
Abstract
There are many different anthropogenic causes of wetland degradation, such as disturbances which affect the physical structure of wetlands, resulting in erosion (altered fire regimes, road and railway building through wetlands, channelization of wetlands), pollution, land-cover change, and climate change. These different types of degradation have various impacts, depending on the type of wetland, soils, biochemistry and other factors. We researched a poorly-studied South African valley-bottom peatland that is dominated by the ecosystem engineer Palmiet: Prionium serratum. We ask the question: what is the impact of degradation by gully erosion, pollution and alien tree invasion on biochemistry and plant community composition of palmiet wetlands? In 39 plots from three palmiet wetlands situated approximately 200 km apart we found that channel erosion, through a loss of alluvium, has probably resulted in leached soils with lower soil organic matter and water content, less able to retain nutrients and cations. Soil leaching is a possible explanation for the groundwater of degraded wetlands having higher electrical conductivity and pH than that of pristine wetlands and a lower soil cation exchange capacity (21.3 ± 5.80–7.7 ± 4.91 meq/100 g). The loss of alluvium typically resulted in a completely new plant community, composed mostly of pioneer species and several alien species. The increase in base saturation (17.5 ± 8.46–30.2 ± 17.85%) and soil pH (4.8 ± 0.51–5.1 ± 0.50) with degradation was hypothesized to be the result of liming practices. Once extremely degraded, i.e. all the alluvium is lost, it is unlikely that these sensitive palmiet wetlands will recover original vegetation communities and lost functions, except on long timescales. We recommend conservation of the few pristine wetlands that remain, and rehabilitation of those that still retain some of their original function.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wetlands make up less than 3% of the area of the globe, and less than 8.6% of land, and yet half of the worlds wetlands have been estimated to have been lost and many more have been degraded (Zedler and Kercher 2005; Nel et al. 2007; Meli et al. 2014; Rebelo et al. 2015; Ferreira and Lacerda 2016; Meng et al. 2017). There are many different types of wetland degradation, and some types of degradation trigger further impacts. Wetland degradation can be grouped loosely into four main categories: disturbance of wetland physical structure, land-cover changes, pollution and climate change (Zedler and Kercher 2005; Meng et al. 2017). Disturbance affecting the physical structure of wetlands may include drainage; either by humans or as a result of other damage (Zedler and Kercher 2005; Watters and Stanley 2007; Krüger et al. 2015), peat excavation (Nsor 2007; Winde 2011; Cabezas et al. 2014), erosion (e.g. channel or gully erosion) (Boardman 2014; Rebelo et al. 2015; de Haan 2016), mechanical disturbance, such as the building of roads or railways across wetlands, and altered fire regimes (Zedler and Kercher 2004). Types of land-cover changes include vegetation changes (Brooks et al. 2003), alien plant invasion (Zedler and Kercher 2004, 2005) or land conversion, for example to agriculture (Rebelo et al. 2015; de Haan 2016). Wetland pollution may be in the form of agricultural runoff, wetland fertilization, or point source pollution (Jordan et al. 2003; Zedler and Kercher 2005; Carpenter and Bennett 2011; Winde 2011). Ultimately the degradation of wetlands results in a loss of biodiversity and ecosystem services (Zedler and Kercher 2005; Meli et al. 2014) which may have economic consequences for local communities (Schuyt 2005).
Different types of wetland degradation affect wetland biochemistry, community composition and ecosystem functioning differently, however commonalities exist. Wetland degradation generally increases soil bulk density and causes a decline in organic matter/carbon content (Salimin et al. 2010; Huo et al. 2013; Sankura et al. 2014; Krüger et al. 2015). In contrast, impact on soil pH depends on the system and type of degradation, in some cases decreasing (Sankura et al. 2014) and in others increasing with degradation (Salimin et al. 2010; Aggenbach et al. 2013; Emsens et al. 2015). Wetland drainage of Northern Hemisphere ecosystems has been cited to cause a chain reaction of impacts, such as the loss of biodiversity, peat decomposition leading to the leaching of nutrients into rivers, as well as dust storms (Zedler and Kercher 2005; Krüger et al. 2015). Wetland drainage may also cause erosion, dramatically altering wetland form, eventually leading to channel formation (Watters and Stanley 2007). It is important to note that gully erosion is hypothesised to be a natural process that has played a role in shaping palmiet wetland formation through cut-and-fill processes leading to valley-floor planing (Job 2014; Pulley et al. 2018). However, it is likely that this natural process has also been accelerated in recent times by anthropogenic activities (Job 2014; Rebelo et al. 2017). In South Africa, accelerated channel erosion has been postulated to be the result of damage caused by roads bisecting wetlands or the landscape in general, however the processes behind this degradation are poorly researched (Boardman 2014; de Haan 2016; Rebelo et al. 2017). Overall wetland degradation has significant consequences for ecosystem service provision; for example the loss of soil organic matter or carbon results in a release of carbon into the atmosphere (Krüger et al. 2015). Gully erosion, on the other hand, increases siltation of dams and increases flood risk to downstream landowners (Job 2014; Rebelo et al. 2015).
South African wetlands and associated river systems are in a critical state, with over 65% estimated to be damaged, and over half destroyed (Nel et al. 2007). One such threatened wetland type, a unique valley-bottom peatland, occurs within the Cape Floristic Region of South Africa. These peatlands are unique because they are dominated by a wetland species endemic to southern Africa and listed as declining on the red list of South African plants: Palmiet (Prionium serratum). Palmiet is thought to be an ecosystem engineer (Sieben 2012). Due to its deep, extensive rooting structure and clonal nature, it is hypothesized to have stabilized river valleys within the Cape Floristic Region, creating a local base level and water ponding, forming unchannelled valley-bottom wetlands and allowing the accumulation of peat beds (Job 2014). Palmiet wetlands are highly threatened with degradation, having declined by almost 31% since the 1940s (Rebelo et al. 2017). The main threats faced by these wetlands are channel erosion, land-cover change (wetlands to agriculture), pollution from agricultural runoff (potentially both fertilizers and effluent from liming) and invasion by alien vegetation. These threats are often operating together and as a result it is very difficult to select sites which exclusively face only certain types of degradation and it is equally difficult to disentangle their effects.
Removal of wetland vegetation for agriculture is perhaps one of the more extreme changes to palmiet wetlands and since wetland vegetation is completely removed and sites are heavily manipulated with fertilizers, liming and irrigation, these impacts were not investigated in this study and these sites were avoided. The second most dramatic impact to these wetlands is channel erosion, which might be expected to draw down the water table in adjacent wetland habitat, resulting in an increase in soil bulk density and a decline in organic matter/carbon content (Salimin et al. 2010; Huo et al. 2013; Sankura et al. 2014; Krüger et al. 2015). This would increase soil oxygen in the root zone making soil nutrients available, which, combined with lower organic matter may result in the release of these nutrients into the groundwater (Zedler and Kercher 2005; Krüger et al. 2015). The impacts of alien plant and tree invasion on palmiet wetlands is difficult to study since in many places alien vegetation has been removed by restoration programmes such as Working for Water. Therefore, we did not explicitly consider these effects in this study. There has been research on the impacts of invasion by alien Acacias on riparian systems, which has shown an increase in nitrogen availability in the soil as well as enhanced phosphorus mineralization rates (Naudé 2012). Impacts from agricultural runoff may include an increase in bioavailable nutrients from fertilizers as well as an increase in pH due to liming practises common in agriculture in the Cape Floristic Region (Beukes et al. 2012). In this study, we ask the question: what is the cumulative impact of this complex, multifaceted wetland degradation on soil and groundwater biochemistry and vegetation community composition and chemistry of palmiet wetlands?
Methods
Study region and wetlands
The Cape Floristic Region has a predominantly mediterranean-type climate characterised by summer drought and winter rainfall resulting from the passage of cold fronts (Midgley et al. 2003). The soils of the Cape Floristic Region are mainly highly leached dystrophic lithosols associated with the sandstone mountains of the Cape Supergroup (Midgley et al. 2003). Three palmiet wetlands were selected as study sites within the Cape Floristic Region: the Theewaterskloof and Goukou wetlands (Western Cape) and the Kromme wetland (Eastern Cape) (Table 1). Despite being situated as much as 470 km apart; these wetlands are remarkably similar in vegetation composition. They occur on low slopes relative to the surrounding mountains, and below elevations of 400 m. Mean annual precipitation is highly variable, highest in the Theewaterskloof catchment and lowest in the Goukou catchment. Mean annual runoff is also highest in the Theewaterskloof catchment but lowest in the Kromme catchment. In the case of the Kromme and Goukou, most of this runoff occurs over a short period of time, during flood events (Job 2014; Rebelo et al. 2015). All three wetlands have accumulated peat layers between 0.5 and 10 m deep (Table 1). In some ways, these systems are similar to European bogs, in that they are highly oligotrophic and have a low pH (Wheeler and Proctor 2000). On the other hand, they are also similar to fens in terms of vegetation composition and hydrology.
Study design
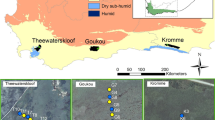
To capture seasonal variation in wetland properties and processes, each wetland was sampled twice: once in September 2014, which was just after winter, and once in March/April 2015, which was just after the summer. Rainfall for the 6 months preceding the first fieldtrip (winter) was average-to-low for all sites (approximating 50% of the MAP for the Theewaterskloof and Goukou), as a result of a drought. Rainfall for the 6 months preceding the second fieldtrip (summer) was far lower for Theewaterskloof (Table 1). For Theewaterskloof and the Kromme, the second campaign represents a drier season, whereas there is no such difference for the Goukou wetland as the rainfall regime is influenced by its proximity to the coast. At each wetland, both a degraded and an undisturbed (pristine) stretch was sampled, yielding a total of six sites. Degraded stretches were characterized by channelization or erosion with subsequent drainage, which was typically accompanied by alien tree or weed invasion (Table 2, Fig. 1). Additionally, degraded sites were often situated in an agricultural context, receiving polluted runoff from adjacent fields. Pristine stretches were selected such that there was no channelization or alien vegetation in the immediate vicinity (at least 300 m–1 km away), though it should be noted that all wetlands are transformed to some degree, with channelization occurring upstream or downstream of pristine fragments.

The location of the 39 plots (circles) and three study wetlands within the Cape Floristic Region (green) of South Africa. Orange borders show wetland fragments that are degraded (often channelized), whereas green borders indicate relatively undisturbed fragments. Blue-filled circles indicate the location of piezometers within the wetlands. See Table A5 for exact locations of plots. (Color figure online)
At each site, cross-sectional transects (100–200 m) were made across the wetland, with six plots (3 × 3 m) placed between 20 and 50 m apart, yielding a total of 36 plots across the six sites (Fig. 1). Transects and plots were chosen in the field to ensure adequate representation of the main vegetation communities. To this end we included one extra pristine site in the Goukou wetland, with only three plots, yielding a final sum of 39 plots. Piezometers (3 m, PVC) were placed adjacent to every second plot, yielding a total of 21 piezometers (Fig. 1).
Sampling
Plant community composition and vegetation analysis
In each plot, all plants were identified to species level where possible and percentage cover was estimated for each species using the Braun-Blanquet Scale (Mueller-Dombois and Ellenberg 1974). Total standing vegetation biomass was sampled from three small, randomly selected subplots of 0.28 × 0.28 m within each plot. This vegetation was weighed after oven drying for 48 h at 70 °C and then ground and homogenised using a mill until it could pass through a 0.5 mm mesh sieve. Plant total carbon and total nitrogen contents were determined by total combustion of 5 mg of each sample on a Flash 2000 CN-analyzer (Thermo Fisher Scientific). Plant total nitrogen and total phosphorus were determined using acid digestion and were measured with a continuous -flow analyzer (CFA) (SKALAR: SAN++) (Walinga et al. 1989). Plant total K, Ca and Mg were analyzed by Inductively Coupled Plasma-emission spectrometry (ICP-OES) (Walinga et al. 1989) after acid digestion of ± 0.3 g of dried and finely ground vegetation with H2SO4–Se-salicylic acid.
Soil sampling and chemical analyses
In each plot, one composite soil sample was taken from 10 points throughout the plot at a depth of 1–10 cm using a hand-held auger of 1 cm in diameter. Soil pH-H2O was measured after adding distilled water to a 10 g soil sample and shaking it for an hour. Additionally, in each plot one bulk density sample was taken of the topsoil using a 100 cm3 metal Kopecky ring. Samples were weighed after oven drying for 48 h at 70 °C and values are expressed as g/cm3. Soil moisture was calculated gravimetrically by weighing ± 20 g of fresh soil before and after drying for 24 h at 105 °C. Soil organic matter content was determined by loss on ignition (4 h at 550 °C). Total phosphorus and nitrogen were analyzed on a CFA. Soft extractions were done on fresh soil to determine NO3−–N, NH4+–N and PO43−–N; samples were extracted and preserved for later analysis on a CFA using AA-EDTA (ammonium acetate–ethylenediaminetetraacetic acid) for PO43−–P and AA-KCl (ammonium acetate–potassium chloride) for NO3−–N and NH4+–N respectively (Houba et al. 1989). Nutrient pools were calculated by multiplying nutrient concentrations by bulk density measurements. Total soil K, Ca and Mg were analyzed on the ICP-OES after acid digestion of ± 0.3 g of dried and finely ground soil with H2SO4–Se-salicylic acid (Walinga et al. 1989).
Cation exchange capacity was determined using the method of Brown (1943) by weighing ± 8 g of soil before and after air drying in an incubator at 40 °C for 48 h. Samples were then sieved through a 2 mm sieve, 25 ml ammonium acetate solution (1 M) was added to 2.5 g of soil and samples were shaken for 1 h. Soil pH was measured and samples were filtered through a 0.45 µm filter before being analyzed for H, Ca, K, Mg, Na, Al, Fe, Mn ions on an ICP-OES. CEC is calculated as the sum of all the ions, and base saturation is calculated as the percentage of base cations (Ca, K, Mg, Na) per CEC. Soil microbial biomass carbon was measured as a proxy for microbial activity in the soil at each site. We used the chloroform fumigation direct extraction protocol for microbial biomass carbon (Martens 1995; Beck et al. 1997).
Groundwater sampling and chemical analyses
Depth to the water table was measured in each piezometer using a sounding device, and the standing water was emptied using a bailer. Once fresh water had refilled the piezometer, a sample was taken for a pH and conductivity reading. Six water samples were taken and filtered (0.45 µm) to test for water quality parameters. The concentration of phosphate (PO43−–P), ammonium (NH4+–N), total phosphorus (P-tot), and total Kjeldahl nitrogen (Kj-N) were measured on a CFA. Concentrations of sodium, magnesium and calcium were measured on the ICP-OES.
Data analysis
We performed an ANOSIM and SIMPER analysis in R, using the ‘Vegan’ package for community ecology (Warton et al. 2012). We used analysis of similarity (ANOSIM) to determine whether vegetation communities from degraded and pristine wetland fragments were separable or not, using the Bray–Curtis dissimilarity index. Next, we performed a similarity percentage analysis (SIMPER) to determine which species characterised degraded and pristine fragments, as well as different wetlands.
To test the effect of degradation and wetland (Theewaterskloof, Goukou, Kromme) on soil, groundwater and vegetation parameters, we fitted linear mixed models taking season into account (2014, 2015) using the lme4 package in R (Bates et al. 2015). Plots were entered as a random effect to account for the dependence between observations from within the same plot. Wetland, degradation, season and the interaction between wetland and degradation were entered as fixed effects. First, the significance of the interaction was tested by comparing the fit of this model to a reduced model with only the three main effects. Where the interaction term was significant, we split the dataset by wetland and tested for the effect of degradation in all three wetlands separately. Where the interaction term was not significant, we excluded it from the model and tested the significance of the main effect: degradation. Significance was tested using an F-test with Kenward-Roger correction for degrees of freedom, as implemented in the “pbKRtest” package of R. All variables, except pH and ratios, were log(x + 1) transformed to fulfill assumptions of normality and homoscedasticity of linear mixed models.
Lastly a Detrended Correspondence Analysis (DCA) was performed on species abundance data, using the “vegan” package in R. Each season was analyzed separately (2014, 2015). Soil and vegetation parameters that differed significantly between degraded and pristine wetland fragments were correlated to the first and second axes, and overlain on the plot.
Results
The effect of degradation on abiotic parameters
Degraded wetland fragments had a significantly higher soil pH and significantly lower soil water content than their pristine counterparts (Table 3). Degraded wetland fragments also tended to have higher bulk densities and lower soil organic matter contents, though these differences were not significant for the Theewaterskloof wetland. In terms of nutrients, pristine wetland soils had higher nutrient (phosphorus and nitrogen) concentrations overall, as well as available nutrients (PO4–P, NH4–N) relative to degraded wetland soils. However, pools of total and available nutrients varied inconsistently among sites and among wetlands with the exception of the NO3–N pool, being significantly higher on degraded wetland fragments. Potassium concentration was significantly higher in pristine wetland soils. Cation exchange capacity was higher, and the base saturation lower in pristine wetland soils, though not significantly so for the Theewaterskloof wetland. Base cations, total soil Ca2+ and Mg2+ concentrations, therefore follow the same trend as cation exchange capacity, as do cations in general, having higher concentrations in pristine relative to degraded soils.
For all groundwater parameters, the interaction effects between the three wetlands were not significant; therefore, only results for pristine and degraded wetlands are shown (Table 4). Relative groundwater depth fluctuated substantially between sampling sessions in pristine wetland fragments (average fluctuations of 0.85–2.08 m), and to a lesser degree in degraded fragments (0.33–0.96 m). However, there was no detectable difference in relative groundwater depth between degraded and pristine sites (Table 4). The groundwater of degraded wetland fragments had a significantly higher pH and conductivity than pristine wetland fragments. There were no significant differences in nutrient levels (total or available) in the groundwater of degraded and pristine wetland fragments, except for biologically available nitrogen (NH4–N) which was significantly higher at degraded sites. Cations (Na+, Ca2+ and Mg2+) were significantly higher in the groundwater of degraded wetland fragments relative to pristine ones.
Vegetation on degraded wetland fragments had a significantly higher phosphorus concentration and a lower N/P ratio in their tissues relative to vegetation on pristine fragments (Table 5). For the Theewaterskloof and Goukou wetlands, total nitrogen followed the same trend; however, there was no difference for the Kromme (Table 5). The relative increase in total nitrogen and phosphorus was greatest in Theewaterskloof. Results of potassium concentrations in plant tissues were conflicting, in some cases higher for degraded wetland fragments (Goukou), in other cases higher for pristine (Kromme). Vegetation on degraded wetland fragments had a higher concentration of base cations (higher Mg2+, significantly higher Ca2+) in their tissues relative to pristine fragments.
The effect of degradation on wetland community composition
Degradation results in a change in vegetation communities in South African palmiet wetlands; results of the SIMPER analysis revealed 82% dissimilarity between degraded and pristine fragments. Pristine palmiet wetland fragments were characterized by patches of two main shrub-dominated vegetation communities: Palmiet (Prionium serratum) communities (mean: 81–86% cover) and those dominated by a mix of other fynbos wetland species. Several species were able to co-exist with P. serratum, albeit at a lower density, and these differentially characterized pristine palmiet wetland patches at each of the three study wetlands. Key species were: Psoralea aphylla, Restio paniculatus, Wachendorfia thyrsiflora (Theewaterskloof), R. paniculatus, Todea barbara (Goukou) and Cliffortia odorata and Helichrysum odoratissimum (Kromme). Fynbos communities in pristine wetland fragments were typically more diverse than Palmiet communities and species more evenly distributed. Some key species characterizing these communities were: Pteridium aquilinum and Isolepis prolifera (Theewaterskloof), and P. aquilinum, R. paniculatus, C. strobilifera, and Epischoenus gracilis (Goukou).
Vegetation communities of degraded palmiet wetland fragments, unlike pristine ones, were highly dissimilar among the three study wetlands, and were typically characterized by pioneer and alien vegetation (lower percentage cover of shrubs, increased trees and graminoids), except where small patches of P. serratum communities persisted (Table 6). Therefore, degraded wetland fragments tended to be more species-rich than pristine fragments. In each of the three wetlands, the key species characterizing degraded fragments were: P. aquilinum, I. prolifera, R. fruticosus, C. strobilifera, Carpha glomerata, Psoralea pinnata, and Laurembergia repens (Theewaterskloof), C. strobilifera, A. mearnsii and W. thyrsiflora (Goukou), and Pennisetum macrourum, C. strobilifera, H. odoratissimum, R. fruticosus, J. lomatophyllus, and I. prolifera (Kromme). Microbial biomass was significantly higher in the pristine wetland soils of the Goukou and Kromme relative to degraded soils, though not significantly so for Theewaterskloof wetland.
Relationship between vegetation composition and abiotic parameters
The results of the DCA, based on vegetation community structure from September 2014, show some degree of separation between degraded and pristine wetland sites and to some extent a grouping of sites from the same wetlands (Fig. 2). The results from 2015 showed similar trends and are displayed in Fig. A1. The degree of separation is noticeably much greater (especially in Fig. A1) for the Kromme wetland than for Goukou and Theewaterskloof wetlands. Degraded wetland sites tend to occupy the upper right quadrant, and pristine sites the lower left quadrant. However, three of the degraded Goukou wetland plots were more similar in community structure to pristine wetlands (plots were sampled in surviving patches of P. serratum), though their soil and groundwater characteristics were closer to those of degraded plots. In general, the first axis may represent a gradient from annuals (graminoids) and small herbaceous plants (P. decipens, L. repens etc.) on the right to longer-lived perennials on the left (T. barbara, P. serratum etc.). This axis seems to be explained (though not significantly) by soil pH, calcium and NH4-N. The second axis seems to capture some element of alien/weedy plant invasion, with weedy species (T. capensis, R. fructicosus) and alien trees (A. mearnsii) at the top of the plot, corresponding with degraded plots. Soil magnesium concentration (r2 = 0.4535, p = 0.001), water content (r2 = 0.3426, p = 0.001), and total vegetation phosphorus (r2 = 0.4660, p = 0.001) correlated weakly with the second axis. Vegetation N/P ratio was strongly positively correlated with many soil parameters: soil water content, potassium, magnesium, calcium and sodium concentration, cation exchange capacity, and soil organic matter. Vegetation tissue calcium and total phosphorus concentrations were positively correlated with soil bulk density, base saturation and nitrate pool.

Detrended Correspondence Analysis (DCA) of the vegetation communities on pristine and degraded wetland fragments of three South African palmiet wetlands sampled in September 2014. Degraded sites are in red, pristine in green. Symbols: filled triangle indicates Theewaterskloof, filled square indicates Goukou, filled circle indicates Kromme. Species names are given in black, and + indicates species with a lower abundance that would have masked by other labels. Abiotic parameters that were interesting or significantly different (bold) between pristine and degraded wetland fragments were overlain and are indicated by the arrows. Soil parameters are in brown, vegetation chemical composition in green. NSp number of species, MBC microbial biomass carbon, BS base saturation, BD bulk density. Stippled circles encompass sites from pristine and degraded wetland fragments. For full species names see supplementary material. (Color figure online)
Discussion
All three selected fragments of degraded palmiet wetlands were degraded by some degree of channel erosion, and all had some level of invasion of alien plants and trees. However only the Goukou and Kromme wetlands were situated in an agricultural context, and therefore were potentially impacted by agricultural effluent. These differences are important to consider, given that results are suggestive that these different types of degradation have had differing impacts on wetland biochemistry.
Possible influence of channel erosion on wetland biochemistry and community composition
Some key soil, groundwater and vegetation parameters differed between degraded and pristine palmiet wetland fragments, regardless of the specific wetland. Therefore, the site (wetland) effect was not significant, despite these wetlands being located far from each other (as much as 470 km apart). Since the main type of degradation that these three wetlands had in common was channel erosion, with concomitant or resultant invasion by alien vegetation; it can be inferred that these universal differences between degraded and pristine wetland patches are mainly attributable to channel erosion. Channel erosion impacts palmiet wetlands through the physical loss of soil, or alluvium during high flow/flood events (Rebelo et al. 2015; de Haan 2016; Rebelo et al. 2017). Under extreme channel erosion, the entire valley floor is removed, leaving only coarse sand behind (e.g. sections of the Kromme wetland) (Rebelo et al. 2017). In earlier stages of degradation by channel erosion, either patches of palmiet wetland vegetation may persist within the degraded fragment, retaining the alluvium within that patch (e.g. the degraded Goukou wetland), or the channel has only cut down through the alluvium in one area, leaving the wetland vegetation in the majority of the wetland fragment more-or-less intact (e.g. the degraded Theewaterskloof wetland). However, despite this gradient in degradation, degradation resulting from channel erosion had some clear impacts on palmiet wetland biochemistry.
Firstly, channel erosion appears to have caused a decrease in soil water content, which may be a result of loss of organic matter (soil water content and organic matter are well correlated; Table A4) and/or wetland drainage. However, water table draw-down was not found to be significant in this study due to considerable seasonal fluctuations. The bulk density of the soil also increased and soil organic matter decreased with degradation, though not significantly for the Theewaterskloof wetland, which is likely due to the earlier stage of degradation relative to the other two wetlands. For example, plots furthest from the channel in the degraded fragment of the Theewaterskloof wetland are impacted by lower soil water content, but the alluvium is not yet lost over the entire valley floor and therefore bulk density and organic matter content has not yet significantly changed. A decline in soil organic matter or carbon with degradation has been found in many wetland studies (Salimin et al. 2010; Huo et al. 2013). Cation exchange capacity and total soil nitrogen (concentration and pools) may also have decreased as a result of the loss of soil organic matter (Table A4). Overall degradation increased soil pH, though, again, these differences in the Theewaterskloof wetland were slight. Conversely, in studies of other wetland systems: restoration was found to increase soil pH of northern temperate fens (Aggenbach et al. 2013; Emsens et al. 2015) and wetland conversion to cropland or plantations in Ethiopia decreased soil pH (Sankura et al. 2014).
