
Abstract
The high genetic variation within indigenous chickens (IC) which provides an opportunity to select superior stock for sustainable production and conservation is under-exploited. This study is aimed at estimating heritability and genetic and phenotypic correlation coefficients of productive and reproductive traits of Ugandan IC as a basis for selection. Data on traits were collected across two consecutive generations, weight (W) and shank length (SL) of chicks at hatching (HW) as well as at 2 (W2; SL2), 4 (W4; SL4), 6 (W6; SL6), 8 (W8; SL8), and 12 (W12; SL12) weeks of growth. Body weights at onset of lay (WFE) were also measured. In addition, egg number (EN-60), egg weight (EW), clutch number (CLN-60), and clutch size (CLS-60) over a period of 60 days were recorded. Genetic parameters were estimated using the univariate animal model analysis with restricted maximum likelihood procedure using the variability package of R, version 4.1.1. Heritability of traits ranged from 0.30 and 0.72 except SL4 (0.02), SL12 (0.14), and EN-60 (0.17). The traits EN-60 and W4 were negatively phenotypically correlated (− 0.49). Body weight at first egg was highly genetically correlated (0.99) with SL8. Egg number was significantly, negatively, and genetically correlated (− 0.96) with SL12. In conclusion, shank length is a potential phenotypic marker when selecting for live weight at onset of lay and egg yield. The shank length could, therefore, permit selection of superior chickens at an early age.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Indigenous chickens (IC) continue to retain a wide range of desirable attributes and contribute to the nutritional and social-economic well-being of rural households in Asia and Africa (Mahendra 2016; Rajkumar 2021). However, the contribution of IC to the well-being of rural households is limited by their innate slow growth and low egg productivity (Wong et al. 2017). On average, the on-set of lay is delayed up to 7.2 months, and hens lay only 36 eggs per year. On the side of cockerels, sexual maturity is delayed up to 5.9 months (Nakkazi et al. 2014; Ssewannyana et al. 2008). The high genetic variation within IC, which provides an opportunity to identify replacement stock with desired productive and reproductive traits, remains under exploited (Adoligbe et al. 2020; Fleming et al. 2016; Lubandi et al. 2018; Magothe et al. 2010; Ssewannyana et al. 2008; Wong et al. 2017). The common practice of genetic improvement through cross breeding with highly specialized breeds has accelerated loss of important adaptation, broodiness, and reproductive traits critical for survival under harsh, rural resource-constrained farming systems (Tadelle et al. 2000; Dana et al. 2010; Semahoro et al. 2018). Unlike cross breeding, selecting within the surviving Ecotypes provides an opportunity to conserve the IC genetic resources and its associated multiple traits (Dana et al. 2010; Osei-Amponsah et al. 2013; Zonuz et al. 2013). In addition, the unique genomic and phenotypic selection signatures acquired by IC over generations for their adaptation to harsh environments need to be conserved (Fleming et al. 2016; Elbeltagy et al. 2019).
Acquisition and expression of traits by the offspring depends on the degree of inheritance and association of responsible genes. Apart from varying across genotypes and environments, genetic parameters change with age and physiological status of chickens (Magothe et al. 2007; Dana et al. 2011; Gwaza et al. 2016; Singh et al. 2018; Chomchuen et al. 2022; Sanda et al. 2022). The variation in heritability of traits with age, therefore, requires timing of selection at a stage when heritability is at its peak (Adeyinka et al. 2006; Osei-Amponsah et al. 2013; Singh et al. 2018).
The association between traits indicates the underlying genetic interactions, and it is always important to know the effect of selecting one trait on others (Sanda et al. 2022). Genetic and phenotypic correlations determine a combination of traits that can be considered simultaneously in a multi-trait selection program (Magothe et al. 2007; Sanda et al. 2022). A moderate to high genetic correlation among traits is required to achieve a high breeding value (Magothe et al. 2007; Niknafs et al. 2012). Negative additive genetic correlations have been reported for egg production and live weight (Ghazikhani et al. 2007). Such negative genetic correlation of traits creates a puzzle for upgrading multipurpose IC reared mostly for both meat and eggs.
Genetic improvement through selection of superior replacement stock requires population-specific genetic parameter estimates of productive and reproductive traits. Genetic and phenotypic parameter estimates guide the setting of breeding objectives, stage of selection, and marker traits in a breeding program. In addition, previous genetic parameter estimation studies targeted a single Ecotype from a selected province (Dana et al. 2011; Norris and Ngambi 2006). Therefore, this study is aimed at estimating the genetic and phenotypic parameters of productive and reproductive traits of a mixed population of IC Ecotypes under a supplemented scavenging system.
