
Abstract
As urbanization accelerates, urban biodiversity conservation is becoming a great concern for the maintenance of urban ecosystem functions. In particular, forest bird communities in urban areas have been recognized as a conservation target because of their functions in food webs and ecosystem services. But our understanding of which local- and landscape-scale factors influence native bird communities within urban green spaces is still insufficient to provide managers with information for effectively planning biodiversity management programs. Here we examine how local habitat characteristics, human disturbance, and habitat connectivity influence the diversity of forest bird communities in 44 small forest patches (0.5–20.0 ha) embedded in an urbanized landscape. Patch size exerted a positive influence on the diversity of most bird functional groups, and it had the greatest effects on total abundance and species richness. The second most important factor was human disturbance. Remnant patches with lower levels of human disturbance had higher diversity than newly established patches where intense human activities occurred more frequently. In addition, vegetation complexity and habitat connectivity were positively related to total species richness and abundance, respectively, but they were less important. Management strategies for the conservation of urban forest birds, therefore, should consider not only local improvements in habitat structure – through increased patch size, reduced human disturbance, and increased vegetation complexity – but also the maintenance of habitat connectivity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The environment in and around an urban green space influences biodiversity, its ecological functions, and the ecosystem services that the biodiversity provides (Sadler et al. 2010). However, despite the fact that most urban green spaces are small and scattered (Forman 2014), most urban planning efforts and ecological research have concentrated on larger or contiguous green patches. Indeed, it has been reported that the ecosystem functions and biodiversity value of small urban green spaces have not been adequately explored (e.g., Shwartz et al. 2013). Although small green spaces may not provide as many resources or refuges for various wildlife species as larger patches, they can form a well–connected network that increases overall urban biodiversity (Shanahan et al. 2011). In addition, the wildlife that inhabits small urban green spaces may increase the ecosystem services provided to urban residents, such as aesthetic enjoyment and recreation (Kong et al. 2007). Therefore, understanding how to plan and manage small green spaces to maintain, or even enhance, biodiversity can be of great benefit for policy-makers, planners, and urban residents (Shwartz et al. 2013).
The size, vegetation structure, human disturbance, and connectivity of green spaces are important factors in maintaining avian biodiversity in urban landscapes (reviewed by Fernández-Juricic and Jokimäki 2001). Among them, green space area has been considered a good indicator of urban bird diversity (Melles et al. 2003; Park and Lee 2000; Zhou and Chu 2012). Accordingly, the first management alternative for urban bird conservation might aim to increase the area of existing green space patches. However, revegetation is both expensive and time-consuming in urban areas. As an alternative, local conditions could be enhanced by discouraging human disturbance and by increasing vegetation complexity (i.e., diversity). Or, at a landscape level, connectivity could be increased through matrix management. Although restoration plans are urgently needed, it is not yet clear which of these factors is more important for determining bird diversity in small green spaces.
This study investigates how local habitat structure, human disturbance, and habitat connectivity interplay to influence bird diversity in a heavily-developed metropolitan area (Seoul and its satellite cities, South Korea). This research focused on forest bird assemblages in small patches (<20 ha) as they are known as ecological indicators of habitat structure and human disturbance (O'Connell et al. 2000). Compared to other vertebrates, birds are easy to monitor and provide a mechanism to explore their responses to urbanization (Koskimies 1989; Minor and Urban 2010). Our findings may invite ecologists and urban planners to develop more thoughtful guidelines on how to design and manage a more sustainable urban ecosystem.
Material and methods
Study area and bird surveys
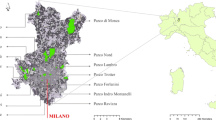
The study was conducted in Seoul and surrounding cities in Gyeonggi Province, South Korea, one of the most densely populated places in the world with a population of over ten million people (KOSTAT 2012). The climate in this area is temperate monsoon, with summer monsoon rainfall (892 mm per year), predominantly dry winter, and an average annual temperature of 12.5 °C. About 30 % of the study area is covered by forests (KFS 2012), and urban remnant forest patches are under pressure of development (Kim 2003). Forty-four forest patches between 0.5 and 20.0 ha in size were selected from throughout the study area (Fig. 1). In order to avoid spatial autocorrelation biases, the patches were separated by a minimum distance of 1 km (Legendre et al. 2002).

