
Abstract
Background and aims
In recent decades, temperate herbaceous wetlands have been widely experiencing shrub encroachment. By now, little has been known about whether such alteration in plant community would generate updated soil organic carbon (SOC) conservation strategies, and thus alter SOC dynamics.
Methods
To assess the temporal dynamics of SOC stock and fractions after shrub encroachment into temperate herbaceous wetlands, we measured floor litter mass, fine root biomass, and SOC stock and fractions (i.e. labile C pool I, labile C pool II, and recalcitrant C pool) across the 0–50 cm soil profile in the open wetlands and shrub islands (Betula fruticose and Salix floderusii) with different basal diameter sizes (small, 10 ~ 15 cm; medium, 15 ~ 20 cm; and large, 20 ~ 30 cm) in the Sanjiang Plain of Northeast China.
Results
Regardless of shrub species, SOC stock initially decreased after shrub expansion, but gradually recovered to the initial level of herbaceous wetlands. Moreover, SOC stock correlated positively with fine root biomass, but showed no significant relationship with floor litter mass. The responses of SOC fractions to shrub expansion mainly occurred in the surface soils (0–15 cm), despite a slight change in recalcitrant C pool in 15–50 cm layer. In the surface soils, labile C pool decreased substantially, whereas recalcitrant C pool and recalcitrant index of SOC elevated with increasing shrub sizes.
Conclusions
Shrub encroachment can recover SOC pool and enhance SOC recalcitrance in temperate herbaceous wetlands, and altered fine root biomass would account for shrub encroachment-induced SOC dynamics.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wetlands play an essential role in global carbon (C) cycle, with a storage of 37% of global terrestrial organic C in only 4 ~ 6% of the earth’s land surface (Roehm 2005). In wetlands, most organic C is stored in the soil, and even a small change in soil organic C (SOC) could have a disproportional influence on the terrestrial C cycle globally (Briggs et al. 2005; Cahoon et al. 2012; Lett et al. 2004). However, this huge soil C pool is vulnerable to land cover changes (Scharlemann et al. 2014). In recent decades, temperate herbaceous wetlands have been widely experiencing shrub encroachment as a result of global warming and human activities such as drainage (Bart et al. 2016; Saintilan and Rogers 2015), with Betula spp. and Salix spp. being commonly expanded shrub species (Castro-Morales et al. 2014; Favreau et al. 2019). By changing litter inputs, root distribution, soil abiotic environment, and decomposer community and activity (Cahoon et al. 2012; Cornelissen et al. 2007; McLaren et al. 2017; Myers-Smith et al. 2011), shrub encroachment can potentially pose a remarkable effect on SOC dynamics and thus soil C sequestration potential in wetlands.
In temperature herbaceous wetlands, SOC accumulation largely results from the imbalance between organic matter inputs and decomposition rates (Aerts et al. 1999; Bernal and Mitsch 2012; Fenner and Freeman 2020; Moore et al. 2007). Because of the substantial differences in functional traits between expanded shrubs and native herbaceous species (Cornelissen et al. 2007; Fenner and Freeman 2020; Zhang et al. 2020), shrub encroachment may change SOC characteristics in these wetlands. First, this shift in plant community can increase primary productivity and consequent organic matter inputs to soils, and the magnitude may increase with shrub size or coverage (Feng and Bao 2018; McKinley and Blair 2008; Montané et al. 2010). Second, shrub expansion can affect organic matter decomposition by altering substrate quality, but the effect may be species-specific (Cornelissen et al. 2007; Zhang et al. 2021). In addition, the intensified interspecific competition for resources between expanded shrubs and native herbaceous species, as well as soil drying, can alter plant root biomass allocation (Wang et al. 2016; Xie et al. 2008), which would cause a redistribution of root-derived organic matter inputs across the soil profile. Therefore, it is expected that shrub encroachment into temperate herbaceous wetlands would potentially affect SOC stock and its vertical distribution. However, the net effect of this shift in plant composition on SOC dynamics is still unclear due to the lack of empirical studies in these wetlands.
