
Abstract
This review presents an overview of the two ways that cyanobacteria, algae, and plants have adapted to high O2 and low CO2 concentrations in the environment. First, the process of photorespiration enables photosynthetic organisms to recycle phosphoglycolate formed by the oxygenase reaction catalyzed by ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco). Second, there are a number of carbon concentrating mechanisms that increase the CO2 concentration around Rubisco which increases the carboxylase reaction enhancing CO2 fixation. This review also presents possibilities for the beneficial modification of these processes with the goal of improving future crop yields.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
The interplay between photosynthetic organisms and atmospheric change
The process of photosynthesis has had a profound effect on the Earth’s atmosphere. It is estimated that the primordial atmosphere was high in CO2 and extremely low in O2 (Fig. 1) (Sage 2004; Govindjee and Shevela 2011; Kasting 1987). As photosynthesis developed, the atmospheric CO2 concentration slowly decreased as plants fixed atmospheric CO2 and carbon was subsequently sequestered in the ground or ocean floor as carboniferous fossils (Fig. 1). Besides the decline in CO2 concentration, a critical change occurred with the evolution of Photosystem II which produces O2. As a consequence of photosynthetic organisms, the partial pressure of CO2 has dropped from an estimated 1,000 mbar in the atmosphere of the primitive earth to 0.4 mbar and the concentration of O2 has risen from almost undetectable levels to approximately 210 mbar in the present day atmosphere (Fig. 1) (Govindjee and Shevela 2011; Sage 2004).

The changes in the O2 and CO2 concentrations in the atmosphere during Earth’s history. Redrawn from Sage (2004)
While oxygenic photosynthetic organisms changed the makeup of the atmosphere, they also had to adapt to these same changes. In the Calvin–Benson–Bassham cycle (CBB cycle), the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) catalyzes the addition of CO2 to ribulose-1,5-bisphosphate (RuBP). Over the Earth’s history, the concentration of the substrate of this reaction, atmospheric CO2, has dropped dramatically. This has led to a decrease in the efficiency of this carboxylation reaction. In addition, the product of the reaction catalyzed by Photosystem II, O2, inhibits the process of CO2 fixation. All aerobic photosynthetic organisms must deal with this present day combination of high O2 and low CO2 conditions.
O2 inhibits photosynthesis and plant growth
The first report that O2 inhibits photosynthesis was made by Warburg in 1920 (Warburg 1920). Later it was shown that lowering the O2 concentration to 2 % resulted in a 20–30 percent increase in biomass and yield in plants (Hesketh 1967; Chollet and Ogren 1975). Conversely, if the O2 concentration was increased to above atmospheric levels, photosynthesis was severely reduced (Turner and Brittain 1962).
Why does O2 inhibit CO2 fixation? The key to answering this question was the molecule glycolate. When Calvin and his colleagues fed Chlorella 14CO2, 14C-labeled glycolate was one of the early products observed (Benson et al. 1950; Schou et al. 1950). However, the amount of labeled glycolate was variable. Sometimes a lot of labeled glycolate was found, whereas in other experiments, very little glycolate was observed. In his review article, Tolbert (1973) noted that glycolate was found when (1) the CO2 concentration was low, (2) the O2 concentration was high, (3) the pH was high, or (4) the light intensity was high. It was also observed that glycolate was evenly labeled on both carbons, glycolate was never labeled before 3-phosphoglycerate. Zelitch and Ochoa (1953) showed the enzyme glycolate oxidase catalyzed the conversion of glycolate to glyoxylate in plant tissues. These observations led to the hypothesis that glycolate was not on the main pathway of carbon fixation, but instead was the result of an O2-dependent side reaction. Feeding algal cells labeled glycolate allowed researchers to detail the pathway of glycolate metabolism, tracing a pathway from glycolate to 3-phosphoglycerate (Fig. 2) (Husic et al. 1987).

Outline of the photorespiratory pathway in higher plants emphasizing major carbon intermediates and compartments. The NH3 released in the mitochondrion is recycled (not shown). The dashed line indicates transport. CBB Cycle Calvin–Benson–Bassham cycle
However, it was not clear how glycolate was formed. The breakthrough came in 1971, when Bowes et al. (1971) reported that Rubisco also catalyzes an oxygenation reaction with RuBP, leading to one molecule of phosphoglycerate and one of phosphoglycolate (1b). Therefore, Rubisco catalyzes these two separate reactions:
It was also clear that chloroplasts had phosphoglycolate phosphatase activity. This enzyme converts phosphoglycolate formed during the oxygenation reaction to glycolate (Fig. 2).
This finding led to the current explanation of how O2 inhibits photosynthesis. Rubisco is able to catalyze an oxygenation reaction as well as a carboxylation reaction with RuBP. It is apparent that CO2 and O2 are competitive substrates. The relative amounts of carboxylation and oxygenation are dependent on the properties of Rubisco and the [CO2]/[O2] ratio at which they take place (Andrews et al. 1973; Lorimer et al. 1973; Spreitzer and Salvucci 2002). The Rubisco specificity factor S C/O describes the enzyme’s catalytic properties for carboxylation and oxygenation and is defined as S C/O = V C K O/V O K C, where V C and V O are the maximal velocities for carboxylation and oxygenation, respectively, and K O and K C are the Michaelis–Menten constants of Rubisco for O 2 and CO 2 , respectively (Jordan and Ogren 1981b). In a typical analysis, the average S C/O for ten C3 plants was about 86 and in ten C4 plants about 79 (see table S1 in Whitney et al. (2011). S C/O indicates the relative specificity for reacting with CO2 versus O2 and can be obtained by determining the slope of the plot of V C/V O versus [CO2]/[O2] when carboxylase and oxygenase activities are measured simultaneously (Jordan and Ogren 1981a). Thus, the relative catalysis of the carboxylase versus oxygenase in vivo depends on S C/O times the ratio of [CO2]/[O2] present (e.g., about 10 μM CO2 vs 280 μM O2 around the enzyme during photosynthesis in C3 species). Obviously, in the primitive atmosphere of high [CO2]/[O2] (Fig. 1), only the carboxylation reaction would occur. In contrast, the Earth’s atmosphere is presently about 20.95 % O2 and 0.039 % CO2, and the ratio of the carboxylation reaction to the oxygenation reaction is about 4 to 1 in most C3 plants (Sharkey 1988).
Photorespiration is a way to recapture carbon potentially lost due to the oxygenation reaction of Rubisco
Photorespiration is defined as the light-dependent consumption of O2 and production of CO2. Clearly photorespiration is counterproductive if a plant gives off CO2, since the purpose of photosynthesis is to capture CO2 and produce sugars. The photorespiratory pathway serves to LIMIT the loss of fixed carbon due to the oxygenase activity of Rubisco. When phosphoglycolate is generated by the oxygenase reaction, there is a potential of losing both of these carbons. In fact, the photorespiratory pathway returns 75 % of the potentially “lost” carbon back to 3-phosphoglycerate and the CBB cycle (Fig. 2) since two phosphoglycolate molecules contain four carbons and are photorespired to 3-phosphoglycerate which contains three carbons.
Several steps characterize photorespiration. The biochemistry of this pathway is very well understood for many different prokaryotic and eukaryotic species and reviewed extensively (Kern et al. 2011; Bauwe et al. 2010; Husic et al. 1987). To summarize the process with a focus on higher plants, the 2-phosphoglycolate that is produced by Rubisco is first dephosphorylated by phosphoglycolate phosphatase to glycolate. Glycolate is transported out of the chloroplast into peroxisomes and oxidized by glycolate oxidase to glyoxylate which is further modified by glutamate–glyoxylate aminotransferase to glycine. Two glycine molecules can be transformed into one serine molecule by a glycine decarboxylase complex in mitochondria (Fig. 2). The one carbon “lost” is the CO2 given off at this step when the two glycines are converted to one serine, one CO2 and one molecule of ammonia. Back in the peroxisome, serine is modified to hydroxypyruvate by serine-glyoxylate aminotransferase and further to glycerate by hydroxypyruvate reductase. Glycerate is transported into the chloroplast where it is phosphorylated by glycerate kinase to 3-phosphoglycerate, the key-molecule that interlinks photorespiration and the CBB cycle (Fig. 2) (Husic et al. 1987).
Is the photorespiratory pathway essential to plants?
The answer is clearly yes. First, if any of the key photorespiratory enzymes are blocked or disrupted by mutation, higher plants either die or grow very slowly under normal atmospheric concentrations of O2 and CO2 (Somerville and Ogren 1982; Blackwell et al. 1988; Timm et al. 2012; Zelitch et al. 2009). Only by keeping the plants in an artificially high CO2 environment can plants defective in the photorespiratory pathway be maintained (Timm et al. 2012). It is clear then, that plants must efficiently recycle 2-phosphoglycolate back to 3-phosphoglycerate to survive under today’s atmospheric conditions. Another potential consequence of the photorespiratory pathway is the loss of ammonia (NH3) when two molecules of glycine are converted to one molecule of serine (see Fig. 2). This ammonia must be reassimilated by the plant, adding to the energetic cost of photorespiration, but this is necessary to insure survival.
Why does Rubisco still have an oxygenase activity?
One question often raised when photorespiration and the CCM are discussed is “Why has not Rubisco evolved to either better fix CO2 or avoid the oxygenase reaction altogether?” While we cannot unequivocally answer this question, at least two reasonable hypotheses have been put forth to explain the persistent oxygenase activity of Rubisco. The first hypothesis is that the mechanism used by Rubisco to catalyze the carboxylation reaction cannot easily be changed through mutation to eliminate the oxygenase reaction. It is important to point out that there are no known CO2 or O2 substrate binding sites on Rubisco. Instead, it is thought that Rubisco stabilizes the formation of the enediol conformation of RuBP, allowing either CO2 or O2 to react with RuBP (Eqs. 1a, 1b) (Cleland et al. 1998; Spreitzer 1993; Spreitzer and Salvucci 2002). The way Rubisco “favors” the carboxylation reaction over the oxygenation reaction is by stabilizing the six carbon intermediate before it is cleaved to form two molecules of 3-phosphoglycerate. Since there are no binding sites for the two substrates, CO2 and O2, amino acid substitution will not favor either of the substrates so the competing reactions are dependent on the concentrations of O2 and CO2. One could imagine a mutation or series of mutations that might prevent the diffusion of O2 to the RuBP binding site. However, since CO2 and O2 are both small uncharged molecules, it is possible that this type of change is extremely unlikely. In spite of these difficulties, there is a wide variety of specificities for CO2 versus O2 for Rubisco from different species. For example, the Rubisco specificity factors (S C/O) of higher plants are in the range of 80–90 while they are in the range of 40–50 in cyanobacteria. The increased specificity seen in higher plants appears to be a result of tighter binding of the six-carbon intermediate by Rubisco. The higher plant enzyme thus favors the carboxylation reaction, but it is also much slower than the cyanobacterial Rubisco (Jordan and Ogren 1981b) although all Rubiscos appear optimized to the natural conditions under which they operate (Tcherkez et al. 2006). The very low catalytic rate of Rubisco especially in higher plants is in part, due to how tightly Rubisco binds the six carbon catalytic intermediate. At this point, the scientific community has not found or engineered a form of Rubisco that has completely eliminated the oxygenase activity.
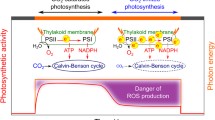
The second hypothesis to explain why the oxygenase activity is persistent in nature is that photosynthetic organisms need an extra outlet for the high energy molecules, namely ATP and NADPH produced by the light-dependent reactions of photosynthesis (Kozaki and Takeba 1996). Considering that the CO2 concentration is so low in nature, there may be times during the day when the production of ATP and NADPH far exceeds the needs of the CBB cycle. This might be particularly true under high light conditions or drought conditions when the stomata are closed, preventing CO2 from entering the leaf. In addition, soil and fresh-water organisms often encounter very low CO2 concentrations as noted earlier. Under these conditions, the electron transport chain could easily become over-reduced, producing superoxide or other reactive oxygen species causing photodamage (Kozaki and Takeba 1996).
It is possible that both of these hypotheses concerning the oxygenation reaction of Rubisco in photosynthesis are valid. Another strategy to reduce the oxygenation reaction of Rubisco implies the active concentrating of CO2 around Rubisco. Clearly there is strong selective pressure in favor of carbon concentrating mechanisms in aquatic environments and C4 photosynthesis has arisen independently in many plant families. However, at the same time, the oxygenation activity of Rubisco occurs to some extent in all species.
Carbon concentrating mechanisms, a way to increase CO2 for fixation
Introduction to carbon concentrating mechanisms (CCMs)
Because O2 and CO2 are competing substrates, two ways to enhance photosynthesis are (1) to reduce the O2 concentration or (2) to increase the CO2 concentration at the site of Rubisco. The reduction of O2 is especially difficult as oxygenic photosynthetic organisms are generating O2 through the action of Photosystem II very close to Rubisco. A common way that photosynthetic organisms reduce the oxygenase reaction is to increase CO2 at the site of Rubisco. By increasing CO2 for Rubisco, the carboxylase reaction is increased and the oxygenase reaction is reduced. Photosynthetic organisms have evolved a number of ways to increase CO2 for Rubisco, including cyanobacterial CCMs, similar CCMs in eukaryotic algae and C4 photosynthesis found in higher plants. In all cases, the CCM increases the CO2 concentration for Rubisco, enhancing CO2 fixation and minimizing the oxygenase reaction.