(a) Map of Seoul and surrounding area with the study sites. Aerial photos of two patch types provided by Daum Kakao Corp, (b) a remnant forest patch, and (c) a new patch that was established in June 30, 1988
In 2012, four repeat bird surveys were conducted at each patch using a line transect method (Bibby et al. 2000; Park and Lee 2000), three times during the breeding season (April-July) and once during the non-breeding season (September-October), between 30 min before sunrise and 4 h after (a total of 176 surveys). At each patch, a single transect line was set up at the length of 0.1 to 2 km according to its patch size, enough to cover the patch area. All birds heard or seen within 25 m of both sides of the transect line were recorded. The time spent on each patch was also dependent on its area but was not less than 20 min duration, which was sufficient to make an exhaustive search for all species. Birds just flying over the site were not included in the count. Urban exploiters (Black-billed Magpie Pica pica, Eurasian Tree Sparrow Passer montanus, and Feral Pigeon Columba livia) were also excluded because their abundances are not dependent upon the types or amount of vegetation (Johnston 2001; Lancaster and Rees 1979).
Species richness and abundance estimates were derived for each patch. Species richness was the total number of recorded bird species within each survey patch, and species abundance was the average number of individuals counted across all surveys at a patch. Each species was assigned to various functional groups based on migratory, nesting and foraging strategy, diet, and habitat use, as well as species rarity (Table 1), according to Lee and Park (1995), field observations, and expert opinion. Diet guild was assigned based on the main foods eaten in the breeding season.
Variables measured
Local-scale habitat characteristics
We measured local habitat attributes, including patch area and vegetation complexity. A forest-cover map for the study area was derived from a biotope map (SMG 2010). The biotope map was originally created by Seoul metropolitan government using aerial photographs and satellite imagery interpretation, and was later classified according to ground-truthed data. To accurately estimate patch area, we digitized the edges of patches using the forest map and fine-resolution (0.5 m) aerial photographs (dated from May 2011) provided by Daum Kakao Corp.
Complexity in vegetation structure and composition are important predictors of bird diversity (Joshi et al. 2012). Hence, we surveyed the vegetation characteristics of patch sites from September to October 2012 when most plants were fully grown. A 100 m transect was randomly located within each site, and vegetation was surveyed within 2 m of both sides of this line, giving a total survey area of 400 m2. The number of transects per patch was based on patch area: 1 transect for <10 ha patches and three transects for 10–20 ha patches (Miller and Cale 2000). Transect measurements for each >10 ha patch were averaged to obtain an overall estimate. Within the transect, we recorded species identity and stem diameter for all woody plants greater than or equal to 5 cm diameter at breast height (DBH). We also recorded canopy cover and presence/absence of shrub, grass, litter layer, and coarse and fine woody debris at 2-m intervals along the transect, for a total of 51 sampling locations. We estimated canopy cover using a densiometer. Each of these observations was then combined to estimate mean canopy cover, shrub cover, ground vegetation cover, leaf litter cover, and cover of fine and coarse woody debris for each transect. The resulting vegetation variables were used to describe each forest patch:
-
(i)
Tree (>5 m height), shrub (1–5 m height), and total woody species richness per 400 m2;
-
(ii)
Woody stem density per 400 m2;
-
(iii)
Average and total woody stem basal area per 400 m2;
-
(iv)
Basal area of hardwood and conifer trees and snags (standing dead trees ≥5 cm DBH) per 400 m2;
-
(v)
Estimates of percent cover of vegetation (canopy, >5 m height; shrub, 1–5 m height; ground, 0–1 m height), leaf-litter, and woody debris (fine woody debris, 1–10 cm diameter; coarse woody debris, >10 cm diameter)
Principal coordinate analysis (PCoA) (Legendre and Legendre 1998) based on a Bray–Curtis similarity distance was performed to combine these multiple vegetation characteristics into fewer explanatory variables. PCoA is an ordination technique, which has some advantage over principal component analysis (PCA). For instance, in PCoA, any ecological distance can be applied, while only Euclidean distance can be used as a similarity measure in PCA. In the study, two-dimensional PCoA was implemented by the package labdsv in R (Roberts 2013). The first and second PCoA axes explained 57.2 and 13.6 % of the total variance in vegetation characteristics, respectively. The resulting first and second scores of the ordination were referred as ‘Veg complexity 1′ and ‘Veg complexity 2′ (Shanahan et al. 2011). Veg complexity 1 was negatively correlated with mean tree basal area and positively correlated with the following variables: total basal area and basal area of hardwood trees and snags; percent cover of shrub, leaf-litter, and fine and coarse woody debris; total woody and shrub plant richness; and woody stem density (Pearson’s r) (p < 0.05). Veg complexity 2 was positively correlated with percent cover of shrub, leaf-litter, and fine and coarse woody debris; total woody, tree, and shrub species richness; and mean tree basal area (Pearson’s r) (p < 0.05). The sites with high values of Veg complexity 1 and 2 had complex forest structures with relatively dense young and less dense mature trees in urban areas, respectively. Thus, the two vegetation complexity measures indicated a gradient of vegetation diversity and basal area heterogeneity.
