
Abstract
The recent incorporation of immune checkpoint inhibitors targeting the PD-1 (programmed cell death receptor 1) and CTLA-4 (cytotoxic T lymphocyte antigen 4) pathways into the therapeutic armamentarium of cancer has increased the need to understand the correlation between the immune system, autoimmunity, and malignant neoplasms. Both autoimmune thyroid diseases and thyroid cancer are common clinical conditions. The molecular pathology of autoimmune thyroid diseases is characterized by the important impact of the PD-1/PD-L1 axis, an important inhibitory pathway involved in the regulation of T-cell responses. Insufficient inhibitory pathways may prone the thyroid tissue to a self-destructive immune response that leads to hypothyroidism. On the other hand, the PD-1/PD-L1 axis and other co-inhibitory pathways are the cornerstones of the immune escape mechanisms in thyroid cancer, which is a mechanism through which the immune response fails to recognize and eradicate thyroid tumor cells. This common mechanism raises the idea that thyroid autoimmunity and thyroid cancer may be opposite sides of the same coin, meaning that both conditions share similar molecular signatures. When associated with thyroid autoimmunity, thyroid cancer may have a less aggressive presentation, even though the molecular explanation of this clinical consequence is unclear. More studies are warranted to elucidate the molecular link between thyroid autoimmune disease and thyroid cancer. The prognostic impact that thyroid autoimmune disease, especially chronic lymphocytic thyroiditis, may exert on thyroid cancer raises important insights that can help physicians to better individualize the management of patients with thyroid cancer.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
More than a century has passed since the first pathological description of malignant neoplasms infiltrated by leukocytes [1, 2]. Several studies have demonstrated the role of the immune system in cancer formation [3,4,5,6]. One such piece of evidence is the frequent coexistence of cancer and autoimmune diseases [7]. However, relatively few molecular mechanisms have been proposed to explain this relationship. The recent incorporation of immune checkpoint inhibitors targeting the PD-1 (programmed cell death receptor 1) and CTLA-4 (cytotoxic T lymphocyte antigen 4) pathways into the therapeutic armamentarium of cancer has increased the need to understand the correlation between the immune system, autoimmunity, and malignant neoplasms [3, 7, 8].
Autoimmune diseases occur when the immune response is directed toward self-antigens [7, 9]. Cancer occurs when the immune response fails to identify and eliminate the malignant cells [5, 7, 10]. These two phenomena, occurring as vectors in opposite directions, suggest that autoimmunity and cancer can be different sides of the same coin [7]. In contrast, data from clinical and epidemiological studies have indicated that the co-occurrence of autoimmune diseases and cancer is a frequent clinical event [11]. For example, patients with advanced-stage melanoma and concomitant vitiligo have better clinical outcomes [12]. One might argue that this concomitance could be a random effect. However, accumulating evidence suggests that cancer and autoimmune diseases share common underlying molecular mechanisms [13]. These data suggest that the concomitance of autoimmunity does not occur directly during the development of cancer. On the contrary, concomitant autoimmunity may help to modulate an antitumor immune response, favoring a better prognosis [14].
Autoimmune thyroid disease is the most common autoimmune disease in the field of internal medicine [15]. Thyroid cancer is also very common, its incidence is increasing, and both clinical conditions seem to be closely associated [15]. Would this relationship be causal? Next, we review the molecular mechanisms underlying autoimmune thyroid disease. Subsequently, we revisit the clinical implications of the co-occurrence of autoimmune thyroid diseases and thyroid cancer, exploring the molecular mechanisms that may help explain the relationship between the two.
2 Physiopathology of autoimmune thyroid disease
Autoimmune thyroid diseases (AITD) such as Graves' disease (GD) and Hashimoto’s thyroiditis (HT) are the most prevalent organ-specific autoimmune conditions in humans [15]. Hashimoto thyroiditis, also known as chronic lymphocytic thyroiditis, is the most prevalent cause of hypothyroidism in adults, particularly in women aged between 45 and 65 years [16, 17]. The NHANES III study found a 4.6% prevalence of hypothyroidism and circulating anti-thyroid peroxidase antibodies in 10% of over 13,000 healthy individuals examined, suggesting that humoral immune response against the thyroid is more prevalent than thyroid dysfunction [18]. A recent meta-analysis involving 103,341 subjects from several European studies found a prevalence of hypothyroidism of 4.94% in the population, of which 93% were cases of subclinical hypothyroidism [19, 20].
The gross pathology of chronic lymphocytic thyroiditis is characterized by increased volume or even diminished and hardened volume [21]. The thyroid capsule is usually intact, and the thyroid parenchyma is pale hue. The thyroid immune microenvironment in chronic lymphocytic thyroiditis is enriched with lymphocytes, plasma cells, and germinal centers [21]. Germinal centers are prototypical tertiary lymphoid organs that form in peripheral and non-lymphoid tissues in response to chronic inflammation. They are collections of immune cells that resemble secondary lymphoid organs, such as lymph nodes [22]. The germinal centers present an upregulation of Lymphotoxin-alpha, CXCR4, CXCR5, CXCL13, and CCL19, all of which are related to the secretion of cytokines that attract immune cells to the milieu [23]. Thyroid follicles undergo an intense process of atrophy, and epithelial cells are noted in the presence of abundant eosinophilic cytoplasm rich in mitochondria. Histopathology is characterized by abundant connective tissue in the thyroid parenchyma, which marks the chronic replacement of follicular cells with fibrous tissue [21].
The molecular pathology is characterized by the important impact of the PD-1/PD-L1 axis [24]. The PD-1/PD-L1 axis is an important inhibitory pathway involved in the regulation of T-cell responses and is frequently involved in autoimmune processes [25]. In fact, PD-L1, but not PD-L2, is expressed in more than 80% of Graves’ disease glands and only in 25% of non-autoimmune glands, suggesting that the PD-1/PD-L1 axis is activated in Graves’ disease [24]. The authors showed that PD-L1 was expressed by thyroid cells in areas that also contain abundant PD-1 positive T cells, but the co-localization in thyroid follicular cells indicated only partial overlap between the smaller areas of PD-L1+ and the larger areas of HLA class II+ expression, suggesting an insufficient containment of the autoimmune process by partial inhibition of PD-L1 in Graves’ disease and chronic lymphocytic thyroiditis glands [24]. Recently, in silico analysis has been used for the molecular characterization of the immune microenvironment of chronic lymphocytic thyroiditis, comparing non-infiltrated thyroids to small focal infiltrated thyroids and extensive lymphoid infiltrated thyroids [26]. PD-1/PD-L1 axis and other inhibitory immune checkpoint receptors expressed by both B and T lymphocytes are broadly represented by the upregulation of genes in the thyroid tissue. Glands with large infiltrates are enriched with follicular helper and memory CD4 T cells and are markedly infiltrated by naive B cells [24, 26].
The immune response against thyroid cells results in the gradual destruction of the thyroid gland, leading to hormonal insufficiency, which is clinically recognized as hypothyroidism [27]. In addition, some autoantibodies may have an affinity for the TSH receptor, exerting an inhibitory function, suggesting that the histological destruction of the thyroid is not a unique mechanism of hormonal insufficiency [28].
Chronic lymphocytic thyroiditis is a multifactorial disease and includes both environmental and endogenous factors. Patients who live in sufficient areas in iodine, as well as increased intake of dietary iodine, are at increased risk of developing chronic lymphocytic thyroiditis [29]. Patients who present some haplotypes of HLA, mutations in TCR receptors or even mutations in genes such as CTLA-4 and PTPN22 appear to add individual risk of developing thyroiditis chronic lymphocytic disease [30]. Clinical risk factors include female sex and comorbidities such as Down Syndrome and Turner Syndrome [31]. It has long been recognized that patients with chronic lymphocytic thyroiditis are more susceptible to the development of other autoimmunities, both endocrine and non-endocrine. It is possible that HT and thyroid cancer share some common molecular pathophysiological mechanisms, as explained below.
