
Abstract
Warming and elevated atmospheric CO2 (eCO2) can elicit contrasting responses on different SOM pools, thus to understand the effects of combined factors it is necessary to evaluate individual pools. Over two years, we assessed responses to eCO2 and warming of SOM pools, their susceptibility to decomposition, and whether these responses were mediated by plant inputs in a semi-arid grassland at the PHACE (Prairie Heating and CO2 Enrichment) experiment. We used long-term soil incubations and assessed relationships between plant inputs and the responses of the labile and resistant pools. We found strong and contrasting effects of eCO2 and warming on the labile C pool. In 2008 labile C was increased by eCO2 and was positively related to plant biomass. In contrast, in 2007 eCO2 and warming had interactive effects on the labile C, and the pool size was not related to plant biomass. Effects of warming and eCO2 in this year were consistent withtreatment effects on soil moisture and temperature and their effects on labile C decomposition. The decomposition rate of the resistant C was positively related to indicators of plant C inputs. Our approach demonstrated that SOM pools in this grassland can have early and contrasting responses to climate change factors. The labile C pool in the mixed-grass prairie was highly responsive to eCO2 and warming but the factors behind such responses were highly dynamic across years. Results suggest that in this grassland the resistant C pool could be negatively affected by increases in plant-production driven available soil C.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
There is a high degree of uncertainty on the combined effects of climate warming and atmospheric CO2 increase on soil organic matter (SOM) dynamics (Pendall et al. 2004; Shen et al. 2009). Through their effects on plant productivity, warming and eCO2 can impact SOM dynamics, both by directly determining inputs and/or by affecting rates of losses. Changes in plant productivity and biomass turnover can alter SOM pools by affecting the rates at which belowground production is incorporated into SOM via root death, slough off and exudation (Crow et al. 2009; Belay-Tedla et al. 2009). Increasing CO2 generally increases above and belowground biomass (Morgan et al. 2004a; Jastrow et al. 2000; de Graaff et al. 2006) as well as root growth rate and turnover (Allard et al. 2005; Norby et al. 2004; Milchunas et al. 2005). Various meta-analyses have found that eCO2 generally increases total soil C pools (Luo et al. 2006; Hungate et al. 2009; Jastrow et al. 2005). Warmer temperatures can also increase or limit above and belowground production, biomass and root turnover (Belay-Tedla et al. 2009; Rustad et al. 2001; Wan et al. 2004, 2005). The flux of recent photosynthate associated with increased productivity and turnover can be a major source of readily available C (Hutsch et al. 2002; Weintraub et al. 2007). For example, recent photosynthate has been found to contribute about 40% of the dissolved organic C (DOC) pool (Högberg and Högberg 2002; Giesler et al. 2007). Increases in the availability of plant-derived labile C due to CO2 elevation and/or warming may stimulate SOM decomposition and reduce the stock of soil C, an effect known as priming (Dijkstra and Cheng 2007a; Fontaine et al. 2007; Kuzyakov 2002). Stimulation of SOM decomposition has been found to be correlated to root biomass (Fu et al. 2002) and leaf biomass (Dijkstra et al. 2006) suggesting that the extent of the stimulation could be controlled by the magnitude of the effect of CO2 and warming on plant productivity.
Warming and eCO2 can have contrasting effects on different SOM pools. Cardon et al. (2001) showed that eCO2 retarded the decay of resistant, slow-turnover SOM while it increased the decomposition of labile, fast-turnover SOM. Cheng et al. (2007) observed that under eCO2 more plant derived C was stored in the resistant pool and that its decay rate decreased. In contrast, Gill et al. (2002) found plant growth at increased CO2 led to losses in resistant SOM, as measured by mineral-associated soil C. Other studies have observed that eCO2 depletes both resistant SOM as well as fractions dominated by fresh plant derived C (de Graaff et al. 2009; Del Galdo et al. 2006). Because the decomposition of labile SOM fractions is theoretically less sensitive to temperature than that of more resistant fractions (Davidson and Janssens 2006) it is likely that the effects of warming will also depend on the SOM pool examined (e.g. Belay-Tedla et al. 2009). Further, the regulation of SOM dynamics by soil moisture might not be equivalent for different pools (Garten et al. 2009). To understand the mechanisms underlying the combined effects of multiple global change factors on long-term soil C storage, effects on individual C pools and their kinetics should be evaluated.