Probably as a result of the increased leaching of the topsoil, the groundwater of degraded wetlands had more base cations (Ca2+, Mg2+, Na+), higher ammonium, higher electrical conductivity and also a higher pH. However, the same trend is not seen for potassium, possibly because it is limiting. The legacy effects of this degradation can also be seen in the vegetation community, with vegetation on degraded fragments having almost double the total phosphorus and total nitrogen than that of pristine fragments as well as a lower N/P ratio. The reason that this may be most pronounced at the Goukou and especially the Theewaterskloof wetland may be a result of the early stage of channel erosion compared to the Kromme wetland where the alluvium has already been washed away at the degraded site. It is possible that in this earlier stage of wetland erosion, decomposition makes nutrients, especially nitrates, available for plant uptake before they are leached into the groundwater or washed away.
Higher soil pH has been shown to result in an increase of phosphorus uptake in vegetation in acid soils (Beukes et al. 2012), in this case driving a vegetation community that is less phosphorus limited. This is evidenced by decreasing vegetation N/P ratio and visibly by the higher incidence of alien weeds and competitive tree species such as Acacia mearnsii and Acacia saligna on degraded fragments (Zedler and Kercher 2004). Alien species exploiting an increase in nutrient availability have been observed in other aquatic ecosystems (Siemann and Rogers 2007; Li et al. 2011). It is probable that productivity (though not standing biomass) is higher on degraded sites due to the higher incidence of annuals, though we did not measure this. The results of the DCA are conflicting: the fact that no variable significantly correlated with the first axis suggests that either the main driver of vegetation community structure is not measured, or that the story is too complex to be untangled by correspondence analysis. One reason for this is that vegetation communities and wetland biochemistry may be out of sync, such that vegetation structure may in some cases still reflect more ‘pristine’ conditions, whereas the soil and groundwater reflect its degraded state. For example, localized channel erosion through a palmiet wetland (in the early stages) may draw down the water table locally, resulting in aerobic soil conditions which may increase decomposition and a change in plant nutrient and water availability. The vegetation community would probably remain the same at this early stage, but would be a legacy of the past soil conditions. With time, these species may be out-competed by other, better-adapted species, or may be eroded out as the gully erosion increases in severity.
Possible influence of agricultural effluent on wetland biochemistry and community composition
Although the impacts of degradation by channel erosion on palmiet wetland soil, groundwater and vegetation seem clear, some factors differed between wetlands, suggesting other explanations account for the patterns emerging. Several soil parameters differed significantly between degraded and pristine wetland fragments, but only for the Goukou and Kromme wetlands, and not for the Theewaterskloof wetland. We hypothesize that these differences are linked to the impacts of agricultural runoff. We did not measure this, however it is known that fertilizers and lime (usually dolomitic lime) are commonly applied to agricultural fields in the Cape Floristic Region of South Africa, particularly irrigated fields, to increase the pH and nutrient availability of extremely acidic and oligotrophic soils (Beukes et al. 2012). Both the Goukou and Kromme catchments have intensive irrigated agriculture adjacent to degraded palmiet wetlands.
Degradation as a result of pollution by agricultural runoff is one plausible explanation for the increased pH on degraded fragments of the Goukou and Kromme wetlands, as increased pH cannot be explained by the mechanisms around channel erosion alone. A characteristic of degradation by pollution of agricultural effluent may be the observed increased base saturation. We hypothesize that this is a result of base cations and hydrogen carbonates from the dolomitic lime leaching into the wetland soils from adjacent irrigated fields, resulting in the increase of base cations (Ca2+, Mg2+, Na+) that are evident in degraded palmiet wetland topsoils. We did not measure soils deeper than 10 cm and therefore it is possible that nutrient availability may differ in deeper soil layers. Although carbonates were not measured, we hypothesize that the carbonates from the dolomitic lime applied to agricultural fields also entered the wetland soils, possibly being the mechanism behind the observed increase in pH. It should be noted that these shifts in vegetation communities are less dramatic than those that occur with the loss of the entire alluvium as a result of channel erosion. It is unclear from this study what impact agricultural practices adjacent to palmiet wetlands are having on wetland biochemistry in terms of fertilizing/liming, and this should be investigated further.
Conclusion
There appear to be two major, compound types of degradation in palmiet wetlands. Channel erosion, often accompanied by invasion of alien plant species, causes a loss of alluvium and in extreme cases, the alluvium and accompanying vegetation of the entire wetland basin is washed away. Remaining soil has higher rates of decomposition as a result of the lowered water table and resultant aerobic conditions, which results in lower organic matter accumulation. A reduction in organic matter is hypothesized to cause a decrease in soil water content and cation exchange capacity, resulting in soil which is highly leached and less able to retain nutrients and cations. We hypothesize that as a result, groundwater had higher conductivity and pH in degraded wetlands. This channel erosion and resultant biochemical changes are associated with a completely new plant community, composed mostly of pioneer species, with patches of the original Prionium serratum wetland vegetation persisting–depending on the severity of degradation. The second type of degradation discussed here, pollution by agricultural effluent, may increase base saturation, possibly as a result of liming practices in agriculture. Loss of soil organic matter implies a release of CO2 into the atmosphere, loss of alluvium implies a decrease in flood attenuation [through the loss of peat and all the vegetation biomass on this peat, both of which would play a significant role in dissipating the force of floods (Rebelo et al. 2015)] and a reduction in cation exchange capacity and bulk density would imply a reduction in the water purification capacity of palmiet wetlands. This suggests a marked reduction in ecosystem service provision with palmiet wetland degradation. Once the pristine wetland soils and vegetation communities are lost, it would take tremendous effort and long timescales to restore these palmiet wetlands. This makes a case for the protection and conservation of remaining pristine patches of this unique South African valley-bottom wetland system.