Materials and methods
Acquisition of eggs and rearing of chickens
One thousand two hundred (1200) fresh fertile IC eggs (1–5 days) were purchased from selected rural households in six agro-ecological zones of Uganda as previously described (Beyihayo et al. 2022). Briefly, eggs from six IC Ecotypes (Madi, Acholi, Lango, Nteso, Nsoga, and Ngisu) were collected for the study. Rural households rearing IC with no access to exotic chicken or eggs were selected. In addition, egg-contributing rural households were selected on the basis of having kept IC for a minimum of 3 years. Ownership of an indigenous mature cock by the household was also considered for higher egg fertility and hatchability. Eggs were packed in paper trays and delivered to Gulu University Poultry Unit for artificial incubation using a commercial incubator operated by an Automatic Computer Controller (Model: XM-18, India). Seven hundred thirteen (713) chicks were hatched, wing tagged, and brooded artificially for 4 weeks using a commercial chick starter (20.3% crude protein, 7.5% crude fiber, 0.5% phosphorus, and 1.1% calcium) purchased from Biyinzika Enterprises Limited. Heat for the brooder was provided by clay pots using charcoal as a source of fuel for a period of 2 weeks. After the brooding period, chickens were reared under a scavenging system and supplemented with 15% and 11% crude protein diets during their growth and laying phases, respectively. Chickens were vaccinated following a schedule recommended for commercial egg-type chickens.
Chickens were sexed at 8 weeks of age into 250 pullets and 241 cockerels. Using body weight at the onset of lay and the number of eggs laid in two consecutive clutches as criteria, 16 out of the 68 hens evaluated were selected to become parents for the second generation. Hens weighing at least 1213 g at the onset of lay, with the potential of laying 16 eggs and above in two consecutive clutches were selected (Figs. 1 and 2). Four cockerels were selected based on live body weight at 12 weeks of age for random mating with the selected hens at each generation (Fig. 3). The selected cockerels weighed between 657 and 992 g at 12 weeks. The ratio of cocks to hens was maintained at 1:4 to increase the chances of fertility for each hen and variation within the population. Unselected chickens were then culled due to limited space. The collection of fertile eggs intended for hatching and raising the next generation was done at or above 32 weeks of age. Fresh fertile eggs collected within 10 days from different selected hens were pooled and incubated at once. The second generation consisted of 181 chicks at hatching. They comprised 55 cockerels and 85 hens at 8 weeks of age and were evaluated following the same protocol as in the first generation.

Indigenous hens—generation one

Indigenous hens—generation two

Indigenous cocks
Data collection
Data were collected across two consecutive generations of indigenous chickens of both sexes raised at the Gulu University Poultry Unit. The live weight of chicks at hatching (HW) as well as at 2 (W2), 4 (W4), 6 (W6), 8 (W8), and 12 (W12) weeks of growth were measured using a portable electronic scale (Model: TS200, Canada). Shank lengths (SL) were also measured at 2 (SL2), 4 (SL4), 6 (SL6), 8 (SL8), and 12 (SL12) weeks of growth using a Tailor’s measuring tape as described by the African Union Inter-African Bureau for Animal Resources (AU-IBAR 2015) field manual. Locally made trap nests were used to collect data on egg production traits. The reproductive traits, such as body weight of hens at on-set of lay (WFE), egg weight (EW), clutch number (CLN-60), clutch size (CLS-60), and number of eggs laid within 60 days (EN-60), were recorded.
Data analysis
Descriptive statistics of productive and reproductive traits were carried out using R software package, version 4.1.1 (R Core Team 2021). Heritability of traits was estimated using equation.
where h2 is the broad sense heritability, \({\sigma}_g^2\) is the genotypic variance, and \({\sigma}_p^2\) is the phenotypic variance.
Genetic analysis was performed using univariate animal model with the restricted maximum likelihood program (Dana et al. 2011).
Correlations among traits were estimated by the animal model for traits i (i = 1 and i = 2) using the variability package of R, version 4.1.1 (R Core Team 2021).
where yi is the vector of observations; bi is the vector of fixed effects of generation; ai is the vector of random direct genetic effects; ei is the vector of random residual effects; and Xi and Zi are the incidence matrices relating the observations to the respective fixed and direct genetic effects for trait i.
