
Abstract
Plant growth-promoting rhizobacteria (PGPR) play an important role in the biodegradation of natural and xenobiotic organic compounds in soil. They can also alter heavy metal bioavailability and contribute to phytoremediation in the presence or absence of synthetic metal chelating agents. In this study, the inhibitory effect of Cd2+ and Ni2+ at different concentrations of Ca2+ and Mg2+, and the influence of the widely used chelator EDTA on growth of the PGPR Pseudomonas brassicacearum in a mineral salt medium with a mixture of four main plant exudates (glucose, fructose, citrate, succinate) was investigated. Therefore, the bacteriostatic effect of Cd2+, Ni2+ and EDTA on the maximum specific growth rate and the determination of EC50 values was used to quantify inhibitory impact. At high concentrations of Ca2+ (800 μmol L-1) and Mg2+ (1,250 μmol L-1), only a small inhibitory effect of Cd2+ and Ni2+ on growth of P. brassicacearum was observed (EC50 Cd2+, 18,849 ± 80 μmol L−1; EC50 Ni2+, 3,578 ± 1,002 μmol L−1). The inhibition was much greater at low concentrations of Ca2+ (25 μmol L−1) and Mg2+ (100 μmol L−1) (EC50 Cd2+, 85 ± 0.5 μmol L−1 and EC Ni2+, 62 ± 1.8 μmol L−1). For the chosen model system, a competitive effect of the ions Cd2+ and Ca2+ on the one hand and Ni2+ and Mg2+ on the other hand can be deduced. However, the toxicity of both, Cd2+ and Ni2+, could be significantly reduced by addition of EDTA, but if this chelating agent was added in stoichiometric excess to the cations, it also exhibited an inhibitory effect on growth of P. brassicacearum.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The rhizosphere is the root–soil contact zone which is most important for the regulation of soil organic matter decomposition and nutrient cycling, particularly due to the biodegradation of natural and xenobiotic organic compounds. It is a highly dynamic area for interactions between plant roots and, among others, beneficial soil microbes such as plant growth-promoting rhizobacteria (PGPR). One of the most important natural influences on microbial communities in the rhizosphere is the release of a wide range of various compounds by plant roots, referred to as exudation (Lugtenberg et al. 1999) or rhizodeposition (Costa 2006). Growing plants release up to 40% of assimilated carbon as exudates into the rhizosphere (Liljeroth et al. 1994; Neumann and Römheld 2000; Kumar et al. 2006). This results in highly active and diverse heterotrophic microbial populations with bacterial densities 10 to 200 times higher than in surrounding bulk soil. In turn, PGPR essentially contribute to plant growth and health. Direct beneficial effects are, for example, biofertilisation (e.g., solubilising mineral nutrients), production of phytohormones, stimulation of root growth, rhizoremediation and enhancing plant stress resistance (Lynch 1990; Bowen et al. 1999; Cook 2000; Jing et al. 2007; Lugtenberg and Kamilova 2009).
The contamination of ecosystems by heavy metals, in particular, soils used for agriculture, represents one of the long-standing major environmental problems due to their prevalent existence and potential toxicity to plants, microorgansims, animals and humans. In this study, the term “heavy metal” refers to the definition of Passow et al. (1961). They classify elements according to their density, in which heavy metals show a density greater than 5 g cm−3.
One of the main roles of rhizobacteria in the phytoremediation of heavy metal-contaminated soil using hyperaccumulating plants is the increased heavy metal mobility and availability to the plant caused by the release of naturally occurring chelating agents, acidification, phosphate solubilization and redox changes (Lynch 1990; Bowen et al. 1999; Whiting et al. 2001; Abou-Shanab et al. 2003; Kamnev 2003; Jing et al. 2007; Zhuang et al. 2007; Khan et al. 2009). An additional increase of heavy metal availability to the plant can be achieved by the addition of a synthetic chelating agent such as EDTA to the contaminated soil. Consequently, chelator-enhanced phytoremediation of heavy metal contaminated soil has gained considerable attention within the last two decades (Alkorta et al. 2004; Lestan and Finzgard 2007). For example, Chen and Cutright (2002) showed that inoculation of sunflower seeds and surrounding soil with heavy metal-resistant rhizosphere bacteria caused a significant increase of heavy metal phytoremediation in the presence of EDTA. The exposure to EDTA without inoculation with rhizobacteria caused an increased specific bioaccumulation, but also a reduced formation of biomass. In the presence of the rhizobacteria, however, a high biomass yield together with an increased accumulation of cadmium, chrome or nickel was observed.
Nevertheless, the multiple and complex interactions between rhizobacteria, heavy metal ions and synthetic chelating agents are still not fully investigated and understood to date. In addition, many rhizobacteria, including xenobiotics-degrading bacteria, are very sensitive to heavy metals (Duxbury 1985; Baath 1989; Jansen et al. 1994; McGrath et al. 1995; Giller et al. 1998; Oliveira and Pampulha 2006; Giller et al. 2009). Amongst other reasons, this toxicity is due to the displacement of essential macroelements such as Ca2+ and Mg2+ from biomolecules, and result in a substantial reduction of viability, activity, diversity and density of microbial rhizosphere populations (Pennanen et al. 1996; Giller et al. 1998; Sandaa et al. 1999; Liao et al. 2005). As the contamination of soils with heavy metals represents one of the long-standing major environmental problems, a better understanding of the manifold interactions between plants, rhizobacteria, heavy metals, essential macroelements and additional synthetic chelating agents is required.
