
Abstract
Nitrogen (N) deposition is predicted to impact on the structure and functioning of Mediterranean ecosystems. In this study, we measured plant species composition, production and root phosphatase activity in a field experiment in which N (0, 10, 20 and 50 kg N ha−1 year−1) was added since October 2007 to a semiarid shrubland in central Spain. The characteristically dominant annual forb element responded negatively to N after ~2.5 and ~3.5 years. In contrast, the nitrophilous element (mainly crucifers) increased with N after ~2.5 and ~5.5 years, a response controlled by between-year variations in rainfall and the heterogeneous distribution of P availability. We also described a hierarchy of factors driving the structure and composition of the plant community: soil fertility was the most important driver, whereas calcareousness/acidity of soils and shrub cover played a secondary role; finally, N deposition contributed to explain a smaller fraction of the total variance, and its effects were predominantly negative, which was attributed to ammonium toxicity. Root phosphatase activity of three species was not responsive to N after ~2.5 years but there was a negative relationship with soil P in two of them. We conclude that increased N deposition in semiarid Mediterranean ecosystems of Europe can contribute to cause a shift in plant communities associated with an increase in the nitrophilous element and with a decline in abundance of various forb species adapted to the local conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Nitrogen (N) deposition threatens plant communities of European Mediterranean ecosystems (Phoenix et al. 2006) but information concerning the main implications on this area is still scarce (Bonanomi et al. 2006; Ochoa-Hueso et al. 2011a). This is due to the lack of field N fertilization experiments using realistic N loads and simulating patterns of dry vs. wet deposition (Ochoa-Hueso et al. 2011a). There is also a lack of gradient studies relating atmospheric N deposition loads with indicators of the ecosystem health in this region (Bobbink et al. 2010; Ochoa-Hueso et al. 2014). Consequently, information from manipulation experiments conducted in semiarid Californian chaparral and coastal sage scrub are usually viewed as models to predict potential consequences of N deposition on natural and semi-natural ecosystems from other areas with a semiarid Mediterranean climate; i.e. Mediterranean Basin, Cape Region, Australia and Chile (Vourlitis and Pasquini 2009; Fenn et al. 2010; Ochoa-Hueso et al. 2011a).
Plant communities from terrestrial ecosystems worldwide have shown shifts in species composition following N inputs that may be related to their strategies for N metabolism and to competition for available resources (Bobbink et al. 2010). The general rule predicts that under increased N availability, plant species adapted to the local conditions are frequently replaced by fast-growing nitrophilous plants (Emmett 2007). Several mechanisms potentially contribute to these shifts, namely: (i) nutritional imbalances (mainly N to phosphorus (P)—Carfrae et al. 2007); (ii) species adapted to low N inherently grow slowly, so are progressively out competed by faster-growing nitrophiles; nitrophiles also tend to produce more seeds, which causes additional impacts on the native soil seed bank (e.g. Fisher et al. 2009; Ochoa-Hueso and Manrique 2010); and (iii) negative effects due to high ammonium levels in soils where nitrate is usually the dominant mineral N form (Bobbink et al. 2010). The increase of plant competition for space and light is considered to only play a small role in the alteration of plant communities in low productivity semiarid sites such as lowland fynbos or shrublands from central Spain (except in the case of unusually wet years; Ochoa‐Hueso and Manrique 2014), where induced nutritional imbalances, increased soil ammonium availability and other soil-mediated indirect effects are usually viewed as the main factors driving responses to soil N enrichment (Ochoa-Hueso et al. 2011a; Bobbink et al. 2010).
Semiarid Mediterranean ecosystems are characterized by a pronounced climatic seasonality, which conditions the dominant form of N deposition; i.e. dry N deposition dominates during rainless periods (accumulating on soil, plant and biological soil crust surfaces) and becomes available as high N concentration pulses with rainfall events (Fenn et al. 2003). In these systems, water is therefore considered the main limiting factor of plant production, whereas N often plays a secondary and regulatory role (Cox and Allen 2008; Ochoa-Hueso and Manrique 2010). Despite the above-mentioned climatic similarities, soils from different Mediterranean areas often differ, which conditions potential responses to increased N deposition (Hobbs and Richardson 1995). One of the most relevant differences is related to the P status of soils, as the biodiversity in these systems is frequently controlled by the soil N to P ratio (Fanelli et al. 2008). The most P-rich soils are normally found in Chile and California and so the existing data from California can only be applied cautiously to the Mediterranean Basin, where soils are usually poorer in P (Hobbs and Richardson 1995). The inorganic P can be supplied to plants by a number of mechanisms, including direct uptake of microbially mineralized P from organic sources or the direct action of plant roots, and this process is catalysed by the phosphatase group of enzymes, including phosphomonoesterases (PME) (Johnson et al. 1999). Therefore, the N to P imbalance induced by anthropogenic N pollution has usually been found to be related to increased root-surface PME activity and to the alteration of arbuscular mycorrhizal fungi (AMF) colonization rates (Phoenix et al. 2003). For example, AMF colonization was reduced in N-enriched California soils that are high in P (Egerton-Warburton and Allen, 2000), whereas the increase in colonization rates with N fertilization in a low-P semiarid shrubland in central Spain was dependent on the availability of potential soil inoculum (Ochoa-Hueso et al. 2013a).