As noted, CCMs increase the concentration of CO2 at the site of Rubisco. In cells lacking CCMs the CO2 concentration in equilibrium with the atmosphere is very low (around 10 μM). Although there may be some membrane selectivity for CO2 transport, overall the permeability for CO2 is relatively high (Tholen and Zhu 2011). Therefore concentrating CO2 around Rubisco is potentially difficult. So what is the mechanism for concentrating CO2? This is accomplished by accumulating the carbon in a form other than CO2. Examples include the charged molecule bicarbonate (HCO3 −) or metabolites like malate or aspartate which do not cross biological membranes as rapidly as CO2. These molecules are transported to the location where Rubisco is present and CO2 is generated there. In cyanobacteria and eukaryotic algae, the accumulated molecule is HCO3 − which is converted to CO2 in the carboxysome or chloroplast pyrenoid respectively through the activity of carbonic anhydrase (CA). In the case of C4 plants, the accumulated malate (or aspartate) is decarboxylated in the bundle sheath cells where Rubisco is located. The CO2 generated from these accumulated molecules is largely fixed by Rubisco due to diffusive resistance limiting its leakage from the cell.
A model for the CO2 concentrating mechanism in cyanobacteria
A general model for concentrating CO2 in cyanobacteria is shown in Fig. 3 (Price et al. 2008). In this model, bicarbonate accumulates within the cytoplasm of the cell and Rubisco is packaged inside the proteinaceous structure called the carboxysome. The accumulated bicarbonate then enters the carboxysome through pores (Klein et al. 2009) in the carboxysome shell (Tsai et al. 2007; Tanaka et al. 2008) and is converted to CO2 by a carbonic anhydrase. This creates a microenvironment with a high CO2 concentration for Rubisco inside the carboxysome (Yeates et al. 2008). Compelling evidence for this model comes from mutant analysis where disruptions of the carboxysome (Ohkawa et al. 1998; Schwarz et al. 1995), loss of the carboxysomal carbonic anhydrase (Price et al. 1992; Price and Badger 1989a, b, 2002; Yu et al. 1992), or targeting Rubisco to the cytoplasm instead of the carboxysome (Orús et al. 1995; Pierce et al. 1989) results in a cell unable to grow on low concentrations of CO2 (0.04 % CO2).

The CCM of cyanobacteria. Left is an electron micrograph of Synechococcus PCC 7942 from (Long et al. 2007), published by permission. The model of inorganic carbon transport in a cyanobacterial cell is shown on the right. HCO3 − is taken up by transporters (unfilled circle) and accumulated in the cytoplasm. A carbonic anhydrase converts HCO3 − to CO2 in the carboxysome (see arrows in micrograph). CO2 is subsequently fixed into 3-phosphoglycerate (3PGA) by Rubisco. Based on the model of Price et al. (2008)
In cyanobacteria, the accumulation of HCO3 − is accomplished by multiple HCO3 − uptake systems (Omata et al. 1999; Shibata et al. 2002; Price et al. 2004). These transporters include a Na+- HCO3 − cotransporter (Ohkawa et al. 1998; Espie and Kandasamy 1994; Ogawa et al. 1998), a low CO2-inducible ABC-type transporter (Omata et al. 1999, 2002) and the BicA HCO3 − transporter (Price et al. 2004). In addition to these HCO3 – uptake systems, CO2 that enters the cell is converted to HCO3 – internally by two proteins associated with the thylakoid membranes (Fig. 3) (Maeda S-i et al. 2002; Price et al. 2002). Studies comparing the genomes of different bacteria reveal that all of these uptake systems may not be present in a given species, but many of the cyanobacteria examined so far have redundant Ci uptake systems.
In cyanobacteria, Rubisco is localized to the carboxysome. Three types of CAs are associated with the carboxysome although usually only one type is found in a given organism (Pena et al. 2010; So et al. 2002b; Yu et al. 1992; Cannon et al. 2010). Carbonic anhydrases are proteins that accelerate the inter-conversion between inorganic carbon species (2).
As an example, one type of carbonic anhydrase is encoded by the CcaA gene. Cyanobacteria with ccaA mutations have normal carboxysomes but require high CO2 for growth (So et al. 2002a; Yu et al. 1992; Fukuzawa et al. 1992). Interestingly, these mutants accumulate HCO3 – to very high levels, indicating that the HCO3 – uptake systems are still operating, but the cell can no longer convert the accumulated HCO3 – to CO2 for fixation. All cyanobacteria studied to date have a carboxysomal CA, and this CA is required for the functioning of the CCM.
A model for the concentration of CO2 in Chlamydomonas, an example of a eukaryotic algal CCM
The CCM in green algae has many similarities to the cyanobacterial model including active HCO3 − accumulation, Rubisco packaging and HCO3 − dehydration to CO2 in the chloroplast (Fig. 4) (Moroney et al. 2011). In a current model of the CCM in Chlamydomonas, a green unicellular alga, bicarbonate is accumulated within the chloroplast stroma and Rubisco is localized to the pyrenoid (Rawat et al. 1996), an electron-dense structure within the chloroplast (Fig. 4). The accumulated bicarbonate is then converted to CO2 by a carbonic anhydrase, CAH3, located within the thylakoid lumen. Evidence for this model comes from mutant analysis where loss of two or more CO2 or HCO3 − transporters (Pollock et al. 2004; Duanmu et al. 2009; Moroney et al. 1989; Spalding et al. 1983b; Wang and Spalding 2006), the CAH3 carbonic anhydrase (Karlsson et al. 1998; Spalding et al. 1983a; Funke et al. 1997), or disruption of the pyrenoid (Ma et al. 2011; Genkov et al. 2010), leads to Chlamydomonas strains that are unable to grow on low concentrations of CO2. In addition, it is important to note that the CCM in Chlamydomonas is inducible (Badger et al. 1980; Somanchi et al. 1998; Moroney and Tolbert 1985). If Chlamydomonas is grown on high CO2 (>1 % CO2 in air), it does not produce the CCM components. However, if Chlamydomonas is switched from high CO2 (>1 % CO2) to low (0.04 % CO2) or very low CO2 (0.01 % CO2), it will acclimate within about 4 h and the affinity of the alga for CO2 and HCO3 − increases by 10–20 fold.

The CCM of the eukaryotic green alga Chlamydomonas reinhardtii. Left is an electron micrograph of C. reinhardtii with the current CCM model is presented on the right. CO2 diffuses into to the cells and is converted by cytoplasmic carbonic anhydrases to HCO3 −. HCO3 − is also taken up into the cytoplasm by specific transporters (unfilled circle). Both inorganic carbon species enter the chloroplast, HCO3 − via active transport, CO2 via diffusion. A carbonic anhydrase is also present in the chloroplast stroma and converts CO2 to HCO3 −, which is transported into the thylakoid lumen. The thylakoids span through the Rubisco containing pyrenoid (P), here the bicarbonate is converted to CO2 which is fixed into 3PGA. A starch shell surrounds the pyrenoid and minimizes CO2 diffusion out of the fixation site
C4 photosynthesis
C4 photosynthesis, a process seen in many higher plants, is another way to increase the CO2 concentration for Rubisco. Most C4 plants have a type of leaf structure known as Kranz anatomy, with two distinct types of photosynthetic cells, the mesophyll cells and the bundle sheath cells (Fig. 5). C4 photosynthesis has arisen independently many times (Sage et al. 2011) predominantly by forming the two-cell type Kranz anatomy (Edwards et al. 2001; Hatch 2002), with the spatial separation of function as illustrated in Fig. 5. The process is called C4 photosynthesis because the first product with fixed CO2 is a four carbon organic acid. In the cytoplasm of mesophyll cells, carbonic anhydrase converts CO2 to HCO3 −, which is added to phosphoenolpyruvate (PEP) by PEP carboxylase to form oxaloacetate which is converted to malate or aspartate (Fig. 5). One reason C4 photosynthesis can concentrate CO2 is because PEP carboxylase has a high affinity for HCO3 −, allowing high rates of fixation even at low CO2 + HCO3 − concentrations and unlike Rubisco, oxygen is not a competing substrate. The C4 acids malate and aspartate are transported from mesophyll to bundle sheath cells. C4 decarboxylases (NADP-malic enzyme, NAD-malic enzyme, or PEP carboxykinase, depending on the C4 biochemical subtype) generate CO2 and a three carbon product, the latter returning to the mesophyll cells for regeneration of PEP. Through the C4 cycle, the CO2 is concentrated around Rubisco which limits photorespiration. The energy required for CO2 fixation in the CCB cycle via Rubisco is 3 ATP and 2 NADPH. In C3 plants, photorespiration results in additional use of energy which increases the cost per CO2 fixed, while in C4 plants there is the added cost of 2 ATP per CO2 delivered to Rubisco by the C4 cycle. C4 photosynthesis is especially advantageous under conditions where photorespiration increases, i.e., when CO2 is limited under water stress due to reduced stomatal conductance, and under higher leaf temperatures where the kinetic properties of Rubisco change favoring an increase in reaction with O2. For further details on evolution and function of C4 see (Christin and Osborne 2013; Ludwig 2013). While it is clear that the mechanisms for C4 photosynthesis and the algal CCMs are very different, they both accomplish the same thing, namely raising the CO2 concentration for Rubisco, thereby favoring the carboxylation reaction at the expense of the oxygenase reaction.

The C4 photosynthetic pathway. a Light micrograph of a leaf cross-section from the C-4 plant Zea mays. GC guard cell, IGS intercellular gas space, MC mesophyll cell, BC bundle sheath cell. b Overview of the C4 photosynthetic pathway. Dashed lines represent plasmodesmata transport. CBB Cycle Calvin–Benson–Bassham cycle, PEP phosphoenolpyruvate, OAA oxaloacetate, C3 pyruvate, C4 malate/aspartate
How widespread are these adaptations in photosynthetic organisms?
Photosynthetic organisms have adapted to the earth’s changing atmosphere including the rise in the concentration of O2 and the decline in CO2 concentration. The process of photorespiration appears to be an ancient adaptation. Recent studies by Bauwe and his colleagues indicate that photorespiration was likely present when the chloroplast endosymbiosis event occurred (Bauwe et al. 2010, 2012; Eisenhut et al. 2008). Not only is the pathway present in cyanobacteria as well as algae and higher plants, the enzymes that make up the pathway are remarkably similar. In higher plants, the chloroplastic enzymes phosphoglycolate phosphatase and glycerate kinase closely align with their cyanobacterial counterparts (Bauwe et al. 2012; Kern et al. 2011).
The origin of the other adaptation, the CCM, appears to be more recent than the appearance of photorespiration but the CCM is also commonly found in nature. Almost all eukaryotic algae and cyanobacteria thus far examined contain a CCM. On the other hand, about four percent of higher plant species conduct C4 photosynthesis. The widespread occurrence of the CCM in aquatic organisms is due to the fact that aquatic photosynthetic organisms commonly face an environment where the dissolved CO2 concentration is 10 μM or less. In addition, the diffusion coefficient of CO2 in water is approximately 10,000 times smaller than the coefficient for CO2 in air also limiting the availability of CO2 to aquatic photosynthetic organisms. In contrast, higher plants live in a variety of environments where water stress may not be a factor and stomates allow ready access of gaseous CO2 to photosynthetic cells.
If a photosynthetic organism has a CCM, is photorespiration still needed?
The answer to this question is yes. To this point in time, the photorespiratory pathway has been found in all C4 plants (Sage et al. 2012; Dai et al. 1993), eukaryotic algae (Moroney et al. 1986) and cyanobacteria studied (Eisenhut et al. 2008; Hagemann et al. 2010) as well as C3 plant species. The requirement for photorespiration in C4 plants was eloquently demonstrated in a maize glycolate oxidase mutant (Zelitch et al. 2009). This question is particularly interesting in the case of cyanobacteria which have the ability to concentrate bicarbonate over a 100-fold. However, even this very high concentration does not completely eliminate the oxygenase reaction. Cyanobacteria with a normal CCM but which cannot process phosphoglycolate grow poorly on low concentrations of CO2 (Eisenhut et al. 2008). To date, all oxygenic photosynthetic organisms have an active photorespiratory pathway.
Future efforts to improve photosynthesis by modifying the photorespiratory pathway and introducing a CCM
It is clear that both the photorespiratory pathway and the various carbon concentrating mechanisms have evolved as a response to the rise in O2 concentration and decline in CO2 concentration in the Earth’s atmosphere. However, C3 plants, including most of the important crop plants, do not employ a CCM. Plant scientists are now exploring the question whether photosynthesis and/or crop yield might be improved by (1) changing Rubisco, (2) altering the photorespiratory pathway, or (3) by introducing a CCM into a C3 plant.
Improving Rubisco
Clearly the cause of the O2 inhibition of CO2 fixation is the oxygenase activity of Rubisco (Eqs. 1a, 1b). If the oxygenase activity could be reduced or eliminated, there might be a significant increase in plant productivity under today’s atmospheric concentrations of CO2 and O2. It has been argued that through evolution, Rubiscos found in nature are optimized to balance catalytic efficiency and the [CO2]/[O2] specificity and therefore the oxygenase activity of Rubisco cannot be eliminated (Whitney et al. 2011; Whitney and Sharwood 2008; Bainbridge et al. 1995). However, there is clear evidence that when comparing Rubiscos from a wide variety of organisms there is a significant variation in the [CO2]/[O2] specificity of the enzyme (Jordan and Ogren 1981b; Whitney et al. 2011). Perhaps a Rubisco from an organism with a high [CO2]/[O2] specificity could be incorporated into a crop plant that normally has a lower [CO2]/[O2] specificity. Possibly as we better understand which amino acids influence the [CO2]/[O2] specificity of Rubisco, an improved Rubisco could be designed and put into crop plants to increase photosynthesis and yield. A caveat to this type of approach is whether the oxygenase is needed to reduce photodamage, as the oxygenase activity does allow for an alternate electron sink. Modifying Rubisco to a point where there is minimal oxygenase activity might lead to a plant that is more susceptible to photodamage.