Strains of Pseudomonas brassicacearum represent an interesting group of PGPR which belong to the major root zone colonisers of Arabidopsis thaliana and Brassica napus (Achouak et al. 2000; Achouak et al. 2004). In this study, P. brassicacearum (type strain CFBP 11706) was chosen as a representative and attractive model rhizobacterium to obtain a better understanding of the interactions between rhizobacteria, metal ions and synthetic chelating agents by investigating the inhibitory effect of Cd2+ and Ni2+, two of the environmentally most important and hazardous heavy metals, and in return, a possible protective influence of Ca2+, Mg2+ and EDTA.
2 Material and Methods
2.1 Strain and Culture Conditions
P. brassicacearum (CFBP 11706) was obtained as a lyophilisate from the “German Collection of Microorganisms and Cell Cultures” (DSMZ) with DSM number 13227.
It was grown on a mineral medium based on the four characteristic plant exudates glucose, fructose (Sigma), citrate (Merck) and succinate (Fluka Biochemika), each at 2 mmol L−1 (so-called 4C medium). Additional components were 6 mmol L−1 (NH4)2SO4, 0.8 mmol L−1 CaCl2, 1.25 mmol L−1 MgSO4, 60 μmol L FeCl3, 1 ml L−1 trace element solution SL6 (according to Pfennig (1974)), and 25 ml L−1 of 1 mol L−1 phosphate buffer (as reported by Pack (1996)). The pH value was adjusted to 7.5, which was identified to be optimal for bacterial growth in preliminary experiments.
Precultivations of P. brassicacearum were conducted in a tenfold diluted tryptic soy broth (TSB) medium (as described by Achouak et al. 2000). Typically, these cultivations were performed in 1,000-mL shaking flasks containing 200 mL of TSB/10 medium. The medium was inoculated with 2 mL of P. brassicacearum cryo-culture and incubated overnight (12 h) at 30°C and 120 min−1. After that, 180 mL of sterile 4C medium (1,000-mL shaking flasks) were inoculated with 20 mL of the preculture. Ni2+, Cd2+ and/or EDTA were added to culture medium directly after inoculation. Cultivations continued at 30°C with a shaking rate of 120 min−1. Bacterial growth was monitored by measuring absorbance at 546 nm, which was adopted from experiments described in literature (Belimov et al. 2007)
Screening experiments were carried out in 96-well microtiter plates containing 200-μL 4C medium per well. For this purpose, 180 mL sterile 4C medium was prepared in 1,000-mL shaking flasks. Subsequently, shaking flasks (TSB/10) were inoculated with 20 ml of the preculture, and 200 μL of this culture were transferred to microtiter wells. Cultivations in 96-well microtiter plates were performed using the microplate reader Sunrise (Tecan). Contaminations could be avoided by sterilisation of the microtiter plates with UV light for 20 min before adding culture medium. Cycles of the microplate reader were adjusted to 15-min measurement intervals and medium agitation intensity in the meantime. Absorbance measurements were conducted at 620 nm. This wavelength differs from the one used in the shaking-flask experiments due to the fact that the microtiter plate reader used in this study only supports certain wavelengths. However, P. brassicacearum measurements showed almost identical values at both wavelengths. All cultivations were performed as triplicates of at least two independent experiments. The respective maximum specific growth rate was evaluated by plotting logarithmical optical density values versus time, followed by linear regression of the data from the exponential growth phase. The slope of this regression line is referred to as maximum specific growth rate μmax [per hour]. It was used as characteristic parameter giving hints on bacteriostatic effects of Cd2+ and Ni2+ on bacterial growth.
2.2 Analytics
After filtration of the samples using 0.2 μm sterile filters, sugars and organic acids in the medium were analysed by High-Performance Liquid Chromatography (HPLC) using a VWR-Hitachi LaChrom Elite system. The HPLC system was equipped with an UV- (detection wavelength 210 nm) and an RI-detector. A Varian Metacarb 87C column was utilised for determination of glucose and fructose concentrations, a Varian Metacarb 67H column served for analysis of citrate and succinate. Sulphuric acid (15 mmol L−1) was used as mobile phase in an isocratic HPLC method. Additionally, glucose was quantified with an YSI 2700 Select Biochemistry Analyzer, allowing quick analysis of cultivation samples with identical concentrations in comparison to the HPLC analyses (data not shown).
2.3 Determination of EC50 Values
In toxicology, the half-maximal effective concentration (EC50) is defined as the concentration of a toxicant that leads to a response halfway between the baseline and the maximum effect. To determine the EC50 value, the logarithmical value of the toxicant concentrations added to the cultivations was plotted against the respective maximum specific growth rate of P. brassicacearum (determined as described in Section “Strain and Culture Conditions”). Afterwards, a linear regression of the data points was performed. From the regression line's equation, the logarithmical concentration of toxicants could be calculated which lead to a half-maximal specific growth rate. From these data, the EC50 values can be derived.