Soil organic C pool is composed of both labile and recalcitrant C fractions, and the long-term SOC storage is regulated by the long-lived recalcitrant C fractions (Trumbore and Czimczik 2008). Generally, SOC composition is related tightly to the characteristics of plant litter including senesced organs and root exudates (Trumbore and Czimczik 2008; Fenner and Freeman 2020). In temperate wetlands, shrub litter is often more lignified and contains a higher concentration of secondary compounds such as polyphenols than herbaceous litter (Cornelissen et al. 2007; Fenner and Freeman 2020; Zhang et al. 2020). Considering that mineral contents are relatively low in wetland soils, especially in the surface organic layer, the labile C fractions are preferentially exhausted by the decomposer community, and the recalcitrant C fractions tend to accumulate in soils (Leifield et al. 2012; Sollins et al. 1996). Thus, SOC pool would become biochemically-resistant to microbial decomposition following shrub expansion, which is beneficial for SOC accumulation in temperate herbaceous wetlands (Fenner and Freeman 2020). Unfortunately, the understanding of SOC recalcitrance responses to shrub encroachment remains limited in these carbon-rich wetlands.
Here, we measured aboveground plant biomass, floor litter mass, fine root biomass, soil bulk density, and SOC concentration and fractions in the open wetlands and two shrub (Salix floderusii and Betula fruticosa) islands with varying sizes in a herbaceous-dominated wetland in the Sanjiang Plain of Northeast China. The main objective of this study was to assess the effects of shrub encroachment on SOC stock and fractions in temperate herbaceous wetlands. We hypothesized that: (1) SOC pool would increase with shrub size due to the enhanced plant productivity and associated plant litter inputs; and (2) because of the greater concentrations of lignin and secondary compounds in shrub litter in relative to herbaceous litter (Fenner and Freeman 2020; Zhang et al. 2020), soil labile C fractions would decrease, but soil recalcitrant C fractions would increase with shrub size.
Materials and methods
Study site
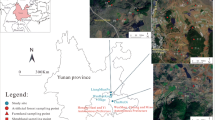
This study was performed in a freshwater wetland (47°35′N, 133°31′E, 56 m a. s. l) with an area of about 150 ha, which is a part of a wetland reserve (~ 200 km2) in the Sanjiang Plain of northeast China. The Sanjiang Plain is one of the largest freshwater wetlands in China, and has a temperate continental monsoon climate. Mean annual temperature is 3 °C, and mean annual precipitation is 560 mm. During the snow-free season, monthly mean water table in this wetland ranges from −14.6 cm to 8.5 cm. According to the US soil taxonomy, the soil is classified as an Inceptisol, and the parent material is quaternary alluvial sediments. Herbaceous-dominated wetlands are the main landscape in this region, and the single dominant species is Deyeuxia angustifolia. Two common deciduous shrubs, B. fruticose and S. floderusii, have been encroaching into the herbaceous wetlands as a result of intensive climate warming and agricultural development in the past decades (Zhang et al. 2021). The initial litter chemical properties of the dominant graminoid species and two expanded shrubs are shown in Tables S1 and S2. More detailed information on this study site is provided in Zhang et al. (2014, 2021).
Sample collection and analyses
Because it is difficult to determine expanded shrub age, we used the size of shrub basal diameter to assess the temporal change in SOC pool and fractions after shrub expansion. According to the distribution of shrub basal diameter in this wetland, both B. fruticose and S. floderusii islands were divided into three categories: small (10 ~ 15 cm in diameter), medium (15 ~ 20 cm in diameter), and large (20 ~ 30 cm in diameter) shrub islands. At each category, four shrub islands per species were randomly selected as replicates, and four corresponding open wetlands (at least 5 m away from any shrub canopy edge) were chosen as controls. The detailed information about shrub basal diameter and height in the selected shrub islands was detailed in Table 1. We set up three 0.5 m × 0.5 m quadrats located on the halfway between shrub base and canopy edge in each shrub island (Fig. 1), and three corresponding quadrats were randomly established in each replicate of the adjacent open wetlands. In the peak growing season (early August) in 2014, we collected herbaceous aboveground biomass and floor litter from three quadrats, mixed to form a composite sample, oven-dried to constant weight, and weighed. Subsequently, one soil core (5 cm in diameter) per quadrat was collected at 0–15, 15–30, and 30–50 cm depths, and then three soil cores per soil layer were mixed to determine SOC concentration and fractions. In addition, we collected soil cores (10 cm in diameter) at 0–15, 15–30, and 30–50 cm depths to determine bulk density and fine root biomass, respectively.