Results
Averages, variance components, and heritability of traits
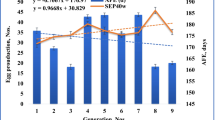
The averages, genetic variance, phenotypic variance, and heritability of productive and reproductive traits of indigenous hens under a supplemented scavenging system are summarized in Table 1. Starting with a hatching weight (HW) of 25.35 g, chicken gained up to 1498.57 g of body weight at on-set of lay (WFE). Shank length (SL) increased from 3.38 cm at 2 weeks to 7.98 cm at 12 weeks of growth. On average, hens laid 19.29 eggs within a period of 60 days, spread across 1.90 clutches of 8.70 eggs each. Heritability ranged from 0.02 to 0.72. Majority of the traits had values above 0.3 which is moderate and adequate for targeted increased genetic improvement. Low heritability values were estimated for trait SL4 (0.02), SL12 (0.14), and EN-60 (0.17). Body weight at 2 weeks of growth (W2) was highly heritable (0.72) compared to other traits in the study.
Genetic and phenotypic correlation of traits
Genetic and phenotypic correlation coefficients of productive and reproductive traits of IC are summarized in Table 2. Most of the phenotypic correlation coefficients were positive, significant, and ranging from moderate to high. The highest significant (P < 0.001) phenotypic correlation coefficient (0.83) was between SL2 and W2. The lowest (P < 0.001) and only negative phenotypic correlation coefficient (− 0.49) was between EN-60 and W4. Weight at first egg (WFE) was highly correlated with W2 (0.56), SL4 (0.40), W6 (0.45), SL6 (0.70), W8 (0.52), SL8 (0.81), and SL12 (0.38). Egg weight (EW) was highly correlated with W2 (0.45), SL6 (0.51), SL8 (0.54), and WFE (0.65). Significant genetic correlation coefficients ranged from − 0.96 (SL12-EN-60) to 1.04 (HW-SL2). Weight at first egg (WFE) was highly (P < 0.01) and positively genetically correlated (0.99) with SL8. Egg number (EN-60) was significantly (P < 0.01) negatively genetically correlated with shank length (SL12).
Discussion
Averages of phenotypic traits
The average body weight at 0, 2, 4, 8, and 12 weeks of age in the present study were higher than that of indigenous chicken Ecotypes in Eastern Uganda (Semahoro et al. 2018). In the current study, chicken were raised under a supplementary scavenging system different from the intensive system used by Semahoro et al. (2018). Probably, the disparity in body weight could be attributed to the management system and nutritional profile of feeds used in the two studies. In the current study, the mean shank length of hens at 8 to 12 weeks of age was higher than the values estimated for Nganda chickens within the same age range in Central Uganda (Semakula et al. 2011). The variation in shank length across different studies is a result of the differences in field guidelines used in collecting data on morphometric measurements (Beyihayo et al. 2022). The body weight of hens at onset of lay was within the range of 1200 and 1500 g reported for adult hens in Eastern and Western ecological zones of Uganda (Ssewannyana et al. 2008). The mean weight of eggs (37.34 g) was, however, lower than the value of 41.13 g recorded for eggs laid by scavenging hens in rural households (Beyihayo et al. 2022). In the current study, eggs were weighed at the on-set of lay, a period characterized by small and lightweight eggs (Anene et al. 2020; Udeh 2009). The average clutch size of 8.70 eggs per clutch in the present study was outside the range of 13 to 15 eggs reported for hens in Northern (Nakkazi et al. 2014), Western, and Eastern (Ssewannyana et al. 2008) ecological zones of Uganda. The small clutch size in the present study could be attributed to the short resting period between clutches as hens were not allowed to incubate and brood chicks. In addition, phenotypic differences in traits could be attributed to variations in feeding regimes and chicken Ecotypes used in various performance studies.