3 Results
3.1 Medium Development and Determination of Kinetic Parameters KS- [mmol L−1] and μmax [per hour].
Sugar and organic acid exudates provide the main carbon and energy sources for rhizosphere colonising microorganisms (Lugtenberg et al. 1999). Prior to the experiments with the 4C medium containing the four characteristic plant exudates glucose, fructose, citrate and succinate, the kinetic parameter KS [millimoles per litre] and μmax [per hour] for growth of P. brassicacearum with each of these compounds (0–2.5 mmol L−1) as a sole source of carbon and energy were determined. The results clearly displayed a Monod-like growth kinetic. Significant differences in the kinetic data for the single components were observed. The maximum growth rates were, in descending order: succinate (0.4 ± 0.04 h−1) > glucose (0.36 ± 0.03 h−1) > fructose (0.18 ± 0.01 h−1) > citrate (0.1 ± 0.01 h−1). Substrate affinity (given by the KS value) increased in reverse order: succinate (0.64 ± 0.13 mmol L−1) < glucose (0.24 ± 0.06 mmol L−1) < fructose (0.1 ± 0.03 mmol L−1) < citrate (0.09 ± 0.03 mmol L−1). At an initial substrate concentration of 2 mmol L−1, growth with each of the single-carbon sources occurred approximately with the respective maximum rate, and so a model 4C medium with 2 mmol L−1 of each of the plant exudates was chosen for the further investigations.
3.2 Influence of Cd2+ and Ni2+ on Growth of P. brassicacearum in a Standard Mineral Medium
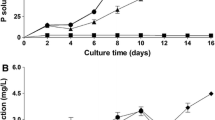
In a previous study with cells of P. brassicacearum strain NFM421, Pagès et al. reported a maximum tolerable Cd2+ concentration of 25 μmol L−1 when using a tenfold diluted TSB/10 medium for the cultivations (Pages et al. 2007). In the present study, however, the synthetic mineral 4C medium was used to specifically investigate the influence of Cd2+ and Ni2+ on growth of P. brassicacearum (type strain CFBP 11706) under various defined conditions. In the first experiments, the mineral salt composition corresponded to that of typical mineral media used for growth experiments; it contained, amongst others, 1.25 mmol L−1 of MgSO4 and 0.8 mmol L−1 of CaCl2 (see Material and Methods), and the concentrations of Cd2+ or Ni2+ were 0 μmol L−1 or adjusted to 100 μmol L−1 and 500 μmol L−1, respectively. Under these conditions, only a slight growth inhibition even at relatively high concentrations of 500 μmol L−1 of Cd2+ (EC50 15,849 ± 80 μmol L−1) or Ni2+ (EC50 3,578 ± 1002 μmol L−1) was observed (Fig. 1a and b). Figure 1 also exhibits a lower maximum optical density in presence of heavy metal ions, but however, in comparison to the high amount of metal ions, this effect is relatively small. Since the inhibitory or toxic effect of heavy metal ions might be caused by their competition with essential macroelements, it was assumed, that the high concentrations of Mg2+ and Ca2+ in the 4C medium could alleviate the toxicity of Cd2+ and Ni2+ based on similar ion radii of Mg2+ (72 pm) and Ni2+ (69 pm), and of Ca2+ (100 pm) and Cd2+ (95 pm) (Lide 1994).

Growth curves of P. brassicacearum based on synthetic mineral salt medium 4C in the presence of: a 0, 100 or 500 μmol L−1 CdCl2; 1,250 μmol L−1 MgSO4 and 800 μmol L−1 CaCl2. b 0, 100 or 500 μmol L−1 NiCl2; 1,250 μmol L−1 MgSO4 and 800 μmol L−1 CaCl2
3.3 Influence of Cd2+ and Ni2+ on Growth of P. brassicacearum at Low Mg2+ and Ca2+ Concentrations
To examine, if the given concentrations of Mg2+ and Ca2+ might have significantly influenced the inhibitory effect of Cd2+ or Ni2+ on the growth of P. brassicacearum, the Mg2+ and Ca2+ concentrations in 4C medium were lowered to a minimum level, which still allowed growth of the microorganisms at the maximum rate. This required a screening experiment which is performed in microtiter cultivations using the standard 4C medium with different concentrations of MgSO4 (0–1 mmol L−1) and CaCl2 (0–0.8 mmol L−1). The results of this experiment are depicted in Fig. 2. As expected, no bacterial growth is observed in the absence of the essential elements Mg2+ and Ca2+. Additionally, the respective maximum growth rates indicate a higher dependency from Mg2+ concentration rather than from Ca2+ concentrations while remaining almost constant even at low concentrations of CaCl2, it declines rapidly at decreased MgSO4 concentrations. As a consequence, the initial concentrations of MgSO4 and CaCl2 in the 4C medium were reduced from 1.25 mmol L−1 to 100 μmol L−1 (MgSO4) and from 0.8 mmol L−1 to 25 μmol L−1 (CaCl2) ensuring an only slightly reduced maximum specific growth rate of approximately 0.4 h−1 (Fig. 2).

Dependency of μmax [per hour] on Mg2+ and Ca2+ concentration, 100 μmol L−1 MgSO4 and 25 μmol L−1 CaCl2 present in 4C medium ensure a maximum specific growth rate of 0.4 h−1
At these lower concentration of Mg2+ (100 μmol L−1) and Ca2+ (25 μmol L−1) in the 4C medium, a strong inhibitory effect of Cd2+ (EC50 85 ± 0.5 μmol L−1) and Ni2+ (EC50 62 ± 1.8 μmol L−1) on the maximum growth rate of P. brassicacearum was observed (Fig. 3a and b). While in the previous experiment at high Mg2+ and Ca2+ concentrations in the medium, inhibition was almost negligible at heavy metal concentrations of 100 μmol L−1 (see Fig. 1), a similar concentration at low Mg2+ and Ca2+ contents led to a dramatic decrease of bacterial growth in both experimental setups. In addition, Fig. 3 and the EC50 values derived from this experiment (given in Table 1) indicate a higher toxicity of Ni2+ compared to Cd2+.