Mediterranean ecosystems are also characterized by a high spatial and temporal heterogeneity in terms of plant cover and soil resources (Gallardo et al. 2000; Cruz et al. 2008; Ochoa-Hueso et al. 2011a), which affects the potential to find consistent responses to N fertilization even across small spatial scales within the same area (Ochoa-Hueso et al. 2011a). For instance, Witkowski (1989a) found that the effects of increased N availability on plant productivity of lowland fynbos were masked by the heterogeneity in initial plant cover and that covariance analysis significantly contributed to clarify results. In addition to the confounding effects of superimposed natural environmental gradients and of the heterogeneous distribution of plant cover, the response of Mediterranean-type ecosystems to increased N availability also depends on the stage of ecological succession (Aber et al. 2003). For instance, Cruz et al. (2008) reported that certain early successional species prefer nitrate and are more susceptible to increased ammonium availability, while certain later successional species tend to be more ammonium tolerant. This question also highlights the potential relative importance of the main form of inorganic N pollution to predict impacts on plant communities (Bobbink et al. 2010; Stevens et al. 2011).
Within this ecological context, we designed a field experiment to identify responses of an Iberian semiarid shrubland to a simulated N deposition gradient. We planned an integrated survey where biomass, community composition and physiological measurements of plants were taken in order to report the main impacts of N fertilization on the plant community. We hypothesized that, being originally dominated by species typical of low-N (or at least low-ammonium) environments, the plant community would experience significant alterations when extra N (in the form of ammonium nitrate) was added (Bobbink et al. 2010). We also hypothesized that responses would be species-specific, ranging from negative for most of the annual plant species typical of the area to positive in the case of the nitrophilous element (Bobbink et al. 2010). In the semiarid portions of central Spain, this nitrophilous element is often characterized by fast-growing annual species within the Cruciferae and Asteraceae family, among which Diplotaxis DC., Sysimbrium L. and Centaurea L. can be cited (Ochoa‐Hueso and Manrique 2014). Finally, we predicted that root-surface PME activity would be enhanced with extra N, with this response being conditioned by the pre-existing P status of soils and the identity of the studied species (Phoenix et al. 2003).
2 Methods
2.1 Study Area
A semiarid Mediterranean shrubland located in a relatively N polluted area ~50 km away from the city of Madrid and within the Nature Reserve “El Regajal-Mar de Ontígola” (central Spain, 40° 00ʹ N, 3° 36ʹ W; mean altitude 580 m above the sea level) was selected for this study. Annual rainfall in the study site is ~425 mm, mainly falling from October to May (Rivas-Martínez 1987). Annual plants are absent most of the year, and a diverse and well-developed late-successional biological soil crust composed by terricolous lichens and mosses (predominantly Pleurochaete squarrosa (Brid.) Lindb) is frequent in the interspaces between shrubs (Ochoa-Hueso et al. 2011b). Dominant shrub species are Quercus coccifera L. (Fagaceae), Rosmarinus officinalis L. (Labiatae), Lithodora fruticosa (L.) Griseb. (Boraginaceae) and Rhamnus lycioides L. (Rhamnaceae). The herbaceous layer includes species such as Sedum sp. L. (Crassulaceae), Asterolinon linum-stellatum (L.) Duby (Primulaceae), Limonium echiodes L. (Mill.) (Plumbaginaceae), Stoibrax dichotomum (L.) Rafin. (Umbelliferae), Iberis crenata Lam. (Cruciferae) or Platycapnos spicata (L.) Bernh. (Fumariaceae). There are no leguminous shrubs and herbs. The shrubland is located at the top and middle parts of a hill. Soils are rich in calcium carbonate and with a slightly basic pH (8.0). Nitrate is the dominant inorganic N form in soils, with mean (2008–2011) soil values ranging from 8.2 to 20.7 mg N kg soil−1 (Ochoa-Hueso et al. 2013b). Ammonium ranges from 4.0 to 7.6 mg N kg soil−1 on average (Ochoa-Hueso et al. 2013b). Extractable P in soils is frequently below 0.5 mg kg−1, and higher values have usually been related to animal activity, mainly rabbits and partridges, and to seasonal processes such as organic matter mineralization (Ochoa-Hueso et al. 2013b). Temporal variation, i.e. seasonality, of other nutrients in the soil is also high (Ochoa-Hueso et al. 2013b). Mean values for other macro- and micronutrients in soils of the study site are reported in Ochoa-Hueso and Manrique (2011) and Ochoa-Hueso et al. (2013b).
2.2 Nitrogen Fertilization Experiment
Twenty-four plots (2.5 × 2.5 m) were established in October 2007 within the shrubland in a six-block design. Plots within each block (four plots per block) were randomly assigned to one of four N treatments (0, 10, 20 and 50 kg N ha−1 year−1; n = 6 replicates per treatment) over the background deposition (wet + dry deposition, ~6.4 kg N ha−1 year−1; Ochoa-Hueso et al. 2013ab). All these treatments, excluding the one receiving 50 kg N ha−1 year−1, are either reasonably falling within the predicted N deposition scenarios for the Mediterranean Basin by 2050 (~12–24 kg N ha−1 yr−1; Phoenix et al. 2006) or within N deposition loads reported in other Mediterranean regions (up to 45 kg N ha−1 year−1; Fenn et al. 2003). Nitrogen was applied once a month from September to June, when the majority of rain occurs, as wet deposition in the form of 2 L ammonium nitrate solution corresponding to N doses of 0 (i.e. only water), 0.019, 0.037 and 0.093 M. We sprayed the whole plot surface, including soil, biological soil crusts and plants. At the end of September, the first month after the dry season, a triple dose of N was added to simulate the accumulated N deposition that is washed into the soil with the first rains (Ochoa-Hueso et al. 2013b).