Human disturbance
Patch type and human population density were included as human disturbance factors that might influence bird species richness and abundance. Two main patch types were considered: newly established and remnant native forest patches (Fig. 1). The remnant patches are sites that have never been cleared for urban development. On the contrary, newly established forest patches, which are mainly urban parks, are vegetation-covered sites that have been created and planted with trees in the late 20th and early 21st centuries. Newly established and remnant patches differed in the severity of human disturbance, as the number of visitors is the major driver that could disturb bird species (Fernández-Juricic and Tellería 2000). For example, newly established sites have more than 500 visitors per day, while there were relatively few visitors (i.e., less than 50 people per day) to remnant patches (W. Kang, personal observation). In the study, a remnant patch was coded as 0 and a newly established patch as 1. We also estimated human population density (person / km2) within a 1 km buffer around the edges of each patch using the BIZ-GIS database (http://www.biz-gis.com/GISDB), based on the 2005 population and housing census in South Korea (KSIS 2005).
Landscape variables
Normalized difference vegetation index (NDVI) was chosen as a larger-scale measure of vegetation, as it is strongly related to the amount of vegetation cover (Purevdorj et al. 1998). NDVI values were estimated from 15-m ASTER imagery from May 9, 2012 and classified in 13 classes ranging from zero (concrete structures) to 12 (high-density vegetation) (Shwartz et al. 2013). The average of the class values (i.e., the average green proportion) for five buffer zones (100–500 m) around a patch was then calculated. Because the five mean NDVI values were highly correlated, hierarchical partitioning (Mac Nally 2002) was performed to select the variables that had the strongest independent influence on bird species richness and abundance, i.e., the mean NDVI in a 100 m buffer zone around the patch.
Patch connectivity at a landscape level was also measured, using a graph-theoretical approach. The connectivity measure was based on the probability of connectivity (PC; see a more detailed description in Saura and Pascual-Hortal (2007)). PC is defined as the probability that two points (organisms) placed randomly in a landscape fall into habitat areas that are reachable from each other (interconnected), given a set of habitat patches and links among them (Saura and Pascual-Hortal 2007). In order to compute PC and its fractions, the links between every two patches i and j first need to be characterized by the probability of dispersal (p ij ), here obtained as a negative exponential function of the Euclidean distance between patches (Bunn et al. 2000; Urban and Keitt 2001). The path with the maximum product probability (p ij *) is then considered the best possible one for the movement of individuals from patch i to j through the network of patches. The importance of a patch as a stepping stone between other patches was estimated through the dPCconnector fraction derived from the PC metric (refer to Saura and Rubio 2010: 526–7 for details on dPCconnector). dPCconnector measures the contribution of a patch to the connectivity between other patches, as an irreplaceable connecting element or stepping stone between them (Saura and Rubio 2010). A certain patch will have a dPCconnector value that is greater than zero only when it meets two criteria: (1) it is part of the best (i.e., maximum product probability) path between other patches in a landscape, and (2) when, after losing that patch, the alternative paths between the remaining patches cannot compensate for the connecting role played by that patch in an intact landscape (Bodin and Saura 2010). The dPCconnector was calculated at distance thresholds of 0.5, 1, 1.5, 2, 3, and 5 km for every patch in the study area. A dispersal probability of 0.05 was defined to correspond to the threshold dispersal distance. Since the six dPCconnector values were highly correlated, we performed hierarchical partitioning (Mac Nally 2002) to select the variable with the greatest independent influence on bird species richness and abundance, i.e., the dPCconnector value at the distance threshold of 500 m.