The direction and magnitude of combined effects of eCO2 and warming are likely to be dependent on ecosystem type (Norby and Luo 2004; Luo et al. 2008). In moisture limited systems such as semi-arid grasslands, the impacts of CO2 and temperature on soil C are likely to be mediated by their effects on soil water availability, which can regulate both decomposition and plant productivity and thus plant inputs. Warming may induce soil drying (Rustad et al. 2001), potentially limiting plant productivity and SOM decomposition (Liu et al. 2009). However, increases in soil moisture associated with enhanced plant water use efficiency under eCO2 (Morgan et al. 2004b) could counteract this effect. For semi-arid grasslands, models predict strong responses of plant production and soil C storage to CO2 elevation and warming as well as interactive effects of the combined factors (Parton et al. 2007; Pepper et al. 2005; Luo et al. 2008), but empirical assessments are lacking. Our field experiment (PHACE: Prairie Heating and CO2 Enrichment), in a semi-arid, native northern mixed grass prairie in Wyoming, USA, was modeled in advance by Parton et al. (2007). Modeling results spanning ten years after the beginning of treatments predict a high dependency of soil C flux on yearly variations in soil moisture availability. A gradual decrease in soil organic C is predicted for plots exposed to eCO2 (with or without warming) due to faster decomposition associated with higher soil moisture under eCO2, which would surpass the effects of increased plant C input.
Here, we present an empirical evaluation of the impacts of eCO2 and warming on SOM at the PHACE experiment. We investigated two main questions: 1) how do eCO2, warming and their combination affect SOM pools and their susceptibility to decomposition? and 2) are the responses of SOM pools and their susceptibility to decomposition associated with plant inputs? To address these questions, we conducted long-term laboratory incubations because they provide a more sensitive indicator of SOM responses to climate change than bulk soil measurements (Pendall and King 2007; Langley et al. 2009). Incubations under optimal conditions allow assessments of SOM pools and kinetics while removing the environmental constraints and confounding effects at play in a field situation (Davidson and Janssens 2006). The rate of microbial respiration during laboratory incubations and the kinetic parameters that can be derived from them can be used to assess changes in the susceptibility of C pools to decomposition due to previous field exposure to experimental treatments (Taneva and Gonzalez-Meler 2008; Collins et al. 2000; Carney et al. 2007). In order to examine the role of plant inputs on SOM pool sizes and their susceptibility to decomposition, we assessed whether there were relationships between indicators of plant inputs (aboveground biomass, fine root biomass and DOC measured at the time of soil collection) and the responses of the labile and resistant pools.
Materials and methods
Study site
The Prairie Heating and CO2 Enrichment (PHACE) experiment at the USDA-ARS High Plains Grassland Research Station (HPGRS) is located about 15 km west of Cheyenne, WY, USA (41° 11′ N, 104° 54′ W; 1930 m.a.s.l.). The ecosystem is a northern mixed grass prairie with a plant community dominated by the C4 grass Bouteloua gracilis (H.B.K) Lag. and two C3 grasses, Hesperostipa comata Trinand Ruprand Pascopyrum smithii (Rydb.). About 20% of the vegetation is composed of sedges and forbs. Annual precipitation is 384 mm, with ca. 60% falling during the growing season, which extends from late March through September. Mean air temperatures are 17.5°C in summer and −2.5°C in winter, with average maximum July temperature of 27°C (meteorological data collected on-site and compiled by HPGRS). Soils at the experimental site are Mollisols (fine-loamy, mesic Aridic Argiustoll, mixed Ascalon and Altvan series), with a pH of 7.0. Average organic soil C content is 1.9% (S.D. = 0.27) at 0–5 cm and 1.5% (S.D. = 0.25) at 5–15 cm. Average carbonate content by weight is 0.0045 % at 0–5 cm (S.D. = 0.062) and 0.2056 % (S.D. = 0.61) at 5–15 cm. The experiment uses Free Air CO2 Enrichment (FACE) technology (Miglietta et al. 2001) installed in 3.4-m diameter rings to produce an atmosphere with a CO2 concentration of 600 ± 40 μmol mol−1 in the eCO2 treatments during the growing season. Infrared heaters (Kimball et al. 2008) attached to a frame 1.5 m above the ground increase air temperature by 1.5°C during daytime and 3°C at night all year round. The plots are thus exposed to four combinations of CO2 and temperature (ambient CO2 and ambient temperature: ct; ambient CO2 and elevated temperature: cT; eCO2 and ambient temperature: Ct; eCO2 and elevated temperature: CT) with five replications. CO2 enrichment began at the start of the growing season (early April) in 2006 and heating began at the start of the growing season in 2007.
