
Abstract
Allometric relationships between easily measurable independent variables like diameter at breast height (DBH) and total height (TH) and biomass of leaf, branch, bark, stem without bark, and total above-ground biomass of Excoecaria agallocha L. were tested. Similarly, relationships between independent variables (DBH and HT); and nutrients (N, P and K) and carbon content in total above-ground biomass were also evaluated. The best fit allometric models were selected by considering the values of parameter of estimation of the models such as R2, CV, Rmse, MSerror, Sa, Sb, and Akaike’s information criterion corrected (AICc). The selected allometric models were Log Biomass = 0.9256 Log DBH2 − 2.133; Log biomass = 1.1656 Log DBH2 − 1.7047; Log biomass = 1.0824 Log DBH2 − 1.7568; Log biomass = 1.0927 Log DBH2 − 1.0275; Log biomass = 1.0996 Log DBH2 − 0.8572 respectively for leaf, branch, bark, stem without bark and total above-ground biomass. Whereas the selected allometric models for nitrogen, phosphorus, potassium and carbon were Log N = 1.0972 Log DBH2 − 3.0845; Log P = 1.0947 Log DBH2 − 5.6790; Log K = 1.0990 Log DBH2 − 3.0370; Log C = 1.1 Log DBH2 − 1.1937 respectively. Highest concentration of nitrogen (16.07 mg/g) and phosphorus (0.05 mg/g) was observed in leaf, while higher concentration (45.95–48.60 %) of carbon was observed in stem and bigger branches. The findings of this study are first-hand information for forest managers and conservation workers for sustainable management of E. agallocha in the Sundarbans of Bangladesh.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mangroves are important and most productive ecosystem of the tropical and sub-tropical forest types (Field 1995). Mangroves are used as nursery and breeding sites for birds, fish, crustaceans, shellfish, reptiles and mammals; a renewable source of wood; accumulation sites for sediment, contaminants, carbon and nutrients; and offer protection against coastal erosion, cyclones and tsunami (Mazda et al. 1997; Alongi 2002; Mahmood et al. 2005, 2008; Mahmood 2014). Sundarbans is the largest single tract of mangrove forest in the world, within the delta of the Ganges, Brahmaputra and Meghna rivers in the Bay of Bengal (Islam and Gnauck 2008). The prominent plant species are Heritiera fomes and Excoecaria agallocha with uneven distributions of Xylocarpus granatum, X. mekongensis, Ceriops decandra, Bruguiera spp., Avicennia spp., Sonneratia spp. Heritirea fomes is the dominant species of the Sundarbans, which alone constitute 21 % of the forest area. While, H. fomes–E. agallocha, E. agallocha–H. fomes and C. decandra dominated forest types constitute 29.7, 14.8 and 14.46 % of the forest area respectively (Siddiqi 2001). E. agallocha is the 2nd most important timber resource of the Sundarbans in terms of abundance, distribution and commercial value. It is a small to medium sized tree (Siddiqi 2001) and found to dominate at the southeastern part of moderate salinity areas of the Sundarbans (Ellison et al. 2000). This species was used as a raw material for newsprint and fuel wood. The density of E. agallocha with diameter at breast height (DBH) of 10 and 15 cm has been decreased to 35 and 44 % during the period of 1985 and 1996 respectively (Hussain and Ahmed 1994). This scenario indicates over-exploitation attitudes of E. agallocha. Irrespectively, it implies that the previous felling scheme was not sustainable for sustainable production of this species. Therefore, there is an urgent need to develop a new felling scheme of E. agallocha on the basis of biomass production and nutrient budgeting.
Biomass studies on mangroves are very important to know the details of successional changes, production and competition in vegetal communities, site productivity, nutrient cycling, nutrient budget, amount of carbon, predicting future change due to climate change and overall to manage the forest in a sustainable way (Komiyama et al. 2008; Mahmood 2014). There are three methods of biomass estimation: area harvest method, mean-tree method and allometric method (Golley et al. 1975; Ketterings et al. 2001). But, the allometric method is frequently used for estimating the forest biomass and this method is less destructive and the most powerful tool for estimating the biomass of a tree from measurable dimensions (Clough and Scott 1989; Mahmood et al. 2004, 2012; Komiyama et al. 2005, 2008). Allometric models for biomass estimation vary greatly among the species and sites (Clough and Scott 1989; Komiyama et al. 2008), it is preferable to use species and site specific models (Ketterings et al. 2001). The allometric method of biomass estimation in mangrove forests has been studied for several decades in different mangrove forest of the world (e.g. Clough and Scott 1989; Komiyama et al. 2000, 2002; Tamai et al. 1986; Mahmood et al. 2004), but there are limited works for the Sundarbans of Bangladesh e.g. C. decandra (Mahmood et al. 2012) and Aegialitis rotundifolia (Siddique et al. 2012). The objectives of this study were to develop allometric models for above-ground biomass, nutrients (N, P and K) and carbon stock in E. agallocha. The study would generate first-hand information for forest managers and conservation workers for sustainable management of the species in the Sundarbans on the basis of biomass production and nutrient budgeting.
Materials and methods
Study area
This study was conducted at E. agallocha dominated forest types of the Sundarbans mangrove forest of Bangladesh, between latitudes of 21°30′ and 22°30′N and longitudes of 89°00′ and 89°55′E. The subtropical climate of the study area is strongly seasonal with 87 % of the mean annual rainfall (1500 mm) falling from May to October. Minimum and maximum temperatures were 18.50–35.20 °C in summer and 12.20–28.80 °C in winter. Soil texture of the study area was silty to sandy clay loam. The bulk density, particle density and porosity varied from 1.18 to 1.27 g/cc, 2.31 to 2.52 g/cc and 46–52 %, respectively. Soil pH, conductivity and salinity were 7.8, 1.19 mS/cm and 5.94 dS/cm respectively (Siddiqi 2001).
Sample collection and processing
Excoecaria agallocha appears as single stem tree in the Sundarbans. A total of 30 individuals of E. agallocha were selected to cover the range of DBH from 2 to 22 cm (Table 1) and this was the representative of DBH range of this species in the Sundarbans (Siddiqi 2001). This DBH range was divided into five classes as 2–6, 6–10, 10–14, 14–18 and 18–22 cm. Six individuals from each class were selected avoiding mechanically or insect damaged or infested with disease. DBH and total height (TH) of the selected individuals were measured and felled at the ground level. The above ground parts of the individual were then separated into leaves, smaller branches (diameter < 2 cm), bigger branches (diameter > 2 cm) and main stems. One 0.5 m long stem section for smaller individuals and a 1 m long section for larger trees were collected from the base, middle and upper portion of the main stem for each individual. These stem sections were debarked in the field to calculate fresh weight ratio of bark and stem wood (Mahmood et al. 2004, 2012). All parts of each individual were weighted (fresh weight) separately in the field and recorded. Ten subsamples (about 0.25 kg) from each part were brought back to the laboratory and oven-dried at 80 °C for 10 days to get the conversion ratio of fresh weight to oven-dried weight. The oven-dried biomass of each part (leaf, branch, bark and stem without bark) of individual tree was estimated from the derived conversion ratio of respective plant part. Finally, mean biomass proportion for each part was also estimated in accordance with the DBH classes.
Nutrients (N, P and K) and carbon in plant part
Ten samples (about 100 g) of leaves, branches, stems and barks were collected from the felled trees and samples were oven-dried at 80 °C until constant weight for 10 days. The oven-dried samples were grounded, processed and acid digestion according to Allen (1974). The digested sample extracts were processed according to Weatherburm (1967) and Timothy et al. (1984) to measure nitrogen and phosphorus concentration in sample extracts respectively using UV–Visible Recording Spectrophotometer (U-2910, HITACHI, Japan). Potassium concentration in sample extracts was also measured by Flame photometer (PFP7, Jenway LTD, England). Carbon content was measured by combustion method by using Leco CR—12 Carbon determinator, USA. Nutrients and carbon concentration in different parts were compared by one-way analysis of variance (ANOVA) followed by Duncan Multiple Range by using SAS (6.12) statistical software. The amount of nutrients and carbon in each part of individual tree was estimated from their concentration and biomass of respective plant parts. While the total amount of nutrient and carbon in total above-ground biomass of individual tree was estimated by summing up their amount in each part.