Data analyses
Seventeen separate generalized linear models (GLMs) were performed, using a log link function assuming a Poisson distribution, to explore the relative influence of local- and landscape-level variables and human disturbance on various aspects of bird diversity. To model abundance of all forest birds, bush nesters, and bush-foraging birds, a negative binomial distribution and a logarithmic link function was used to explain over-dispersion in the observed data. The Poisson and negative binomial models were implemented using the package stats (Chambers and Hastie 1992) and MASS (Venables and Ripley 2002), respectively, in R (R Core Team 2013). No analysis was conducted for abundance of house nesters, aerial foragers, or carnivores, as there was an insufficient number of species or individuals in these groups (Table 1).
A model-averaging approach based on information criteria was adopted for model selection (Burnham and Anderson 2002). First, all models were ranked according to the AICc (corrected Akaike Information Criterion) using the MuMIn package in R (Barton 2014). Variables included in the most parsimonious models with ΔAICc values below 4 were identified by averaging their estimated coefficients and associated standard errors weighted by each model’s AICc (Burnham and Anderson 2002). Finally, coefficients and standard errors for the variables that had p-values <0.05 were presented. We also present the adjusted R-squared value for each model, which was calculated as the average of the adjusted R-squared values in the most parsimonious models. Except vegetation complexity measures and dummy variables, all measurements were log-transformed (log[x + 1]) to improve normality. Before executing multivariate regressions, multicollinearity among independent variables was tested by performing Pearson’s correlations to ensure that no variables were strongly correlated (|r | < 0.53).
Alongside a model-averaging procedure, this study used a hierarchical partitioning approach to quantify the independent contribution of each explanatory variable to the response variables’ total species richness and total bird abundance (Chevan and Sutherland 1991). R-squared (r 2) was used as a goodness-of-fit measure. A randomization procedure was performed with 1,000 iterations to determine the statistical significance of independent effects (Mac Nally 2002). The package hier. part in R was used in the analysis (Walsh and Mac Nally 2013).
Results
In total, 46 bird species and 1,925 individual birds were observed across our study sites, with an average of 11.7 species ± 6.0 SD per patch (Table 1). Although the majority of the observed birds were common species, four rare migrant species (comprising 11 individuals) were also observed. The observed species were almost evenly divided between migrants and residents. Most species were canopy nesters (n = 15), cavity nesters (n = 11), or ground nesters (n = 9); only one species was a bush nester although it was very abundant (282 observations). Twenty-five species were canopy foragers, five were bush foragers, and seven were ground foragers. The majority of the species were classified as insectivores, while two species were granivores and three were omnivores. More species preferred interior habitat, or were edge species than were habitat generalists.
Local habitat, landscape variables, and human disturbance explained most of the variance in total species richness, bird abundance, and abundances of different guilds. Among local habitat characteristics, patch area was positively correlated with total species richness, total bird abundance, and abundances of different guilds, except for the ground-foraging and granivore bird guilds (Table 2). Veg complexity 2 showed positive effects on the total species richness and abundances of migrant and ground-nesting birds (Table 2).
Patch type, which is classified according to the presence or levels of human disturbance, was also important in explaining the variance of both richness and abundance of birds. For example, total species richness and abundance were significantly higher in the remnant patches than in the newly established forest patches (Table 2). Furthermore, resident, canopy-, cavity-, and bush-nesting, bush- and ground-foraging, insectivore and granivore, and edge species were more common in the remnant patches. One landscape variable exerted significant influences on the attributes of urban forest bird communities. dPCconnector measured at a distance threshold of 500 m had positive effects on the total bird abundance and abundances of resident, bush-nesting, insectivore, and edge species (Table 2). On the contrary, Veg complexity 1, mean NDVI 100 m, and human population density showed no significant influence on bird species diversity (Table 2).
Patch area was the most important variable for total species richness, followed by patch type and Veg complexity 2 (Fig. 2a). It also showed the greatest effect on total bird abundance, followed by patch type and dPCconnector 500 m (Fig. 2b).