Heritability of traits
Heritability values of 0.72, 0.67, 0.30, and 0.51 in the current study were close to the values of 0.66, 0.55, 0.52, and 0.63 for body weight at 2, 4, 6, and 8 weeks, respectively estimated for local chickens in Ghana (Osei-Amponsah et al. 2013). The heritability of body weight at 4 and 8 weeks of age in the current study is similar to the values estimated for Indian Ghagus chicken (Haunshi et al. 2022). Like in the current study, a similar body weight heritability value of 0.55 was estimated for Fars chickens at 12 weeks of age but lower values of 0.25 and 0.23 were reported for Iranian Azarbaijan and Esfahan Ecotypes (Ghazikhani et al. 2007). Likewise, body weight heritability of 0.72 and 0.67 in the current study differed from the estimates of 0.45 and 0.38 at 2 and 4 weeks of age, respectively (Chomchuen et al. 2022). However, a similar heritability of 0.3 at week 6 of growth was recently reported (Chomchuen et al. 2022). In related studies, Adeyinka et al. (2006) and Norris and Ngambi (2006) reported similar heritability values at hatching and a low value at the fourth week of growth. The decline in body weight heritability with increasing age was attributed to the corresponding reduction in maternal effect (Tongsiri et al. 2019). Heritability estimates for shank length in the present study ranged from 0.02 and 0.58 and are in agreement with the values of 0.57, 0.3, and 0.58 at 2, 6, and 8 weeks of age, respectively estimated for local chickens in Ghana (Osei-Amponsah et al. 2013).
The heritability value of 0.47 for egg weight in the current study was similar to 0.46 estimated for Mazandaran chicken but varied from 0.64, 0.22, and 0.25 associated with Far, Azarbaijan, and Esfahan chicken Ecotypes, respectively (Ghazikhani et al. 2007). The estimated heritability of egg number (0.17) was close to 0.15 reported by Tongsiri et al. (2019) and within a range of 0.16 to 0.18 for Mazandaran and Esfahan chickens in Iran (Ghazikhani et al. 2007). The heritability value of 0.17 for egg number highly varied from the average of 0.36 and 0.32 estimated for Ethiopian Horro chickens (Dana et al. 2011) and Iranian Fars chickens (Ghazikhani et al. 2007). The variation in heritability estimates of traits reported in different studies is attributed to the method of estimation (Ndung et al. 2020) and number of generations considered (Haunshi et al. 2022). The trait of body weight could be adequately improved by direct selection except for egg number which is associated with low heritability. Tongsiri et al. (2019) suggested the use of a multiple trait selection index method for improved egg number.
Genetic correlation of traits
Egg number and shank length at 12 weeks of age are negatively genetically correlated. The negative genetic correlation implies that hens with short shanks at 12 weeks are more likely to lay more eggs, and the reverse is true. On the other hand, shank length at 12 weeks of age is positively genetically correlated with hatching weight, implying that light chicks at the time of hatching are expected to have short shanks at 12 weeks of age and more likely to lay more eggs compared to heavy chicks. This implies that hens can probably be indirectly selected for large clutch size using shank length at 12 weeks as a phenotypic marker trait.
In the current study, egg number and weight at onset of lay were negatively genetically correlated with a coefficient of − 0.26. Likewise, Ghazikhani et al. (2007) and Tongsiri et al. (2019) reported a similar negative value of genetic correlation between egg number and weight at onset of lay. Additionally, Zonuz et al. (2013) reported negative genetic correlations of egg number and body weight at 0 (− 0.04), 8 (− 0.09), and 12 (− 0.14) weeks of growth. The antagonistic pleiotropic relationship involving three candidate genes, namely, CPEB3, MAST2, and CACNA1H, is responsible for the negative cross-phenotypic association between body weight and egg production traits (Tarsani et al. 2021). Probably, a gene controlled biological mechanism in light hens directs abundant nutrients for egg production as compared to body maintenance. Unlike most studies, Dana et al. (2011) reported positive genetic correlation between egg number and body weight excluding a negative correlation (− 0.16 and − 0.54) at 2 and 6 weeks of age. The positive genetic correlation could be due to errors introduced by the high mortality of 13 to 29% reported during the laying period. The genetic correlation coefficient between weight at onset of lay and egg weight was 0.908 and was higher than 0.6 reported by (Tongsiri et al. 2019). However, negative genotypic correlation coefficients were reported for egg number against egg weight (− 0.39) and weight at onset of lay (− 0.28) among local chickens in Nigeria (Oleforuh-okoleh 2011).