Field measurements and sample processing
In each experimental plot, soil volumetric water content is monitored hourly at 10 and 20 cm (Envirosmart sensors, Sentek Sensor Technologies, Stepney, Australia). Soil temperature is also measured hourly at 3 and 10 cm using thermocouples. Daily means of soil water content and temperature were averaged across the six-week periods prior to soil sampling (Shim et al. 2009). Standing fine root biomass and aboveground plant biomass were assessed at the peak of the growing season (late July, close to the time of soil collection) in 2007 and 2008. Fine roots (<1 mm) were removed from soil samples (see below). Aboveground biomass was obtained by clipping an area of 0.75 m2 in each plot.
In June 2007 and July 2008 three soil cores (15-cm deep; 3-cm diameter) were collected from each plot, divided into 0–5 and 5–15 cm depths, consistent with pretreatment protocols, and composited into one sample per depth. Fresh soils were sieved to 2 mm and further inspected to remove visible litter and root material. To obtain a measure of DOC, a 15-g subsample of fresh soil was extracted by shaking in 30 ml of 0.05 M K2SO4 for 1 hour. The filtered extract was analyzed for total C (Shimadzu TOC-VCPN, Shimadzu Scientific Instruments, Wood Dale, IL). Bulk soil organic C content was determined for a subsample of air-dried soil which was ground to powder and analyzed with a Costech EA 1108 Element Analyzer. For incubations we used soil from 0 cm to 15 cm in depth, which contains very low and mostly undetectable levels of carbonates (see Study site). However, prior to analyses, and for consistency, all soil samples and extracts from our experiment are routinely treated to remove carbonates as deeper soils have higher contents that require removal. Extracts were treated with 1 M H3PO4 (1 μl per 10 ml of extract) and soil samples were shaken with 1 M H3PO4 (Sherrod et al. 2002).
Incubations
On the day after collection, subsamples (15–20 g) of root/litter-free 2-mm sieved soil were placed in polystyrene beakers which had small holes punched into the bases and pre-combusted glass fiber filters in the bottom for drainage (Townsend et al. 1997). Sub-samples of soils collected from three and five experimental replicates were incubated in 2007 and 2008 respectively. Deionized water was added to each beaker to reach 60% of the soil’s field capacity. Each beaker was placed into a 500-ml canning jar with a lid modified to hold a 1.5-cm, blue butyl rubber stopper. Soils were incubated at 25°C in a controlled environment incubator. Headspace samples (30 ml) were collected at 1, 2, 5, 8, 9, 12, 22, 33, 43, 79 and 107 days of incubation in 2007 and at 1, 2, 5, 9, 13, 22, 30, 45, 78 and 114 days in 2008. Prior to headspace sampling jars were opened to allow equilibration with ambient air and then closed and flushed with CO2-free air for 2 min to bring the headspace CO2 concentration to zero. Jars were then immediately placed in the incubator for periods ranging from 24 h during the early days of incubation to 50 hours at the final sampling date, to allow approximately 1000 μmol mol−1 CO2 to accumulate. CO2 production rate per hour was calculated based on the length of time after flushing. Four empty jars were used as controls. Headspace samples were analyzed for CO2 concentration with an infrared gas analyzer (Li-Cor 820, LICOR Inc. Lincoln, NE) calibrated with four standard gases over the concentration range 377–4500 μmol mol−1.
To estimate C kinetic parameters, measured CO2 production rates over time were fitted to a two pool decay model in which a pool of labile C decays exponentially and a pool of resistant C decays at a constant rate (Dijkstra and Cheng 2007a; Wedin and Pastor 1993):
Where Rt = respiration rate at time t (μg C g soil−1 day−1); Ri = initial rate of decomposition of the labile pool (at the start of the incubation; mg C g soil−1 day−1); kl = decay rate of C in the labile pool (day−1); Rr = decomposition rate of C in resistant pool (mg C g soil−1 day−1). The size of the labile pool (Cl) was defined as Ri/kl. Curve fitting was done using the single, 3 parameter exponential decay equation in Sigma Plot 10.0. To account for pre-experimental soil C conditions, Ri, Cl and Rr were standardized by dividing the estimated values by total soil C content, measured for each plot and at each depth. The labile pool is reported as mg C g soil C−1 and the decomposition rates of the labile and resistant pools as mg C g soil C−1 day−1.