The independent contribution of each variable to model fit for (a) the total bird species richness and (b) abundance data, as determined by hierarchical partitioning. The model includes all seven independent variables indicated in the figure. An asterisk (*) indicates the variables for which its independent percentage contribution to overall model fit was significant at p < 0.05
Discussion and conclusions
Conserving biodiversity in urban areas has become a high priority (Alvey 2006). The results showed that even small (<20 ha) forest patches in a highly urbanized and densely populated region can support a significant functional diversity of bird communities. The birds recorded in this study correspond to nearly 70 % of the forest bird species observed in 10 large forest patches (over 100 ha) in the Seoul region (Park and Lee 2000). In addition, the birds sampled accounted for 60 % of the total forest bird species observed in the continuous non-disturbed deciduous forest (ca. 2,240 ha) in central Korea (Choi et al. 2006). More importantly, rare migrant species were also detected in the small urban forest fragments, even though the majority of forest birds sampled were common species. Both rare and common species are essential for biodiversity conservation, since they all contribute to ecosystem functioning and urban ecosystem health (Gaston 2010; Lyons et al. 2005). The co-occurrence of these species indicates that small forest patches not only shelter groups of common species but also act as havens for rare migrating birds within the urban landscape.
The positive influence of patch area on the abundances of most functional groups of bird species (Table 2) is notable, as it had the greatest effects on both overall species richness and abundance (Fig. 2). Several studies have already suggested that patch area is the best predictor of bird species richness and diversity in vegetation fragments (e.g., Carbó-Ramírez and Zuria 2011; Drinnan 2005). In this study, only a small range of patch sizes (0.5–20 ha) was considered, and patch area still appeared to have the greatest effect on both the total species richness and abundance (Fig. 2). However, this strong relationship between patch area and bird community diversity is typically related to a size threshold of ca. 3.5–5 ha, the point at which species richness rapidly declines (Drinnan 2005). This study also identified a patch size threshold for forest birds of ca. 3.5 ha (data not shown). It is, therefore, feasible to conclude that small changes in the area of urban forest remnants may increase the species richness and functional diversity of birds, and forest remnants should be a minimum of 3.5 ha.
Forest habitats in urban areas, especially small patches, generally experience a higher degree of human use compared to rural forests. Such use leads to direct or indirect disturbance, through activities such as walking either on or off trails, which can be detrimental to forest bird species (Fernández-Juricic 2004). The degree of human disturbance in the focal patches, as measured by patch type, also showed a strong effect on the richness and abundance of birds (Table 2). In fact, patch type was the second most influential factor for both overall species richness and abundance (Fig. 2). Remnant patches with a low level of human disturbance had higher species richness and abundance than newly established patches where more human activities occurred. In particular, the abundances of bush–nesting and bush– and ground–feeding species, which are expected to be most heavily affected by human disturbance, were related to patch type. High levels of human visitation to urban forest fragments may decrease temporal and spatial resource availability, particularly throughout the breeding season (Fernández-Juricic 2000b). As human intrusion into habitats increases, the probability of local colonization would decrease (Fernández-Juricic 2000b), which may lead to functional homogenization of bird communities by including a higher proportion of exotic or disturbance–tolerant species (Devictor et al. 2007). Thus, managers might want to consider controlling human disturbance in newly established patches. Although species responses to disturbance are complex and variable (Blumstein et al. 2005), establishing fencing to keep people away from potentially sensitive centers of bird activity, even on a small scale, will likely lessen the impacts to bird communities in urban forests (Ikuta and Blumstein 2003).
Vegetation complexity can increase forest bird species richness (Evans et al. 2009; Husté et al. 2006). In general, mature trees and high complexity of vegetation cover provide more diverse habitats (Karr and Roth 1971). Especially in urban habitats, the presence of a shrub layer has been shown to be important to bird species diversity, especially low-nesting species (Burr and Jones 1968; Tilghman 1987). In this study, the vegetation complexity variable, Veg complexity 2, which was positively correlated with percent cover of shrub and mean tree basal area, had a positive effect on total species richness and abundances of migrant and ground-nesting birds (Table 2). Vegetation complexity would increase the probabilities of occupation by forest-nesting birds, and in particular those species that nest on the ground or in low shrubs, presumably because of a higher degree of the availability of superior nesting and feeding sites (Fernández-Juricic 2000a). Thus, increasing the vegetation complexity of urban forest fragments could help improve local bird diversity.