2.3 Destructive Plant Sampling (2010 and 2013 Growing Seasons)
Each plot was subdivided into a destructive (north side) and a nondestructive (south side) sampling half, each of 1.25 × 2.5 m in size. During May 2010, total above-ground biomass of annual herbaceous species was harvested in the destructive half of each plot. The plant material was transported to the lab in paper bags, oven-dried at 65 °C to constant weight, and then weighed. Individuals of three selected annual forbs (I. crenata, L. echioides and S. dichotomum) and of annual grasses (overall considered as functional group and including species such as Desmazeria rigida (L.) Tutin) were also separately weighed. These species were chosen among all the annual species present at the study site because they were present and abundant enough across most of the experimental plots, whereas the rest of the annual species were too scarce for subsequent statistical analyses. In early June 2013, and in order to investigate in detail the response of the nitrophilous element to increased N, we harvested all individuals of Diplotaxis siifolia G. Kunze, Sysimbrium commune (Speg.) Romanczuk and Centaurea melitensis L. present in each plot as described before. The rest of the vegetation, with the exception of the local crucifer I. crenata, which was also collected in order to compare responses, remained untouched. Sampling campaigns were conducted in May/early June as this time of the year coincides at the study site with the end of the spring growing season, which effectively starts at the end of February/beginning of April.
2.4 Nondestructive Plant Sampling (2011 Growing Season)
During May 2011, the density of every vascular plant species present (including annual species and small chamaephytes such as Helianthemum violaceum (Cav.) Pers.) was consistently recorded in the nondestructive half of each plot. Each half was gridded into six 0.5 × 0.5 m squares, leaving a perimeter band of 0.5 m toward south and 0.25 m toward east and west. Three squares in alternating disposition were sampled. Each square was subdivided into 25 10 × 10 cm sub-squares, where individual counts were made. Sampling in 2011 was separately conducted for plants living in the interspaces and under shrubs, and these microsites (interspace vs. shrub) were considered separately in the statistical analyses (see below).
2.5 Root-Surface PME Activity (2010 Growing Season)
The PME activity determinations (considered as a potential indicator of N saturation and of an induced N to P imbalance, Phoenix et al. 2003) were conducted only in 2010 on fresh root samples of three different vascular plant species, two annual forbs (I. crenata and L. echioides) and current-year seedlings of the dominant woody species (R. officinalis). In May 2010, roots from different individuals of each species from each plot were collected and pooled in plastic vials previously filled with deionized water. Between two and ten roots per species were included depending on plant material availability. Dry weight of roots per plot was approximately 40, 20 and 5 mg for I. crenata, L. echioides and R. officinalis, respectively. Once all sampling was finished, root material was rapidly transported to the lab. In the lab, vials were emptied of water in order to eliminate soil particles and dirt. Once the roots were clean, root-surface PME activity was determined by measuring the release of p-nitrophenol (p-NP) from p-nitrophenyl phosphate (p-NPP). Two millilitres of 0.1 M maleate buffer at pH 6.5 and 0.5 ml of substrate were added to the fresh roots. Vials were incubated at 37 °C for 90 min. The reaction was stopped by cooling rapidly to 4 °C for 15 min; 0.5 M CaCl2 and 2 ml of 0.5 M NaOH were then added, and the mixture centrifuged at 3000 rpm for 15 min. The amount of p-NP was determined using a spectrophotometer (Shimadzu UV-1603) at 398 nm (Johnson et al. 1999). After the spectrophotometrical determinations, roots were oven-dried at 65 °C to constant weight and then weighted to present the phosphatase activity in a time and dry weight basis (Phoenix et al. 2003).
2.6 Soil Sampling and Chemical Analysis
In order to investigate the existing relationships between soil chemical properties and plant community composition and physiology, soil samples were taken in spring 2010 and 2011 immediately before the monthly N addition. Eight to ten cylinders, 0–4 cm depth and 2 cm width, were removed from each plot using a soil corer and bulked in a composite sample. Once in the lab, samples were air-dried for several days and stored at room temperature in the dark prior to analysis. Chemical analyses (organic matter, organic and extractable N, total and extractable P, extractable cations and pH) were conducted at the Unit of Analysis of the Centre for Environmental Sciences (CSIC, Madrid, Spain) and have already been published in Ochoa-Hueso et al. (2013b).