References
Aggenbach CJS, Backx H, Emsens WJ, Grootjans AP, Lamers LPM, Smolders AJP, Stuyfzand PJ, Wolejko L, Van Diggelen R (2013) Do high iron concentrations in rewetted rich fens hamper restoration? Preslia 85:405–420
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting Linear mixed-effects models using lme4. J Stat Softw 67:1–48
Beck T, Joergensen RG, Kandeler E, Makeschin F, Nuss E, Oberholzer HR, Scheu S (1997) An inter-laboratory comparison of ten different ways of measuring soil microbial biomass C. Soil Biol Biogeochem 29:1023–1032
Beukes DJ, Fyfield TP, Jezile GG, Mapumulo TC (2012) Comparison of medium-term organic and inorganic fertiliser application on leaf nutrient concentration and yield of maize in rural agriculture in the Mbizana area, Eastern Cape province, South Africa. S Afr J Plant Soil 29:127–133. https://doi.org/10.1080/02571862.2013.777131
Boardman J (2014) How old are the gullies (dongas) of the Sneeuberg uplands, Eastern Karoo, South Africa? CATENA 113:79–85. https://doi.org/10.1016/j.catena.2013.09.012
Brooks AP, Brierley GJ, Millar RG (2003) The long-term control of vegetation and woody debris on channel and flood-plain evolution: insights from a paired catchment study in southeastern Australia. Geomorphology 51:7–29. https://doi.org/10.1016/S0169-555X(02)00323-9
Brown IC (1943) A rapid method of determining exchangable hydrogen and total exchangable bases of soils. Soil Sci 56:353–357
Cabezas A, Pallasch M, Schönfelder I, Gelbrecht J, Zak D (2014) Carbon, nitrogen, and phosphorus accumulation in novel ecosystems: Shallow lakes in degraded fen areas. Ecol Eng 66:63–71. https://doi.org/10.1016/j.ecoleng.2013.10.037
Carpenter SR, Bennett EM (2011) Reconsideration of the planetary boundary for phosphorus. Environ Res Lett 6:14009. https://doi.org/10.1088/1748-9326/6/1/014009
de Haan V (2016) The Effects of Erosion-control Structures and Gully Erosion on Groundwater Dynamics along the Kromrivier, Eastern Cape, South Africa. Masters Thesis. Stockholm University
Emsens WJ, Aggenbach CJS, Smolders AJP, van Diggelen R (2015) Topsoil removal in degraded rich fens: can we force an ecosystem reset? Ecol Eng 77:225–232. https://doi.org/10.1016/j.ecoleng.2015.01.029
Ferreira AC, Lacerda LD (2016) Degradation and conservation of Brazilian mangroves, status and perspectives. Ocean Coast Manag 125:38–46. https://doi.org/10.1016/j.ocecoaman.2016.03.011
Houba R, Van der Lee JJ, Novozamsky I, Wallinga I (1989) Soil and Plant Analysis, a Series of Syllabi. Part 5. Wageningen
Huo L, Chen Z, Zou Y, Lu X, Guo J, Tang X (2013) Effect of Zoige alpine wetland degradation on the density and fractions of soil organic carbon. Ecol Eng 51:287–295. https://doi.org/10.1016/j.ecoleng.2012.12.020
Job N (2014) Geomorphic origin and dynamics of deep, peat-filled, valley bottom wetlands dominated by palmiet (Prionium serratum)—a case study based on the Goukou Wetland, Western Cape. Rhodes University, Eastern Cape
Jordan TE, Whigham DF, Hofmockel KH, Pittek MA (2003) Nutrient and sediment removal by a restored wetland receiving agricultural runoff. J Environ Qual 32:1534. https://doi.org/10.2134/jeq2003.1534
Kotze DC (2015) A survey of organic soils in the upper Riviersonderend catchment. WWF, South Africa
Krüger JP, Leifeld J, Glatzel S, Szidat S, Alewell C (2015) Biogeochemical indicators of peatland degradation—a case study of a temperate bog in northern Germany. Biogeosciences 12:2861–2871. https://doi.org/10.5194/bg-12-2861-2015
Li H, Lei GC, Zhi YB, An SQ, Huang HP, Ouyang Y, Zhao L, Deng ZF, Liu YH (2011) Nitrogen level changes the interactions between a native (Scirpus triqueter) and an exotic species (Spartina anglica) in coastal China. PLoS ONE 6:10. https://doi.org/10.1371/journal.pone.0025629
Martens R (1995) Current methods for measuring microbial biomass C in soil: Potentials and limitations. Biol Fertil Soils 19:87–99
Meli P, Benayas JRM, Balvanera P, Ramos MM (2014) Restoration enhances wetland biodiversity and ecosystem service supply, but results are context- dependent : a meta-analysis. PLoS ONE 9:e93507. https://doi.org/10.1371/journal.pone.0093507
Meng W, He M, Hu B, Mo X, Li H, Liu B, Wang Z (2017) Status of wetlands in China: a review of extent, degradation, issues and recommendations for improvement. Ocean Coast Manag 146:50–59. https://doi.org/10.1016/j.ocecoaman.2017.06.003
Middleton BJ, Bailey AK (2008) Water resources of South Africa, 2005 study (WR2005). Pretoria, South Africa
Midgley GF, Hannah L, Millar D, Thuiller W, Booth A (2003) Developing regional and species-level assessments of climate change impacts on biodiversity in the Cape Floristic Region. Biol Conserv 112:87–97. https://doi.org/10.1016/S0006-3207(02)00414-7