Growth of P. brassicacearum based on mineral salt medium 4C in the presence of: a 0, 25, 75, 125 or 250 μmol L−1 CdCl2; 100 μmol L−1 MgSO4 and 25 μmol L−1 CaCl2. b 0, 25, 75, 125 or 250 μmol L−1 NiCl2; 100 μmol L−1 MgSO4 and 25 μmol L−1 CaCl2
3.4 Combined Effect of Cd2+ and Ni2 on Growth of P. brassicacearum at Low Mg2+ and Ca2+ Concentrations
Plants and soil microorganisms growing in heavy metal contaminated soils are usually exposed to mixtures of different pollutants. After studying the growth inhibiting impact of Cd2+ or Ni2+ separately, the combined effect of these metals at different particular concentrations is investigated. The curve progression of Fig. 4 shows a sharp decline of μmax [per hour] with increasing concentrations of Cd2+ and Ni2+ present in 4C medium. Bacterial growth of P. brassicacearum was strongly inhibited by equimolar mixtures of Ni2+ and Cd2+ (EC50 Cd2+/Ni2+ 40 ± 0.4 μmol L−1). The combined toxicity effect of Cd2+ and Ni2+ on μmax values [per hour] of P. brassicacearum can therefore be described as additive. The course of isolines illustrates again the higher toxicity of Ni2+ towards the bacteria than the same concentration of Cd2+.

Combined effect of heavy metal toxicity on μmax [per hour] of P. brassicacearum 4C medium in presence of CdCl2 and NiCl2; Ni2+ (75 μmol L−1): μmax = 0.24 h−1; Cd2+ (75 μmol L−1): μmax = 0.32 h−1; Ni2+ and Cd2+ (each 75 μmol L−1): μmax, combined = 0.14 h−1; reference (no heavy metal) μmax = 0.53 h−1
3.5 Effect of EDTA in the Absence or Presence of Cd2+ or Ni2+, Respectively
As EDTA is widely used in remediation processes for heavy metal contaminated soils (Maki et al. 2009), the effect of EDTA on Cd2+ and Ni2+ toxicity for growth of P. brassicacearum was examined. For this purpose, screening experiments in microtiter scale were carried out using the developed 4C medium with low concentrations of Ca2+ and Mg2+, varying concentrations of Cd2+ or Ni2+ (0–200 μmol L−1), respectively, and of non-complexed EDTA (0–1,000 μmol L−1). As indicated by Fig. 5a and b, no significant inhibitory of Cd2+ or Ni2+ was observed when EDTA was present at respective stoichiometric concentrations (EC50 Cd2+/EDTA 19,953 ± 1,852 μmol L−1; EC50 Ni2+/EDTA 4,148 ± 616 μmol L−1). At equimolar concentrations of EDTA and Cd2+ (Fig. 5a) or Ni2+ (Fig. 5b), the maximum growth rates were in the range between 0.4 and 0.53 h−1, independent of the respective heavy metal concentration. This indicates that the heavy metals are completely masked by the formation of strong and non-inhibitory EDTA chelates. However, not only an excess concentration of either heavy metal ions (EC50 Cd2+ 85 ± 0.5 μmol L−1; EC50 Ni2+ 62 ± 1.8 μmol L−1, see Fig. 5 along x-coordinate) but also of non-complexed EDTA (EC50 EDTA 116 ± 4.6 μmol L−1, see Fig. 5 along y-coordinate) resulted in decreasing maximum specific growth rates of P. brassicacearum.

Influence of chelating agent EDTA on heavy metal toxicity: a Variation of CdCl2 (0–200 μmol L−1) and EDTA (0–1,000 μmol L−1). b Variation of NiCl2 (0–200 μmol L−1) and EDTA (0–1,000 μmol L−1)
Table 1 summarises the EC50 values which could be obtained by graphical determination. As discussed before it becomes obvious, that high concentrations of either Mg2+ or Ca2+ significantly reduce toxicity of the heavy metal ions Cd2+ and Ni2+ towards P. brassicacearum. The aforementioned higher impact of nickel in comparison to cadmium also is described in this table, while the addition of both heavy metal ions results in an increased effect in comparison to addition of only one heavy metal species. Additionally, the EC50 values for the experiments containing EDTA are depicted in Table 1.
In Table 2 some physicochemical properties of the considered metal ions and observations concerning toxicity are listed. The toxic effects are subdivided into three classes: absence of EDTA, equimolar concentrations and excess of EDTA. Each class showed different impact on bacterial growth.
The similar ion radii of Ni2+ and Mg2+ on the one hand and Cd2+ and Ca2+ on the other hand (see Table 2) as well as similar ionic indices lead to the conclusion that toxic effects are based on competitive interactions. The comparably high complexation constants (log KB) of heavy metal ions with EDTA explain the fact that equimolar concentrations of heavy metal ions and EDTA result in almost no inhibition effect on P. brassicacearum: heavy metal complexation is preferred compared to complexation of Ca2+ and Mg2+. The toxicity of both compounds is masked due to the complexation.