2.7 Statistical Analyses
A principal component analysis (PCA) was used to summarize plant community composition (2011) data. Analysis of variance (ANOVA) was used for the effects of simulated N pollution (fixed factor), block (random factor, without interactions) and microsite (fixed factor, only in 2011) on plant biomass production (2010 and 2013), plant community composition (including PCA components, 2011) and diversity (Shannon-Wiener index, Hʹ) and root-surface PME activity (2010). Soil extractable P, assumed to be an important regulator of the ability of N-loving plants to invade formerly nutrient-poor habitats in the context of increased N deposition (Ochoa‐Hueso and Manrique 2014), was tested as a covariate (ANCOVA) in the response of the nitrophytic element to N addition in 2013. Least significant difference tests were used for post-hoc multiple comparisons. Simple linear and nonlinear regressions were used to relate PCA components and root-surface PME activity to soil variables (spring 2011 and 2010, respectively) and simulated N deposition loads. Plant biomass in 2013 was also correlated with selected soil chemical variables. The values used in this analysis, as well as in the covariance analysis, were averages of all sampling campaigns conducted between 2008 and 2011 (Ochoa-Hueso et al. 2013a) and are therefore assumed to accurately represent the average chemical properties of each plot. When necessary, data were transformed as indicated to fit assumptions for normality. Data were analysed using SPSS17.0 (SPSS Inc., Chicago, IL, USA).
3 Results
3.1 Destructive Plant Sampling (2010)
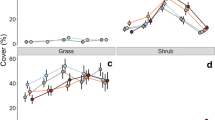
Limonium echioides was consistently reduced by N addition regardless of the N dose applied (Table 1; Fig. 1). Similarly, I. crenata and S. dichotomum showed a nonsignificant trend towards a decrease in biomass with N application, whereas grass biomass tended to increase linearly with N, despite its low contribution to the total plant biomass (Fig. 1). Total plant biomass also tended to increase with N additions, but this effect was again nonsignificant (Fig. 1). When pooled together, the biomass of the three dominant annual plant species that were separately analysed (i.e. L. echioides, S. dichotomum and I. crenata) was reduced by N, whereas the pooled biomass of other forbs (i.e. excluding these three species) increased marginally (P = 0.09) in the 50 kg N ha−1 year−1 treatment (Fig. 1). This effect was largely due to the colonization of the experimental plots by different species of yellow crucifers such as D. siifolia and S. commune and a variety of other species in the Asteraceae familiy (e.g. C. melitensis) from the surrounding areas but without any clear consistent pattern (Table S2).

Nitrogen fertilization effects on total, grass, forb and specific productivity in 2010. Standard error bars are shown. n = 6. Units are biomass (g) per destructive half. Different lower case letters indicate significant differences between N treatments
3.2 Nondestructive Plant Sampling (2011)
Total plant density was reduced by simulated N deposition in spring 2011 (Table 2). In contrast, plant diversity (Hʹ) remained unchanged in response to 3.5 years of N fertilization (Table 2), despite the negative trend observed in the interspaces (data not shown). The PCA with the log-transformed abundance of the ten most common species showed four main components accounting for 75.5 % of the total variance (Table S1). These ten species were: A. linum-stellatum, C. melitensis, H. violaceum, Herniaria hirsuta L., H. petraea, I. crenata, L. echioides, Pistorinia hispanica (L.) DC., Sedum album L. and Teucrium pseudochamaepytis L. The component 4 (accounting for 10.5 % of the total variance; Table S1) was marginally (P = 0.07) affected by simulated N deposition (Table 3; Fig. 2), whereas the rest of the components were unaffected by N (Table 3). Statistically significant differences were between 0–10 and 50 kg N ha−1 year−1. Out of the ten most common species, and following the eigenvalues of the PCA, three species responded negatively to N (A. linum-stellatum, H. violaceum and P. hispanica), whereas six showed no clear response to intermediate doses (L. echioides, C. melilotii, H. hirsuta, H. petraea, I. crenata and S. album) and one showed positive effects of high N doses (T. pseudochamaepytis) (Table S1). The component 1 segregated all these species into two clearly separated groups (Fig. 2) and was positively related to soil fertility; in particular to: soil organic N content (N = 48; R 2 = 0.63; P < 0.01), K availability (N = 48; R 2 = 0.61; P < 0.01), organic matter content (N = 48; R 2 = 0.60; P < 0.01), nitrate availability (N = 48; R 2 = 0.56; P < 0.01), nitrate to ammonium ratio (N = 48; R 2 = 0.48; P < 0.01) and total P (N = 48; R 2 = 0.35; P < 0.01). The component 2 showed a gradient of preference for high Ca availability (N = 48; R 2 = 0.47; P < 0.01) and soil pH (N = 48; R 2 = 0.42; P < 0.01) values. The component 3 was weakly related to Mg availability (N = 48; R 2 = 0.10; P ≤ 0.05) but was highly affected by the presence of shrub cover (Table 3), suggesting that shrub cover, and not Mg, was the main driver of this component. Finally, the component 4 was negatively related to simulated N deposition loads (N = 48; R 2 = 0.13; P = 0.02; Fig. 2) and nitrate availability (N = 48; R 2 = 0.12; P < 0.05).

PCA biplot showing the effects of simulated N deposition on PCA_2011 components. The relative influence of simulated N deposition on each species (grey dots) is also represented as the eigenvalues (see Table 1 for species identity). Standard error bars are shown. n = 12. 0 N = control; 10 N = 10 kg N ha−1 year−1; 20 N = 20 kg N ha−1 year−1; 50 N = 50 kg N ha−1 year−1. Different lower case letters indicate significant differences between N treatments
3.3 Destructive Plant Sampling (2013)
Nitrogen fertilization increased the biomass of yellow crucifers and also their overall biomass per individual (Table 4; Fig. 3). The biomass response was, however, conditioned by the heterogeneous distribution of P availability; i.e. only those plots with higher inorganic P values were able to support increased growth under high N fertilization conditions (Fig. 3). In contrast to the trend observed in 2010, total biomass of I. crenata was also increased by N additions (Table 4). In contrast, plants in the Asteraceae famility did not respond to our fertilization treatments (Table 4), despite the trend towards an increase in density, biomass and biomass per individual (data not shown). Total biomass production of these species was, overall, positively related to nitrate and phosphate availability, nitrate to ammonium ratio and soil organic matter content (Table 5; Fig. 3).