Improving the photorespiratory pathway
If the oxygenase activity of Rubisco cannot be avoided, perhaps the recycling of the carbon in glycolate could be improved. Presently there are no known metabolic pathways that can improve upon the 75 % carbon recovery rate of the photorespiratory pathway. However, there have been attempts to improve on the recapture of the 25 % of the carbon “lost” as CO2. In a recent experiment, three bacterial genes were introduced into tobacco to create the tartronic semialdehyde pathway in the tobacco chloroplast (Fig. 6) (Kebeish et al. 2007; Peterhansel and Maurino 2011). The advantage of this pathway over the existing pathway is that (1) there are fewer steps (3 vs 5) and (2) the CO2 is released in the chloroplast stroma instead of the mitochondria (compare Figs. 2, 6), and (3) no additional nitrogen needs to be reassimilated. The idea is that CO2 released in the chloroplast has a better chance to be refixed than CO2 released in the mitochondria. The fact that nitrogen does not have to be reassimilated means the plant uses less energy with this pathway. Plants containing this pathway have been generated and are now being tested under different growth conditions (Peterhansel and Maurino 2011). This strategy may lead to better growth of the modified plants.

Introducing a CCM into a C3 plant
Scientists have begun to consider the possibility of what might happen if a CCM was introduced into a C3 crop plant. A major international project was begun a few years ago to create a C4 rice plant. Since C4 photosynthesis has arisen independently many times in nature, it is reasonable to think that C4-like traits could be selected for, particularly in important C3 crop species (Furbank et al. 2009; von Caemmerer and Evans 2010). The type of traits that are being selected for include increased expression of specific C4 enzymes or the development of anatomical characteristics associated with Kranz anatomy. Clearly with the regulation of so many genes being different in C3 and C4 plants, the transition from C3 to C4 photosynthesis would normally take many generations. However, the process might be accelerated by introducing C4 genes along with promoters that insure the appropriate expression into rice cultivars to facilitate the conversion of photosynthesis to a more C4-like process. von Caemmerer and colleagues (2012) recently published a progress report on an effort to develop C4 rice.
Another way to introduce a CCM into a C3 plant is to incorporate components of the algal CCMs into higher plants. This approach has been championed by Price and Badger and their colleagues (Price et al. 2008, 2011). The method entails putting either cyanobacterial or algal HCO3 − transporters into C3 plants. In theory, these transporters would help deliver CO2 to Rubisco enhancing photosynthesis. Initially the proteins would be targeted to the chloroplast envelope although it is possible that transporters on the plasma membrane would help as well (Price et al. 2011; Weber and von Caemmerer 2010). There is a report in the literature that the introduction of the ictb gene (thought to code for a protein involved in HCO3 − transport in cyanobacteria) into Arabidopsis and tobacco enhanced both photosynthesis at low CO2 and improved water balance (Lieman-Hurwitz et al. 2003). This report underscores a second possible benefit of incorporating CCM genes into higher plants which is improved water usage. When plants acquire CO2 through open stomates, a great deal of water vapor is lost. In fact on a mass basis, a typical transpiration ratio is 400 water molecules lost for every CO2 molecule fixed (Nobel 2009). If a plant could operate more efficiently with less CO2, it also might need less water.
The human population continues to grow while crop yields using conventional plant are not keeping pace (Zhu et al. 2010). A significant challenge to plant scientists will be to improve crop yield to feed the Earth’s growing population. One potential way to improve crop yield is to use our knowledge of photosynthetic processes to develop ways to improve photosynthesis and potentially increase crop productivity. Scientists are now poised to increase plant production and yield by engineering photorespiration and enhancing CO2 delivery to Rubisco. The authors would like to thank Drs. Govinjee and Joliot for their contributions to our understanding of photosynthesis. Their work has helped make it possible to design the approaches described in this closing section.
Abbreviations
- CA:
-
Carbonic anhydrase
- CBB cycle:
-
Calvin–Benson–Bassham cycle
- CCM:
-
Carbon (dioxide) concentrating mechanism
- PEP:
-
Phosphoenolpyruvate
- Rubisco:
-
Ribulose-1,5-bisphosphate carboxylase/oxygenase
- RuBP:
-
Ribulose-1,5-bisphosphate
References
Andrews TJ, Lorimer GH, Tolbert NE (1973) Ribulose diphosphate oxygenase. 1. Synthesis of phosphoglycolate by fraction-1 protein of leaves. Biochem-Us 12(1):11–18. doi:10.1021/Bi00725a003
Badger M, Kaplan A, Berry J (1980) Internal inorganic carbon pool of Chlamydomonas reinhardtii: evidence for a carbon dioxide-concentrating mechanism. Plant Physiol 66(3):407
Bainbridge G, Madgwick P, Parmar S, Mitchell R, Paul M, Pitts J, Keys AJ, Parry MAJ (1995) Engineering Rubisco to change its catalytic properties. J Exp Bot 46:1269–1276
Bauwe H, Hagemann M, Fernie AR (2010) Photorespiration: players, partners and origin. Trends Plant Sci 15(6):330–336. doi:10.1016/j.tplants.2010.03.006
Bauwe H, Hagemann M, Kern R, Timm S (2012) Photorespiration has a dual origin and manifold links to central metabolism. Curr Opin Plant Biol 15(3):269–275. doi:10.1016/j.pbi.2012.01.008
Benson AA, Bassham JA, Calvin M, Goodale TC, Haas VA, Stepka W (1950) The path of carbon in photosynthesis. 5. Paper chromatography and radioautography of the products. J Am Chem Soc 72(4):1710–1718. doi:10.1021/Ja01160a080