4 Discussion and Conclusions
The present study focuses on the interactive effects of environmentally important heavy metals (Cd2+ and Ni2+), essential macroelements (Ca2+ and Mg2+) and a synthetic metal chelating agent (EDTA) on growth of a representative PGPR (P. brassicacearum) in mineral salts media. Complex growth media, soil waters and sediment pore waters can contain a wide variety of dissolved inorganic and organic constituents which may drastically alter the speciation of metals in solutions and thus mask the effective interactions between heavy metals, essential cations and additional chelators. In order to avoid or minimise such undesired interactions, the investigations are carried out under defined conditions in liquid cultures with mineral salts media containing either one or all of four plant exudate compounds (glucose, fructose, succinate and citrate, each 2 mmol L−1).
Although laboratory conditions do not adequately represent field settings, they are necessary to gain a better understanding of the influence of major essential cations and chelating agents on the toxicity of heavy metals towards rhizobacteria. Independent of the actual chemical speciation of Cd2+ and Ni2+ in the mineral media used for this study, the defined and comparable conditions should thus supply significant information about the interactive influence of Ca2+ and Mg2+ at different concentrations as well as of EDTA on growth of P. brassicacearum and on the inhibitory effect caused by Cd2+ and Ni2+. Since the toxic effect of Cd2+ on P. brassicacearum has been characterised as a bacteriostatic inhibition of growth, but not as a bacteriocidal inactivation (Pages et al. 2007), the respective maximum specific growth rate μmax [per hour] of this organism is used as an indicator for the magnitude of toxicity caused by Cd2+, Ni2+ and EDTA.
Assuming that the displacement of essential metal ions from biomolecules by heavy metal ions does not necessarily result in the inactivation of the biomolecule, the inhibitory effect of Cd2+ and Ni2+ on growth of P. brassicacearum should thus, at least in part, be based on their competition with Ca2+ and Mg2+. According to this, the respective concentrations of Ca2+ and Mg2+ are supposed to have an important influence on the heavy metal toxicity. This assumption is confirmed by comparative investigations which were carried out using two different growth media which contained the same components but a different concentration of Ca2+ and Mg2+, respectively. When a mineral salt medium containing 1.25 mmol L−1 of Mg2+ and 0.8 mmol L−1 of Ca2+ is used, only a minor inhibitory effect of Cd2+ and Ni2+ on growth of P. brassicacearum is observed even at a high concentration of 500 μmol L−1 of Cd2+ (EC50 15,849 ± 80 μmol L−1) or Ni2+ (EC50 3.578 ± 1,002 μmol L−1). In contrast, at minimum concentrations of Mg2+ (100 μmol L−1) and Ca2+ (25 μmol L−1) which still enable growth at a maximum rate in the absence of heavy metals, an expected strong toxic effect is detected (EC50 Cd2+ 85 ± 0.5 μmol L−1; EC50 Ni2+ 62 ± 1.8 μmol L−1). This is in accordance with other studies describing a protective effect of Ca2+ or Mg2+ (respectively, the water hardness) against heavy metal toxicity on pro- and eukaryotic organisms (Aimal and Khan 1984; Hong et al. 1995; Heijerick et al. 2003; Slaveykova et al. 2009). Although it is known that the toxicity may be reduced if dissolved cations such as Ca2+ or Mg2+ compete with metal cations for uptake into the organism (Campbell 1995; Santore et al. 2002), the observed differences in the EC50 values for Cd2+ and Ni2+ at different Ca2+ or Mg2+ concentrations (Table 1) are surprisingly high and indicate that the influence of essential macroelements on the heavy metal toxicity is even stronger than expected.
Although phosphate, which was used as pH buffer in this medium, tends to form insoluble salts with certain metal species and hence, reduces the effective concentration of bioavailable heavy metal ions, control experiments with different concentrations of phosphate or HEPES buffer instead of phosphate gave no hint of a strong chelation or potential precipitation of insoluble Cd2+ or Ni2+ salts (data not shown).
In contrast to Cd2+, the transition metal nickel is an essential cofactor for a number of enzymatic reactions. At elevated concentrations, however, Ni2+ is also highly toxic (Babich and Stotzky 1983). Under artificial conditions, i.e. at concentrations that are much higher than in the natural environment, Ni2+ (ion radius, 95 pm) may be taken up by microbial Mg2+ (ion radius, 72 pm) transportation systems such as the fast and unspecific CorA magnesium transporter described for Gram-negative bacteria like Salmonella typhimurium (Tao et al. 1995; Eitinger and Friedrich 1997; Kehres et al. 1998; Smith and Maguire 1998). In these systems, Mg2+ seems to be a competitive inhibitor of Ni2+ transport so that the concentration of Mg2+ may have a significant impact on Ni2+ toxicity.