Allometric models for above-ground biomass, nutrients and carbon stock
Allometric relationships between independent variables (DBH and TH) and oven-dry biomass of plant parts (leaf, branch, bark, stem without bark, total above-ground biomass); nutrients (N, P and K) and carbon stock in above-ground biomass were evaluated. Linear regression equations (y = a + bX) with different transformation (e.g. square root, Log10 and ln) of independent and dependent variables were used in these allometric relationships. A total of eight linear models with 64 regression equations with combination of DBH and TH were tested for each part (Table 2). Significance tests of regression equations were tested by using SAS (6.12) statistical software. The best fit allometric models were selected by considering those equations having the highest R2 with lowest values of CV, Rmse, MSerror, Sa, Sb and AICc, where R2 = Coefficient of determination; CV = Co-variance, Rmse = Root mean square error; MSerror = Mean square error; Sa = Standard error of intercept ‘‘a’’; Sb = Standard error of regression coefficient ‘‘b’’; AICc = Akaike’s information criterion corrected (Chave et al. 2005; Soares and Schaeffer-Novelli 2005; Siddique et al. 2012). A correction factor for allometric models was also estimated to solve the deformation during the log10 and ln transformation of variable values and back to the initial values (Sprugel 1983).
Results
Biomass proportion, nutrients and carbon
The mean biomass proportion of plant parts was found to vary with DBH classes. Comparatively higher proportion of stem biomass (67.34 %) was observed at lowest DBH class of 2.1–6 cm while higher proportion of branch biomass was observed at higher DBH classes (Table 3). Nutrients (N, P and K) and carbon concentration significantly (p < 0.05) varied among the plant parts. Highest concentration of nitrogen (16.07 mg/g) and phosphorus (0.05 mg/g) was observed in leaf and highest concentration (16.72 mg/g) of potassium was detected in smaller branches. Higher concentration (45.95–48.60 %) of carbon was observed in woody parts (stem and bigger branches) of E. agallocha (Table 4).
Allometric models
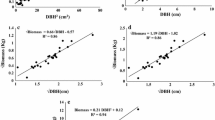
A total of eight linear models with 64 regression equations with combination of DBH and TH were tested for each plant part, which yield a total of 320 equations for estimating biomass of plant parts (Table 2). All the equations were significant (p < 0.05), but 290 equations were excluded considering the value of co-efficient of determination (R2) less than 0.84 for leaf, 0.96 for branch, 0.99 for bark, stem without bark and total above ground biomass. The selected allometric models were Leaf: Log Biomass = 0.9256 Log DBH2 − 2.133; Branch: Log biomass = 1.1656 Log DBH2 − 1.7047; Bark: Log biomass = 1.0824 Log DBH2 − 1.7568; Stem without bark: Log biomass = 1.0927 Log DBH2 − 1.0275; Total above-ground biomass: Log biomass = 1.0996 Log DBH2 − 0.8572 and that have been marked with “*” in the Table 5.
Nutrients (N, P and K) and carbon content in plant parts and total above-ground biomass were estimated and allometric equations were determined for their stock in total above-ground biomass. A total of 8 linear models with 64 regression equations with combination of DBH and TH were tried for the stock of nutrients and carbon in total above-ground biomass, which yield a total of 256 equations (Table 2). All the equations were significant (p < 0.05) and 240 equations were excluded considering the value of co-efficient of determination (R2) less than 0.99 for Nitrogen, Phosphorus, Potassium and Carbon. The selected allometric models were Nitrogen: Log N = 1.0972 Log DBH2 − 3.0845; Phosphorus: Log P = 1.0947 Log DBH2 − 5.6790; Potassium: Log K = 1.0990 Log DBH2 − 3.0370; and Carbon: Log C = 1.1 Log DBH2 − 1.1937 that have been marked with “*” in the Table 6.
Discussion
Biomass proportion in plant parts is important to understand the photosynthate allocation to plant growth (Kozlowski et al. 1991). Similar to the study, higher proportion of biomass in stem was observed with lower DBH classes of Bruguiera Parviflora in Malaysia (Mahmood et al. 2004), Rhizophora apiculata and R. stylosa in north-eastern Australia (Clough 1992). Biomass proportion in plant parts of mangrove trees were not only affected by the geographical location and microclimates but also related to forest structure, species-specific architecture (Tomlinson 1986; Steinke et al. 1995) and stages of development (seedlings, saplings and trees) (Mahmood et al. 2004).
The trend of nitrogen, phosphorus and potassium in different parts of the studied species was similar to that of C. decandra (Mahmood et al. 2012), R. apiculata (Ong et al. 1984), Avicennia spp., Bruguiera spp. and Ceriops spp. (Aksornkoae and Khemnark 1984) and B. parviflora (Mahmood et al. 2006). Physiologically more active tissue (leaf, flower) usually contain higher concentration of nutrients (Binkley 1986) and woody parts (stem and bigger branches) contain higher concentration of carbon (Mahmood 2014). Nutrients and carbon concentration in plant parts vary with the structural component of plant cell (Kaakinen et al. 2004), species, physiological age of the tissue, position of the tissue in plant, available form of nutrients in the substrate, concentration of other nutrients, climatic and soil edaphic factors may be the reason for variation in nutrient concentration in plant parts (Mahmood 2004).
Biomass estimation allometric models of mangroves commonly use total height, DBH, Girth at Breast Height (GBH) and Girth at Collar Height (GCH) as independent variables (Clough and Scott 1989; Gong and ong 1990; Cienciala et al. 2006; Mahmood et al. 2012; Siddique et al. 2012). Tree height measurement is complicated, time consuming and involves error in measurement. But, DBH is easy to measure and more acceptable physical parameter of trees (Komiyama et al. 2002). Accurate measurement of independent variables in biomass models will increase its acceptability and applicability (Overman et al. 1994). Similar consideration was made for the present study and the recommended allometric model of leaf biomass includes DBH as independent variable (Log Biomass = 0.9256 Log DBH2 − 2.133) instead of the model (Log biomass = 0.5994 Log DBH2 × HT2 − 2.4814) with higher R2 and AICc values (Table 5). In a comparison with the present study, the common allometric biomass model of mangroves (Komiyama et al. 2005) showed similar amount of total above-ground biomass for the individual having DBH of 2–5 cm. Conversely, the common model gives about 16–60 % over estimation of total above-ground biomass for trees of higher DBH (range 6–22 cm) (Fig. 1). This difference in biomass estimation of the present study and Komiyama et al. (2005) may be due to the variation in biophysical condition of the study areas (Bangladesh, Indonesia and Thailand). Furthermore, the common allometric model did not include E. agallocha as sample tree and individual having DBH less than 5 cm. Irrespectively, the variation in estimated biomass may also be related to the differences of coefficients in allometric models (Mahmood et al. 2004). Even the coefficients value of the same species may vary with localities and depends on site quality, tree density, species composition and management practices (Clough 1992; Steinke et al. 1995). In conclusion, the common allometric model is not applicable for E. agalloch in the Sundarbans and species specific allometric equations are required for accurate estimation of tree biomass. The findings of this study will be useful to estimate biomass, nutrients and carbon stock for single stem E. agallocha with its wide range of DBH (2–22 cm), which finally contributes in planning for future management and sustainable exploitation of this species.

Estimated total above-ground biomass of Excoecaria agallocha
References
Aksornkoae S, Khemnark C (1984) Nutrient cycling in mangrove forest of Thailand. In: Soepadmo E, Rao AN, Macintosh DJ (eds) Proceedings of the Asian symposium on mangrove environment research and management. University of Malaya, Kuala Lumpur, pp 545–557
Allen SE (1974) Chemical analysis of ecological materials. Blackwell Scientific, Oxford
Alongi DM (2002) Present state and future of the world’s mangrove forests. Environ Conserv 29(3):331–349
Binkley D (1986) Forest nutrition management. Wiley, New York
Chave J, Andalo C, Brown S, Cairns MA, Chambers JQ, Eamus D, Folster H, Fromard F, Higuchi N, Kira T, Lescure JP, Nelson BW, Ogawa H, Puig H, Riera B, Yamakura T (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145:87–99
Cienciala E, Černý M, Tatarinov F, Apltauer J, Exnerová Z (2006) Biomass functions applicable to Scots pine. Trees 20:483–495
Clough BF (1992) Tropical Mangrove ecosystem, coastal and estuarine studies 41. In: Robertson AI, Alongi DM (eds) Primary productivity and growth of mangrove forests. American Geophysical Union, Washington, pp 225–250
Clough BF, Scott K (1989) Allometric relationships for estimating above-ground biomass in six mangrove species. For Ecol Manag 27:117–127
Ellison AM, Mukherjee BB, Karim A (2000) Testing patterns of zonation in mangroves: scale dependence and environmental correlates in the Sundarbans of Bangladesh. J Ecol 88:813–824
Field CD (1995) Journey amongst mangroves. International Society of Mangrove Ecosystems, Japan
Golley BF, Mc Ginnis TJ, Clements GR, Child IG, Duever JM (1975) Mineral cycling in a tropical moist forest ecosystem. University of Georgia Press, Athens
Gong WK, Ong JE (1990) Plant biomass and nutrient flux in a managed Mangrove forest in Malaysia. Estuar Coast Shelf Sci 31:519–530
Hussain Z, Ahmed I (1994) Management of the forest resources. In: Hussain Z, Acharya G (eds) Mangroves of the Sundarbans, 2nd edn. IUCN, Bangkok
Islam SN, Gnauck A (2008) Mangrove wetland ecosystems in Ganges–Brahmaputra delta in Bangladesh. Front Earth Sci China 2(4):439–448
Kaakinen S, Jolkkonen A, Iivonen S, Vapaavuori E (2004) Growth, allocation and tissue chemistry of Picea abies seedlings affected by nutrient supply during the second growing season. Tree Physiol 24:707–719
Ketterings QM, Coe R, Noordwijk MV, Amagau Y, Palm CA (2001) Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forest. For Ecol Manag 146:199–209
Komiyama A, Havanond S, Srisawatt W, Mochida Y, Fujimoto K, Ohnishi T, Ishihara S, Miyagi T (2000) Top/root biomass ratio of a secondary mangrove (Ceriops tagal (Perr.) C. B. Rob.) forest. For Ecol Manag 139:127–134
Komiyama A, Jintana V, Sangtiean T, Kato S (2002) A common allometric equation for predicting stem weight of mangroves growing in secondary forests. Ecol Res 17:415–418
Komiyama A, Poungparn S, Kato S (2005) Common allometric equations for estimating the tree weight of mangroves. J Trop Ecol 21:471–477
Komiyama A, Ong JE, Poungparn S (2008) Allometry, biomass, and productivity of mangrove forest: a review. Aquat Bot 89:128–137
Kozlowski TT, Kramer JP, Pallardy SG (1991) The physiological ecology of woody plants. Academic Press, San Diego
Mahmood H (2004) Biomass, Litter Production and Selected Nutrients in Bruguiera Parviflora (Roxb.) Wight & Arn. Dominated Mangrove Forest Ecosystem at Kuala Selangor, Malaysia. PhD thesis, University Putra Malaysia, Seri Kembangan, unpublished
Mahmood H (2014) Carbon pools and fluxes in Bruguiera parviflora dominated naturally growing mangrove forest of Peninsular Malaysia. Wetl Ecol Manag 22(1):15–23