Diagram showing the locations of sampling quadrats in shrub islands
To determine soil bulk density, soil cores were oven-dried at 105 °C until constant mass, and weighed. To measure fine root biomass, soil cores were rinsed with distilled water on a 0.5 mm sieve, and the living fine roots (< 2 mm) were oven-dried at 65 °C to constant weight, and weighed. In each replicate, soil bulk density and fine root biomass were the mean values of three quadrats. To determine SOC concentration and fractions, soil samples were oven-dried at 65 °C after picking out plant roots and fauna with hand, ground (< 0.15 mm), and stored for chemical analyses. Soil organic C concentration was measured with the dry combustion method on a multi N/C 2100 analyzer (Analytik Jena, Germany). Given that the majority of SOC stored in the organic horizon in the wetlands of the study region (Zhang et al. 2008), we divided SOC into three fractions with the two-step acid hydrolysis approach (Rovira and Vallejo 2002): (1) Labile C pool I (LC I), obtained by hydrolysis with 2.5 M H2SO4 at 105 °C for 30 min; (2) Labile C pool II (LC II), obtained by hydrolysis with 13 M H2SO4 at room temperature overnight, and then with 1 M H2SO4 at 105 °C for 3 h; and (3) Recalcitrant C pool (RC), the unhydrolyzed residues. Organic C concentration of LC I, LC II, and RC was analyzed with the method described above. The LC I pool comprises either plant- or microbial-derived non-cellulosic polysaccharides, the LC II pool mainly consists of cellulose, and the RC pool is primarily comprised of recalcitrant organic compounds such as lignin, fats, waxes, and resins (Rovira and Vallejo 2002). The recalcitrant index (RI, %) was expressed as the proportion of RC in total SOC, and could indicate the degree of biochemical recalcitrance (Rovira and Vallejo 2002, 2007).
Calculation and statistical analyses
Considering the effects of shrub expansion on soil compactness, we used equivalent soil mass to calculate SOC stock according to Lee et al. (2009) and Mao et al. (2010). First, we calculated soil mass in the fixed layer (Mi, Mg ha−1, i = 1, ..., n) using the following formula:
where BDi is the bulk density of the ith soil layer (g cm−3), Di the thickness of ith soil layer (cm), and 100 is a unit conversion factor. Second, we calculated SOC stock in the fixed layer (Ci, fixed, Mg ha−1) as follows:
where Oi is the SOC concentration of ith soil layer (mg g−1), and 0.001 is a unit conversion factor. Last, we calculated the equivalent SOC stock (Cequiv, Mg ha−1) in the fixed layer as follows:
where Mequiv is the equivalent soil mass in the fixed layer, Madd is the additional soil mass that was used to calculate Mequiv in the fixed layer, and Obottom is the SOC concentration of Madd. In this study, we used the largest Mi value in the 0–50 cm layer as the equivalent mass (i.e. the value in the open wetlands), and used bulk density and SOC concentration in the 30–50 cm layer to calculate Madd and Cequiv.
The data were analyzed statistically in R 4.0.2 (R Core Team 2020) with the accepted significance level of 0.05. Data were tested for normality with the Shapiro–Wilk test. Non-normal data were logarithmically transformed before statistical analyses, but the raw untransformed data were reported in figures and tables. Linear mixed models in the “lmerTest” package were performed to assess how bulk density, SOC concentration, and recalcitrant index were affected by shrub species, category, soil layer, and their interactions (as fixed effects), and the replicate (as a random effect). One-way analysis of variance in the “multcomp” package, followed by Tukey’s (or Tamhane’s when equal variances not assumed) multiple comparison test, was used to examine the effects of shrub category on herbaceous aboveground biomass, floor litter mass, fine root biomass, bulk density, SOC concentration and fractions, SOC stock, and RI. Simple linear regression was performed to reveal the relationship between SOC stock and floor litter mass as well as fine root biomass.
Results
Herbaceous aboveground biomass, floor litter mass, and fine root biomass
Both B. fruticosa and S. floderusii islands generally had lower herbaceous aboveground biomass than open wetlands, and the magnitudes increased with shrub size (Table 1). However, no significant difference in floor litter mass was observed between shrub islands and open wetlands (Table 1). In addition, fine root biomass in each soil layer generally varied among categories for both B. fruticosa and S. floderusii islands (Table 1). Among the four categories, the large shrub islands had highest fine root biomass in 0–15 cm soil layer (Table 1). Regardless of shrub species, fine root biomass generally increased with increasing shrub size at both 15–30 cm and 30–50 cm depths (Table 1). In the 0–50 cm profile, only large B. fruticosa islands had higher fine root biomass than the open wetlands, whereas both medium and large S. floderusii islands had greater fine root biomass than the open wetlands (Table 1).
Soil bulk density and organic C concentration and stock
Shrub species, category, and soil layer had significant effects on soil bulk density, and the category effect varied with soil layer (Table 2). For both B. fruticosa and S. floderusii, large shrub islands had lower soil bulk density than open wetlands in both 0–15 cm and 15–30 cm soil layers, although no significant difference in 30–50 cm layer (Table 3).
Soil organic C concentration varied significantly with shrub species, category, soil layer, and the interactions of soil layer with shrub species and category (Table 2). In both 0–15 cm and 30–50 cm layers, small B. fruticosa islands had lower SOC concentration, whereas large B. fruticosa islands generally had higher SOC concentration than open wetlands (Table 3). Similarly, SOC concentration in 0–15 cm layer was lower in the small S. floderusii islands than in the open wetlands, despite no significant difference between open wetlands and S. floderusii islands with medium or large size (Table 3). In 15–30 cm layer, SOC concentration was higher in the large S. floderusii islands than in the open wetlands (Table 3). In addition, SOC concentration did not differ at 15–30 cm depth among B. fruticosa categories, and at 30–50 cm depth among S. floderusii categories (Table 3).
For both B. fruticosa and S. floderusii, SOC stock varied significantly among categories (Fig. 2). Soil organic C stock was always lowest in the small category, and gradually increased with increasing shrub basal diameter (Fig. 2). No significant difference in SOC stock was observed between open wetlands and large shrub islands (Fig. 2). In the 0–50 cm soil layer, SOC stock showed a positive relationship with fine root biomass, but did not correlate significantly with floor litter mass (Fig. 3).

Soil organic C stock in relation to shrub species and category in a temperate herbaceous wetland of Northeast China. The means (SE) with different letters are significantly different (P < 0.05, n = 4) for the same shrub species

Simple linear relationship between soil organic C stock and litter mass as well as fine root biomass in both open wetlands and shrub islands in a temperate herbaceous wetland of Northeast China
Labile and recalcitrant C pools
Both LC I and LC II concentrations varied with shrub species, category, soil layer, and the interaction between category and soil layer (Table 2). Irrespective of shrub species, small and medium shrub islands had the lowest LC I and LC II in 0–15 cm soil layer among the four categories (Fig. 4a and d). Moreover, LC I in 0–15 cm soil layer was lower in the large shrub islands than in the open wetlands (Fig. 4a and d). However, there was no significant difference in LC I and LC II among the four categories for any shrub islands in 15–30 cm and 30–50 soil layers (Fig. 4b, c, e, and f).

Soil organic C fractions (labile C I, LC I; labile C II, LC II; and recalcitrant C, RC) in relation to shrub species and category in a temperate herbaceous wetland of Northeast China. The recalcitrant indexes (RI) are shown in the inset graphs. The means (SE) with different letters are significantly different (P < 0.05, n = 4) for the same shrub species
Shrub category and soil layer significantly interacted to affect RC concentration and RI value (Table 2). In 0–15 cm layer, both B. fruticosa and S. floderusii islands generally had higher RC concentrations than open wetlands, and the magnitudes increased with shrub size (Fig. 4a and d). Accordingly, the RI value was higher in the shrub islands than in the open wetlands in 0–15 cm soil layer (Fig. 4a and d). Compared with open wetlands, only large S. floderusii islands had higher RC concentration and RI value in 15–30 cm layer (Fig. 4b and e), and only small B. fruticosa islands had lower RC concentration and RI value in 30–50 cm layer (Fig. 4c and f).
Discussion
Inconsistent with the first hypothesis, SOC stock initially decreased by 14.5 ~ 16.8% after shrub encroachment, but gradually recovered to the initial level as the shrub grew in this herbaceous-dominated wetland. In mesic grasslands, shrub encroachment was observed to promote organic C accrual in soils (McKinley and Blair 2008; Montané et al. 2010), and the magnitude increased with increasing shrub size (Feng and Bao 2018). Moreover, the positive relationship between SOC stock and fine root biomass indicated that SOC dynamics after shrub encroachment was primarily caused by the altered amount of root litter inputs to soils. Several studies also have pointed out that belowground litter inputs contribute more than aboveground inputs to soil C accrual in diverse ecosystems (e.g. Berhongaray et al. 2018; Rasse et al. 2005; Sokol and Bradford 2018). These findings imply that shrub encroachment can recover SOC pool primarily via enhanced root litter inputs, and highlight that shrub size should be considered to accurately assess the consequence of shrub encroachment on SOC dynamic in temperate herbaceous wetlands. Given the greater primary productivity of shrubs than herbs, such a shift in the growth form of dominant species can enhance ecosystem C sink potential in temperate wetlands.