At comparable concentrations, Ni2+ exhibits a stronger inhibitory effect (EC50 62 ± 1.8 μmol L−1) on growth of P. brassicacearum than Cd2+ (EC50 85 ± 0.5 μmol L−1). This higher toxicity of Ni2+ could possibly be explained by a missing or weak detoxification mechanism, or by its competition with the metabolically higher importance of Mg2+ compared to that of Ca2+. Magnesium acts as cofactor in more than 300 enzymatically catalysed reactions, affecting energetic metabolism, transmembrane ion movements, e.g. Na,K-ATPase, and synthesis of DNA and RNA (Jorgensen et al. 2003; Haber 2004). This would also explain why, in the absence of heavy metal ions, the dependency of the maximum growth rate on the Mg2+ concentration is significantly higher than on the calcium concentration (Fig. 2). Hence, inhibition of Mg2+-based reactions by structural related Ni2+ seems to have a greater influence on bacterial metabolism than inhibition of Ca2+-based reactions by Cd2+. The additive toxic effect (EC50 Cd2+/Ni2+ 40 ± 0.4 μmol L−1) on μmax [per hour] of P. brassicacearum by facing Ni2+ and Cd2+ corroborate the hypothesis of different target sites within bacterial cells.
However, the poor biodegradability of EDTA becomes advantageous for investigations of metal-chelating effects on the inhibition of growth of rhizobacteria, as its total concentration remains constant during the experiments. When EDTA is added to P. brassicacearum cultivations in 4C medium containing toxic nickel or cadmium concentrations, a noticeable increase in the EC50 values is observed (Fig. 5a and b). At equal concentration of heavy metal ions and EDTA, growth of P. brassicacearum is not inhibited which can be explained by the formation of the corresponding Cd- or Ni-EDTA chelates. Compared to Ca- or Mg-EDTA, these are thermodynamically highly stable (compare complexation constants KB in Table 2), leading to very low concentrations of free heavy metal ions in the solution. In summary, this results in a negligible or highly reduced impact of Cd2+ and Ni2+. However, if concentrations of EDTA exceed that of Cd2+ or Ni2+, the chelator also exhibits an inhibitory effect, most probably due to removal of Ca2+ and Mg2+ from the outer cell membrane which affects membrane integrity (Chavez de Paz et al. 2010).
It was shown for a representative PGPR that the heavy metal toxicity depends essentially—even stronger than expected—on environmental conditions like concentrations of Mg2+ and Ca2+, as well as on the presence of chelating agents such as EDTA. Similar ion radii of Mg2+ and Ca2+ and their toxic antagonists Ni2+ and Cd2+ can be assumed responsible for the observed competition, either in uptake mechanism into cells, in toxic effects at cellular targets or for detoxification mechanisms.
Figure 6 summarises the results of this study and illustrates the interactions between the essential divalent cations Ca and Mg, the heavy metal ions Cd and Ni, the chelator EDTA and the observed plant-growth promoting bacteria P. brassicacearum, which could be observed in this study .

Summary of mutual interactions between heavy metals Cd2+ and Ni2+, essential divalent cations Ca2+ and Mg2+, chelator EDTA and plant-growth promoting rhizobacteria P. brassicacearum which could be observed in this study
Additionally and as a further conclusion, the application of EDTA for remediation of heavy metal contaminated soil can be seen from two sides: on the one hand, EDTA has proven to build highly stable complexes with both heavy metal ions tested in this study, leading to a reduced toxicity onto soil bacteria such as P. brassicacearum. On the other hand, EDTA itself poses toxic properties towards bacteria in the uncomplexed form. Besides its poor biodegradability, this observation should lead to careful evaluation of synthetic chelating agents for soil remediation purposes.
For future approaches regarding interactions between soil bacteria and contaminants such as complexing agents and heavy metal ions, a transfer of the experiments discussed in this work into a soil system, e.g. soil column or lysimeter, might be a feasible advance.
References
Abou-Shanab, R. A., Angle, J. S., Delorme, T. A., Chaney, R. L., van Berkum, P., Moawad, H., et al. (2003). Rhizobacterial effects on nickel extraction from soil and uptake by Alyssum murale. The New Phytologist. doi:10.1007/s11274-007-9464-x.
Achouak, W., Sutra, L., Heulin, T., Meyer, J. M., Fromin, N., Degraeve, S., et al. (2000). Pseudomonas brassicacearum sp. nov. and Pseudomonas thivervalensis sp. nov., two root-associated bacteria isolated from Brassica napus and Arabidopsis thaliana. International Journal of Systematic and Evolutionary Microbiology, 50, 9–18.
Achouak, W., Conrod, S., Cohen, V., & Heulin, T. (2004). Phenotypic variation of Pseudomonas brassicacearum as a plant root-colonization strategy. Molecular Plant-Microbe Interactions. doi:10.1094/MPMI.2004.17.8.872.
Aimal, M., & Khan, A. U. (1984). Effect of water hardness on the toxicity of cadmium to microorganisms. Water Resources. doi:10.1016/0043-1354(84)90122-2.
Alkorta, I., Hernández-Allica, J., Becerril, J. M., Amezaga, I., Albizu, I., Onaindia, M., et al. (2004). Chelate-enhanced phytoremediation of soils polluted with heavy metals. Rev Environmental Science and Biotechnology. doi:10.1023/B:RESB.0000040057.45006.34.
Baath, E. (1989). Effects of heavy metals in soil on microbial processes and populations (a review). Water, Air, and Soil Pollution. doi:10.1007/BF00279331.
Babich, H., & Stotzky, G. (1983). Influence of chemical speciation on the toxicity of heavy metals to the microbiota. In J. O. Nriagu (Ed.), Aquatic Toxicology (pp. 1–46). New York: Wiley.