Mahmood H, Saberi O, Japar Sidik B, Misri K, Rajagopal S (2004) Allometric relationships for estimating above and below-ground biomass of saplings and trees of Bruguiera parviflora (Wight and Arnold). Malays Appl Biol 33(1):37–45
Mahmood H, Misri K, Japar Sidik B, Saberi O (2005) Sediment accretion in a protected mangrove forest at Kuala Selangor. Malays Pak J Biol Sci 8(1):149–151
Mahmood H, Saberi O, Misri K, Japar Sidik B (2006) Seasonal variation in concentrations of N, P and K in different components of Bruguiera parviflora (Wight and Arnold) at three growth stages in Malaysia. Indian J For 29(2):149–155
Mahmood H, Saberi O, Japar Sidik B, Misri K (2008) Net primary productivity of Bruguiera parviflora (Wight & Arn.) dominated mangrove forest at Kuala Selangor Malaysia. For Ecol Manag 255:179–182
Mahmood H, Siddique MRH, Bose A, Limon SH, Saha S, Chowdhury MRK (2012) Allometry, above-ground biomass and nutrient distribution in Ceriops decandra (Griffith) Ding Hou dominated forest types of the Sundarbans mangrove forest, Bangladesh. Wetl Ecol Manag 20:539–548
Mazda Y, Magi M, Kogo M, Hong PN (1997) Mangroves as a coastal protection from waves in the Tong King Delta Vietnam. Mangrove Salt Marshes 1:127–135
Ong JE, Gong WK, Wong CH (1984) Seven years of productivity studies in a Malaysian managed mangrove forest, then what? In: Bardsley KN, Davie JDS, Woodroffe CD (eds) Coastal and tidal wetlands of the Australian monsoon region. Australian National University, Darwin, pp 213–223
Overman JPM, Witte HJL, Saldarriaga JG (1994) Evaluation of regression models for above ground biomass determination in Amazon rainforest. J Trop Ecol 10:207–218
Siddiqi NA (2001) Mangrove forestry in Bangladesh. University of Chittagong, Institute of Forestry & Environmental Science, Chittagong
Siddique MRH, Mahmood H, Chowdhury MRK (2012) Allometric relationship for estimating above-ground biomass of Aegialitis rotundifolia Roxb. of Sundarbans mangrove forest, in Bangladesh. J For Res 23(1):23–28
Soares MLG, Schaeffer-Novelli Y (2005) Above-ground biomass of mangrove species. I. Analysis of models. Estuar Coast Shelf Sci 65:1–18
Sprugel DG (1983) Correcting for bias in log-transformed allometric equations. Ecology 64(1):209–210
Steinke DT, Ward CJ, Rajh A (1995) Forest structure and biomass of mangroves in the Mgeni estuary, South Africa. Hydrobiologia 295:159–166
Tamai S, Nakasuga T, Tabuchi R, Ogino K (1986) Standing biomass of mangrove forests in southern Thailand. J Jpn Soc 68:384–388
Timothy RP, Carol ML (1984) A manual of chemical and biological methods for seawater analysis. Pergamon press, New York
Tomlinson PB (1986) The botany of mangroves. Cambridge University Press, Cambridge
Weatherburm MW (1967) Phenol-hypochlorite reaction for determination of ammonia. Anal Chem 39(8):971–974
Acknowledgments
We are thankful to the Ministry of Science and Technology of Bangladesh for this research Grant. We are also indebted to Forestry and Wood Technology Discipline and Forest Department for their logistic support and continuous adherences.
Funding
Ministry of Science and Technology of Bangladesh for the research grant
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hossain, M., Siddique, M.R.H., Saha, S. et al. Allometric models for biomass, nutrients and carbon stock in Excoecaria agallocha of the Sundarbans, Bangladesh. Wetlands Ecol Manage 23, 765–774 (2015). https://doi.org/10.1007/s11273-015-9419-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-015-9419-1