RNA can be classified into two main types: protein-coding RNA (messenger RNA, mRNA) and non-coding RNA (ncRNA). Even though these do not encode proteins, they play a variety of important roles in gene regulation, RNA processing, and other cellular functions. ncRNAs can be further classified according to their length. Long ncRNAs (lncRNAs), short ncRNAs (sncRNAs), and miRNAs, that are the most widely studied short ncRNAs. miRNA can be assessed in different compartments of the body of patients with AITD [32]. Bernecker et al. [33] investigated miRNAs amplified from peripheral blood mononuclear cells (PBMCs) and purified CD4+ and CD8+ T cells of patients with Grave’s disease or Hashimoto’s thyroiditis, and compared healthy controls. Both GD and HT showed significantly decreased expression of miRNA 200a_1 and miRNA 200a2* in CD4+ T cells and CD8+ T cells. The exact role of this miRNA in CD4+ and CD8+ cells remain unclear. Some studies have suggested that miRNA200a promotes cell proliferation, metastasis, epithelial-mesenchymal transition, and anticancer drug resistance in many histological types of cancer [34]. However, it is not clear whether the decrease in the expression of miRNA 200a influences the autoimmunity process. GD and HT showed significantly decreased expression of miRNAs 155_2 and 155*_1 in HT in CD8+ T cells. Thai et al. [35] showed that miRNA-155 plays an important role in the immune system, by specifically regulating T-helper cell differentiation and the germinal center construction, to produce an optimal T-cell-dependent antibody response. MiRNA 155 (miR-155) is the most expressed miRNA in the thymus and spleen, and its role has already been demonstrated in mature peripheral T cells [36]. In addition, miRNA 155 is the most expressed miRNA in the thymus and spleen, and its role has already been demonstrated in mature peripheral T cells. In the thymus, miRNA 155 may promote migration and chemotaxis, which are crucial for immune development [36]. miRNA 155 may be involved in central tolerance to autoantigens, justifying the potential role of this molecule in autoimmune thyroid diseases. Martinez-Hernandez et al. [37] performed an analysis of the miRNA integrated with gene expression, and discovered that atypical ciliogenesis can be a novel mechanism that is involved in the molecular pathology of autoimmune thyroid diseases.
Exosomes are small cell membrane vesicles that enclose genetic material, mainly microRNAs. These vesicles are produced by cells of different origins and can target cells at long distances, which, once they receive these exosomes, accept the miRNA content, and have different modulated genes. Thus, exosomes are powerful communication mechanisms between cells and are fundamental to many biological processes [38]. Exosomes generated from immune and non-immune cells play essential roles in the immunopathogenesis of autoimmune [39], infectious, and neoplastic diseases [40, 41]. Plasma from patients with autoimmune thyroid diseases is enriched with exosomes derived from platelets and depleted in exosomes derived from leukocytes and endothelium [42]. The exosomes of these patients are full of miR-146a and miR-155 [43]. These microRNAs target the IL-8 and SMAD4 genes, which are reduced in PBMCs [43]. When subjected to functional assays, exosomes from patients with autoimmune thyroid diseases were able to inhibit the differentiation of regulatory T lymphocytes, which is consistent with the decrease in peripheral tolerance of these patients [43]. On the other hand, exosomes from patients with autoimmune thyroid diseases promoted the expression of interferon-γ by CD4+ lymphocytes and induced the differentiation of Th17 lymphocytes, a subset of lymphocytes strongly involved in immunopathogenesis of autoimmune thyroid diseases [43].
3 Epidemiological and clinical aspects of thyroid autoimmunity and differentiated thyroid cancer
Both autoimmune thyroid diseases and thyroid cancer are common clinical conditions. Epidemiological series indicate an increasing incidence of both in the last decade, suggesting that they are associated [11, 44, 45]. However, it is still controversial whether these increases are just a chance phenomenon or whether they share common molecular mechanisms.
The first question is whether patients with autoimmune thyroid diseases present a higher risk of thyroid cancer. The most frequent association is observed with papillary thyroid cancer (PTC). The risk of medullary thyroid cancer appears to be anecdotal, which is further discussed below. It is generally accepted that patients with chronic low-grade inflammation are at risk of malignant neoplasms, including in the endocrine glands [46]. In a cross-sectional study, Keefe et al. [47] investigated 458 nodules of 385 pediatric patients. Autoimmune thyroid diseases were independently associated with an increased risk of thyroid cancer, mainly papillary thyroid cancer with unaggressive histology. The authors did not find any association between the extent of local, regional, or distant disease and thyroid cancer diagnosis. In a retrospective investigation, Janús et al. [48] confirmed the coexistence of PTC and autoimmune thyroid diseases in almost 50% of patients. The percentage of autoimmune thyroid diseases increased with age according to the pubertal stage. The same group prospectively evaluated this cohort of pediatric patients [49]. They observed that the use of thyroid US focused on the search for developing nodules in the routine follow-up of patients with autoimmune thyroid diseases may not only help to detect thyroid malignancies earlier but may also influence the aggressiveness of cancer management and reduce the side effects of 131I therapy. However, this observation was based on a small cohort of pediatric patients and more investigations are warranted to extrapolate this conclusion to clinical practice.
Regarding the care of adults, as early as 1955, Dailey et al. [50] reported a high frequency of chronic thyroiditis in pathological samples of differentiated thyroid tumors, specifically papillary thyroid carcinomas. Although large prospective series have failed to demonstrate an increase in the incidence of thyroid cancer among patients with AITD [51], the coexistence of both phenomena has been widely reported, suggesting that the association is not a fortuitous coincidence [52]. A large systematic review with meta-analysis that included 38 studies and more than 10,000 patients demonstrated that histologically proven chronic lymphocytic thyroiditis was observed more frequently in PTCs than in benign thyroid diseases and other carcinomas, reinforcing that this co-occurrence may reflect common pathophysiological origins [53]. Interestingly, another recent systematic review with meta-analysis that examined 11 case-control studies and 12 cohort studies revealed that chronic lymphocytic thyroiditis is a risk not only for thyroid cancer but also for breast cancer, lung cancer, cancer of the digestive system, urogenital cancer, blood cancer, and prolactinoma [11].
The second most important question is whether concurrent chronic lymphocytic thyroiditis is associated with aggressiveness and prognosis in patients with papillary thyroid carcinoma. We previously investigated 173 patients with differentiated thyroid cancer who underwent surgery complemented by radioiodine ablation [45]. Analysis of the prognostic factors revealed that the presence of antibodies and the previous history of autoimmune disease correlated negatively with the occurrence of death, metastasis, and/or recurrence [45]. Our data suggests that autoimmune activity against the thyroid gland, even against the non-malignant thyroid gland, may exert a protective effect on the outcome of differentiated thyroid carcinoma patients [54].
In a systematic review with meta-analysis, Lee et al. [53] confirmed that PTCs with coexisting chronic lymphocytic thyroiditis were significantly related to female patients, multifocal involvement, no extrathyroidal extension, no lymph node metastasis, and long recurrence-free survival. In a retrospective analysis of PTC with no extrathyroidal extension, Marotta et al. [55] also observed that chronic lymphocytic thyroiditis was associated with female gender, smaller tumor size, lower rate of aggressive PTC variants, and less frequent post-surgery radio-iodine administration. Consequently, patients with thyroid autoimmunity presented a higher rate of clinical remission [56]. However, some authors have reported an association between concomitant chronic lymphocytic thyroiditis and aggressive characteristics. These reports are restricted to specific groups, such as young patients, and do not report an implication in poor prognosis [57, 58]. Almost ten years after Lee's article, Osborne et al. [59] published an updated systematic review looking at the clinical impact of chronic lymphocytic thyroiditis on papillary thyroid carcinoma. At this time, 41,646 patients were included. Patients with concomitant thyroiditis more often had multifocal tumors and were considered for total thyroidectomy [60]. The presence of concomitant thyroiditis was associated with smaller tumor size, reduced angioinvasion, and decreased occurrence of lymph node metastases [59]. It is noteworthy that patients with chronic lymphocytic thyroiditis may have enlarged lymph nodes due to the inflammatory process, and caution should be exercised when indicating neck dissection [61]. In addition, the presence of antithyroid antibodies should alert the physician to possible interference in the interpretation of serum thyroglobulin values, since antithyroid autoantibodies affect thyroglobulin immunometric assays, causing falsely low results [62].
Among thyroid antibodies, the role of antithyroglobulin antibody (TgAb) stands out, which is useful in monitoring patients with papillary thyroid carcinoma to assess the initial treatment response [63, 64], as TgAb production depends on the amount of thyroid tissue secreting thyroglobulin [65]. Thus, with the destruction of thyroid tissue or thyroidectomy, a decrease in the levels of this antibody is expected. Therefore, it is possible to classify the initial treatment response with thyroidectomy using TgAb levels as one of the criteria: in patients with an excellent response, TgAb becomes undetectable, whereas, in patients with an indeterminate response, these levels remain stable or decrease but are still detectable [63]. In patients with an incomplete response, if imaging remains unchanged but TgAb levels continue to rise, it is considered an incomplete response, indicating ongoing biochemical disease [63]. Thus, in patients where TgAb concentration decreases by more than 50% after surgery, the risk of persistence/recurrence is very low; the risk increases if levels do not drop below 50% and maybe even higher if TgAb concentration increases [64, 66,67,68,69,70]. It is important to consider that TgAb may transiently increase postoperatively as an apparent immune response to surgery and may also increase after radioiodine ablation [71]. It is worth to note that the serum concentration of TgAb immediately after total thyroidectomy should not determine the need for radioactive iodine ablation, and the trend during follow-up can influence the risk of recurrence [72].