Nitrogen fertilization effects on yellow crucifer biomass in spring 2013 (a). Standard error bars are shown. n = 6. Units are biomass (g) per destructive half. Relationships between yellow crucifer biomass production and P availability per N treatment (b) and yellow crucifer biomass and nitrate in soil (c) are also shown
3.4 Root-Surface PME Activity
There were no significant effects of N treatments on root-surface PME activity of any of the three analysed species (Table 1). Inducible root-surface PME activity greatly varied among species with seedlings of rosemary shrubs (244.7 ± 27.8 (SE) μmol p-nitrophenyl g root−1 min−1) having values ~2-fold higher than those of I. crenata (119.1 ± 12.4 (SE) μmol p-nitrophenyl g root−1 min−1) and ~8-fold higher than those of L. echioides (29.6 ± 5.2 (SE) μmol p-nitrophenyl g root−1 min−1) (Table 1). We found, however, significant relationships between PME activity of R. officinalis roots and available P (negative logarithmic regression: R 2 = 0.23; P = 0.02) and between PME activity of L. echioides roots and total soil P (negative linear regression: R 2 = 0.25; P = 0.04). In addition, there was an inverse relationship between L. echioides biomass production and its PME activity (linear regression: R 2 = 0.35; P = 0.02), suggesting a trade-off between growth and allocation of resources for nutrient uptake. Additional linear relationships between root-surface PME activities and selected soil parameters are shown in Table 6.
4 Discussion
This study has shown some potential impacts of increased N pollution on the native plant community from a European semiarid Mediterranean shrubland. These impacts include species-specific responses in terms biomass production (spring 2010 and 2013) and a significant alteration of the plant community structure (spring 2011). Similarly, previous results (spring 2009) from the same N manipulation experiment had already shown a reduction in the number of individuals of two common annual plant species (L. echioides and A. linum-stellatum) and an overall decrease in rosemary cover in response to N addition (Ochoa-Hueso et al. 2013a). This evidence demonstrates that the negative effects of increased N fertilization on the native annual and woody plant species appeared soon after the initiation of the experiment and also that this type of response (i.e. reduced either plant biomass or density) is consistent throughout the years, although the identity of the species affected may vary between years (L. echioides and A. linum-stellatum in 2009; L. echioides in 2010 and; A. linum-stellatum, H. violaceum and P. hispanica and total annual plant density in 2011) mostly likely due to interaction with varying climatic pressures (Rao et al. 2009; Southon et al. 2013) and the local soil chemistry. We also suggest that these negative effects are primarily caused by increased ammonium availability in sites with formerly hardly any ammonium (Boudsocq et al. 2012; Ochoa-Hueso et al. 2013a). In contrast to these consistent negative effects, the biomass production of the nitrophilous element only increased in response to N in 2010 and 2013, suggesting that between-year variations are important in order to predict responses to N deposition in low-productivity semiarid shrublands (Gutknecht et al. 2012; Ladwig et al. 2012a). Given that the spring of 2010, and particularly of 2013, was the wettest of the 4 years at the study site (59.6, 144.4, 97.1 and 187.7 mm between 1 March and 31 May in 2009, 2010, 2011 and 2013, respectively), we predict that the most negative effects of increased N deposition in terms of plant invasion by N-loving species in semiarid Mediterranean environments will take place during unusually wet years when water limitation is alleviated (Ladwig et al. 2012; Ochoa‐Hueso and Manrique 2014). Responses of the nitrophytic element (particularly crucifers) in 2013 were attributed to high concentrations of nitrate and also dependent on the heterogeneous availability of inorganic P, which supports previous findings that strongly suggested the key role of this particular nutrient in the probability of a given ecosystem to become invaded in response to N deposition (Ochoa‐Hueso and Manrique 2014).
In contrast to our results, only 1 year of N fertilization (40 kg N ha−1) in a Portuguese Mediterranean maquis dominated by early successional summer semi-deciduous plant species (i.e. species that occasionally shed their leaves in summer in response to high temperatures and drought) resulted in ruderal and other herbaceous species growing more in N-fertilized than in control plots (Dias et al. 2011), whereas 2 years of nutrient addition (50 kg N ha−1, 5 kg P ha−1 and a micronutrient solution in a fully factorial combination) in a South-African fynbos showed that herbaceous element, i.e. restioids, graminoids and annuals, was more responsive to N than the remaining woody vegetation in terms of increased productivity (Witkowski 1989). Fertilization with P and other micronutrients also exerted a significant effect on annual plant production of certain functional groups (Witkowski 1989), supporting the role of phosphate and possibly certain micronutrients inter-playing with the extra N and affecting responses to N fertilization (Ochoa-Hueso et al. 2013b). In our study site, soil fertility, calcareousness/acidity of soils and shrub cover explained, in order of importance, a higher percentage of the variation found in the 2011 annual plant community data than simulated N deposition did. However, given that all these variables are susceptible to be altered by increased N deposition (Horswill et al. 2008; Ochoa-Hueso et al. 2013ab), it is predicted that some of the alterations of the plant community found in this study could be mediated, at least partially, by changes in the soil chemistry and the abundance of shrubs (Bobbink et al. 2010; Ochoa-Hueso et al. 2013a). Contrary to our study, alterations of plant community composition in a Californian coastal sage scrub, dominated by summer semi-deciduous species, in response to additions of 50 kg N ha−1 year−1 for 5 years, were only related to changes in the dominant shrubs but not of herbaceous species (Allen et al. 1998; Vourlitis and Pasquini 2009), whereas the same N manipulation design in a Californian chaparral dominated by evergreen sclerophytes, i.e. late successional species, did not result in plant community changes (Vourlitis and Pasquini 2009).