Soil organic C dynamic is primarily controlled by the balance between litter inputs to soils and C emission from soils (Jackson et al. 2017; Lohila et al. 2011). Given that floor litter mass and fine root biomass in the small-size shrub islands were comparable with that in the open wetlands, the initial decrease in soil C stock could be primarily attributed to shrub encroachment-induced soil C loss through the following mechanisms. First, shrub encroachment into herbaceous wetlands generally aggravates soil drying and enhances oxygen diffusion (Bart et al. 2016; Saintilan and Rogers 2015), thereby accelerating microbial mineralization of organic C in the upper soil layer (Sulman et al. 2013). Second, shrub encroachment increased the allocation of fine root biomass toward deep soils, which could produce a priming effect on organic C mineralization by providing labile organic C fractions via root exudates and turnover (Cheng et al. 2013; Moore et al. 2020). In this study, the initial decrease in SOC concentration at both 0–15 and 30–50 cm depths as well as unchanged SOC concentration at 15–30 cm depth further confirmed these potential mechanisms. As the shrubs grew, enhanced C loss from soils would be offset by increased organic matter inputs to soils, especially for fine root-derived organic matter. Meanwhile, compared with herbaceous species, shrubs often produce plant litter containing recalcitrant organic compounds such as lignin and polyphenols (Tables S1 and S2), which would favor organic C accumulation in soils through its inhibiting effect on microbial growth and activity. Together, SOC stock eventually recovered to the initial level of herbaceous wetlands in the large-size shrub islands.
Soil labile C fractions decreased, and recalcitrant C fractions increased only in 0–15 cm layer after shrub encroachment (Fig. 4). Moreover, the response of recalcitrant C fractions in the subsurface soils to shrub expansion varied with shrub species, despite unchanged labile C fractions (Fig. 4). Thus, the results partly supported the second hypothesis. In general, the biochemical recalcitrance of organic C is tightly related to plant trait identity (Ahmed et al. 2016). Compared with herbaceous species, shrub litter was more lignified, and contained higher concentrations of secondary compounds such as total polyphenol and condensed tannins in this wetland (Tables S1 and S2). Accordingly, shrub encroachment could replenish SOC pool primarily by altering the quantity of recalcitrant C fractions, especially in the surface soils. These observations imply that the divergent responses of labile and recalcitrant C fractions across soil profiles should be fully considered to assess the consequence of shrub encroachment on SOC stability in graminoid-dominated wetlands.
In wetlands, shrub encroachment is often associated with soil drying and elevated oxygen availability in the surface soils (Fenner and Freeman 2020). Accordingly, this shift in plant composition can unlock soil enzyme latch and enhance the activities of hydrolytic and oxidative enzymes, especially phenoloxidase activity (Fenner and Freeman 2011, 2020). In the surface organic layer, labile C fractions is not associated as organo-mineral complexes due to the relatively low mineral content, and thus is susceptible to microbial degradation (Leifield et al. 2012; Rovira and Vallejo 2002). Moreover, expanded shrubs produced more biochemically-recalcitrant plant litter than herbaceous species (Tables S1 and S2), which could protect organic C from microbial attack, especially under aerobic conditions (Fenner and Freeman 2020). Taking together, soil microbial decomposer could preferentially use the labile C fractions, leading to a net loss of labile C after shrub encroachment. Similarly, recalcitrant C accumulated in surface soils because of the selective preservation (Sollins et al. 1996), and the magnitude increased with elevating shrub size. These findings suggest that shrub encroachment will cause a shift of SOC accumulation strategy from anaerobic protection to biochemical protection in the surface soils in the herbaceous-dominated wetlands.
In the subsurface soils, the response of recalcitrant C pool to shrub encroachment was species-specific in this wetland. Compared with surface soils, SOC dynamics in the subsurface soils are largely influenced by root litter inputs rather than aboveground plant litter inputs (Bernal et al. 2016). Thus, the different responses of soil recalcitrant C pools in the subsurface soils would be explained by the contrasting allocation patterns of fine root biomass between S. floderusii and B. fruticosa islands. At 15–30 cm depth, fine root biomass increased only after S. floderusii encroachment in this wetland, and the magnitudes increased with increasing shrub size (Table 1). Moreover, S. floderusii produced fine roots with relatively higher total polyphenols and condensed tannins than the graminoid species (Table S2). Consequently, soil recalcitrant C pool increased with shrub size following S. floderusii expansion in this wetland.