Statistical analyses
To determine the effects of depth, CO2 and warming treatments on field soil moisture, temperature,and estimated parameters (size of the labile C pool, the rates of decomposition of the labile and resistant pool) over the course of incubation we used a split plot, three-factor ANOVA with CO2, warming and depth as fixed effects and replicate and plot × depth as random effects. Each one of the incubations was analyzed as an independent experiment. We preferred this approach to a single model analysis as there was a small (6%) difference in the duration of the incubation and slightly more rapid drying of the 2007 soils in the last few weeks of incubation. These differences could have influenced the magnitudeof some of the estimated parameters across all treatments, but were not expected to have influenced treatment effects. Consequently, our analyses and interpretations focus on the treatment effects observed in each year and the variables driving them rather than on the comparison of absolute magnitudes of the pools and rates across years. Aboveground plant biomass, root biomass, DOC and average daily soil water content means for the six weeks prior to soil sampling were linearly regressed against the estimated pool sizes and kinetic parameters. Significant effects and relationships are reported at p < 0.05 unless otherwise stated. All statistical analyses were performed with JMP (version 8.0; SAS Institute, Cary, N.C. USA).
Results
Effects of field treatments on soil temperature and moisture
Average soil volumetric water content ranged from 8 to 13% at 0–10 cm depth, and 12 to 18% at 10–20 cm depth (Table 1). Over the six weeks previous to mid-season sampling in 2007 and 2008, soil water content was significantly greater (by 2.1% water content on average; Table 1) in eCO2 than ambient CO2 plots (p = 0.03 for 2007 and p = 0.002 for 2008). Warming significantly decreased water content prior to all sampling dates by 2.0% on average (p = 0.04 for 2007 and p = 0.006 for 2008; Table 1). No interactions between treatment and depth were observed for soil moisture.
eCO2 significantly decreased soil temperature by 0.2°C in the period before sampling in 2007 and 0.3°C in 2008 (p = 0.07, p = 0.0001 respectively). Warming significantly increased soil temperature during all periods (Table 1). The impact of warming depended on soil depth (significant interaction of warming and soil depth, p = 0.0004 for 2007 and p = 0.0003 for 2008), as the increase in soil temperature was greater at 3 cm than at 10 cm (2.8°C vs 2.1°C on average in 2007 and 2.1°Cvs 1.8°C in 2008).
Plant biomass and DOC
eCO2 increased fine root biomass in 2007 and 2008 and aboveground biomass in 2008 (Table 2). There was no main effect of warming, but the combination of eCO2 and warming tended to enhance fine root biomass in 2008 (marginally significant CO2 x warming interaction; Table 2). eCO2 had no detectable effect on DOC while warming tended to increase DOC concentration in 2008 (Table 2). Aboveground plant biomass and fine root biomass were positively correlated (Fig. 1). The concentration of DOC in soil was positively correlated to both aboveground plant biomass and root biomass (Fig. 2).

Linear relationship between above-ground plant biomass and fine root biomass (0–15 cm depth) at the time of peak biomass (late July) in 2007 and 2008.ct: ambient CO2 and ambient temperature; cT: ambient CO2 and elevated temperature; Ct: elevated CO2 and ambient temperature; CT: elevated CO2 and elevated temperature

Linear relationship between field measurements of dissolved organic C in soil (0–15 cm; DOC) and above-ground plant biomass a and fine root biomass (0–15 cm) b at the time of peak biomass in 2007 and 2008. ct: ambient CO2 and ambient temperature; cT: ambient CO2 and elevated temperature; Ct: elevated CO2 and ambient temperature; CT: elevated CO2 and elevated temperature
Estimated pool sizes and decomposition rates
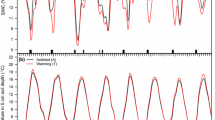
The proportion of labile C and its initial decomposition rates showed strong interactive effects of CO2 and warming in the 0–5 cm layer in soils sampled in 2007. Warming increased the proportion of total C that was labile in soils from ambient CO2 plots but decreased it in eCO2 soils (Fig. 3a). In 2007 the decomposition rate of the labile C was lower with warming in ambient CO2 soils, while it increased under warming in eCO2 soils (Fig. 3c). In 2008, the labile proportion was greater in eCO2 soils and this effect was consistent across depths (Fig. 3b). The resistant pool decomposition rate was not significantly affected by the CO2 and warming treatments or by depth in either year (Fig. 3e, f).

Labile pool size (a, b), its initial decomposition rate (c, d) and the decomposition rate of the resistant pool (e, f) in soils collected in June 2007 and July 2008. Values were estimated through non-linear regression of observed respiration rates under incubation conditions (see Methods) and are presented per gram of soil C to account for pre-experimental soil C concentrations. Values are means standard error. ct: ambient CO2 and ambient temperature; cT: ambient CO2 and elevated temperature; Ct: elevated CO2 and ambient temperature; CT: elevated CO2 and elevated temperature. p values of treatment effects from split-plot 3-factor ANOVA (CO2, warming and depth) are shown only when effects were considered significant
The proportion of labile C was positively related to aboveground biomass at both 0–5 cm and 5–15 cm depths and to fine root biomass at 0–5 cm in 2008 (Fig. 4). In contrast, the labile C in soils from 2007 was not related to plant biomass or DOC (regression plots not shown). The initial rate of decomposition of the labile pool was positively related to soil water content at both depths (p = 0.04, r 2 = 0.34 at 0–5 cm and p = 0.03, r 2 = 0.39 at 5–15 cm) in soils from 2007, and at 5–15 cm in 2008 (p = 0.02, r 2 = 0.27) (regression plots not shown). Significant relationships between the resistant pool decomposition rate and the indicators of plant input were observed in both years: the resistant pool decomposition rate was positively related to aboveground plant biomass, root biomass and DOC in 2007 and with DOC in 2008 (Fig. 5).

Linear relationships between the estimated pool of labile C at 0–5 cm and 5–15 cm soils (as mg C g soil C−1) and field measured above-ground biomass and fine root biomass for soils collected close to the time of aboveground peak biomass in 2008. ct: ambient CO2 and ambient temperature; cT: ambient CO2 and elevated temperature; Ct: elevated CO2 and ambient temperature; CT: elevated CO2 and elevated temperature

Linear relationships between the estimated rate of decomposition of the resistant C pool from 0–5 and 5–15 cm soils (as mg C g soil C−1 day−1) and field measured aboveground biomass, root biomass and dissolved organic C (DOC) for soils collected close to the time of aboveground peak biomass in 2008. ct: ambient CO2 and ambient temperature; cT: ambient CO2 and elevated temperature; Ct: elevated CO2 and ambient temperature; CT: elevated CO2 and elevated temperature
Discussion
We evaluated the responses to eCO2 and warming of SOM pools and their susceptibility to decomposition in a semi-arid grassland and whether these responses were mediated by plant inputs. We used aboveground biomass (in this system, equivalent to aboveground net primary productivity) and fine root biomass as measures of plant inputs to soil. Our results demonstrated that positive aboveground productivity responses to eCO2 and warming treatments translated into greater belowground biomass, which in turn was associated with an enlarged pool of soil DOC (Figs. 1 and 2). An important source of DOC is root exudation (Högberg and Högberg 2002; Giesler et al. 2007), which is, in turn, proportional to root biomass (Jones et al. 2009) and fixed C (Pinton et al. 2001). Together, these results indicate that aboveground biomass, fine root biomass and DOC could be used as indicators of plant C inputs to soil.
The marked treatment effectson the labile C observed in both years demonstrated the sensitive nature of this pool to CO2 and warming. Contrasting effects across years indicate the dynamic nature of the factors involved in the regulation of this pool. Our results suggest that plant input was more important in determining the size of the labile pool in 2008 while treatment effects on soil moisture and temperature and their impacts on decomposition dominated the effects in 2007. The greater importance of plant inputs in 2008 is supported by positive relationships between the labile C pool size and plant biomass (Fig. 4). A greater role of plant inputs in controlling the labile C pool in 2008 than in 2007 is also consistent with the positive effect of eCO2 on aboveground biomass observed in 2008 (Table 2). A significant increase in aboveground biomass is expected to increase the input of recent photosynthate to soil which apparently enlarged the labile C pool in 2008. In contrast, in 2007, labile C was not related to plant biomass variables and there were no strong effects of treatments on plant biomass. Observed interactive effects of warming and CO2 on the labile C pool in 2007 are, instead, consistent with the effects of eCO2 and warming on soil moisture and temperature and with their effects on decomposition rates. The higher content of labile C in ambient CO2 with warming (Fig. 3a; cT) in 2007 could be explained by decreased labile C decomposition by microbes due to soil drying (Liu et al. 2009). In eCO2 with warming (Fig. 3a; CT), the effect of drying on the labile pool was likely offset by greater soil moisture due to higher plant water use efficiency under eCO2 (Table 1) (Morgan et al. 2004b), thus allowing enhanced decomposition of the labile C with increased temperature (Kirschbaum 2006).
When considering all treatments in each year, the decomposition rate of the resistant C pool was positively related to plant (above and belowground) biomass in 2007 and to the concentration of DOC in soil in 2007 and 2008 (Fig. 5). Although in 2008 DOC was the only variable with detectable relationships with the resistant pool, both root and aboveground biomass were positively related to DOC (Fig. 2) and labile C in this year (Fig. 4). The positive correlations between plant biomass variables with DOC and labile C suggest that enhanced plant inputs increased the concentration of readily available C in soil. Together, these observations suggest that, at least under incubation conditions, the susceptibility to decomposition of the resistant pool was greater in soils that had greater concentrations of plant derived inputs and thus greater concentrations of available C. Increased available C in soil has been linked to enhanced decomposition of SOM via the priming effect (Fontaine et al. 2007; Kuzyakov 2010). Thus, the positive relationships between the decomposition rate of the resistant pool with plant biomass and DOC that we observed suggest that greater concentrations of readily available C in soil could have made the resistant C pool more susceptible to decomposition, in other words, that the resistant pool in these soils has the potential to be primed by increased labile C under field conditions.
A disadvantage of incubation conditions is that soil C pools are decoupled from the root system. Thus it is not possible to directly measure stimulation of SOM decomposition resulting from root C inputs, or ‘rhizosphere priming’ (Kuzyakov 2002). However, by measuring aboveground and root biomass as well as DOC at the time of soil collection and immediately setting up controlled incubations of freshly collected soils to determine labile C pools we were able to link field treatment impacts on plant inputs with C availability in soil. Stimulation/suppression of SOM decomposition is often assessed by measuring total CO2 fluxes and not directly by assessing decomposition rates of individual pools and thus it is difficult to determine whether the extra C respired is derived from microbial turnover, labile soil C or stable OM (Blagodatskaya and Kuzyakov 2008). However, a change in the dynamics of long-lived, resistant SOM pools has the highest potential to impact long term C storage. With an incubation approach, we were able to estimate the rate of decomposition of the resistant pool, in particular, as opposed to bulk SOM, under the range of labile C availability generated by the experimental treatments.
eCO2 has been hypothesized to cause SOM depletion via the priming effect because it can promote plant production and thus plant C input to soil (Carney et al. 2007). Studies have found evidence of accelerated decomposition of older SOM under eCO2 (Pendall et al. 2003) as well as of pools of total and resistant SOM under eCO2 (Langley et al. 2009; Hoosbeek et al. 2004), which have been attributed to priming. Other climate factors, however, might contribute to C depletion by altering the availability of labile C. For example, changes in moisture availability due to either warming or CO2 could contribute to changes in labile C availability and thus impact the extent of SOM priming (Dijkstra and Cheng 2007b). Our observations under incubation conditions suggest that the stable SOM in soils from the mixed-grass prairie is sensitive to plant C inputs, which in turn were related to available C in soil. Based on the positive relationships observed between indicators of plant inputs and the decomposition rate of the resistant C at PHACE, we speculate that if increased C availability were to occur and persist over multiple growing seasons it could lead to a gradual depletion of the resistant C pool. Using the DayCent model, Parton et al. (2007) predicted that despite increased plant inputs, eCO2 alone and in combination with warming will gradually decrease total soil organic C at PHACE, due to enhanced decomposition with greater soil moisture (a mechanism that can be accounted for by the DayCent model). Changes in the dynamics of long-lived, resistant SOM pools have the highest potential to impact long term C storage. Our results suggest an additional mechanism by which soil C, the resistant pool in particular, could be depleted in this system, highlighting the need to include the influence of labile C on the dynamics of long-lived, resistant SOM pool, in the modeling of long term responses of soil pools to climate change (Heimann and Reichstein 2008; Chapin et al. 2009).
Changes in bulk soil C pools are usually detected only after long exposures to global change treatments. By assessing individual C pools, our study demonstrated that SOM pools in the semi-arid mixed grass prairie can have early responses to eCO2 and warming and that different pools can show contrasting effects. Our results demonstrated that the labile C pool is highly responsive to eCO2 and warming but that the main factors driving the responses of this pool are highly dynamic. Plant inputs were more important in controlling the size of the labile pool in the year when aboveground biomass responded to eCO2, which highlighted the strong link between plant biomass responses to climate factors and labile C availability in soil. Our assessment of the decomposition rate of the resistant C pool suggested that the stable SOM in soils from the mixed-grass prairie is sensitive to C availability and particularly that it could be negatively affected by increases in readily available C driven by global change impacts on plant productivity. An in situ evaluation of the response of resistant SOM under field conditions is thus warranted.
References
Allard V, Newton PCD, Lieffering M, Soussana JF, Carran RA, Matthew C (2005) Increased quantity and quality of coarse soil organic matter fraction at elevated CO2 in a grazed grassland are a consequence of enhanced root growth rate and turnover. Plant Soil 276(1–2):49–60
Belay-Tedla A, Zhou XH, Su B, Wan SQ, Luo YQ (2009) Labile, recalcitrant, and microbial carbon and nitrogen pools of a tallgrass prairie soil in the us great plains subjected to experimental warming and clipping. Soil Biol Biochem 41(1):110–116
Blagodatskaya E, Kuzyakov Y (2008) Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review. Biol Fertil Soils 45(2):115–131
Cardon ZG, Hungate BA, Cambardella CA, Chapin FS, Field CB, Holland EA, Mooney HA (2001) Contrasting effects of elevated CO2 on old and new soil carbon pools. Soil Biol Biochem 33(3):365–373
Carney KM, Hungate BA, Drake BG, Megonigal JP (2007) Altered soil microbial community at elevated CO2 leads to loss of soil carbon. Proc Natl Acad Sci USA 104(12):4990–4995
Chapin FS, McFarland J, McGuire AD, Euskirchen ES, Ruess RW, Kielland K (2009) The changing global carbon cycle: linking plant-soil carbon dynamics to global consequences. J Ecol 97(5):840–850
Cheng L, Leavitt SW, Kimball BA, Pinter PJ, Ottmane MJ, Matthias A, Wall GW, Brooks T, Williams DG, Thompson TL (2007) Dynamics of labile and recalcitrant soil carbon pools in a sorghum free-air CO2 enrichment (FACE) agroecosystem. Soil Biol Biochem 39(9):2250–2263
Collins HP, Elliott ET, Paustian K, Bundy LC, Dick WA, Huggins DR, Smucker AJM, Paul EA (2000) Soil carbon pools and fluxes in long-term corn belt agroecosystems. Soil Biol Biochem 32(2):157–168
Crow SE, Lajtha K, Filley TR, Swanston CW, Bowden RD, Caldwell BA (2009) Sources of plant-derived carbon and stability of organic matter in soil: implications for global change. Glob Chang Biol 15(8):2003–2019
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440(7081):165–173
Del Galdo I, Oechel WC, Cotrufo MF (2006) Effects of past, present and future atmospheric CO2 concentrations on soil organic matter dynamics in a chaparral ecosystem. Soil Biol Biochem 38(11):3235–3244
de Graaff MA, van Groenigen KJ, Six J, Hungate B, van Kessel C (2006) Interactions between plant growth and soil nutrient cycling under elevated CO2: a meta-analysis. Glob Chang Biol 12(11):2077–2091
de Graaff MA, Van Kessel C, Six J (2009) Rhizodeposition-induced decomposition increases N availability to wild and cultivated wheat genotypes under elevated CO2. Soil Biol Biochem 41(6):1094–1103
Dijkstra FA, Cheng WX (2007a) Interactions between soil and tree roots accelerate long-term soil carbon decomposition. Ecol Lett 10(11):1046–1053
Dijkstra FA, Cheng WX (2007b) Moisture modulates rhizosphere effects on C decomposition in two different soil types. Soil Biol Biochem 39(9):2264–2274
Dijkstra FA, Cheng WX, Johnson DW (2006) Plant biomass influences rhizosphere priming effects on soil organic matter decomposition in two differently managed soils. Soil Biol Biochem 38(9):2519–2526
Fontaine S, Barot S, Barre P, Bdioui N, Mary B, Rumpel C (2007) Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 450(7167):277–U210
Fu SL, Cheng WX, Susfalk R (2002) Rhizosphere respiration varies with plant species and phenology: a greenhouse pot experiment. Plant Soil 239(1):133–140
Garten CT, Classen AT, Norby RJ (2009) Soil moisture surpasses elevated CO2 and temperature as a control on soil carbon dynamics in a multi-factor climate change experiment. Plant Soil 319(1–2):85–94
Giesler R, Hogberg MN, Strobel BW, Richter A, Nordgren A, Hogberg P (2007) Production of dissolved organic carbon and low-molecular weight organic acids in soil solution driven by recent tree photosynthate. Biogeochemistry 84(1):1–12
Gill RA, Polley HW, Johnson HB, Anderson LJ, Maherali H, Jackson RB (2002) Nonlinear grassland responses to past and future atmospheric CO2. Nature 417(6886):279–282
Heimann M, Reichstein M (2008) Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 451(7176):289–292
Högberg MN, Högberg P (2002) Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytol 154(3):791–795
Hoosbeek MR, Lukac M, van Dam D, Godbold DL, Velthorst EJ, Biondi FA, Peressotti A, Cotrufo MF, de Angelis P, Scarascia-Mugnozza G (2004) More new carbon in the mineral soil of a poplar plantation under free air carbon enrichment (POPFACE): cause of increased priming effect? Glob Biogeochem Cycles 18(1)
Hungate BA, van Groenigen KJ, Six J, Jastrow JD, Lue YQ, de Graaff MA, van Kessel C, Osenberg CW (2009) Assessing the effect of elevated carbon dioxide on soil carbon: A comparison of four meta-analyses. Glob Chang Biol 15(8):2020–2034
Hutsch BW, Augustin J, Merbach W (2002) Plant rhizodeposition—an important source for carbon turnover in soils. J Plant Nutr Soil Sci-Zeitschrift Fur Pflanzenernahrung Und Bodenkunde 165(4):397–407
Jastrow JD, Miller RM, Owensby CE (2000) Long-term effects of elevated atmospheric CO2 on below-ground biomass and transformations to soil organic matter in grassland. Plant Soil 224(1):85–97
Jastrow JD, Miller RM, Matamala R, Norby RJ, Boutton TW, Rice CW, Owensby CE (2005) Elevated atmospheric carbon dioxide increases soil carbon. Glob Chang Biol 11(12):2057–2064
Jones DL, Nguyen C, Finlay RD (2009) Carbon flow in the rhizosphere: carbon trading at the soil-root interface. Plant Soil 321(1–2):5–33
Kimball BA, Conley MM, Wang S, Lin X, Luo C, Morgan J, Smith D (2008) Infrared heater arrays for warming ecosystem field plots. Glob Chang Biol 14(2):309–320
Kirschbaum MUF (2006) The temperature dependence of organic-matter decomposition—still a topic of debate. Soil Biol Biochem 38(9):2510–2518
Kuzyakov Y (2002) Review: factors affecting rhizosphere priming effects. J Plant Nutr Soil Sci-Zeitschrift Fur Pflanzenernahrung Und Bodenkunde 165(4):382–396
Kuzyakov Y (2010) Priming effects: interactions between living and dead organic matter. Soil Biol Biochem 42(9):1363–1371
Langley JA, McKinley DC, Wolf AA, Hungate BA, Drake BG, Megonigal JP (2009) Priming depletes soil carbon and releases nitrogen in a scrub-oak ecosystem exposed to elevated CO2. Soil Biol Biochem 41(1):54–60
Liu WX, Zhang Z, Wan SQ (2009) Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob Chang Biol 15(1):184–195
Luo YQ, Hui DF, Zhang DQ (2006) Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: a meta-analysis. Ecology 87(1):53–63
Luo YQ, Gerten D, Le Maire G, Parton WJ, