubacq JP, Thomas JC (1987) The effects of nitrogen deficiency on pigments and lipids of Cyanobacteria. Plant Physiol 83(4):838–843
Lu X (2010) A perspective: photosynthetic production of fatty acid-based biofuels in genetically engineered cyanobacteria. Biotechnol Adv 28:742–746. https://doi.org/10.1016/j.biotechadv.2010.05.021
Maffei G, Bracciale MP, Broggi A, Zuorro A, Santarelli ML, Lavecchia R (2018) Effect of an enzymatic treatment with cellulase and mannanase on the structural properties of Nannochloropsis microalgae. Bioresour Technol 249:592–598. https://doi.org/10.1016/j.biortech.2017.10.062
Maheshwari N, Kumar M, Thakur IS, Srivastava S (2018) Carbon dioxide biofixation by free air CO2 enriched (FACE) bacterium for biodiesel production. J CO2 Util 27:423–432. https://doi.org/10.1016/j.jcou.2018.08.010
Maslova IP, Mouradyan EA, Lapina SS, Klyachko-Gurvich GL, Los DA (2004) Lipid fatty acid composition and thermophilicity of cyanobacteria. Russ J Plant Physiol 51:353–360. https://doi.org/10.1023/B:RUPP.0000028681.40671.8d
Matyash V, Liebisch G, Kurzchalia TV, Shevchenko A, Schwudke D (2008) Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J Lipid Res 49(5):1137–1146. https://doi.org/10.1194/jlr.d700041-jlr200
Milledge JJ, Heaven S (2014) Methods of energy extraction from microalgal biomass: a review. Rev Environ Sci Biotechnol 13:301–320. https://doi.org/10.1007/s11157-014-9339-1
Molazadeh M, Ahmadzadeh H, Pourianfar HR, Lyon S, Rampelotto PH (2019) The use of microalgae for coupling wastewater treatment with CO2 biofixation. Front Bioeng Biotechnol 7:42. https://doi.org/10.3389/fbioe.2019.00042
Mondal M, Goswami S, Ghosh A (2017) Production of biodiesel from microalgae through biological carbon capture: a review. 3 Biotech 7:99. https://doi.org/10.1007/s13205-017-0727-4
Mostafa SSM (2012) Microalgal biotechnology: prospects and applications. Plant Sci. https://doi.org/10.5772/53694
Nagappan S, Bhosale R, Duc Nguyen D, Pugazendhi A, Tsai PC, Chang SW, Ponnusamy VK, Kumar G (2020) Nitrogen-fixing cyanobacteria as a potential resource for efficient biodiesel production. Fuel 279:118440. https://doi.org/10.1016/j.fuel.2020.118440
Nagarkar S, Williams GA, Subramanian G, Saha SK (2004) Cyanobacteria-dominated biofilms: a high quality food resource for intertidal grazers. Hydrobiologia 512:89–95. https://doi.org/10.1007/978-94-007-0944-7_12
Oliveira DT, Vasconcelos CT, Feitosa AMT, Aboim JB, Oliveir AN, Xavier LP, Santos AS, Gonçalves EC, Filho GNR, Nascimento LAS (2018) Lipid profile analysis of three new Amazonian cyanobacteria as potential sources of biodiesel. Fuel 234(5):785–788. https://doi.org/10.1016/j.fuel.2018.07.080
Palanisamy KM, Paramasivam P, Jayakumar S, Maniam GP, Rahim AMH, Govindan N (2021) Economical cultivation system of microalgae Spirulina platensis for lipid production. IOP Conf Ser 641:12022. https://doi.org/10.1088/1755-1315/641/1/012022
Patel VK, Sundaram S, Patel AK (2018) Characterization of seven species of cyanobacteria for high-quality biomass production. Arab J Sci Eng 43:109–121. https://doi.org/10.1007/s13369-017-2666-0
Peng Z, Feng L, Wang X (1864) Miao X (2019) Adaptation of Synechococcus sp. PCC 7942 to phosphate starvation by glycolipid accumulation and membrane lipid remodeling. Biochim Biophys Acta Mol Cell Biol Lipids 12:158522. https://doi.org/10.1016/j.bcab.2020.101588
Pienkos PT, Darzins A (2009) The promise and challenges of microalgal-derived biofuels. Biofuels Bioprod Biorefin 3(4):431–440. https://doi.org/10.1002/bbb.159
Politaeva N, Smyatskaya Y, Toumi A, Oparina A (2018) Concentration of Chlorella sorokiniana microalga biomass at combined usage of coagulants and flocculants. Eurasian Chem-Technol J 20(3):243–247. https://doi.org/10.18321/ectj728
Qv XY, Zhou QF, Jiang JG (2014) Ultrasound-enhanced and microwave-assisted extraction of lipid from Dunaliella tertiolecta and fatty acid profile analysis. J Sep Sci 37(20):2991–2999. https://doi.org/10.1002/jssc.201400458
Radakovits R, Jinkerson RE, Darzins A, Posewitz MC (2010) Genetic engineering of algae for enhanced biofuel production. Eukaryot Cell 9(4):486–501
Rajeshwari KR, Rajashekhar M (2011) Biochemical composition of seven species of cyanobacteria isolated from different aquatic habitats of Western Ghats, Southern India. Braz Arch Biol Technol Curitiba 54(5):849–857. https://doi.org/10.1590/S1516-89132011000500001
Rathore DS, Kumar A, Kumar HD (1993) Lipid content and fatty acid composition in N2-fixing cyanobacterium Anabaena doliolum as affected by molybdenum deficiency. World J Microbiol Biotechnol 9:508–510. https://doi.org/10.1007/bf00386284
Ren X, Zhao X, Turcotte F, Deschênes JS, Tremblay R, Jolicoeur M (2017) Current lipid extraction methods are significantly enhanced adding a water treatment step in Chlorella protothecoides. Microb Cell Fact 16(1):26. https://doi.org/10.1186/s12934-017-0633-9
Richardson JW, Johnson MD, Outlaw JL (2012) Economic comparison of open pond raceways to photo bio-reactors for profitable production of algae for transportation fuels in the southwest. Algal Res 1(1):93–100. https://doi.org/10.1016/j.algal.2012.04.001
Rittmann BE, Krajmalnik-Brown R, Halden RU (2008) Pre-genomic, genomic and post-genomic study of microbial communities involved in bioenergy. Nat Rev Microbiol 6(8):604–612. https://doi.org/10.1038/nrmicro1939
Rosgaard L, de Porcellinis AJ, Jacobsen JH, Frigaard N, Sakuragi Y (2012) Bioengineering of carbon fixation, biofuels, and biochemicals in cyanobacteria and plants. J Biotechnol 162:134–147. https://doi.org/10.1016/j.jbiotec.2012.05.006
Ruffing AM, Jones HD (2012) Physiological effects of free fatty acid production in genetically engineered Synechococcus elongatus PCC 7942. Biotechnol Bioeng 109(9):2190–2199. https://doi.org/10.1002/bit.24509
Ruffing AM, Trahan CA, Jones HDT (2013) Genetic engineering of cyanobacteria as biodiesel feedstock. Sandia report. https://core.ac.uk/download/pdf/207094185.pdf
Ryckebosch E, Bruneel C, Muylaert K, Foubert I (2012) Microalgae as an alternative source of omega- 3 long chain polyunsaturated fatty acids. Lipid Technol 24(6):128–130. https://doi.org/10.1002/lite.201200197
Sajjadi B, Raman AAA, Arandiyan H (2016) A comprehensive review on properties of edible and non-edible vegetable oil-based biodiesel: composition, specifications and prediction models. Renew Sustain Energy Rev 63:62–92. https://doi.org/10.1016/j.rser.2016.05.035
Salvo RD, Reich A, Dykes HWH, Teixeira RJR (2010) Lipid extraction from microalgae using a single ionic liquid. Patent No. 8450111 US 12/970, 484
Samarasinghe N, Fernando S, Faulkner B (2012) Effect of high pressure homogenization on aqueous phase solvent extraction of lipids from Nannochloris Oculata microalgae. J Energy Nat Resour 1(1):1–7. https://doi.org/10.11648/j.jenr.20120101.11
Santos-Merino M, Garcillán-Barcia MP, da Cruz F (2018) Engineering the fatty acid synthesis pathway in Synechococcus elongatus PCC 7942 improves omega-3 fatty acid production. Biotechnol Biofuels Sep 5(11):239. https://doi.org/10.1186/s13068-018-1243-4
Sarsekeyeva F, Usserbaeva A, Zayadan B, Mironov K, Sidorov R, Kozlova A, Kupriyanova E, Sinetova M, Los D (2014) Isolation and characterization of a new cyanobacterial strain with a unique fatty acid composition. Adv Microbiol 4:1033–1043. https://doi.org/10.4236/aim.2014.415114
Sharma KK, Schuhmann H, Schenk PM (2012) High lipid induction in microalgae for biodiesel production. Energies 5:1532. https://doi.org/10.1016/j.bej.2013.03.006
Shashirekha V, Sridharan MR, Swamy M (2008) Biosorption of trivalent chromium by free and immobilized blue green algae: kinetics and equilibrium studies. J Environ Sci Health A Toxicol Hazard Subst Environ Eng 43:390–401. https://doi.org/10.1080/10934520701795608
Sheng J (2011) Downstream processing of Synechocystis for biofuel production. Thesis. Arizona State University. December. https://ui.adsabs.harvard.edu/abs/2011PhDT.......282S/abstract
Sheng J, Vannela R, Rittmann BE (2011) Evaluation of methods to extract and quantify lipids from Synechocystis PCC 6803. Bioresour Technol 102:1697–1703. https://doi.org/10.1016/j.biortech.2010.08.007
Silva CSP, Silva-Stenico ME, Fiore MF, Castro HF, Rós PCM (2014) Optimization of the cultivation conditions for Synechococcus sp. PCC7942 (cyanobacterium) to be used as feedstock for biodiesel production. Algal Res 3:1–7. https://doi.org/10.1016/j.algal.2013.11.012
Singh JS (2015) Plant-microbe interactions: a viable tool for agricultural sustainability. Appl Soil Ecol 92:45–46. https://doi.org/10.1016/j.apsoil.2015.03.004
Singh JS (2015) Microbes: the chief ecological engineers in reinstating equilibrium in degraded ecosystems. Agric Ecosyst Environ 203:80–82. https://doi.org/10.1016/j.agee.2015.01.026
Soydemir G, Keris-Sen UD, Sen U (2016) Biodiesel production potential of mixed microalgal culture grown in domestic wastewater. Bioprocess Biosyst Eng 39:45–51. https://doi.org/10.1007/s00449-015-1487-3
Sukenik A, Yamaguchi Y, Livne A (1993) Alterations in lipid molecular species of the marine eustigma tophyte Nannochlorosis Sp.1. J Phycol 29:620–626. https://doi.org/10.1111/j.0022-3646.1993.00620.x
Sun A, Davis R, Starbuck M, Ben-Amotz A, Pate R, Pienkos PT (2011) Comparative cost analysis of algal oil production for biofuels. Energy 36(8):5169–5179. https://doi.org/10.1016/j.energy.2011.06.020
Temina M, Rezankova H, Rezanka T, Dembitsky VM (2007) Diversity of the fatty acids of the Nostoc species and their statistical analysis. Microbiol Res 162(4):308–321. https://doi.org/10.1016/j.micres.2006.01.010
The BEFS Analytical Framework (2010) http://www.fao.org/3/i1968e/i1968e.pdf
Topare NS, Raut SJ, Renge VC, Khedkar SV, Chavan YP, Bhagat SL (2011) Extraction of oil from algae by solvent extraction and oil expeller method. Int J Sci 9(4):1746–1750
Torres CM, Ríos SD, Torras C, Salvadó J, Mateo-Sanz JM, Jiménez L (2013) Microalgaebased biodiesel: a multicriteria analysis of the production process using realistic scenarios. Bioresour Technol 147:7–16. https://doi.org/10.1016/j.biortech.2013.07.145
Tran HL, Hong SJ, Lee CG (2009) Evaluation of extraction methods for recovery of fatty acids from Botryococcus braunii LB 572 and Synechocystis sp. PCC 6803. Biotechnol Bioprocess Eng 14:187–192. https://doi.org/10.1007/s12257-008-0171-8
Uduman N, Bourniquel V, Danquah MK, Hoadley AFA (2011) A parametric study of electrocoagulation as a recovery process of marine microalgae for biodiesel production. Chem Eng J 174(1):249–257. https://doi.org/10.1016/j.cej.2011.09.012
Vandamme D, Foubert I, Muylaert K (2013) Flocculation as a low-cost method for harvesting microalgae for bulk biomass production. Trends Biotechnol 31(4):233–239. https://doi.org/10.1016/j.tibtech.2012.12.005
Wahidin S, Idris A, Shaleh SRM (2013) The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour Technol 129:11. https://doi.org/10.1016/j.biortech.2012.11.032
Wahlen BD, Willis RM, Seefeldt LC (2011) Biodiesel production by simultaneous extraction and conversion of total lipids from microalgae, cyanobacteria, and wild mixed-cultures. Bioresour Technol 102(3):2724–2730. https://doi.org/10.1016/j.biortech.2010.11.026
Wang X, Xiong X, Sa N (2016) Metabolic engineering of enhanced glycerol-3-phosphate synthesis to increase lipid production in Synechocystis sp. PCC 6803. Appl Microbiol Biotechnol 100:6091–6101. https://doi.org/10.1007/s00253-016-7521-9
Wang Q, Oshita K, Takaoka M (2021) Effective lipid extraction from undewatered microalgae liquid using subcritical dimethyl ether. Biotechnol Biofuels 14:17. https://doi.org/10.1186/s13068-020-01871-0
Wu X, Joyce EM, Mason TJ (2011) The effects of ultrasound on cyanobacteria. Harmful Algae 10(6):738–743. https://doi.org/10.1016/j.hal.2011.06.005
Yalcin D, Hoffmann W (2020) Growth, lipid content, and fatty acid profile of freshwater cyanobacteria Dolichospermum affine (Lemmermann) by using modified nutrient media. Aquacult Int 28:1371–1388. https://doi.org/10.1007/s10499-020-00531-2
Acknowledgements
This work was funded by the Ministry of Education and Science of the Republic of Kazakhstan in the framework of the Projects: AP09260077, AP08052402, AP08052481; SA would like to thank the Distinguished Scientists Fellowship Program, King Saud University, Saudi Arabia, for their support. SIA were supported by the grant from Russian Science Foundation (No: 19-14-00118).
Funding
The authors declare no competing financial interests.
Author information
Authors and Affiliations
Contributions
AKS, BDK, Conceptualization, methodology, software; AKS, BDK, BKZ, DKK, SA analysis, and investigation, resources, data curation; SIA, AKS, BDK writing -original draft preparation; AKS, SIA project administration; BDK, AKS, BKZ, SA, SIA, All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sadvakasova, A.K., Kossalbayev, B.D., Zayadan, B.K. et al. Potential of cyanobacteria in the conversion of wastewater to biofuels. World J Microbiol Biotechnol 37, 140 (2021). https://doi.org/10.1007/s11274-021-03107-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11274-021-03107-1