4 Molecular phenomena behind thyroid autoimmunity and differentiated thyroid cancer
The synchronous appearance of chronic lymphocytic thyroiditis and papillary thyroid cancer suggests a molecular and immunological link between the two entities. Indeed, many malignancies and chronic inflammation share etiopathogenic components. It is well established that infectious agents such as Helicobacter pylori and hepatitis C virus may promote both chronic inflammation and malignant transformation (i.e., gastric cancer and hepatocellular carcinoma, respectively [73]). Hepatitis C virus may infect thyroid cells [74] and trigger the production and secretion of interleukine-8 [75], a pro-inflammatory cytokine related to both thyroid autoimmunity and thyroid cancer [76]. We previously demonstrated that an inherited cancer-prone genetic profile may favor HHV-7 infection and maintenance, which, in turn, may initiate and perpetuate autoimmune in the thyroid gland [77]. We further investigated the cosmopolitan viral milieu in the thyroid cancer microenvironment and observed no evidence of HSV-2 or CMV DNA, but found EBV DNA sequences in 16% of thyroid cancer samples [78]. EBV infection may correlate to both chronic lymphocytic thyroiditis and thyroid tumors [79]. The presence of high EBV copy numbers in thyroid neoplasms and the fact that EBER expression is restricted to malignant cells suggest a strong association between this virus and the malignant transformation of follicular cells [78]. Thyroid cancer cells are permissive to EBV infection and the virus can distort oncogene expression, reinforcing that these effects may be related to different patterns of tumor progression [80].
Considering the immunologic link between chronic lymphocytic thyroiditis and thyroid cancer, it is important to distinguish leukocyte infiltration in the tumor from concomitant thyroiditis. Patients with concomitant chronic lymphocytic thyroiditis show a different pattern of leukocyte tumor infiltration, characterized by an enrichment of macrophages, CD3+, CD8+, CD20+, and FoxP3+ cells [81], suggesting that concomitant autoimmunity may influence the immune microenvironment of thyroid cancer [82]. Papillary thyroid carcinomas with concomitant chronic lymphocytic thyroiditis may be enriched with cytotoxic lymphocytes, reflecting a local antitumor immune activity that may help explain the protective effect of concomitant chronic lymphocytic thyroiditis on the progression of papillary thyroid carcinoma [83].
Thyroid parenchyma affected by autoimmunity is rich in pro-inflammatory cytokines such as IL-17, IL-21, and IL-22 [84, 85]. Follicular thyroid cells can produce IL-17 in both autoimmunity and neoplastic contexts [86, 87]. All these pro-inflammatory cytokines are related to the differentiation of Th17 and Th22 lymphocytes, which are essential to the chronification of inflammatory processes in the thyroid gland [84]. RORγt is a central molecule to Th17 differentiation. Zeng et al. [88] demonstrated that papillary thyroid cancer cells could induce initial T lymphocytes to differentiate into Th17 cells by the production of RORγt. It may independently modulate both the presence of concurrent autoimmunity and the antitumor immune response, since its expression in the tumor microenvironment of PTC patients with or without concurrent autoimmunity may inhibit lymph node metastasis [87]. Therefore, RORγt production by tumor cells may favor the migration of immune cells to the thyroid gland, exerting an antitumor activity that improves the prognosis of these patients [87].
Another pathway that may link autoimmune thyroid disease and thyroid cancer is the inflammasome pathway [89]. IL-1β is an important pro-inflammatory mediator produced by endangered tissues, such as infected tissues, whose activity is controlled at different stages, such as expression, maturation, and secretion [90]. Inflammasomes control the maturation and secretion of IL-1β [91]. Inflammasomes can induce a programmed cell death called pyroptosis [92]. Some NOD-like receptors (NLR) family members have been reported to exhibit inflammasome activity in vitro [93]. Expression of NLR family pyrin domain containing 1 (NLRP1), NLRP3, CARD-domain containing 4 (NLRC4), absent in melanoma 2 (AIM2), the apoptosis-associated speck-like protein that contains a caspase recruitment domain (ASC), caspase-1, IL-1β, and IL-18 mRNA, and protein were significantly increased in thyroid tissues from patients with thyroiditis [94]. Enhanced posttranslational maturation of caspase-1, IL-18, and IL-1β was also observed. In addition, the clustering of pyroptosis-related genes were differentially expressed in thyroid cancer [94]. The pyroptosis-related risk signature may be an effective predictor of prognosis in thyroid cancer, suggesting a common mechanism shared by both chronic lymphocytic thyroiditis and thyroid cancer [95].
In addition to immunological mechanisms, thyroid autoimmunity and thyroid cancer have common molecular triggers. More than 20 years have passed since the first description of RET/PTC rearrangement activation in chronic lymphocytic thyroiditis [96], although this finding has not been uniformly replicated by other authors [97]. Likewise, papillary thyroid carcinoma and chronic lymphocytic thyroiditis co-express some other genes (i.e., TSHR, BACH2, FOXE1, RNASET2, CTLA4, PTPN22, IL2RA) and share several somatic mutations [98]. Bioinformatics and mRNA expression analysis identified that LTF and CCL21 expression is significantly increased in HT, but decreased in PTC, suggesting an underlying association between both [99]. LTF is an iron-binding protein that induces B cell migration and maturation and was previously described in chronic lymphocytic thyroiditis and papillary thyroid carcinoma [100,101,102]. CCL21 is a chemokine that binds to receptor CCR7 and was previously described to be involved in the promotion of tissue invasion, cell proliferation thyroid carcinoma growth, and dissemination [103].
Subhi et al. [104] performed an expression microarray analysis of papillary thyroid carcinoma samples with and without chronic lymphocytic thyroiditis looking for differentially expressed genes in both conditions. Genes associated with oxidative stress, reactive oxygen species, DNA damage, DNA repair, cell cycle, and apoptosis were also deregulated in both chronic lymphocytic thyroiditis and papillary thyroid carcinoma, reinforcing the conception that inflammatory response may promote alterations in a myriad of biofunctions related to cell proliferation and transformation.
Non-coding RNAs do not participate in protein synthesis, but they are important regulators of the expression of coding RNAs [105]. lncRNAs regulate gene expression by epigenetic, transcriptional, and post-transcriptional mechanisms [106]. miRNAs regulate gene expression by repressing translation or degrading target mRNA molecules [107]. We call endogenous competitive RNA (ceRNA) the molecular regulatory mechanism by which a lncRNA can competitively interact with a miRNA and inhibit its function [108]. Zhang et al. [109] constructed a ceRNA network based on miRNA-mRNA interactions by fusing with lncRNA-microRNA interactions collected from 112 patients with PTC, including 35 cases with coexisting HT. A total of 76 pairs of lncRNA-miRNA-mRNA ceRNA were obtained and their validation showed that mRNA levels of CXCL10, CXCL9, CCL5, and CCR2 were notably higher in tumor tissues from patients with PTC and HT than those of PTC without TH [109].
5 Particularities of medullary thyroid carcinoma
There is an extensive body of information and knowledge about the coexistence of papillary thyroid carcinoma, and chronic lymphocytic thyroiditis. We know that autoimmune thyroid disease is associated with lymphoma, papillary carcinoma and oncocytic thyroid cell neoplasms. However, there are few reports on the coexistence of medullary thyroid carcinoma and chronic lymphocytic thyroiditis [110,111,112,113]. Most of the articles are case reports of patients who have a collision between medullary thyroid carcinoma and papillary thyroid carcinoma with thyroiditis in the adjacent normal parenchyma [114, 115]. In a medical records review of 863 patients undergoing total thyroidectomy, Zayed et al. [116] suggested that there might be an association between medullary carcinomas and chronic lymphocytic thyroiditis, particularly in women, as 3 out of 9 patients with medullary carcinomas had coexisting Hashimoto's thyroiditis. An etiopathogenic hypothesis resides in the role of stem cells. Animal model studies suggest that the thyroid gland exposed to acute injury may be prone to regenerative activity with mobilization of resident stem cells [117]. These stem cells may be a common progenitor of papillary [118] and medullary thyroid carcinoma [119]. However, more investigations need to be done to clarify this potential mechanism.