Phenotypic correlation of traits
Weight at onset of lay and shank length at 8 weeks of age in the current study were moderate, positively correlated, and in agreement with estimates reported for Ghagus chicken in India (Haunshi et al. 2022). Therefore, heavier hens at onset of lay can be indirectly selected using shank length at 8 weeks as a marker trait. The phenotypic and genotypic correlation coefficients between shank length and live weight were positive, significant, and similar to the values of 0.86 and 0.94, respectively, reported for local chickens in Ghana (Osei-Amponsah et al. 2013). The same study (Osei-Amponsah et al. 2013) reported high phenotypic correlation coefficients between body weight and shank length at 6 (0.93) and 12 (0.87) weeks of age. In a recent study involving Alpha and Noiler chicken genotypes, relatively lower genetic correlations of 0.59 and 0.71 were reported for body weight and shank length at 8 and 12 weeks of age, respectively (Sanda et al. 2022). Genetic correlations are indicators of pleiotropic association and chromosomal linkage of genes affecting a pair of traits (Sanda et al. 2022). The disparities in phenotypic and genotypic correlation coefficients reported from different studies are due to the influence of the chicken breeds, rearing environment (Chomchuen et al. 2022; Sanda et al. 2022).
Conclusions
Genetic improvement of IC with respect to shank length and body weight at onset of lay as well as egg weight but not egg number can be achieved through direct selection of superior chickens. Shank lengths at 8 and 12 weeks of age are potential phenotypic markers when selecting for both high body weight at onset of lay and egg yield. Breeders could, for instance, undertake selection of hens for higher body weight at onset of lay based on shank length. Light hens with short shanks are more likely to lay more eggs per clutch. Trait pairs including SL8-WFE, EW-WFE, SL2-W2, and SL8-W8 are, therefore, positively correlated and can be considered together in a breeding program. The significant positive phenotypic correlation between shank length and body weight permits indirect selection and improvement of the later. Further studies are required to estimate the genetic parameters of other reproductive traits such as broodiness, hatchability, and mothering ability in indigenous hens.
Data availability
The datasets generated are available from the corresponding author on request.
References
Adeyinka, I.A., Oni O.O., Nwagu, B.I. and Adeyinka, F.D., 2006. Genetic parameter estimates of body weights of naked neck broiler chickens. International Journal of Poultry Science, 5(6), 589–592. https://doi.org/10.3923/ijps.2006.589.592
Adoligbe, C., Fernandes, A., Osei-amponsah, R. and Adje, N.D., 2020. Native chicken farming : A tool for wealth creation and food security in Benin. International Journal of Poultry Science, 11, 146–162. https://doi.org/10.5897/IJLP2020.0716
Anene, D. O., Akter, Y., Thomson, P.C., Groves, P., and O’shea, C.J., 2020. Variation and association of hen performance and egg quality traits in individual early-laying isa brown hens. Animals, 10(9), 1–14. https://doi.org/10.3390/ani10091601
AU-IBAR, 2015. Pictorial field guide for linear measurements of animal genetic resources. Nairobi, Kenya: African Union Inter-African Bureau for Animal Resources (AU-IBAR)
Beyihayo, G.A., Ndyomugyenyi, E.K., Echodu, R. and Kugonza, D., 2022. In-situ morphological characterization of indigenous chicken Ecotypes in Uganda. Ecological Genetics and Genomics, 24(June). 100129. https://doi.org/10.1016/j.egg.2022.100129
Chomchuen, K., Tuntiyasawasdikul, V., and Chankitisakul, V., 2022. Comparative study of phenotypes and genetics related to the growth performance of crossbred Thai indigenous ( KKU1 vs . KKU2 ) chickens under hot and humid conditions. Veterinary Science, 9(6), 263. https://doi.org/10.3390/vetsci9060263
Dana, N., van der Waaij, L.H., Dessie, T. et al. 2010. Production objectives and trait preferences of village poultry producers of Ethiopia: implications for designing breeding schemes utilizing indigenous chicken genetic resources. Tropical Animal Health and Production, 42, 1519–1529. https://doi.org/10.1007/s11250-010-9602-6
Dana, N., Waaij, E.H., Arendonk, J.A. and Van, M., 2011. Genetic and phenotypic parameter estimates for body weights and egg production in Horro chicken of Ethiopia. Tropical Animal Health and Production, 43, 21–28. https://doi.org/10.1007/s11250-010-9649-4