The consistent and negative effects of simulated N deposition on L. echioides biomass production and rosemary cover (this study, Ochoa-Hueso and Manrique 2010, and Ochoa-Hueso et al. 2013a) contrasted with the lack of significant effects of N addition on their root-surface PME activity, a potential functional response of plant species to avoid the negative consequences of an N to P imbalance in a system where P is limiting (Phoenix et al. 2003). This is opposite to results in Phoenix et al. (2003), who found increased PME activity in roots of seedlings of three herbaceous species from temperate calcareous grasslands in the UK. Also opposite to our study, growth of the plant species analysed by Phoenix et al. (2003) was either unaffected or favored by N fertilization. In another study, Pilkington et al. (2008) also found increased root-surface PME activity of the woody species Calluna vulgaris (L.) Hull. with increasing simulated levels of N pollution and the same results of increased root-surface PME in C. vulgaris have been obtained by Kritzler and Johnson (2010). Two of the three species studied here (L. echioides and seedlings of rosemary shrubs) were, in contrast, able to regulate their PME activity depending on soil P status, showing significant relationships between the PME activity and either total or available soil P, respectively. Other studies in Calluna heathlands have also found an inverse relationship between root-surface PME activity and extractable P (Johnson et al. 1999). The different reliance on P forms as an environmental clue between the annual plant and the shrub species could be tentatively attributed to the different microenvironment occupied by these plants and, finally, to the possibility of resource partitioning for soil P (Turner 2008). Whereas L. echioides usually lives isolated in the interspaces between shrubs, where inputs of labile organic matter are low, the microenvironment generated by the mother plant on rosemary seedlings is favorable for an active nutrient cycling, and therefore they may have access to a more reliable source of P. Whether the responses found are related to mycorrhizal, microbial or direct root secretion has not been investigated here, but the question of the different degrees of response of these three mechanisms to simulated N pollution remains as an interesting area of research for the future.
5 Conclusions
In this study, we have shown that the biomass and density of certain common annual (e.g. L. echioides) and chamaephytic (H. violaceum) plant species found in semiarid shrublands from central Spain had decreased after ~2.5 and ~3.5 years of experimental N manipulation and that this response translated into significant alterations of the local annual plant community. Therefore, we predict that the increasing threat of N deposition will cause a shift in local plant communities from semiarid Mediterranean ecosystems that will be mainly associated with a species-specific decline of local forb species and tentatively attribute this response to an overall increase in the nitrophilous element (particularly crucifers) and to high levels of soil ammonium. However, we did not find any evidence of potential impacts of an induced N to P imbalance, as suggested by the lack of up-regulation of root-surface PME activities. Finally, given the evident implication of P status of soils in terms of probability of ecosystem invasion by nitrophilous plants and grasses (Ceulemans et al. 2013; Fanelli et al. 2008; Ochoa‐Hueso and Manrique 2014) and in the response of plant communities to increased N deposition, future experiments of P addition will help to disentangle the role of this scarce nutrient in the impacts of N deposition on relatively polluted semiarid Mediterranean-type ecosystems.
References
Aber, J. D., Goodale, C. L., Ollinger, S. V., Smith, M.-L., Magill, A. H., Martin, M. E., Hallett, A., & Stoddard, J. L. (2003). Is nitrogen deposition altering the nitrogen status of Northeastern forests? Bioscience, 53, 375–389.
Allen, E. B., Padgett, P. E., Bytnerowicz, A., & Minnich, R. A. (1998). Nitrogen deposition effects on coastal sage vegetation of southern California (pp. 131–140). In: Proceedings of the International Symposium on Air Pollution and Climate Change Effects on Forest Ecosystems. US Department of Agriculture Forest Service Pacific Southwest Research Station, Riverside, California, USA.
Bobbink, R., Hicks, K., Galloway, J., Spranger, T., Alkemade, R., Ashmore, M., Bustamante, M., Cinderby, S., Davidson, E., Dentener, F., Emmet, B., Erisman, J.-W., Fenn, M., Gilliam, F., Nordin, A., Pardo, L., & De Vries, W. (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl, 20, 30–59.
Bonanomi, G., Caporaso, S., & Allegrezza, M. (2006). Short-term effects of nitrogen enrichment litter removal and cutting on a Mediterranean grassland. Acta Oecol, 30, 419–425.
Boudsocq, S., Niboyet, A., Lata, J. C., Raynaud, X., Loeuille, N., Mathieu, J., et al. (2012) Plant preference for ammonium versus nitrate: a neglected determinant of ecosystem functioning?. The American Naturalist 180(1), 60–69.