6 Closing remarks
Autoimmune thyroid disease and thyroid cancer coexist in a significant number of patients. Most studies suggest that patients with papillary carcinoma who have concomitant chronic lymphocytic thyroiditis tend to have a less aggressive clinical picture at diagnosis, and a better prognosis at follow-up. The clinical impact of concomitant thyroiditis suggests that immunomodulation occurs in these patients favoring an immune microenvironment profile more prone to an effector antitumor response. However, further studies are needed to understand the molecular mechanisms that determine how immune effector cells are affected by autoimmunity. These studies may help tailor patient management and even reveal new therapeutic targets. Understanding the molecular relationship between thyroid cancer and thyroiditis may reveal new clinically useful biomarkers for diagnosing thyroid cancer, assessing disease severity and stratifying its risk, predicting, and guiding clinical decision

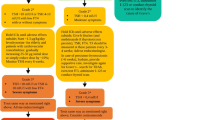
Figure 1 shows the molecular mechanism of the antigen presentation and activation/inhibition of T cells. The PD-1/PD-L1 axis is one of the co-inhibitory molecules that can be coexpressed in the context of antigen presentation. When the antigen molecule is presented by tumor cells together with co-inhibitory molecules, an inhibitory sign is triggered at the T cell. Then, the T cell activity is suppressed and the immune response fails to eradicate cancer. A similar mechanism occurs in autoimmunity. The insufficient expression of co-inhibitory molecules turns the immune response out of control. Then, tolerance to self-antigen is abrogated and the destructive immune response takes place in the healthy tissues

Figure 2 shows the common mechanisms related to both chronic lymphocytic thyroiditis and papillary thyroid carcinoma. Recent literature suggests that several mechanisms can cause both chronic lymphocytic thyroiditis and papillary thyroid carcinoma. Mechanisms can be classified into immune mechanisms and non-immune mechanisms. Among the immune mechanisms, we highlight some chemokines and interleukins that perform pro-inflammatory functions, in addition to playing a role in thyroid tumorigenesis. Among the non-immune mechanisms, we can highlight some viral infections, some non-coding RNAs and some genes commonly expressed by both conditions. It is noteworthy that other mechanisms may also be connecting these two diseases. It is possible, for example, that patients with chronic lymphocytic thyroiditis are subjected for long periods to a state of thyroid hormone insufficiency and TSH excess. Excess TSH, which is a naturally trophic hormone for thyroid cells, can facilitate cell proliferation and eventually lead to malignant cell transformation
References
Le Coulant P, Leuret JP, Texier L, Kermarec J, Maleville J, Aubertin J. A case of systemic amyloidosis with plasma cell infiltration and secondary epithelioma of the liver. Presse Med. 1893;68:820–822.
Korniluk A, Koper O, Kemona H, Dymicka-Piekarska V. From inflammation to cancer. Ir J Med Sci. 2017;186(1):57–62. https://doi.org/10.1007/s11845-016-1464-0.
Abbott M, Ustoyev Y. Cancer and the Immune System: The History and Background of Immunotherapy. Semin Oncol Nurs. 2019;35(5): 150923. https://doi.org/10.1016/j.soncn.2019.08.002.
Gonzalez H, Hagerling C, Werb Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018;32(19–20):1267–84. https://doi.org/10.1101/GAD.314617.118.
Loose D, Van De Wiele C. The immune system and cancer. Cancer Biother Radiopharm. 2009;24(3):369–76. https://doi.org/10.1089/cbr.2008.0593.
Hiam-Galvez KJ, Allen BM, Spitzer MH. Systemic immunity in cancer. Nat Rev Cancer. 2021;21(6):345–59. https://doi.org/10.1038/s41568-021-00347-z.
Sakowska J, et al. Autoimmunity and Cancer—Two Sides of the Same Coin. Front Immunol. 2022;13(May):1–22. https://doi.org/10.3389/fimmu.2022.793234.
De Sousa Linhares A, Leitner J, Grabmeier-Pfistershammer K, Steinberger P. Not All Immune Checkpoints Are Created Equal. Front Immunol. 2018;9:1–15, 2018. https://doi.org/10.3389/fimmu.2018.01909.
Ceccarelli F, Agmon-Levin N, Perricone C. Genetic Factors of Autoimmune Diseases 2017. J Immunol Res. 2017. Hindawi. https://doi.org/10.1155/2017/2789242.
Schmidt J et al. Neoantigen-specific CD8 T cells with high structural avidity preferentially reside in and eliminate tumors. Nat Commun. 2023;14(1). https://doi.org/10.1038/s41467-023-38946-z.
Hu X, et al. Cancer Risk in Hashimoto’s Thyroiditis: a Systematic Review and Meta-Analysis. Front Endocrinol (Lausanne). 2022;13(July):1–10. https://doi.org/10.3389/fendo.2022.937871.
Quaglino P, et al. Vitiligo is an independent favourable prognostic factor in stage III and IV metastatic melanoma patients: Results from a single-institution hospital-based observational cohort study. Ann Oncol. 2010;21(2):409–14. https://doi.org/10.1093/annonc/mdp325.
Boi F, Pani F, Mariotti S. Thyroid Autoimmunity and Thyroid Cancer: Review Focused on Cytological Studies. European Thyroid Journal. 2017;6(4):178–86. https://doi.org/10.1159/000468928.
Hua C, et al. Association of Vitiligo With Tumor Response in Patients With Metastatic Melanoma Treated With Pembrolizumab. JAMA dermatology. 2016;152(1):45–51. https://doi.org/10.1001/jamadermatol.2015.2707.
Hu X, Chen Y, Shen Y, Tian R, Sheng Y, Que H. Global prevalence and epidemiological trends of Hashimoto’s thyroiditis in adults: A systematic review and meta-analysis. Front Public Heal. 2022;10. https://doi.org/10.3389/fpubh.2022.1020709.
Ragusa F, et al. Hashimotos’ thyroiditis: Epidemiology, pathogenesis, clinic and therapy. Best Pract Res Clin Endocrinol Metab. 2019;33(6): 101367. https://doi.org/10.1016/j.beem.2019.101367.
Diab N et al. Prevalence and Risk Factors of Thyroid Dysfunction in Older Adults in the Community. Sci Rep. 2019;9(1):13156. https://doi.org/10.1038/s41598-019-49540-z.
Hollowell JG, et al. Serum TSH, T4, and thyroid antibodies in the United States population (1988 to 1994): National Health and Nutrition Examination Survey (NHANES III). J Clin Endocrinol Metab. 2002;87(2):489–99. https://doi.org/10.1210/jcem.87.2.8182.
Garmendia Madariaga A, Santos Palacios S, Guillén-Grima F, Galofré JC. The incidence and prevalence of thyroid dysfunction in Europe: A meta-analysis. J Clin Endocrinol Metab. 2014;99(3):923–931. https://doi.org/10.1210/jc.2013-2409.
Mendes D, Alves C, Silverio N, Marques FB. Prevalence of Undiagnosed Hypothyroidism in Europe: A Systematic Review and Meta-Analysis. Eur Thyroid J. 2019;8(3):130–43. https://doi.org/10.1159/000499751.
Ralli M, et al. Hashimoto’s thyroiditis: An update on pathogenic mechanisms, diagnostic protocols, therapeutic strategies, and potential malignant transformation. Autoimmun Rev. 2020;19(10): 102649. https://doi.org/10.1016/j.autrev.2020.102649.
Bery AI, Shepherd HM, Li W, Krupnick AS, Gelman AE, Kreisel D. Role of tertiary lymphoid organs in the regulation of immune responses in the periphery. Cell Mol Life Sci. 2022;79(7):1–18. https://doi.org/10.1007/s00018-022-04388-x.
Ferrari SM, et al. Chemokines in thyroid autoimmunity. Best Pract Res Clin Endocrinol Metab. 2023;37(2):101773. https://doi.org/10.1016/j.beem.2023.101773.
Álvarez-Sierra D, et al. Analysis of the PD-1/PD-L1 axis in human autoimmune thyroid disease: Insights into pathogenesis and clues to immunotherapy associated thyroid autoimmunity. J Autoimmun. 2019;103:102285. https://doi.org/10.1016/j.jaut.2019.05.013.
Zhang S, Wang L, Li M, Zhang F, Zeng X. The PD-1/PD-L pathway in rheumatic diseases. J Formos Med Assoc. 2021;120(1 Pt 1):48–59. https://doi.org/10.1016/j.jfma.2020.04.004.
Álvarez-Sierra D, et al. Lymphocytic Thyroiditis Transcriptomic Profiles Support the Role of Checkpoint Pathways and B Cells in Pathogenesis. Thyroid. 2022;32(6):682–93. https://doi.org/10.1089/thy.2021.0694.
Salazar-Viedma M, Vergaño-Salazar JG, Pastenes L, D’Afonseca V. Simulation Model for Hashimoto Autoimmune Thyroiditis Disease. Endocrinol (United States). 2021;162. https://doi.org/10.1210/endocr/bqab190.
McLachlan SM, Rapoport B. Thyrotropin-blocking autoantibodies and thyroid-stimulating autoantibodies: potential mechanisms involved in the pendulum swinging from hypothyroidism to hyperthyroidism or vice versa. Thyroid. 2013;23(1):14–24. https://doi.org/10.1089/thy.2012.0374.
Franco JS, Amaya-Amaya J, Anaya JM. Thyroid disease and autoimmune diseases. In: Anaya JM, Shoenfeld Y, Rojas-Villarraga A, et al., editors. Autoimmunity: From Bench to Bedside [Internet]. Bogota (Colombia): El Rosario University Press; 2013 Jul 18. Chapter 30. Available from: https://www.ncbi.nlm.nih.gov/books/NBK459466/.
Kaykhaein MA, et al. Association of CTLA4 (rs4553808) and PTPN22 (rs2476601) gene polymorphisms with Hashimoto’s thyroiditis disease: A case-control study and an In-silico analysis. Meta Gene. 2020;24:100693. https://doi.org/10.1016/j.mgene.2020.100693.
Aversa T, et al. Peculiarities of autoimmune thyroid diseases in children with Turner or Down syndrome: An overview. Ital J Pediatr. 2015;41(1):1–5. https://doi.org/10.1186/s13052-015-0146-2.
Martínez-Hernández R, Marazuela M. MicroRNAs in autoimmune thyroid diseases and their role as biomarkers. Best Pract Res Clin Endocrinol Metab. 2023;37(2): 101741. https://doi.org/10.1016/j.beem.2023.101741.
Bernecker C, et al. MicroRNAs miR-146a1, miR-155-2, and miR-200a1 are regulated in autoimmune thyroid diseases. Thyroid. 2012;22(12):1294–5. https://doi.org/10.1089/thy.2012.0277.
Klicka K, Grzywa TM, Mielniczuk A, Klinke A, Włodarski PK. The role of miR-200 family in the regulation of hallmarks of cancer. Front Oncol. 2022;12: 965231. https://doi.org/10.3389/fonc.2022.965231.
Thai T-H, et al. Regulation of the germinal center response by microRNA-155. Science. 2007;316(5824):604–8. https://doi.org/10.1126/science.1141229.
Tanaka PP, et al. miR-155 exerts posttranscriptional control of autoimmune regulator (Aire) and tissue-restricted antigen genes in medullary thymic epithelial cells. BMC Genom. 2022;23(1):404. https://doi.org/10.1186/s12864-022-08631-4.
Martínez-Hernández R, et al. Integrated miRNA and mRNA expression profiling identifies novel targets and pathological mechanisms in autoimmune thyroid diseases. EBioMedicine. 2019;50:329–42. https://doi.org/10.1016/j.ebiom.2019.10.061.
Wortzel I, Dror S, Kenific CM, Lyden D. Exosome-Mediated Metastasis: Communication from a Distance. Dev Cell. 2019;49(3):347–60. https://doi.org/10.1016/j.devcel.2019.04.011.
Mirzaei R, et al. The pathogenic, therapeutic and diagnostic role of exosomal microRNA in the autoimmune diseases. J Neuroimmunol. 2021;358: 577640. https://doi.org/10.1016/j.jneuroim.2021.577640.
Chaudhari P, Ghate V, Nampoothiri M, Lewis S. Multifunctional role of exosomes in viral diseases: From transmission to diagnosis and therapy. Cell Signal. 2022;94: 110325. https://doi.org/10.1016/j.cellsig.2022.110325.
Dai J, et al. Exosomes: key players in cancer and potential therapeutic strategy. Signal Transduct Target Ther. 2020;5(1):145. https://doi.org/10.1038/s41392-020-00261-0.
Rodríguez-Muñoz A, et al. Circulating microvesicles regulate treg and Th17 differentiation in human autoimmune thyroid disorders. J Clin Endocrinol Metab. 2015;100(12):E1531–9. https://doi.org/10.1210/jc.2015-3146.
Rodríguez-Muñoz A, et al. Circulating Microvesicles Regulate Treg and Th17 Differentiation in Human Autoimmune Thyroid Disorders. J Clin Endocrinol Metab. 2015;100(12):E1531-9. https://doi.org/10.1210/jc.2015-3146.
Boi F, Lai ML, Marziani B, Minerba L, Faa G, Mariotti S. High prevalence of suspicious cytology in thyroid nodules associated with positive thyroid autoantibodies. Eur J Endocrinol. 2005;153(5):637–42. https://doi.org/10.1530/eje.1.02020.
Lisboa Souza S, Vera Montalli da Assumpçêo L, Sterian Ward L. Impact of previous thyroid autoimmune diseases on prognosis of patients with well-differentiated thyroid cancer. Thyroid. 2003;13(5)491–495. https://doi.org/10.1089/105072503322021160.
Mussa A, Matarazzo P, Corrias A. Papillary thyroid cancer and autoimmune polyglandular syndrome. J Pediatr Endocrinol Metab. 2015;28(7–8):793–5. https://doi.org/10.1515/jpem-2014-0268.
Keefe G, et al. Autoimmune Thyroiditis and Risk of Malignancy in Children with Thyroid Nodules. Thyroid. 2022;32(9):1109–17. https://doi.org/10.1089/thy.2022.0241.
Januś D, et al. Ultrasound, laboratory and histopathological insights in diagnosing papillary thyroid carcinoma in a paediatric population: a single centre follow-up study between 2000–2022. Front Endocrinol (Lausanne). 2023;14(May):1–17. https://doi.org/10.3389/fendo.2023.1170971.
Januś D, et al. Ultrasound evolution of parenchymal changes in the thyroid gland with autoimmune thyroiditis in children prior to the development of papillary thyroid carcinoma – a follow-up study. Front Endocrinol (Lausanne). 2023;14(April):1–13. https://doi.org/10.3389/fendo.2023.1172823.
Dailey ME, Lindsay S, Skahen R. RELATION OF THYROID NEOPLASMS TO HASHIMOTO DISEASE OF THE THYROID GLAND. AMA Arch Surg. 1955;70(2):291–7. https://doi.org/10.1001/archsurg.1955.01270080137023.
Holm L-E, Blomgren H, Löwhagen T. Cancer Risks in Patients with Chronic Lymphocytic Thyroiditis. N Engl J Med. 1985;312(10):601–4. https://doi.org/10.1056/NEJM198503073121001.
Oh CM, et al. Increased prevalence of chronic lymphocytic thyroiditis in Korean patients with papillary thyroid cancer. PLoS One. 2014;9(6). https://doi.org/10.1371/journal.pone.0099054.
Lee JH, Kim Y, Choi JW, Kim YS. The association between papillary thyroid carcinoma and histologically proven Hashimoto’s thyroiditis: A meta-analysis. Eur J Endocrinol. 2013;168(3):343–9. https://doi.org/10.1530/EJE-12-0903.
Cunha LL, Ward LS. Concurrent lymphocytic thyroiditis is associated to less aggressive papillary thyroid carcinomas. Eur Arch Oto-Rhino-Laryngology. 2012;269(2):699–700. https://doi.org/10.1007/s00405-011-1764-y.
Marotta V, et al. Hashimoto’s thyroiditis predicts outcome in intrathyroidal papillary thyroid cancer. Endocr Relat Cancer. 2017;24(9):485–93. https://doi.org/10.1530/ERC-17-0085.
Xu S, et al. Prevalence of Hashimoto Thyroiditis in Adults With Papillary Thyroid Cancer and Its Association With Cancer Recurrence and Outcomes. JAMA Netw Open. 2021;4(7):e2118526. https://doi.org/10.1001/jamanetworkopen.2021.18526.
Babli S, Payne RJ, Mitmaker E, Rivera J. Effects of chronic lymphocytic thyroiditis on the clinicopathological features of papillary thyroid cancer. Eur Thyroid J. 2018;7(2):95–101. https://doi.org/10.1159/000486367.