Belimov, A. A., Dodd, I. C., Safronova, V. I., Hontzeas, N., & Davies, W. J. (2007). Pseudomonas brassicacearum strain Am3 containing 1-aminocyclopropane-1-carboxylate deaminase can show both pathogenic and growth-promoting properties in its interaction with tomato. Journal of Experimental Botany. doi:10.1093/jxb/erm010.
Bowen, G. D., Rovira, A. D., & Donald, L. S. (1999). The rhizosphere and its management to improve plant growth. In: Advances in agronomy, vol 66. Academic Press, pp 1-102.
Campbell, P. G. C. (1995). Interactions between trace metals and aquatic organisms: a critique of the free-ion activity model. In A. Tessier & D. R. Turner (Eds.), Metal speciation and bioavailability in aquatic systems (pp. 45–102). London: Wiley.
Chavez de Paz, L. E., Bergenholtz, G., & Svensäter, G. (2010). The effects of antimicrobials on endodontic biofilm bacteria. Journal of Endodontia. doi:10.1016/j.joen.2009.09.017.
Chen, H., & Cutright, T. (2002). The interactive effects of chelator, fertilizer, and rhizobacteria for enhancing phytoremediation of heavy metal contaminated soil. Journal of Soils Sediments. doi:10.1007/BF02991040.
Cook, R. J. (2000). Advances in plant health management in the twentieth century. Annual Review of Phytopathology, 38, 95–116.
Costa R. d. S. (2006). Microbial community structure and function in the rhizosphere of crop plants: a molecular study. Dissertation, Technische Universität Braunschweig
Duxbury, T. (1985). Ecological aspects of heavy metal responses in microorganisms. In K. C. Marshall (Ed.), Advances in Microbial Ecology (Vol 8th ed., pp. 185–236). New York: Plenum.
Eitinger, T., & Friedrich, B. (1997). Microbial nickel transport and incorporation into hydrogenases. In G. Winkelmann & C. J. Carrano (Eds.), Transition metals in microbial metabolism (pp. 235–256). Amsterdam: Harwood.
Giller, K. E., Witter, E., & McGrath, S. P. (1998). Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: a review. Soil Biology & Biochemistry. doi:10.1016/S0038-0717(97)00270-8.
Giller, K. E., Witter, E., & McGrath, S. P. (2009). Heavy metals and soil microbes. Soil Biology & Biochemistry. doi:10.1016/j.soilbio.2009.04.026.
Haber, P. (2004). Magnesium update. Acta Medica Austriaca, 31, 37–39.
Heijerick, D. G., Janssen, C. R., & Coen, W. M. D. (2003). The combined effects of hardness, pH, and dissolved organic carbon on the chronic toxicity of Zn to D. magna: Development of a surface response model. Archives of Environmental Contamination and Toxicology. doi:10.1007/s00244-002-2010-9.
Hong, H. B., Brown, L. R., & Kim, J. K. (1995). Effect of zine and calcium on the intracellularly uptake of cadmium and growth of Escherichia coli. Journal of Microbiology, 33, 302–306.
Jansen, E., Michels, M., Til, M., & Doelman, P. (1994). Effects of heavy metals in soil on microbial diversity and activity as shown by the sensitivity-resistance index, an ecologically relevant parameter. Biology and Fertility of Soils. doi:10.1007/BF00336319.
Jing, Y. D., He, Z. L., & Yang, X. Z. E. (2007). Role of soil rhizobacteria in phytoremediation of heavy metal contaminated soils. J Zhejiang Univ-Sc B, 8, 192–207. doi:10.1631/jzus.2007.B0192.
Jorgensen, P. L., Hakansson, K. O., & Karlish, S. J. D. (2003). Structure and Mechanism of Na,K-ATPase: Functional Sites and Their Interactions. In: Annual Review of Physiology, pp 817-849. doi:10.1146/annurev.physiol.65.092101.142558.
Kamnev, A. A. (2003). Phytoremediation of heavy metals: an overview. In: Fingerman M, Nagabhushanam R (ed) Recent Advances in Marine Biotechnology. Science Publishers Inc, Vol 8, Bioremediation, Enfield (NH) USA, pp 269-317.
Kehres, D. G., Lawyer, C. H., & Maguire, M. E. (1998). The CorA magnesium transporter gene family. Microbial & Comparative Genomics. doi:10.1089/omi.1.1998.3.151.
Khan, M. S., Zaidi, A., Wani, P. A., & Oves, M. (2009). Role of plant growth promoting rhizobacteria in the remediation of metal contaminated soils. Environmental Chemistry Letters. doi:10.1007/s10311-008-0155-0.
Kumar, R., Pandey, S., & Pandey, A. (2006). Plant roots and carbon sequestration. Current Science, 91, 885–890.
Lestan, D., & Finzgard, N. (2007). Leaching of Pb contaminated soil using ozone/UV treatment of EDTA extractants. Separation Science and Technology. doi:10.1080/01496390701290367.
Liao, M., Luo, Y. K., Zhao, X. M., & Huang, C. Y. (2005). Toxicity of cadmium to soil microbial biomass and its activity: Effect of incubation time on Cd ecological dose in a paddy soil. J Zhejiang Univ-Sc B, 6, 324–330. doi:10.1631/jzus.2005.B0324.
Lide, D. R. (1994). CRC handbook of chemistry and physics (75th edition). Boca Raton, Florida: CRC Press.