Carfrae, J. A., Sheppard, L. J., Raven, J. A., Leith, I. D., & Crossley, A. (2007). Potassium and phosphorus additions modify the response of Sphagnum capillifolium growing on a Scottish ombrotrophic bog to enhanced nitrogen deposition. Appl Geochem, 22, 1111–1121.
Ceulemans, T., Merckx, R., Hens, M., & Honnay, O. (2013). Plant species loss from European semi natural grasslands following nutrient enrichment–is it nitrogen or is it phosphorus?. Global Ecology and Biogeography, 22(1), 73–82.
Cox, R. D., & Allen, E. B. (2008). Composition of soil seed banks in southern California coastal sage scrub and adjacent exotic grassland. Plant Ecol, 198, 37–46.
Cruz, C., Bio, A. M. F., Jullioti, A., Tavares, A., Dias, T., & Martins-Loução, M. A. (2008). Heterogeneity of soil surface ammonium concentration and other characteristics related to plant specific variability in a Mediterranean-type ecosystem. Environ Pollut. doi:10.1016/jenvpol200712071–10.
Dias, T., Malveiro, S., Martins-Loução, M. A., Sheppard, L. J., & Cruz, C. (2011). Linking N-driven biodiversity changes with soil N availability in a Mediterranean ecosystem. Plant and soil, 341(1-2), 125–136.
Egerton-Warburton, L. M., & Allen, E. B. (2000). Shifts in arbuscular mycorrhizal communities along an anthropogenic nitrogen deposition gradient. Ecological Applications, 10(2), 484–496.
Emmett, B. A. (2007). Nitrogen saturation of terrestrial ecosystems: some recent findings and their implications for our conceptual framework. Water Air Soil Pollut: Focus, 7(1–3), 99–109.
Fanelli, G., Lestini, M., & Sauli, A. S. (2008). Floristic gradients of herbaceous vegetation and P/N ratio in soil in a Mediterranean area. Plant Ecol, 194, 231–242.
Fenn, M. E., Baron, J. S., Allen, E. B., Rueth, H. M., Nydick, K. R., Geiser, L., Bowman, W. D., Sickman, J. O., Meixner, T., Johnson, D. W., & Neitlich, P. (2003). Ecological effects of nitrogen deposition in the Western United States. Bioscience, 53, 404–420.
Fenn, M. E., Allen, E. B., Weiss, S. B., Jovan, S., Geiser, L. H., Tonnesen, G. S., Johnson, R. F., Rao, L. E., Gimeno, B. S., Yuan, F., Meixner, T., & Bytnerowicz, A. (2010). J Environ Manag, 91, 2404–2423.
Fisher, J., Lorenagan, W. A., Dixon, K., & Veneklaas, E. J. (2009). Soil seed bank compositional change constrains biodiversity in an invaded species-rich woodland. Biol Conserv, 142, 256–29.
Gallardo, A., Rodríguez-Saucedo, J. J., Covelo, F., & Fernández-Alés, R. (2000). Soil nitrogen heterogeneity in a Dehesa ecosystem. Plant Soil, 222, 71–82.
Gutknecht, J. L., Field, C. B., & Balser, T. C. (2012). Microbial communities and their responses to simulated global change fluctuate greatly over multiple years. Global Change Biology, 18(7), 2256–2269.
Hobbs, R. J., & Richardson, D. M. (1995). Mediterranean-type ecosystems: opportunities and constraints for studying the function of biodiversity (pp. 1–42). In: Davis, G.W., & Richardson, D. Mediterranean-type ecosystems. The function of biodiversity. Springer-Verlag, Berlin, Heidelberg.
Horswill, P., O’Sullivan, O., Phoenix, G. K., Lee, J. A., & Leake, J. R. (2008). Base cation depletion eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition. Environ Pollut, 155, 336–349.
Johnson, D., Leake, J. R., & Lee, J. A. (1999). The effects of quantity and duration of simulated pollutant nitrogen deposition on root-surface phosphatase activities in calcareous and acid grasslands: a bioassay. New Phytol, 141, 433–442.
Kritzler, U. H., & Johnson, D. (2010). Mineralisation of carbon and plant uptake of phosphorus from microbially-derived organic matter in response to 19 years of simulated nitrogen deposition. Plant Soil, 326, 311–319.
Ladwig, L. M., Collins, S. L., Swann, A. L., Xia, Y., Allen, M. F., & Allen, E. B. (2012). Above-and belowground responses to nitrogen addition in a Chihuahuan Desert grassland. Oecologia, 169(1), 177–185.
Ochoa-Hueso, R., & Manrique, E. (2010). Nitrogen fertilization and water supply affect germination and plant establishment of the soil seed bank present in a semi-arid Mediterranean scrubland. Plant Ecol, 210, 263–273.
Ochoa-Hueso, R., & Manrique, E. (2011). Effects of nitrogen deposition and soil fertility on cover and physiology of Cladonia foliacea (Huds) Willd a lichen of biological soil crusts from Mediterranean Spain. Environ Pollut, 159, 449–457.
Ochoa‐Hueso, R., & Manrique, E. (2014). Impacts of altered precipitation, nitrogen deposition and plant competition on a Mediterranean seed bank. J Veg Sci. doi:10.1111/jvs.12183.