Blackwell RD, Murray AJS, Lea PJ, Kendall AC, Hall NP, Turner JC, Wallsgrove RM (1988) The value of mutants unable to carry out photorespiration. Photosynth Res 16(1–2):155–176. doi:10.1007/Bf00039491
Bowes G, Ogren WL, Hageman RH (1971) Phosphoglycolate production catalyzed by ribulose diphosphate carboxylase. Biochem Biophys Res Commun 45(3):716–722. doi:10.1016/0006-291X(71)90475-X
Cannon GC, Heinhorst S, Kerfeld CA (2010) Carboxysomal carbonic anhydrases: Structure and role in microbial CO2 fixation. Biochim Biophys Acta (BBA)—Proteins & Proteomics 1804(2):382–392. doi:10.1016/j.bbapap.2009.09.026
Chollet R, Ogren WL (1975) Regulation of photorespiration in C3 and C4 species. Bot Rev 41(2):137–179. doi:10.1007/Bf02860828
Christin P-A, Osborne C (2013) The recurrent assembly of C4 photosynthesis, an evolutionary tale. Photosynthesis research (in press)
Cleland WW, Andrews TJ, Gutteridge S, Hartman FC, Lorimer GH (1998) Mechanism of Rubisco: the carbamate as general base. Chem Rev 98(2):549–561. doi:10.1021/Cr970010r
Dai ZY, Ku MSB, Edwards GE (1993) C4 photosynthesis—the CO2-concentrating mechanism and photorespiration. Plant Physiol 103(1):83–90
Duanmu D, Miller AR, Horken KM, Weeks DP, Spalding MH (2009) Knockdown of limiting-CO2-induced gene HLA3 decreases HCO3 − transport and photosynthetic Ci affinity in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 106(14):5990–5995. doi:10.1073/pnas.0812885106
Edwards GE, Furbank RT, Hatch MD, Osmond CB (2001) What does it take to be C-4? Lessons from the evolution of C-4 photosynthesis. Plant Physiol 125(1):46–49. doi:10.1104/Pp.125.1.46
Eisenhut M, Ruth W, Haimovich M, Bauwe H, Kaplan A, Hagemann M (2008) The photorespiratory glycolate metabolism is essential for cyanobacteria and might have been conveyed endosymbiontically to plants. Proc Natl Acad Sci USA 105(44):17199–17204. doi:10.1073/pnas.0807043105
Espie G, Kandasamy R (1994) Monensin inhibition of Na+-dependent HCO3 − transport distinguishes it from Na+-independent HCO3 −transport and provides evidence for Na+/HCO3 −symport in the cyanobacterium Synechococcus UTEX 625. Plant Physiol 104(4):1419–1428
Fukuzawa H, Suzuki E, Komukai Y, Miyachi S (1992) A gene homologous to chloroplast carbonic anhydrase (icfA) is essential to photosynthetic carbon dioxide fixation by Synechococcus PCC7942. Proc Natl Acad Sci USA 89(10):4437–4441
Funke RP, Kovar JL, Weeks DP (1997) Intracellular carbonic anhydrase is essential to photosynthesis in Chlamydomonas reinhardtii at atmospheric levels of CO2. Demonstration via genomic complementation of the high-CO2-requiring mutant ca-1. Plant physiol 114(1):237–244
Furbank RT, von Caemmerer S, Sheehy J, Edwards G (2009) C-4 rice: a challenge for plant phenomics. Funct Plant Biol 36(10–11):845–856. doi:10.1071/Fp09185
Genkov T, Meyer M, Griffiths H, Spreitzer RJ (2010) Functional hybrid Rubisco enzymes with plant small subunits and algal large subunits. J Biol Chem 285(26):19833–19841. doi:10.1074/jbc.M110.124230
Govindjee, Shevela D (2011) Adventures with cyanobacteria: a personal perspective. Front Plant Sci 2:28. doi:10.3389/fpls.2011.00028
Hagemann M, Eisenhut M, Hackenberg C, Bauwe H (2010) Pathway and importance of photorespiratory 2-phosphoglycolate metabolism in cyanobacteria. Adv Exp Med Biol 675:91–108. doi:10.1007/978-1-4419-1528-3_6
Hatch MD (2002) C-4 photosynthesis: discovery and resolution. Photosynth Res 73(1–3):251–256. doi:10.1023/A:1020471718805
Hesketh J (1967) Enhancement of photosynthetic CO2 assimilation in absence of oxygen as dependent upon species and temperature. Planta 76(4):371–374. doi:10.1007/Bf00387543
Husic DW, Husic HD, Tolbert NE (1987) The oxidative photosynthetic carbon-cycle or C2 cycle. Crit Rev Plant Sci 5(1):45–100. doi:10.1080/07352688709382234
Jordan DB, Ogren WL (1981a) A sensitive assay procedure for simultaneous determination of ribulose-1,5-bisphosphate carboxylase and oxygenase activities. Plant Physiol 67(2):237–245. doi:10.1104/Pp.67.2.237
Jordan DB, Ogren WL (1981b) Species variation in the specificity of ribulose-biphosphate carboxylase-oxygenase. Nature 291(5815):513–515. doi:10.1038/291513a0
Karlsson J, Clarke AK, Chen Z-Y, Hugghins SY, Park Y-I, Husic HD, Moroney JV, Samuelsson G (1998) A novel [alpha]-type carbonic anhydrase associated with the thylakoid membrane in Chlamydomonas reinhardtii is required for growth at ambient CO2. EMBO J 17(5):1208–1216
Kasting JF (1987) Theoretical constraints on oxygen and carbon-dioxide concentrations in the precambrian atmosphere. Precambrian Res 34(3–4):205–229. doi:10.1016/0301-9268(87)90001-5
Kebeish R, Niessen M, Thiruveedhi K, Bari R, Hirsch HJ, Rosenkranz R, Stabler N, Schonfeld B, Kreuzaler F, Peterhansel C (2007) Chloroplastic photorespiratory bypass increases photosynthesis and biomass production in Arabidopsis thaliana. Nat Biotechnol 25(5):593–599. doi:10.1038/Nbt1299
Kern R, Bauwe H, Hagemann M (2011) Evolution of enzymes involved in the photorespiratory 2-phosphoglycolate cycle from cyanobacteria via algae toward plants. Photosynth Res 109(1–3):103–114. doi:10.1007/s11120-010-9615-z
Klein MG, Zwart P, Bagby SC, Cai F, Chisholm SW, Heinhorst S, Cannon GC, Kerfeld CA (2009) Identification and structural analysis of a novel carboxysome shell protein with implications for metabolite transport. J Mol Biol 392(2):319–333. doi:10.1016/j.jmb.2009.03.056
Kozaki A, Takeba G (1996) Photorespiration protects C3 plants from photooxidation. Nature 384(6609):557–560. doi:10.1038/384557a0
Lieman-Hurwitz J, Rachmilevitch S, Mittler R, Marcus Y, Kaplan A (2003) Enhanced photosynthesis and growth of transgenic plants that express ictB, a gene involved in HCO3 − accumulation in cyanobacteria. Plant Biotechnol J 1(1):43–50. doi:10.1046/j.1467-7652.2003.00003.x
Long BM, Badger MR, Whitney SM, Price GD (2007) Analysis of carboxysomes from Synechococcus PCC7942 reveals multiple Rubisco complexes with carboxysomal proteins CcmM and CcaA. J Biol Chem 282(40):29323–29335. doi:10.1074/jbc.M703896200