Elbeltagy, A.R., Bertolini, F., Fleming, D.S., Van Goor, A., Ashwell, C.M., Schmidt, C.J., Kugonza, D.R., Lamont, S.J. and Rothschild, M.F., 2019. Natural selection footprints among African chicken breeds and village ecotypes. Frontiers in Genetics, 10, 1–16. https://doi.org/10.3389/fgene.2019.00376
Fleming, D.S., Koltes, J.E., Markey, A.D., Schmidt, C.J., Ashwell, C.M., Rothschild, M.F., Persia, M.E., Reecy, J.M. and Lamont, S.J., 2016. Genomic analysis of Ugandan and Rwandan chicken ecotypes using a 600 k genotyping array. BMC Genomics, 17(1), 1–16. https://doi.org/10.1186/s12864-016-2711-5
Ghazikhani, A.S., Nejati, A.J. and Mehrabani, Y.H., 2007. Animal Model Estimation of Genetic parameters for most important economic traits in Iranan native fowls. Pakistan Journal of Biological Sciences, 10(16), 2787-2789. https://scialert.net/abstract/?doi=pjbs.2007.2787.2789
Gwaza, D.S., Dim, N.I. and Momoh, O.M., 2016. Genetic improvement of egg production traits by direct and indirect selection of egg traits in Nigerian local chickens. Advancements in Genetic Engineering, 5, 1-7. https://doi.org/10.4172/2169-0111.1000148
Haunshi, S., Ullengala, R., Prince, L. L., Ramasamy, K., Gurunathan, K., Devatkal, S. and Chatterjee, R. N., 2022. Genetic parameters of growth traits, trend of production and reproduction traits, and meat quality status of Ghagus, an indigenous chicken of India. Tropical Animal Health and Production, 54(3), 170. https://doi.org/10.1007/s11250-022-03166-y
Lubandi, C., Lwasa, S., Kugonza, D.R., Babigumira, B.M., Semahoro, F., Nadiope, G. and Okot, M.W., 2018. Analysis of the indigenous chicken value chain in Uganda. African Journal of Rural Development, 3 (3), 895-912.
Magothe, T.M., Muhuyi, W.B. and Kahi, A.K., 2010. Influence of major genes for crested-head, frizzle-feather and naked-neck on body weights and growth patterns of indigenous chickens reared intensively in Kenya. Tropical Animal Health and Production, 42(2), 173–183. https://doi.org/10.1007/s11250-009-9403-y
Magothe, T.M., Muhuyi, W.B. and Kahi, A.K., 2007. Genetic Parameter Estimates for Egg and Body Weights of Indigenous Chicken in Kenya. https://lrrd.cipav.org.co›lrrd27›shar27178
Mahendra, K.P., 2016. Importance of indigenous breeds of chicken for rural economy and their improvements for higher production performance. Scientifica 2016, 2604685. https://doi.org/10.1155/2016/2604685
Nakkazi, C., Kayitesi, A., Mulindwa, H.E., Kugonza, D.R., Okot. M.W., 2014. The status of local chicken (Gallus domesticus) production in Northern Uganda. LRRD http://www.lrrd.org/lrrd26/11/nakk26198.html
Ndung, C.W., Okeno, T.O. and Muasya, T.K., 2020. Pooled parameter estimates for traits of economic importance in indigenous chicken in the tropics. Livestock Science, 104102. https://doi.org/10.1016/j.livsci.2020.104102
Niknafs, S. and Nejati-javaremi, A., 2012. Estimation of genetic parameters for body weight and egg production traits in Mazandaran native chicken. Tropical Animal Health and Production, 44, 1437–1443. https://doi.org/10.1007/s11250-012-0084-6
Norris, D. and Ngambi, J.W., 2006. Genetic parameter estimates for body weight in local Venda chickens. Tropical Animal Health and Production, 38(7–8), 605–609. https://doi.org/10.1007/s11250-006-4420-6
Oleforuh-okoleh, V.U., 2011. Estimation of genetic parameters and selection for egg production traits in a Nigerian local chicken ecotype. Journal of Agricultural and Biological Science, 6 (12), 2006-2009.
Osei-Amponsah, R., Kayang, B. and Naazie, A., 2013. Phenotypic and genetic parameters for production traits of local chickens in Ghana. Animal Genetic Resources/Resources Génétiques Animales/Recursos Genéticos Animales 53, 45-50. https://doi.org/10.1017/S2078633613000271
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Rajkumar, U., Rama Rao, S.V., Raju, M. V. L. N. and Chatterjee, R.N., 2021. Backyard poultry farming for sustained production and enhanced nutritional and livelihood security with special reference to India: a review. Tropical Animal Health and Production, 53, 176. https://doi.org/10.1007/s11250-021-02621-6
Sanda, A.J., Bemji, M., Wheto, A., Oso, O., Sanda, M.O. and Olowofeso, O., 2022. Genetic and phenotypic correlation coefficients of quantitative traits in two chicken genotypes. Genetika 54(1), 307-328. https://doi.org/10.2298/GENSR2201307S
Semahoro, F., Kugonza, D.R.D., Babigumira, B.M.B., Walusimbi S., Nadiope, G. and Okot, M.W., 2018. Growth performance of chicken genotypes based on indigenous populations of Eastern Uganda. Proc WCGALP pp557 New Zealand.
Semakula, J., Lusembo, P., Kugonza, D.R., Mutetikka, D., Ssennyonjo, J. and Mwesigwa, M., 2011. Estimation of live body weight using zoometrical measurements for improved marketing of indigenous chicken in the Lake Victoria basin of Uganda. LRRD 23 Article #170. http://www.lrrd.org/lrrd23/8/sema23170.htm
Singh, M.K., Kumar, S., Sharma, R.K., Singh, S.K., Singh, B. and Singh, D.V., 2018. Research article heritability estimates of adult body weight and egg production traits in indigenous Uttara chickens. Genetics 10(2):357-359. https://doi.org/10.9735/0975-2862.10.2.357-359
Ssewannyana, E., Ssali, A., Kasadha, T., Dhikusooka, M., Kasoma, P., Kalema, J., Kwatotyo, B.A. and Aziku, L., 2008. On-farm characterization of indigenous chickens in Uganda. Journal of Animal & Plant Sciences, 1(2), 33–37. http://www.m.elewa.org/JAPS/2008/1.2/1.pdf
Tadelle, Y. and Alemu Peters, K.J., 2000. Indigenous chickens in Ethiopia: Genetic potential and attempts at improvement. Poultry Science Journal, 56(1), 45-54. https://doi.org/10.1079/WPS20000005
Tarsani, E., Kranis, A., Maniatis, G., Hager-Theodorides, A.L. and Kominakis, A., 2021. Detection of loci exhibiting pleiotropic effects on body weight and egg number in female broilers. Scientific Reports, 11(1), 7441-7452. https://doi.org/10.1038/s41598-021-86817-8. PMID: 33811218; PMCID: PMC8018976.
Tongsiri, S., Jeyaruban, G.M., Hermesch, S., Werf, J.H. and Van Der, J., 2019. Genetic parameters and inbreeding effects for production traits of Thai native chickens. Asian-Australas Journal of Animal Science, 32(7), 930–938.
Udeh, I., 2009. Influence of weight grouping on the short term egg production of two strains of layer type chicken. Animal Research International, 4(3), 741–744. https://doi.org/10.4314/ari.v4i3.48684
Wong, J.T., de Bruyn, J., Bagnol, B., Grieve, H., Li, M., Pym, R. and Alders, R.G., 2017. Small-scale poultry and food security in resource-poor settings: A review. Global Food Security, https://doi.org/10.1016/j.gfs.2017.04.003
Zonuz, A.Y., Alijani, S., Mohammadi, H., Rafat, A. and Kia, H.D., 2013. Estimation of genetic parameters for productive and reproductive traits in Esfahan native chickens. Journal of Livestock Science and Technologies, 1(2), 34-38. https://doi.org/10.22103/jlst.2013.5501
Acknowledgements
The authors sincerely appreciate Aber Cinderella, Akello Stella Josephine, and Ben Kaggwa for their assistance in rearing the chickens.
Funding
This work was supported by the African Union (AU) through the Native Chicken Project (grant number AURG II-2-115-2018).
Author information
Authors and Affiliations
Contributions
GAB and DRK conceptualized the study and methodology. MWO, FDA, and DRK mobilized resources. GAB collected and analyzed data and prepared initial draft of manuscript. EKN, DRK, and RO reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
The animal study protocol was approved by the Research Ethics Committee of Gulu University (GUREC-2020-18; dated: 03/12/2020). The research protocol was registered with the Uganda National Council for Science and Technology (UNCST) for final clearance (NO. A154ES; dated: 15/11/2021).
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Beyihayo, G.A., Kugonza, D.R., Ndyomugyenyi, E.K. et al. Genetic and phenotypic parameter estimates for selection within Ugandan indigenous chickens. Trop Anim Health Prod 55, 100 (2023). https://doi.org/10.1007/s11250-023-03513-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11250-023-03513-7