Ochoa-Hueso, R., Allen, E. B., Branquinho, C., Cruz, C., Dias, T., Fenn, M. E., Manrique, E., Pérez-Corona, M. E., Sheppard, L. J., & Stock, W. D. (2011a). Nitrogen deposition effects on Mediterranean-type ecosystems: an ecological assessment. Environ Pollut. doi:10.1016/j.envpol.2010.12.019.
Ochoa-Hueso, R., Hernandez, R. R., Pueyo, J. J., & Manrique, E. (2011b). Spatial distribution and physiology of biological soil crusts from semi-arid central Spain are related to soil chemistry and shrub cover. Soil Biol Biochem. doi:10.1016/j.soilbio.2011.05.010.
Ochoa-Hueso, R., Pérez-Corona, M. E., & Manrique, E. (2013a). Impacts of simulated N deposition on plants and mycorrhizae from Spanish semiarid Mediterranean shrublands. Ecosystems, 16, 838–851.
Ochoa-Hueso, R., Stevens, C. J., Ortiz-Llorente, M. J., & Manrique, E. (2013b). Soil chemistry and fertility alterations in response to N application in a semiarid Mediterranean shrubland. Sci Total Environ, 452–453, 78–86.
Ochoa-Hueso, R., Arróniz-Crespo, M., Bowker, M. A., Maestre, F. T., Pérez-Corona, M. E., Theobald, M. R., Vivanco, M. G., & Manrique, E. (2014). Biogeochemical indicators of elevated nitrogen deposition in semiarid Mediterranean ecosystems. Environ Monit Assess, 186, 5831–5842.
Phoenix, G. K., Booth, R. E., Leake, J. R., Read, D. R., Grime, J. P., & Lee, J. A. (2003). Simulated pollutant nitrogen deposition increases P demand and enhances root-surface phosphatase activities of three plant functional types in a calcareous grassland. New Phytol, 161, 279–289.
Phoenix, G. K., Hicks, W. K., Cinderby, S., Kuylenstierna, J. C. I., Stock, W. D., Dentener, F. J., Giller, K. E., Austin, A. T., Lefroy, R. D. B., Gimeno, B. S., Ashmore, M. S., & Ineson, P. (2006). Atmospheric nitrogen deposition in world biodiversity hotspots: the need for a greater global perspective in assessing N deposition impacts. Glob Chang Biol, 12, 470–476.
Pilkington, M. G., Caporn, S. J. M., Carroll, J. A., Cresswell, N., Lee, J. A., Ashenden, T. W., Brittain, S. A., & Turner, B. L. (2008). Resource partitioning for soil phosphorus: a hypothesis. J Ecol, 96, 698–702.
Rao, L. E., Parker, D. R., Bytnerowicz, A., & Allen, E. B. (2009). Nitrogen mineralization across an atmospheric nitrogen deposition gradient in Southern California deserts. Journal of arid environments, 73(10), 920–930.
Rivas-Martínez, S. (1987) Memoria del mapa de series de vegetación de España. ICONA, Madrid.
Southon, G. E., Field, C., Caporn, S. J., Britton, A. J., & Power, S. A. (2013). Nitrogen deposition reduces plant diversity and alters ecosystem functioning: field-scale evidence from a nationwide survey of UK heathlands. PLoS one,8(4), e59031.
Stevens, C. J., Duprè, C., Dorland, E., Gaudnik, C., Gowing, D. J., Bleeker, A., et al. (2011). The impact of nitrogen deposition on acid grasslands in the Atlantic region of Europe. Environmental Pollution, 159(10), 2243–2250.
Turner, B. L. (2008). Resource partitioning for soil phosphorus: a hypothesis. Journal of Ecology, 96(4), 698–702.
Vourlitis, G. L., & Pasquini, S. C. (2009). Experimental dry-season N deposition alters species composition in southern Californian Mediterranean-type shrublands. Ecology, 90, 2183–2189.
Witkowski, E. F. T. (1989). Response to nutrient additions by the plant growth forms of sand-plain lowland fynbos South Africa. Vegetatio, 79, 89–97.
Acknowledgments
This research was financially supported by the Spanish Ministerio de Educación y Ciencia (CGL-2009-11015 and REMEDINAL-2 S2009/AMB-1783). ROH was also funded by an FPU fellowship (AP2006-04638). CJS was funded by a Leverhulme Early Career Fellowship, the European Science Foundation Nitrogen in Europe programme and the Percy Sladen Memorial Fund. We are very thankful to Octavio Cedenilla, Daniel de la Puente, Cristina Paradela and Christopher Long for helping with the lab and field work. Prof. Alfredo Polo and Dr. J.C. García-Gil are thanked for kindly allowing us to use their spectrophotometer and enzyme substrates. Dr. Jason Kaye provided helpful comments on an earlier version of the manuscript. Finally, we want to specially mention Prof. Esteban Manrique and Dr. M. Esther Pérez-Corona for their personal involvement within this project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ochoa-Hueso, R., Stevens, C.J. European Semiarid Mediterranean Ecosystems are Sensitive to Nitrogen Deposition: Impacts on Plant Communities and Root Phosphatase Activity. Water Air Soil Pollut 226, 5 (2015). https://doi.org/10.1007/s11270-014-2278-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-014-2278-1