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, Hoboken, pp 93–135
Naudé M (2012) Fynbos Riparian Biogeochemistry and Invasive Australian Acacias
Nel JL, Roux DJ, Maree G, Kleynhans CJ, Moolman J, Reyers B, Rouget M, Cowling RM (2007) Rivers in peril inside and outside protected areas: a systematic approach to conservation assessment of river ecosystems. Divers Distrib 13:341–352. https://doi.org/10.1111/j.1472-4642.2007.00308.x
Nsor AC (2007) Plant community distribution and diversity, and threats to vegetation of the Kromme River Peat Basins, Eastern Cape Province. Rhodes University, South Africa
Pulley S, Ellery WN, Lagesse JV, Schlegel PK, McNamara SJ (2018) Gully erosion as a mechanism for wetland formation: an examination of two contrasting landscapes. Land Degrad Dev 29:1–12. https://doi.org/10.1002/ldr.2972
Rebelo AJ, Le Maitre DC, Esler KJ, Cowling RM (2015) Hydrological responses of a valley-bottom wetland to land-use/land-cover change in a South African catchment: Making a case for wetland restoration. Restor Ecol 23:829–841. https://doi.org/10.1111/rec.12251
Rebelo AJ, Scheunders P, Esler KJ, Meire P (2017) Detecting, mapping and classifying wetland fragments. Remote Sens Appl Soc Environ 8:212–223
Salimin MI, Gandaseca S, Ahmed OH, Majid NMA (2010) Comparison of selected chemical properties of peat swamp soil before and after timber harvesting. Am J Environ Sci 6:164–167. https://doi.org/10.3844/ajessp.2010.164.167
Sankura H, Lemma B, Ram N (2014) Effect of changing natural forest and wetland to other land uses on soil properties and stocks of carbon and nitrogen in South Ethiopia. Carpathian J Earth Environ Sci 9:259–265
Schuyt KD (2005) Economic consequences of wetland degradation for local populations in Africa. Ecol Econ 53:177–190. https://doi.org/10.1016/j.ecolecon.2004.08.003
Sieben EJJ (2012) Plant functional composition and ecosystem properties: the case of peatlands in South Africa. Plant Ecol 213:809–820. https://doi.org/10.1007/s11258-012-0043-3
Siemann E, Rogers WE (2007) The role of soil resources in an exotic tree invasion in Texas coastal prairie. J Ecol 95:689–697. https://doi.org/10.1111/j.1365-2745.2007.01253.x
Walinga I, Van Vark W, Houba VJG, Van Der Lee JJ (1989) Plant analysis procedures. Soil and Plant Analysis, Part 7. Wageningen, NL
Warton DI, Wright TW, Wang Y (2012) Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol Evol 3:89–101
Watters JR, Stanley EH (2007) Stream channels in peatlands: The role of biological processes in controlling channel form. Geomorphology 89:97–110. https://doi.org/10.1016/j.geomorph.2006.07.015
Wheeler BD, Proctor MCF (2000) Ecological gradients, subdivisions and terminology of north-west European mires. J Ecol 88:187–203. https://doi.org/10.1046/j.1365-2745.2000.00455.x
Winde F (2011) Peatlands as filters for polluted mine water?—a case study from an uranium-contaminated karst system in South Africa—part II: examples from literature and a conceptual filter model. Water 3:291–322
Zedler JB, Kercher S (2004) Causes and consequences of invasive plants in wetlands: opportunities, opportunists, and outcomes. CRC Crit Rev Plant Sci 23:431–452. https://doi.org/10.1080/07352680490514673
Zedler JB, Kercher S (2005) Wetland resources: status, trends, ecosystem services, and restorability. Annu Rev Environ Resour 30:39–74. https://doi.org/10.1146/annurev.energy.30.050504.144248
Acknowledgements
A.J.R. gratefully acknowledges the following organizations for funding: The Erasmus Mundus Programme (European Commission), Research Fund of the University of Antwerp (Belgium), the National Research Foundation (South Africa) and a Consolidoc Grant (Stellenbosch University). Thank you to Byron-Mahieu van der Linde for assistance in the field and Tom van der Spiet for lab assistance. We also wish to thank two anonymous reviewers for helpful suggestions which improved the manuscript. Research was conducted under permit number: 0052-AAA008-00019 (Cape Nature).
Author information
Authors and Affiliations
Contributions
AJR conceived the study, performed data analysis, wrote the manuscript and KJE, PM, WE, contributed and edited it.
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Rebelo, A.J., Emsens, WJ., Meire, P. et al. The impact of anthropogenically induced degradation on the vegetation and biochemistry of South African palmiet wetlands. Wetlands Ecol Manage 26, 1157–1171 (2018). https://doi.org/10.1007/s11273-018-9638-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-018-9638-3