Liljeroth, E., Kuikman, P., & Van Veen, J. A. (1994). Carbon translocation to the rhizosphere of maize and wheat and influence on the turnover of native soil organic matter at different soil nitrogen levels. Plant and Soil. doi:10.1007/BF00046394.
Lugtenberg, B., & Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annual Review of Microbiology. doi:10.1146/annurev.micro.62.081307.162918.
Lugtenberg, B. J. J., Kravchenko, L. V., & Simons, M. (1999). Tomato seed and root exudate sugars: composition, utilization by Pseudomonas biocontrol strains and role in rhizosphere colonization. Environmental Microbiology. doi:10.1046/j.1462-2920.1999.00054.x.
Lynch, J. M. (1990). The Rhizosphere. New York: Wiley.
Maki, Y., Shibata, J., Murayama, N., Nishimura, Y., & Ueshima, Y. (2009). Treatment of heavy metal contaminated soil with chelating agent. Japanese Source, 35, 258–263.
McGrath, S. P., Chaudri, A. M., & Giller, K. E. (1995). Longterm effects of metals in sewage sludge on soils, microorganisms and plants. Journal of Industrial Microbiology & Biotechnology. doi:10.1007/BF01569890.
Neumann, G., & Römheld, V. (2000). The release of root exudates as affected by the plant physiological status. In Z. Varanini, Z. Nannipieri, & R. Pinton (Eds.), Biochemistry and organic substances at the soil-plant interface. The Rhizosphere: CRC Press.
Oliveira, A., & Pampulha, M. E. (2006). Effects of long-term heavy metal contamination on soil microbial characteristics. Journal of Bioscience and Bioengineering. doi:10.1263/jbb.102.157.
Pack, H. (1996). Schwermetalle in Abwasserströmen: Biosorption und Auswirkungen auf eine schadstoffabbauende Bakterienkultur. Paderborn: FIT-Verlag für Innovation und Technologietransfer.
Pages, D., Sanchez, L., Conrod, S., Gidrol, X., Fekete, A., Schmitt-Kopplin, P., et al. (2007). Exploration of intraclonal adaptation mechanisms of Pseudomonas brassicacearum facing cadmium toxicity. Environmental Microbiology. doi:10.1111/j.1462-2920.2007.01394.x.
Passow, H., Rothstein, A., & Clarkson, T. W. (1961). The general pharmacology of heavy metals. Pharmacology Reviews, 13, 185–224.
Pennanen, T., Frostegard, A., Fritze, H., & Baath, E. (1996). Phospholipid fatty acid composition and heavy metal tolerance of soil microbial communities along two heavy metal-polluted gradients in coniferous forests. Applied and Environmental Microbiology. doi:0099-2240/96/$04.0010.
Pfennig, N. (1974). Rhodopseudomonas globiformis, sp. n., a new species of the Rhodospirillaceae. Archives of Microbiology. doi:10.1007/BF00446317.
Ringbom, A. (1963). Complexation in analytical chemistry. New York: Wiley.
Sandaa, R. A., Torsvik, V., Enger, A., Daae, F. L., Castberg, T., & Hahn, D. (1999). Analysis of bacterial communities in heavy metal-contaminated soils at different levels of resolution. FEMS Microbiology Ecology. doi:10.1111/j.1574-6941.1999.tb00652.x.
Santore, R. C., Mathew, R., Paquin, P. R., & DiToro, D. (2002). Application of the biotic ligand model to predicting zinc toxicity to rainbow trout, fathead minnow, and Daphnia magna. Comparative Biochemistry and Physiology. doi:10.1016/S1532-0456(02)00106-0.
Slaveykova, V. I., Dedieu, K., Parthasarathy, N., & Hajdu, R. (2009). Effect of competing ions and complexing organic substances on the cadmium uptake by the soil bacterium Sinorhizobium meliloti. Environmental Toxicology and Chemistry, 28, 741–748.
Smith, R. L., & Maguire, M. E. (1998). Microbial magnesium transport: unusual transporters searching for identity. Molecular Microbiology. doi:10.1046/j.1365-2958.1998.00810.x.
Tao, T., Snavely, M. D., Farr, S. G., & Maguire, M. E. (1995). Magnesium transport in Salmonella typhimurium: mgtA encodes a P-type ATPase and is regulated by Mg2+ in a manner similar of the mgtB P-type ATPase. Journal of Bacteriology, 177, 2654–2662.
Whiting, S. N., de Souza, M. P., & Terry, N. (2001). Rhizosphere bacteria mobilize Zn for hyperaccumulation by Thlaspi caerulescens. Environmental Science & Technology. doi:10.1021/es001938v.
Zhuang, X., Chen, J., Shim, H., & Bai, Z. (2007). New advances in plant growth-promoting rhizobacteria for bioremediation. Environment International. doi:10.1016/j.envint.2006.12.005.
Acknowledgements
The authors would like to thank the German Research Foundation for financial support (grant of the International Graduate College 802) as well as Y. Göcke, S. Hübner and T. Namuth for their experimental support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Krujatz, F., Haarstrick, A., Nörtemann, B. et al. Assessing the Toxic Effects of Nickel, Cadmium and EDTA on Growth of the Plant Growth-Promoting Rhizobacterium Pseudomonas brassicacearum . Water Air Soil Pollut 223, 1281–1293 (2012). https://doi.org/10.1007/s11270-011-0944-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11270-011-0944-0