Contrary to total species richness and abundance of migrant birds, we found no effect of vegetation complexity on total abundance and abundance of residents. A possible explanation for this result is that most resident birds were canopy- and cavity-nesters (ca. 80 %), and only about 18 % of resident birds were low-nesting species. In addition, resident and low-nesting birds accounted for about 88 and 19 % of the total bird abundance, respectively. Therefore, the vegetation complexity variable may show no significant influence on total abundance and abundance of resident birds.
Spatial arrangement of habitat fragments and its effect on species movement are another crucial issue in landscapes where habitat comprises <30 − 40 % of the total land cover (Andren 1994; Fahrig 2001). A high level of connectivity may favor higher abundances of local populations and, therefore, may reduce the extinction risk of species (Brooker et al. 1999; Haas 1995). In our landscape, which has approximately 30 % forest cover, habitat connectivity had a critical effect on total abundance and abundances of some bird groups (Table 2). This was particularly true for insectivorous birds, which is consistent with previous research indicating that insectivorous birds are particularly susceptible to both reduced patch size and decreased connectivity (e.g., Martensen et al. 2008). Because abundance of insects is negatively influenced by forest fragmentation (Gonzalez et al. 1998), their abundance may increase with habitat connectivity. This could explain why the abundance of insectivorous birds was correlated with habitat connectivity.
Total bird abundance was positively influenced by habitat connectivity, but not total species richness. A possible explanation is that landscape connectivity increases local population density and thereby possibly reduces extinction rates (Steffan-Dewenter 2003). Thus, we might expect that connectivity has a greater influence on species abundance. Moreover, the increasing abundance of certain bird species in well-connected urban forest patches resulted in strong competition for nesting sites with other species and, thus, an insignificant relationship between connectivity and species richness.
Urban forests offer many essential ecological services, including habitat provision for birds and other wildlife species, as well as benefits to humans (MA 2005). Thus, conservation of urban forests is vital for healthy ecosystems. These results elucidate important local and landscape factors affecting forest bird communities in urban environments, which serve as reasonable surrogates for assessing urban biodiversity (Shwartz et al. 2013). Overall, this study showed that the diversity of bird species in an urban landscape is mainly influenced by patch size and human disturbance and, to a lesser extent, by vegetation complexity and connectivity. Local improvements to habitat structure—through increased patch area, reduced human disturbance, or increased vegetation complexity—could positively contribute to local species diversity. Moreover, preserving and promoting connectivity may enhance regional bird biodiversity. This management strategy would require the identification of crucial regions and gaps for connectivity between existing urban fragments so as to establish new habitats and corridors (i.e., revegetation) with more effective, ecological functions. These prioritized recommendations would greatly benefit forest bird communities of urban areas, and would allow for a more sustainable urban environment for human health and well-being.
References
Alvey AA (2006) Promoting and preserving biodiversity in the urban forest. Urban For Urban Green 5:195–201
Andren H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Barton K (2014) MuMIn: multi-model inference. R Package Version 1.10.5. R Foundation for Statistical Computing, Vienna, Austria, http://cran.r-project.org/web/packages/MuMIn/index.html. Accessed 11 Sep 2014
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2000) Bird census techniques, 2nd edn. Academic Press, London
Blumstein DT, Fernández-Juricic E, Zollner PA, Garity SC (2005) Inter-specific variation in avian responses to human disturbance. J Appl Ecol 42:943–953. doi:10.1111/j.1365-2664.2005.01071.x
Bodin Ö, Saura S (2010) Ranking individual habitat patches as connectivity providers: integrating network analysis and patch removal experiments. Ecol Model 221:2393–2405. doi:10.1016/j.ecolmodel.2010.06.017