Lee I, Kim HK, Soh EY, Lee J. The Association Between Chronic Lymphocytic Thyroiditis and the Progress of Papillary Thyroid Cancer. World J Surg. 2020;44(5):1506–13. https://doi.org/10.1007/s00268-019-05337-9.
Osborne D, et al. Hashimoto’s Thyroiditis Effects on Papillary Thyroid Carcinoma Outcomes: A Systematic Review. Cureus. 2022;14(8). https://doi.org/10.7759/cureus.28054.
Hanege FM, Tuysuz O, Celik S, Sakallıoglu O, Solmaz OA. Hashimoto’s thyroiditis in papillary thyroid carcinoma: A 22-year study. Acta Otorhinolaryngol Ital. 2021;41(2):142–5. https://doi.org/10.14639/0392-100X-N1081.
Min Y, et al. Preoperatively Predicting the Central Lymph Node Metastasis for Papillary Thyroid Cancer Patients With Hashimoto’s Thyroiditis. Front Endocrinol (Lausanne). 2021;12(July):1–11. https://doi.org/10.3389/fendo.2021.713475.
Dong S, Xia Q, Wu YJ. High TPOAb levels (> 1300 IU/mL) indicate multifocal PTC in Hashimoto’s thyroiditis patients and support total thyroidectomy. Otolaryngol Head Neck Surg. 2015;153(1):20-6. https://doi.org/10.1177/0194599815581831.
Ward LS, Scheffel RS, Hoff AO, Ferraz C, Vaisman F. Treatment strategies for low-risk papillary thyroid carcinoma: a position statement from the Thyroid Department of the Brazilian Society of Endocrinology and Metabolism (SBEM). Arch Endocrinol Metabolism. 2022;66(4):522–532, 2022. Brazil. https://doi.org/10.20945/2359-3997000000512.
Ernaga-Lorea A, et al. Prognostic value of change in anti-thyroglobulin antibodies after thyroidectomy in patients with papillary thyroid carcinoma. Clin Transl Oncol Off Publ Fed Spanish Oncol Soc Natl Cancer Inst Mex. 2018;20(6):740–744. https://doi.org/10.1007/s12094-017-1782-3.
Chiovato L, et al. Disappearance of humoral thyroid autoimmunity after complete removal of thyroid antigens. Ann Intern Med. 2003;139(5 Pt 1):346–51. https://doi.org/10.7326/0003-4819-139-5_part_1-200309020-00010.
Kim WG, et al. Change of serum antithyroglobulin antibody levels is useful for prediction of clinical recurrence in thyroglobulin-negative patients with differentiated thyroid carcinoma. J Clin Endocrinol Metab. 2008;93(12):4683–9. https://doi.org/10.1210/jc.2008-0962.
Spencer C, Fatemi S. Thyroglobulin antibody (TgAb) methods - Strengths, pitfalls and clinical utility for monitoring TgAb-positive patients with differentiated thyroid cancer. Best Pract Res Clin Endocrinol Metab. 2013;27(5):701–12. https://doi.org/10.1016/j.beem.2013.07.003.
Tsushima Y, et al. Prognostic significance of changes in serum thyroglobulin antibody levels of pre- and post-total thyroidectomy in thyroglobulin antibody-positive papillary thyroid carcinoma patients. Endocr J. 2013;60(7):871–6. https://doi.org/10.1507/endocrj.ej12-0410.
Yamada O, et al. Changes in serum thyroglobulin antibody levels as a dynamic prognostic factor for early-phase recurrence of thyroglobulin antibody-positive papillary thyroid carcinoma after total thyroidectomy. Endocr J. 2014;61(10):961–5. https://doi.org/10.1507/endocrj.ej14-0275.
Hsieh C-J, Wang P-W. Sequential changes of serum antithyroglobulin antibody levels are a good predictor of disease activity in thyroglobulin-negative patients with papillary thyroid carcinoma. Thyroid. 2014;24(3):488–93. https://doi.org/10.1089/thy.2012.0611.
Görges R, et al. Development and clinical impact of thyroglobulin antibodies in patients with differentiated thyroid carcinoma during the first 3 years after thyroidectomy. Eur J Endocrinol. 2005;153(1):49–55. https://doi.org/10.1530/eje.1.01940.
Zavala LF, et al. In properly selected patients with differentiated thyroid cancer, antithyroglobulin antibodies decline after thyroidectomy and their sole presence should not be an indication for radioiodine ablation. Arch Endocrinol Metab. 2019;63(3):293–9. https://doi.org/10.20945/2359-3997000000123.
de Martel C, Georges D, Bray F, Ferlay J, Clifford GM. Global burden of cancer attributable to infections in 2018: a worldwide incidence analysis. Lancet Glob Heal. 2020;8(2):e180–90. https://doi.org/10.1016/S2214-109X(19)30488-7.
Blackard JT, Kong L, Huber AK, Tomer Y. Hepatitis C virus infection of a thyroid cell line: Implications for pathogenesis of hepatitis C virus and thyroiditis. Thyroid. 2013;23(7):863–70. https://doi.org/10.1089/thy.2012.0507.
Akeno N, Blackard JT, Tomer Y. HCV E2 protein binds directly to thyroid cells and induces IL-8 production: A new mechanism for HCV induced thyroid autoimmunity. J Autoimmun. 2008;31(4):339–44. https://doi.org/10.1016/j.jaut.2008.08.001.
Liotti F, et al. Interleukin-8, but not the Related Chemokine CXCL1, Sustains an Autocrine Circuit Necessary for the Properties and Functions of Thyroid Cancer Stem Cells. Stem Cells. 2017;35(1):135–46. https://doi.org/10.1002/stem.2492.
Leite JL, Bufalo NE, Santos RB, Romaldini JH, Ward LS. Herpesvirus type 7 infection may play an important role in individuals with a genetic profile of susceptibility to Graves’ disease. Eur J Endocrinol. 2010;162(2):315–21. https://doi.org/10.1530/EJE-09-0719.
Almeida JFM, et al. Investigation on the association between thyroid tumorigeneses and herpesviruses. J Endocrinol Invest. 2017;40(8):823–9. https://doi.org/10.1007/s40618-017-0609-y.
Assaad SN, Meheissen MA, Elsayed ET, Alnakhal SN, Salem TM. Study of Epstein–Barr virus serological profile in Egyptian patients with Hashimoto’s thyroiditis: A case-control study. J Clin Transl Endocrinol. 2020;20:100222. https://doi.org/10.1016/j.jcte.2020.100222.
Almeida JFM, et al. Epstein–Barr virus induces morphological and molecular changes in thyroid neoplastic cells. Endocrine. 2020;69(2):321–30. https://doi.org/10.1007/s12020-020-02253-0.
Cunha LL, et al. Infiltration of a mixture of immune cells may be related to good prognosis in patients with differentiated thyroid carcinoma. Clin Endocrinol (Oxf). 2012;77(6):918–25. https://doi.org/10.1111/j.1365-2265.2012.04482.x.
Sulaieva O, Selezniov O, Shapochka D, Belemets N. Heliyon Hashimoto ’ s thyroiditis attenuates progression of papillary thyroid carcinoma : deciphering immunological links. Heliyon. 2020;6:e03077. https://doi.org/10.1016/j.heliyon.2019.e03077.
Cunha LL, et al. CD8+ tumour-infiltrating lymphocytes and COX2 expression may predict relapse in differentiated thyroid cancer. Clin Endocrinol (Oxf). 2015;83(2):246–53. https://doi.org/10.1111/cen.12586.
Vitales-Noyola M, et al. Pathogenic Th17 and Th22 cells are increased in patients with autoimmune thyroid disorders. Endocrine. 2017;57(3):409–17. https://doi.org/10.1007/s12020-017-1361-y.
Guan LJ, et al. Increased IL-21/IL-21R expression and its proinflammatory effects in autoimmune thyroid disease. Cytokine. 2015;72(2):160–165. https://doi.org/10.1016/j.cyto.2014.11.005.
Zake T, Skuja S, Kalere I, Konrade I, Groma V. Heterogeneity of tissue IL-17 and tight junction proteins expression demonstrated in patients with autoimmune thyroid diseases. Med (United States). 2018;97(25):1–6. https://doi.org/10.1097/MD.0000000000011211.
Cunha LL, et al. RORγt may Influence the Microenvironment of Thyroid Cancer Predicting Favorable Prognosis. Sci Rep. 2020;10(1):1–12. https://doi.org/10.1038/s41598-020-60280-3.