In 30–50 cm soil layer, fine root biomass increased with shrub size after B. fruticosa expansion. In the deep soil layer, microbial growth and activity are often limited by fresh organic matter inputs (Lützow et al. 2006). Consequently, the increased fine root biomass after B. fruticosa expansion could stimulate microbial decomposition of existing stabilized soil organic matter (Cheng et al. 2013; Moore et al. 2020; Pausch et al. 2016), thereby decreasing the amount of recalcitrant C in the 30–50 cm depth. As shrub grew, increased fine root turnover could offset organic C loss, and recalcitrant C accumulated in the deep soils. Therefore, recalcitrant C pool initially decreased after B. fruticosa encroachment, but gradually recovered to the level of the open wetlands. These results indicate that altered fine root biomass after shrub expansion would be an overriding factor controlling the dynamics of SOC fractions in deep soil in temperate herbaceous-dominated wetlands.
In temperate wetlands, SOC dynamics are influenced by a series of abiotic and biotic factors such as plant litter inputs, water-logging and associated anaerobic conditions, nutrient availability, and soil microbial community and activity (Aerts et al. 1999; Roehm 2005; Bernal and Mitsch 2012; Fenner and Freeman 2020). In this study, we compared the differences in amounts of aboveground litter and fine roots among open wetlands and shrub islands, and observed the important role of fine root biomass in driving shrub expansion-induced alterations of SOC stock in this temperate herbaceous wetland. However, shrub expansion is often associated with soil drying, altered microclimates, and soil microbial growth and composition (Fenner and Freeman 2020; Zhang et al. 2021), which would exert a substantial influence on SOC dynamics in the herbaceous wetlands. To fully understand the underlying mechanisms causing these changes in SOC stock and fractions, further studies should pay attention to the dynamics of abiotic factors and soil microbial properties after shrub expansion in temperate herbaceous wetlands.
Conclusions
In summary, SOC stock decreased initially after shrub encroachment, but gradually recovered to the level of open wetlands as the shrub grew. Moreover, the effects of shrub expansion on SOC fractions occurred mainly in the surface soils, despite a slight change in recalcitrant C fractions in the subsurface soils. In the surface soils, labile C fractions decreased dramatically, but recalcitrant C fractions exhibited a pronounced increasing trend after shrub encroachment. These results indicate that, despite unchanged SOC stock, shrub encroachment eventually enhanced SOC stability by increasing recalcitrant C fractions. Thus, shrub encroachment would mitigate warming- and drying-induced SOC loss mainly via increased recalcitrant C pool in temperate herbaceous-dominated wetlands.
Compared with floor litter mass, the positive relationship between fine root biomass and SOC stock indicates that fine root-derived organic matter may exert a stronger influence on SOC dynamics than aboveground litter inputs in this wetland. Moreover, altered fine root biomass after shrub expansion would be a key factor causing the changes in recalcitrant C pool in subsurface soils in herbaceous-dominated wetlands. Unfortunately, we did not separate shrub roots from graminoid roots, which could not discriminate the potential source of organic C in subsurface soils. Therefore, further studies regarding the effects of shrub encroachment on SOC dynamics should take fine roots into special consideration, including species composition, the vertical pattern across soil profiles, root exudates, and turnover rates in temperate herbaceous-dominated wetlands.
References
Aerts R, Verhoeven JTA, Whigham DF (1999) Plant-mediated controls on nutrient cycling in temperate fens and bogs. Ecology 80:2170–2181
Ahmed IU, Smith AR, Jones DL, Godbold DL (2016) Tree species identity influences the vertical distribution of labile and recalcitrant carbon in a temperate deciduous forest soil. For Ecol Manag 359:352–360
Bart D, Davenport T, Yantes A (2016) Environmental predictors of woody plant encroachment in calcareous fens are modified by biotic and abiotic land-use legacies. J Appl Ecol 53:541–549
Berhongaray G, Cotrufo MF, Jassens IA, Ceulemans R (2018) Below-ground carbon inputs contribute more than above-ground inputs to soil carbon accrual in a bioenergy poplar plantation. Plant Soil 434:363–378
Bernal B, Mitsch WJ (2012) Comparing carbon sequestration in temperate freshwater wetland communities. Glob Chang Biol 18:1636–1647
Bernal B, Mckinley DC, Hungate BA, White PM, Mozdzer TJ, Megonigal JP (2016) Limits to soil carbon stability; deep, ancient soil carbon decomposition stimulated by new labile organic inputs. Soil Biol Biochem 98:85–94
Briggs JM, Knapp AK, Blair JM, Heisler JL, Hoch GA, Lett MS, Mccarron JK (2005) An ecosystem in transition: causes and consequences of the conversion of Mesic grassland to shrubland. Bioscience 55:243–254
Cahoon SM, Sullivan PF, Shaver GR, Welker JM, Post E, Holyoak M (2012) Interactions among shrub cover and the soil microclimate may determine future Arctic carbon budgets. Ecol Lett 15:1415–1422
Castro-Morales LM, Quintana-Ascencio PF, Fauth JE, Ponzio KJ, Hall DL (2014) Environmental factors affecting germination and seedling survival of carolina willow (Salix Caroliniana). Wetlands 34:469–478
Cheng W, Parton WJ, Gonzalez-Meler MA, Phillips R, Asao S, McNickle GG, Brzostek E, Jastrow JD (2013) Synthesis and modeling perspectives of rhizosphere priming. New Phytol 201:31–44
Cornelissen JH, van Bodegom PM, Aerts R, Callaghan TV, van Logtestijn RS, Alatalo J, Chapin FS, Gerdol R, Gudmundsson J, Gwynn-Jones D, Hartley AE, Hik DS, Hofgaard A, Jonsdottir IS, Karlsson S, Klein JA, Laundre J, Magnusson B, Michelsen A, Molau U, Onipchenko VG, Quested HM, Sandvik SM, Schmidt IK, Shaver GR, Solheim B, Soudzilovskaia NA, Stenstrom A, Tolvanen A, Totland O, Wada N, Welker JM, Zhao X, Team MOL (2007) Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol Lett 10:619–627
Favreau M, Pellerin S, Poulin M (2019) Tree encroachment induces biotic differentiation in Sphagnum-dominated bogs. Wetlands 39:841–852
Feng D, Bao W (2018) Shrub encroachment alters topsoil C:N:P stoichiometric ratios in a high-altitude forest cutover. iForest 11:594–599
Fenner N, Freeman C (2011) Drought-induced carbon loss in peatlands. Nat Geosci 4:895–900
Fenner N, Freeman C (2020) Woody litter protects peat carbon stocks during drought. Nat Clim Chang 10:363–369
Jackson RB, Lajtha K, Crow SE, Hugelius G, Kramer MG, Pineiro G (2017) The ecology of soil carbon: pools, vulnerabilities, and biotic and abiotic controls. Annu Rev Ecol Evol Syst 48:419–445
Lee J, Hopmans JW, Rolston DE, Baer SG, Six J (2009) Determining soil carbon stock changes: simple bulk density corrections fail. Agric Ecosyst Environ 134:251–256
Leifield J, Steffens M, Galego-Sala A (2012) Sensitivity of peatland carbon loss to organic matter quality. Geophys Res Lett 39:L14704
Lett MS, Knapp AK, Briggs JM, Blair JM (2004) Influence of shrub encroachment on aboveground net primary productivity and carbon and nitrogen pools in a Mesic grassland. Can J Bot 82:1363–1370
Lohila A, Minkkinen K, Aurela M, Tuovinen JP, Penttilä T, Ojanen P, Laurila T (2011) Greenhouse gas flux measurements in a forestry-drained peatland indicate a large carbon sink. Biogeosciences 8:3203–3218
Lützow M, Kogel-Knabner I, Ekschmitt K, Matzner E, Guggenberger G, Marschner B, Flessa H (2006) Stabilization of organic matter in temperate soils: mechanisms and their relevance under different soil conditions - a review. Eur J Soil Sci 57:426–445
Mao R, Zeng DH, Hu YL, Li LJ, Yang D (2010) Soil organic carbon and nitrogen stocks in an age-sequence of poplar stands planted on marginal agricultural land in Northeast China. Plant Soil 332:277–287
McKinley DC, Blair JM (2008) Woody plant encroachment by Juniperus virginiana in a Mesic native grassland promotes rapid carbon and nitrogen accrual. Ecosystems 11:454–468