Brooker L, Brooker M, Cale P (1999) Animal dispersal in fragmented habitat: measuring habitat connectivity, corridor use, and dispersal mortality. Conserv Ecol 3:4
Bunn A, Urban D, Keitt T (2000) Landscape connectivity: a conservation application of graph theory. J Environ Manag 59:265–278
Burnham K, Anderson D (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Burr RM, Jones R (1968) The influence of parkland habitat management on birds in Delaware. T N Am Wildl Nat Res 33:299–306
Carbó-Ramírez P, Zuria I (2011) The value of small urban greenspaces for birds in a Mexican city. Landsc Urban Plann 100:213–222
Chambers JM, Hastie TJ (1992) Statistical models in S. Chapman and Hall, London
Chevan A, Sutherland M (1991) Hierarchical partitioning. Am Stat 45:90–96
Choi CY, Nam HY, Hur WH, Lee WS, Kim HJ, Hwang GY (2006) Edge preference of forest-dwelling birds in the temperate deciduous forests. J Ecol Field Biol 29:191–203 (in Korean with English abstract)
Devictor V, Julliard R, Couvet D, Lee A, Jiguet F (2007) Functional homogenization effect of urbanization on bird communities. Conserv Biol 21:741–751
Drinnan IN (2005) The search for fragmentation thresholds in a southern Sydney suburb. Biol Conserv 124:339–349
Evans KL, Newson SE, Gaston KJ (2009) Habitat influences on urban avian assemblages. Ibis 151:19–39
Fahrig L (2001) How much habitat is enough? Biol Conserv 100:65–74
Fernández-Juricic E (2000a) Avifaunal use of wooded streets in an urban landscape. Conserv Biol 14:513–521
Fernández-Juricic E (2000b) Local and regional effects of pedestrians on forest birds in a fragmented landscape. Condor 102:247–255
Fernández-Juricic E (2004) Spatial and temporal analysis of the distribution of forest specialists in an urban-fragmented landscape (Madrid, Spain): implications for local and regional bird conservation. Landsc Urban Plann 69:17–32
Fernández-Juricic E, Jokimäki J (2001) A habitat island approach to conserving birds in urban landscapes: case studies from southern and northern Europe. Biodivers Conserv 10:2023–2043
Fernández-Juricic E, Tellería JL (2000) Effects of human disturbance on spatial and temporal feeding patterns of Blackbird Turdus merula in urban parks in Madrid, Spain. Bird Study 47:13–21
Forman RTT (2014) Urban ecology: science of cities. Cambridge University Press, Cambridge
Gaston KJ (2010) Valuing common species. Science 327:154–155
Gonzalez A, Lawton J, Gilbert F, Blackburn T, Evans-Freke I (1998) Metapopulation dynamics, abundance, and distribution in a microecosystem. Science 281:2045–2047
Haas CA (1995) Dispersal and use of corridors by birds in wooded patches on an agricultural landscape. Conserv Biol 9:845–854
Husté A, Selmi S, Boulinier T (2006) Bird communities in suburban patches near Paris: determinants of local richness in a highly fragmented landscape. Ecoscience 13:249–257
Ikuta LA, Blumstein DT (2003) Do fences protect birds from human disturbance? Biol Conserv 112:447–452
Johnston RF (2001) Synanthropic birds of North America. In: Marzluff JM, Bowman R, Donnelly RE (eds) Avian ecology and conservation in an urbanizing world. Kluwer Academic, Norwell, Massachusetts, pp 49–67
Joshi KK, Bhatt D, Thapliyal A (2012) Avian diversity and its association with vegetation structure in different elevational zones of Nainital district (Western Himalayan) of Uttarakhand. Int J Biodivers Conserv 4:364–376
Karr JR, Roth RR (1971) Vegetation structure and avian diversity in several New World areas. Am Nat 105:423–435
Kim J (2003) Forest fragmentation effects from urban expansion in Seoul and its satellite cites. Master degree thesis, Seoul National University (in Korean with English abstract)
Kong F, Yin H, Nakagoshi N (2007) Using GIS and landscape metrics in the hedonic price modeling of the amenity value of urban green space: a case study in Jinan City, China. Landsc Urban Plann 79:240–252
Korea Forest Service (KFS) (2012) Statistical yearbook of forestry. 468 p (in Korean)
Korean Statistical Information Service (KSIS) (2005) The 2005 population and housing census of Korea (in Korean)
Koskimies P (1989) Birds as a tool in environmental monitoring. Ann Zool Fenn 26:153–166
Lancaster RK, Rees WE (1979) Bird communities and the structure of urban habitats. Can J Zool 57:2358–2368
Lee W, Park C-R (1995) Analysis of changes on the forest environment and the bird community in terms of guild. Korean J Environ Ecol 18:397–408 (in Korean with English abstract)
Legendre P, Dale MR, Fortin MJ, Gurevitch J, Hohn M, Myers D (2002) The consequences of spatial structure for the design and analysis of ecological field surveys. Ecography 25:601–615
Legendre P, Legendre L (1998) Numerical ecology. 2nd english edn. Elsevier Science, Amsterdam
Lyons KG, Brigham CA, Traut BH, Schwartz MW (2005) Rare species and ecosystem functioning. Conserv Biol 19:1019–1024. doi:10.1111/j.1523-1739.2005.00106.x
Mac Nally R (2002) Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodivers Conserv 11:1397–1401
Martensen AC, Pimentel RG, Metzger JP (2008) Relative effects of fragment size and connectivity on bird community in the Atlantic Rain Forest: implications for conservation. Biol Conserv 141:2184–2192. doi:10.1016/j.biocon.2008.06.008
Melles S, Glenn S, Martin K (2003) Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient. Conserv Ecol 7:5
Millennium Ecosystem Assessment (MA) (2005) Ecosystems and human well-being. Island Press
Miller JR, Cale P (2000) Behavioral mechanisms and habitat use by birds in a fragmented agricultural landscape. Ecol Appl 10:1732–1748
Minor E, Urban D (2010) Forest bird communities across a gradient of urban development. Urban Ecosyst 13:51–71
O'Connell TJ, Jackson LE, Brooks RP (2000) Bird guilds as indicators of ecological condition in the central Appalachians. Ecol Appl 10:1706–1721. doi:10.1890/1051-0761(2000)010[1706:bgaioe]2.0.co;2
Park CR, Lee WS (2000) Relationship between species composition and area in breeding birds of urban woods in Seoul, Korea. Landsc Urban Plann 51:29–36
Purevdorj T, Tateishi R, Ishiyama T, Honda Y (1998) Relationships between percent vegetation cover and vegetation indices. Int J Remote Sens 19:3519–3535
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/. Accessed 11 Sep 2014
Roberts D (2013) Labdsv: ordination and multivariate analysis for ecology. R package version 1.6-1. R Foundation for Statistical Computing, Vienna, http://cran.r-project.org/web/packages/labdsv/index.html. Accessed 11 Sep 2014
Sadler J, Bates A, Hale J, James P (2010) Bringing cities alive: the importance of urban green spaces for people and biodiversity. In: Gaston KJ (ed) Urban ecology. Cambridge University Press, Cambridge, pp 230–260
Saura S, Pascual-Hortal L (2007) A new habitat availability index to integrate connectivity in landscape conservation planning: comparison with existing indices and application to a case study. Landsc Urban Plann 83:91–103
Saura S, Rubio L (2010) A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 33:523–537
Seoul Metropolitan Government (SMG) (2010) The city biotope map of Seoul, Korea
Shanahan DF, Miller C, Possingham HP, Fuller RA (2011) The influence of patch area and connectivity on avian communities in urban revegetation. Biol Conserv 144:722–729. doi:10.1016/j.biocon.2010.10.014
Shwartz A, Muratet A, Simon L, Julliard R (2013) Local and management variables outweigh landscape effects in enhancing the diversity of different taxa in a big metropolis. Biol Conserv 157:285–292
Statistics Korea (KOSTAT) (2012) Population projections for provinces: 2010–2040. Korea
Steffan-Dewenter I (2003) Importance of habitat area and landscape context for species richness of bees and wasps in fragmented orchard meadows. Conserv Biol 17:1036–1044
Tilghman NG (1987) Characteristics of urban woodlands affecting breeding bird diversity and abundance. Landsc Urban Plann 14:481–495
Urban D, Keitt T (2001) Landscape connectivity: a graph-theoretic perspective. Ecology 82:1205–1218
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer-Verlag, New York
Walsh C, Mac Nally R (2013) Hier.Part: hierarchical Partitioning, R package version 1.0-4. R Foundation for Statistical Computing, Vienna, http://cran.r-project.org/web/packages/hier.part/index.html. Accessed 11 Sep 2014
Zhou D, Chu LM (2012) How would size, age, human disturbance, and vegetation structure affect bird communities of urban parks in different seasons? J Ornithol 153:1101–1112
Acknowledgments
We appreciate the advice on bird fieldwork offered by Ms. Hyunsook Yang. We also thank GoWoon Kim (Seoul National University) for English correction of the manuscript and Bumsuk Seo (University of Bayreuth) and two anonymous reviewers for their helpful comments. This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (No. 2011–0024289).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kang, W., Minor, E.S., Park, CR. et al. Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst 18, 857–870 (2015). https://doi.org/10.1007/s11252-014-0433-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-014-0433-5