Lorimer GH, Andrews TJ, Tolbert NE (1973) Ribulose Diphosphate Oxygenase. 2. Further Proof of Reaction-Products and Mechanism of Action. Biochem-Us 12(1):18–23. doi:10.1021/Bi00725a004
Ludwig M (2013) Evolution of the C4 photosynthetic pathway: events at the cell and molecular levels. Photosynthesis research (in press)
Ma Y, Pollock SV, Xiao Y, Cunnusamy K, Moroney JV (2011) Identification of a novel gene, CIA6, required for normal pyrenoid formation in Chlamydomonas reinhardtii. Plant Physiol 156(2):884–896. doi:10.1104/pp.111.173922
Moroney JV, Tolbert NE (1985) Inorganic carbon uptake by Chlamydomonas reinhardtii. Plant Physiol 77(2):253–258
Moroney JV, Wilson BJ, Tolbert NE (1986) Glycolate metabolism and excretion by Chlamydomonas Reinhardtii. Plant Physiol 82(3):821–826. doi:10.1104/Pp.82.3.821
Moroney JV, Husic HD, Tolbert NE, Kitayama M, Manuel LJ, Togasaki RK (1989) Isolation and characterization of a mutant of Chlamydomonas reinhardtii deficient in the CO2 concentrating mechanism. Plant Physiol 89(3):897–903
Moroney JV, Ma Y, Frey WD, Fusilier KA, Pham TT, Simms TA, DiMario RJ, Yang J, Mukherjee B (2011) The carbonic anhydrase isoforms of Chlamydomonas reinhardtii: intracellular location, expression, and physiological roles. Photosynth Res 109(1–3):133–149. doi:10.1007/s11120-011-9635-3
Nobel P (2009) Physicochemical and environmental plant physiology, 4th edn. Elsevier, Oxford
Ogawa T, Katoh A, Sonoda M (1998) Molecular mechanisms of CO2 concentration and proton extrusion in cyanobacteria. Paper presented at the Stress Responses of Photosynthetic Organisms, Amsterdam
Ohkawa H, Sonoda M, Katoh H, Ogawa T (1998) The use of mutants in the analysis of the CO2-concentrating mechanism in cyanobacteria. Botany 76(6):1035–1042
Omata T, Price GD, Badger MR, Okamura M, Gohta S, Ogawa T (1999) Identification of an ATP-binding cassette transporter involved in bicarbonate uptake in the cyanobacterium Synechococcus sp. strain PCC 7942. Proc Natl Acad Sci USA 96(23):13571–13576
Omata T, Yamaguchi O, Takahashi Y, Nishimura T (2002) Structure, function and regulation of the cyanobacterial high-affinity bicarbonate transporter, BCT1. Funct Plant Biol 29(3):151–159
Orús M, Rodriguez M, Martinez F, Marco E (1995) Biogenesis and ultrastructure of carboxysomes from wild type and mutants of Synechococcus sp. strain PCC 7942. Plant Physiol 107(4):1159–1166
Pena KL, Castel SE, de Araujo C, Espie GS, Kimber MS (2010) Structural basis of the oxidative activation of the carboxysomal gamma-carbonic anhydrase, CcmM. Proc Natl Acad Sci USA 107(6):2455–2460. doi:10.1073/pnas.0910866107
Peterhansel C, Maurino VG (2011) Photorespiration redesigned. Plant Physiol 155(1):49–55. doi:10.1104/pp.110.165019
Pierce J, Carlson TJ, Williams JG (1989) A cyanobacterial mutant requiring the expression of ribulose bisphosphate carboxylase from a photosynthetic anaerobe. Proc Natl Acad Sci USA 86(15):5753–5757
Pollock SV, Prout DL, Godfrey AC, Lemaire SD, Moroney JV (2004) The Chlamydomonas reinhardtii proteins CCP1 and CCP2 are required for long-term growth, but are not necessary for efficient photosynthesis, in a low-CO2 environment. Plant Mol Biol 56(1):125–132. doi:10.1007/s11103-004-2650-4
Price GD, Badger MR (1989a) Expression of human carbonic anhydrase in the cyanobacterium Synechococcus PCC7942 creates a high CO2-requiring phenotype : evidence for a central role for carboxysomes in the CO2 concentrating mechanism. Plant Physiol 91(2):505–513
Price GD, Badger MR (1989b) Isolation and characterization of high CO2-requiring-mutants of the cyanobacterium Synechococcus PCC7942: two phenotypes that accumulate inorganic carbon but are apparently unable to generate CO2 within the carboxysome. Plant Physiol 91(2):514–525. doi:10.1104/pp.91.2.514
Price G, Badger M (2002) Advances in understanding how aquatic photosynthetic organisms utilize sources of dissolved inorganic carbon for CO2 fixation. Funct Plant Biol 29(3):117–121
Price GD, Coleman JR, Badger MR (1992) Association of carbonic anhydrase activity with carboxysomes isolated from the cyanobacterium Synechococcus PCC7942. Plant Physiol 100(2):784–793
Price G, Maeda S-I, Omata T, Badger M (2002) Modes of active inorganic carbon uptake in the cyanobacterium, Synechococcus sp. PCC7942. Funct Plant Biol 29:131–149
Price GD, Woodger FJ, Badger MR, Howitt SM, Tucker L (2004) Identification of a SulP-type bicarbonate transporter in marine cyanobacteria. Proc Natl Acad Sci USA 101(52):18228–18233. doi:10.1073/pnas.0405211101
Price GD, Badger MR, Woodger FJ, Long BM (2008) Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J Exp Bot 59(7):1441–1461. doi:10.1093/Jxb/Erm112
Price GD, Badger MR, von Caemmerer S (2011) The prospect of using cyanobacterial bicarbonate transporters to improve leaf photosynthesis in C-3 crop plants. Plant Physiol 155(1):20–26. doi:10.1104/pp.110.164681
Rawat M, Henk MC, Lavigne LL, Moroney JV (1996) Chlamydomonas reinhardtii mutants without Ribulose-1,5-bisphosphate carboxylase-oxygenase lack a detectable pyrenoid. Planta 198(2):263–270
Sage RF (2004) The evolution of C4 photosynthesis. New Phytol 161(2):341–370. doi:10.1111/j.1469-8137.2004.00974.x
Sage RF, Christin PA, Edwards EJ (2011) The C-4 plant lineages of planet earth. J Exp Bot 62(9):3155–3169. doi:10.1093/Jxb/Err048
Sage RF, Sage TL, Kocacinar F (2012) Photorespiration and the evolution of C-4 photosynthesis. Annu Rev Plant Biol 63(63):19–47
Schou L, Benson AA, Bassham JA, Calvin M (1950) The path of carbon in photosynthesis. 11. The role of glycolic acid. Physiol Plantarum 3(4):487–495. doi:10.1111/j.1399-3054.1950.tb07676.x
Schwarz R, Reinhold L, Kaplan A (1995) Low activation state of ribulose-1,5-bisphosphate carboxylase/oxygenase in carboxysome-defective Synechococcus mutants. Plant Physiol 108(1):183–190. doi:10.1104/pp.108.1.183