McLaren JR, Buckeridge KM, van de Weg MJ, Shaver GR, Schimel JP, Gough L (2017) Shrub encroachment in Arctic tundra: Betula nana effects on above- and belowground litter decomposition. Ecology 98:1361–1376
Montané F, Romanyà J, Rovira P, Casals P (2010) Aboveground litter quality changes may drive soil organic carbon increase after shrub encroachment into mountain grasslands. Plant Soil 337:151–165
Moore TR, Bubier JL, Bledzki L (2007) Litter decomposition in temperate peatland ecosystems: the effect of substrate and site. Ecosystems 10:949–963
Moore JAM, Sulman BN, Mayes MA, Patterson CM, Classen AT, Sayer E (2020) Plant roots stimulate the decomposition of complex, but not simple, soil carbon. Funct Ecol 34:899–910
Myers-Smith IH, Forbes BC, Wilmking M, Hallinger M, Lantz T, Blok D, Tape KD, Macias-Fauria M, Sass-Klaassen U, Lévesque E (2011) Shrub expansion in tundra ecosystems: dynamics, impacts and research priorities. Environ Res Lett 6:045509
Pausch J, Loeppmann S, Kuehnel A, Forbush K, Kuzyakov Y, Cheng W (2016) Rhizosphere priming of barley with and without root hairs. Soil Biol Biochem 100:74–82
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed 6 Aug 2020
Rasse DP, Rumpel C, Dignac MF (2005) Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 269:341–356
Roehm CL (2005) Respiration in wetland ecosystems. Oxford University Press, New York
Rovira P, Vallejo VR (2002) Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil an acid hydrolysis approach. Geoderma 107:109–141
Rovira P, Vallejo VR (2007) Labile, recalcitrant, and inert organic matter in Mediterranean forest soils. Soil Biol Biochem 39:202–215
Saintilan N, Rogers K (2015) Woody plant encroachment of grasslands: a comparison of terrestrial and wetland settings. New Phytol 205:1062–1070
Scharlemann JPW, Tanner EVJ, Hiederer R, Kapos V (2014) Global soil carbon: understanding and managing the largest terrestrial carbon pool. Carbon Manag 5:81–91
Sokol NW, Bradford MA (2018) Microbial formation of stable soil carbon is more efficient from belowground than aboveground input. Nat Geosci 12:46–53
Sollins P, Homann P, Caldwell BA (1996) Stabilization and destablization of soil organic matter: mechanisms and controls. Geoderma 74:65–105
Sulman BN, Desai AR, Mladenoff DJ (2013) Modeling soil and biomass carbon responses to declining water table in a wetland-rich landscape. Ecosystems 16:491–507
Trumbore SE, Czimczik CI (2008) An uncertain future for soil carbon. Science 321:1455–1456
Wang P, Mommer L, van Ruijven J, Berendse F, Maximov TC, Heijmans MMPD (2016) Seasonal changes and vertical distribution of root standing biomass of graminoids and shrubs at a Siberian tundra site. Plant Soil 407:55–65
Xie Y, Luo W, Wang K, Ren B (2008) Root growth dynamics of Deyeuxia angustifolia seedlings in response to water level. Aquat Bot 89:292–296
Zhang W, Xiao H, Tong C, Su Y, Xiang W, Huang D, Syers JK, Wu J (2008) Estimating organic carbon storage in temperate wetland profiles in Northeast China. Geoderma 146:311–316
Zhang X, Mao R, Gong C, Yang G, Lu Y (2014) Effects of hydrology and competition on plant growth in a freshwater marsh of Northeast China. J Freshw Ecol 29:117–128
Zhang X, Wang Y, Jiang W, Mao R (2020) Effect of expanded shrub litter on decomposition of graminoid litter in a temperate freshwater marsh. Plant Soil 451:409–418
Zhang X, Jiang W, Jiang S, Tan W, Mao R (2021) Differential responses of litter decomposition in the air and on the soil surface to shrub encroachment in a graminoid-dominated temperate wetland. Plant Soil. https://doi.org/10.1007/s11104-021-04893-1
Acknowledgements
This study was financed by the National Natural Science Foundation of China (Nos. 41671091 and 31570479). We thank three anonymous reviewers and the editor for their helpful comments on this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Zucong Cai .
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zhang, X., Jiang, S., Jiang, W. et al. Shrub encroachment balances soil organic carbon pool by increasing carbon recalcitrance in a temperate herbaceous wetland. Plant Soil 464, 347–357 (2021). https://doi.org/10.1007/s11104-021-04975-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-021-04975-0