Zeng R, et al. Positive effect of RORγt on the prognosis of thyroid papillary carcinoma patients combined with hashimoto’s thyroiditis. Am J Transl Res. 2018;10(10):3011–24.
Heidari Z, et al. Association of IL-1 β, NLRP3, and COX-2 Gene Polymorphisms with Autoimmune Thyroid Disease Risk and Clinical Features in the Iranian Population. Biomed Res Int. 2021;2021. https://doi.org/10.1155/2021/7729238.
Kaneko N, Kurata M, Yamamoto T, Morikawa S, Masumoto J. The role of interleukin-1 in general pathology. Inflamm Regen. 2019;39:12. https://doi.org/10.1186/s41232-019-0101-5.
Chan AH, Schroder K. Inflammasome signaling and regulation of interleukin-1 family cytokines. J Exp Med. 2020;217(1). https://doi.org/10.1084/jem.20190314.
Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduct Target Ther. 2021;6(1). https://doi.org/10.1038/s41392-021-00507-5.
Babamale AO, Chen ST. Nod-like receptors: Critical intracellular sensors for host protection and cell death in microbial and parasitic infections. Int J Mol Sci. 2021;22(21). https://doi.org/10.3390/ijms222111398.
Guo Q, et al. Cytokine Secretion and Pyroptosis of Thyroid Follicular Cells Mediated by Enhanced NLRP3, NLRP1, NLRC4, and AIM2 Inflammasomes Are Associated With Autoimmune Thyroiditis. Front Immunol. 2018;9:1197. https://doi.org/10.3389/fimmu.2018.01197.
Wu P, Shi J, Sun W, Zhang H. Identification and validation of a pyroptosis-related prognostic signature for thyroid cancer. Cancer Cell Int. 2021;21(1):1–16. https://doi.org/10.1186/s12935-021-02231-0.
Sheils OM, O’Leary JJ, Uhlmann V, Lüttich K, Sweeney EC. ret/PTC-1 activation in Hashimoto thyroiditis. Int J Surg Pathol. 2000;8(3):185–9. https://doi.org/10.1177/106689690000800305.
Mechler C, et al. Papillary thyroid carcinoma: 6 Cases from 2 families with associated lymphocytic thyroiditis harbouring RET/PTC rearrangements. Br J Cancer. 2001;85(12):1831–7. https://doi.org/10.1054/bjoc.2001.2187.
Zhang L, Zhou L, Feng Q, Li Q, Ge M. Mutation of Hashimoto’s Thyroiditis and Papillary Thyroid Carcinoma Related Genes and the Screening of Candidate Genes. Front Oncol. 2021;11(December):1–11. https://doi.org/10.3389/fonc.2021.813802.
Liu C, Pan Y, Li Q, Zhang Y. Bioinformatics analysis identified shared differentially expressed genes as potential biomarkers for hashimoto’s thyroiditis-related papillary thyroid cancer. Int J Med Sci. 2021;18(15):3478–87. https://doi.org/10.7150/ijms.63402.
Dong H, et al. Lactoferrin-containing immunocomplex mediates antitumor effects by resetting tumor-associated macrophages to M1 phenotype. J Immunother Cancer. 2020;8(1). https://doi.org/10.1136/jitc-2019-000339.
Afeltra A, Paggi A, De Rosa FG, Manfredini P, Addessi MA, Amoroso A. Antineutrophil cytoplasmic antibodies in autoimmune thyroid disorders. Endocr Res. 1998;24(2):185–94. https://doi.org/10.1080/07435809809135527.
Chen J, Cao H, Lian M, Fang J. Five genes influenced by obesity may contribute to the development of thyroid cancer through the regulation of insulin levels. PeerJ. 2020;8:1–12. https://doi.org/10.7717/peerj.9302.
Sancho M, et al. Expression and function of the chemokine receptor CCR7 in thyroid carcinomas. J Endocrinol. 2006;191(1):229–38. https://doi.org/10.1677/joe.1.06688.
Sulaieva O, et al. Hashimoto’s thyroiditis attenuates progression of papillary thyroid carcinoma: deciphering immunological links. Heliyon. 2020;6(1):1–21. https://doi.org/10.1016/j.heliyon.2019.e03077.
Statello L, Guo CJ, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118. https://doi.org/10.1038/s41580-020-00315-9.
Gao N, et al. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front Oncol. 2020;10(December):1–13. https://doi.org/10.3389/fonc.2020.598817.
O’Brien J, Hayder H. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol (Lausanne). 2018;9:1–12. https://doi.org/10.3389/fendo.2018.00402.
Liu Y, et al. Competitive endogenous RNA is an intrinsic component of EMT regulatory circuits and modulates EMT. Nat Commun. 2019;10(1):1637. https://doi.org/10.1038/s41467-019-09649-1.
Zhang Y, Tian Y. Comprehensive analysis of lncRNA-mediated ceRNA regulatory networks and key genes associated with papillary thyroid cancer coexistent with Hashimoto’s thyroiditis. BMC Endocr Disord. 2022;22(1):1–11. https://doi.org/10.1186/s12902-022-01173-6.
Dasgupta S, Chakrabarti S, Mandal PK, Das S. Hashimoto’s Thyroiditis and Medullary Carcinoma of Thyroid. JNMA J Nepal Med Assoc. 2014;52(194):831–3.
Weiss LM, Weinberg DS, Warhol MJ. Medullary carcinoma arising in a thyroid with Hashimoto’s disease. Am J Clin Pathol. 1983;80(4):534–8. https://doi.org/10.1093/ajcp/80.4.534.
De Pasquale L, Sommaruga L, Andreani S, Bastagli A. Hashimoto’s thyroiditis and medullary carcinoma in the same gland: a purely random occurrence? Chir Ital. 2004;56(4):557–62.
Mousa U, Gursoy A, Ozdemir H, Moray G. Medullary thyroid carcinoma in a patient with Hashimoto’s thyroiditis diagnosed by calcitonin washout from a thyroid nodule. Diagn Cytopathol. 2013;41(7):644–6. https://doi.org/10.1002/dc.21850.
SamieeRad F, Emami A. Synchronous Occurrence of Papillary Thyroid Carcinoma and Medullary Carcinoma in the Setting of Hashimoto’s Thyroiditis and Multi Nodular Goiter. Iran J Pathol. 2022;27(1):91–6. https://doi.org/10.30699/ijp.2021.527288.2606.
Abdullah AM, Ali RM, Salih KM, Mohammed KK, Kakamad FH, Salih AM. Synchronous occurrence of papillary thyroid microcarcinoma, medullary thyroid carcinoma and Hashimoto thyroiditis in a single thyroid: A case report with literature review. Int J Surg Case Rep. 2022;93:106888. https://doi.org/10.1016/j.ijscr.2022.106888.
Zayed AA, et al. Is Hashimoto’s thyroiditis a risk factor for medullary thyroid carcinoma? Our experience and a literature review. Endocrine. 2015;48(2):629–36. https://doi.org/10.1007/s12020-014-0363-2.
Ma R, Morshed SA, Latif R, Davies TF. A Stem Cell Surge During Thyroid Regeneration. Front Endocrinol (Lausanne). 2021;11(January):1–11. https://doi.org/10.3389/fendo.2020.606269.
Davies TF, Latif R, Minsky NC, Ma R. Clinical review: The emerging cell biology of thyroid stem cells. J Clin Endocrinol Metab. 2011;96(9):2692–702. https://doi.org/10.1210/jc.2011-1047.
Zhu W, Hai T, Ye L, Cote GJ. Medullary thyroid carcinoma cell lines contain a self-renewing CD133+ population that is dependent on ret proto-oncogene activity. J Clin Endocrinol Metab. 2010;95(1):439–44. https://doi.org/10.1210/jc.2009-1485.
Singh B, Shaha AR, Trivedi H, Carew JF, Poluri A, Shah JP. Coexistent Hashimoto’s thyroiditis with papillary thyroid carcinoma: impact on presentation, management, and outcome. Surgery. 1999;126(6):1070–7. https://doi.org/10.1067/msy.2099.101431.
Huang B-Y, Hseuh C, Chao T-C, Lin K-J, Lin J-D. Well-differentiated thyroid carcinoma with concomitant Hashimoto’s thyroiditis present with less aggressive clinical stage and low recurrence. Endocr Pathol. 2011;22(3):144–9. https://doi.org/10.1007/s12022-011-9164-9.