Sharkey TD (1988) Estimating the rate of photorespiration in leaves. Physiol Plantarum 73(1):147–152. doi:10.1111/j.1399-3054.1988.tb09205.x
Shibata M, Ohkawa H, Katoh H, Shimoyama M, Ogawa T (2002) Two CO2 uptake systems in cyanobacteria: four systems for inorganic carbon acquisition in Synechocystis sp. strain PCC6803. Funct Plant Biol 29(2/3):123–129
S-i Maeda, Badger MR, Price GD (2002) Novel gene products associated with NdhD3/D4-containing NDH-1 complexes are involved in photosynthetic CO2 hydration in the cyanobacterium, Synechococcus sp. PCC7942. Mol Microbiol 43(2):425–435. doi:10.1046/j.1365-2958.2002.02753.x
So AK, John-McKay M, Espie GS (2002a) Characterization of a mutant lacking carboxysomal carbonic anhydrase from the cyanobacterium Synechocystis PCC6803. Planta 214(3):456–467
So AKC, Cot SSW, Espie GS (2002b) Characterization of the C-terminal extension of carboxysomal carbonic anhydrase from Synechocystis sp. PCC6803. Funct Plant Biol 29(3):183–194
Somanchi A, Handley E, Moroney J (1998) Chlamydomonas reinhardtii cDNAs upregulated in low-CO2 conditions: expression and analyses. Botany 76(6):1003–1009
Somerville CR, Ogren WL (1982) Genetic-modification of photo-respiration. Trends Biochem Sci 7(5):171–174. doi:10.1016/0968-0004(82)90130-X
Spalding MH, Spreitzer RJ, Ogren WL (1983a) Carbonic anhydrase-deficient mutant of Chlamydomonas reinhardii requires elevated carbon dioxide concentration for photoautotrophic growth. Plant Physiol 73(2):268–272
Spalding MH, Spreitzer RJ, Ogren WL (1983b) Reduced inorganic carbon transport in a CO2-requiring mutant of Chlamydomonas reinhardii. Plant Physiol 73(2):273–276
Spreitzer RJ (1993) Genetic dissection of Rubisco structure and function. Annu Rev Plant Physiol 44:411–434. doi:10.1146/annurev.pp.44.060193.002211
Spreitzer RJ, Salvucci ME (2002) Rubisco: structure, regulatory interactions, and possibilities for a better enzyme. Annu Rev Plant Biol 53:449–475. doi:10.1146/annurev.arplant.53.100301.135233
Tanaka S, Kerfeld CA, Sawaya MR, Cai F, Heinhorst S, Cannon GC, Yeates TO (2008) Atomic-level models of the bacterial carboxysome shell. Science 319(5866):1083–1086. doi:10.1126/science.1151458
Tcherkez GGB, Farquhar GD, Andrews TJ (2006) Despite slow catalysis and confused substrate specificity, all ribulose bisphosphate carboxylases may be nearly perfectly optimized. Proc Natl Acad Sci USA 103(19):7246–7251. doi:10.1073/pnas.0600605103
Tholen D, Zhu XG (2011) The mechanistic basis of internal conductance: a theoretical analysis of mesophyll cell photosynthesis and CO2 diffusion. Plant Physiol 156(1):90–105. doi:10.1104/pp.111.172346
Timm S, Mielewczik M, Florian A, Frankenbach S, Dreissen A, Hocken N, Fernie AR, Walter A, Bauwe H (2012) High-to-Low CO2 Acclimation Reveals Plasticity of the Photorespiratory Pathway and Indicates Regulatory Links to Cellular Metabolism of Arabidopsis. PLoS One 7(8):e42809. doi:10.1371/journal.pone.0042809
Tolbert NE (1973) Glycolate biosynthesis. Curr Top Cell Regul 7:21–50
Tsai Y, Sawaya MR, Cannon GC, Cai F, Williams EB, Heinhorst S, Kerfeld CA, Yeates TO (2007) Structural analysis of CsoS1A and the protein shell of the Halothiobacillus neapolitanus carboxysome. PLoS Biol 5(6):1345–1354. doi:10.1371/journal.pbio.0050144
Turner JS, Brittain EG (1962) Oxygen as a factor in photosynthesis. Biol Rev 37(1):130–170. doi:10.1111/j.1469-185X.1962.tb01607.x
von Caemmerer S, Evans JR (2010) Enhancing C-3 photosynthesis. Plant Physiol 154(2):589–592. doi:10.1104/pp.110.160952
von Caemmerer S, Quick WP, Furbank RT (2012) The development of C-4 rice: current progress and future challenges. Science 336(6089):1671–1672. doi:10.1126/science.1220177
Wang Y, Spalding MH (2006) An inorganic carbon transport system responsible for acclimation specific to air levels of CO2 in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 103(26):10110–10115. doi:10.1073/pnas.0603402103
Warburg O (1920) Über die Geschwindigkeit der photochemischen Kohlensäurezersetzung in lebenden Zellen. II. Biochemische Zeitschrift 103:188–217
Weber APM, von Caemmerer S (2010) Plastid transport and metabolism of C-3 and C-4 plants - comparative analysis and possible biotechnological exploitation. Curr Opin Plant Biol 13(3):257–265. doi:10.1016/j.pbi.2010.01.007
Whitney SM, Sharwood RE (2008) Construction of a tobacco master line to improve Rubisco engineering in chloroplasts. J Exp Bot 59(7):1909–1921. doi:10.1093/Jxb/Erm311
Whitney SM, Houtz RL, Alonso H (2011) Advancing our understanding and capacity to engineer nature’s CO2-sequestering enzyme Rubisco. Plant Physiol 155(1):27–35. doi:10.1104/pp.110.164814
Yeates TO, Kerfeld CA, Heinhorst S, Cannon GC, Shively JM (2008) Protein-based organelles in bacteria: carboxysomes and related microcompartments. Nat Rev Microbiol 6(9):681–691. doi:10.1038/nrmicro1913
Yu J-W, Price GD, Song L, Badger MR (1992) Isolation of a putative carboxysomal carbonic anhydrase gene from the cyanobacterium Synechococcus PCC7942. Plant Physiol 100(2):794–800. doi:10.1104/pp.100.2.794
Zelitch I, Ochoa S (1953) Oxidation and reduction of glycolic and glyoxylic acids in plants. 1. Glycolic acid oxidase. J Biol Chem 201(2):707–718
Zelitch I, Schultes NP, Peterson RB, Brown P, Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize in normal air. Plant Physiol 149(1):195–204. doi:10.1104/pp.108.128439
Zhu XG, Long SP, Ort DR (2010) Improving photosynthetic efficiency for greater yield. Ann Rev Plant Biol 61(61):235–261. doi:10.1146/annurev-arplant-042809-112206
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Moroney, J.V., Jungnick, N., DiMario, R.J. et al. Photorespiration and carbon concentrating mechanisms: two adaptations to high O2, low CO2 conditions. Photosynth Res 117, 121–131 (2013). https://doi.org/10.1007/s11120-013-9865-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11120-013-9865-7