Kashima K, et al. Chronic thyroiditis as a favorable prognostic factor in papillary thyroid carcinoma. Thyroid. 1998;8(3):197–202. https://doi.org/10.1089/thy.1998.8.197.
Kim EY, et al. Coexistence of chronic lymphocytic thyroiditis is associated with lower recurrence rates in patients with papillary thyroid carcinoma. Clin Endocrinol (Oxf). 2009;71(4):581–6. https://doi.org/10.1111/j.1365-2265.2009.03537.x.
Song E, et al. Influence of coexistent Hashimoto’s thyroiditis on the extent of cervical lymph node dissection and prognosis in papillary thyroid carcinoma. Clin Endocrinol (Oxf). 2018;88(1):123–8. https://doi.org/10.1111/cen.13475.
Kwak HY, et al. Does papillary thyroid carcinoma have a better prognosis with or without Hashimoto thyroiditis? Int J Clin Oncol. 2015;20(3):463–73. https://doi.org/10.1007/s10147-014-0754-7.
Yoon Y-H, Kim HJ, Lee JW, Kim JM, Koo BS. The clinicopathologic differences in papillary thyroid carcinoma with or without co-existing chronic lymphocytic thyroiditis. Eur Arch oto-rhino-laryngology Off J Eur Fed Oto-Rhino-Laryngological Soc Affil with Ger Soc Oto-Rhino-Laryngology - Head Neck Surg. 2012;269(3):1013–7.
Jeong JS, et al. Coexistence of chronic lymphocytic thyroiditis with papillary thyroid carcinoma: clinical manifestation and prognostic outcome. J Korean Med Sci. 2012;27(8):883–9. https://doi.org/10.3346/jkms.2012.27.8.883.
Girardi FM, Barra MB, Zettler CG. Papillary thyroid carcinoma: does the association with Hashimoto’s thyroiditis affect the clinicopathological characteristics of the disease? Braz J Otorhinolaryngol. 2015;81(3):283–7. https://doi.org/10.1016/j.bjorl.2014.04.006.
Ryu YJ, Yoon JH. Chronic lymphocytic thyroiditis protects against recurrence in patients with cN0 papillary thyroid cancer. Surg Oncol. 2020;34:67–73. https://doi.org/10.1016/j.suronc.2020.03.008.
Molnár C, et al. Thyroid Carcinoma Coexisting with Hashimoto’s Thyreoiditis: Clinicopathological and Molecular Characteristics Clue up Pathogenesis. Pathol Oncol Res. 2019;25(3):1191–7. https://doi.org/10.1007/s12253-019-00580-w.
Konturek A, Barczyński M, Wierzchowski W, Stopa M, Nowak W. Coexistence of papillary thyroid cancer with Hashimoto thyroiditis. Langenbeck’s Arch Surg. 2013;398(3):389–94. https://doi.org/10.1007/s00423-012-1021-x.
Liang J, et al. Clinical analysis of Hashimoto thyroiditis coexistent with papillary thyroid cancer in 1392 patients. Acta Otorhinolaryngol Ital organo Uff della Soc Ital di Otorinolaringol e Chir Cerv-facc. 2017;37(5):393–400. https://doi.org/10.14639/0392-100X-1709.
Zhu F, Bin Shen Y, Li FQ, Fang Y, Wu YJ. The Effects of Hashimoto Thyroiditis on Lymph Node Metastases in Unifocal and Multifocal Papillary Thyroid Carcinoma: A Retrospective Chinese Cohort Study. Medicine (Baltimore). 2016;95(6):e2674. https://doi.org/10.1097/MD.0000000000002674.
Jara SM, et al. The relationship between chronic lymphocytic thyroiditis and central neck lymph node metastasis in North American patients with papillary thyroid carcinoma. Surgery. 2013;154(6):1272. https://doi.org/10.1016/j.surg.2013.07.021.
Zhu Y. The clinicopathologic differences of central lymph node metastasis in predicting lateral lymph node metastasis and prognosis in papillary thyroid cancer associated with or without Hashimoto’s thyroiditis. Tumour Biol J Int Soc Oncodevelopmental Biol Med. 2016;37(6):8037–45. https://doi.org/10.1007/s13277-015-4706-2.
Kim KW, et al. Elevated risk of papillary thyroid cancer in Korean patients with Hashimoto’s thyroiditis. Head Neck. 2011;33(5):691–5. https://doi.org/10.1002/hed.21518.
Cappellacci F, et al. Association between hashimoto thyroiditis and differentiated thyroid cancer: A single-center experience. Front Oncol. 2022;12: 959595. https://doi.org/10.3389/fonc.2022.959595.
Cordioli MICV, Cury AN, Nascimento AO, de Oliveira AK, Mello M, Saieg MA. Study of the histological profile of papillary thyroid carcinomas associated with Hashimoto’s thyroiditis. Arq Bras Endocrinol Metabol. 2013;57(6):445–9. https://doi.org/10.1590/s0004-27302013000600006.
Lun Y, et al. Hashimoto’s thyroiditis as a risk factor of papillary thyroid cancer may improve cancer prognosis. Otolaryngol Neck Surg Off J Am Acad Otolaryngol Neck Surg. 2013;148(3):396–402. https://doi.org/10.1177/0194599812472426.
Kurukahvecioglu O, Taneri F, Yüksel O, Aydin A, Tezel E, Onuk E. Total thyroidectomy for the treatment of Hashimoto’s thyroiditis coexisting with papillary thyroid carcinoma. Adv Ther. 2007;24(3):510–6. https://doi.org/10.1007/BF02848773.
Kim HS, Choi YJ, Yun J-S. Features of papillary thyroid microcarcinoma in the presence and absence of lymphocytic thyroiditis. Endocr Pathol. 2010;21(3):149–53. https://doi.org/10.1007/s12022-010-9124-9.
Kebebew E, Treseler PA, Ituarte PHG, Clark OH. Coexisting Chronic Lymphocytic Thyroiditis and Papillary Thyroid Cancer Revisited. World J Surg. 2001;25(5):632–7. https://doi.org/10.1007/s002680020165.
Ahn D, et al. Clinical relationship between Hashimoto’s thyroiditis and papillary thyroid cancer. Acta Oncol. 2011;50(8):1228–34. https://doi.org/10.3109/0284186X.2011.602109.
Dobrinja C, et al. Coexistence of chronic lymphocytic thyroiditis and papillary thyroid carcinoma. Impact on presentation, management, and outcome. Int J Surg. 2016;28(Suppl 1):S70-4. https://doi.org/10.1016/j.ijsu.2015.12.059.
Consorti F, Loponte M, Milazzo F, Potasso L, Antonaci A. Risk of malignancy from thyroid nodular disease as an element of clinical management of patients with Hashimoto’s thyroiditis. Eur Surg Res Eur Chir Forschung Rech Chir Eur. 2010;45(3–4):333–7. https://doi.org/10.1159/000320954.
Asanuma K, Sugenoya A, Kasuga Y, Itoh N, Kobayashi S, Amano J. The relationship between multiple intrathyroidal involvement in papillary thyroid carcinoma and chronic non-specific thyroiditis. Cancer Lett. 1998;122(1–2):177–80. https://doi.org/10.1016/s0304-3835(97)00398-4.
Schäffler A, et al. Coexistent thyroiditis is associated with lower tumour stage in thyroid carcinoma. Eur J Clin Invest. 1998;28(10):838–44. https://doi.org/10.1046/j.1365-2362.1998.00363.x.
Acknowledgement
The authors thank FAPESP and CNPq for grants and funding for research.
Funding
This work did not receive any specific grant from any funding agency in the public, commercial, or not-for-profit sector. Fundação de Amparo à Pesquisa do Estado de São Paulo, 2021/02752-6, 2021/02752-6, 2021/02752-6, 2021/02752-6, Thyroid Department, Sociedade Brasileira de Endocrinologia e Metabologia.
Author information
Authors and Affiliations
Contributions
VASV, FRB, LSW, and LLC wrote the main manuscript text, participated in conception and design, critical review of the literature and data, composition of the manuscript, and final approval. VASV prepared the figures. VASV and FRB prepared the table. LSW and LLC promoted clinical and translational orientation. VASV, FRB, and LLC prepared the selection of notable articles for review, and critical review of the literature and data. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
dos Santos Valsecchi, V.A., Betoni, F.R., Ward, L.S. et al. Clinical and molecular impact of concurrent thyroid autoimmune disease and thyroid cancer: From the bench to bedside. Rev Endocr Metab Disord 25, 5–17 (2024). https://doi.org/10.1007/s11154-023-09846-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11154-023-09846-w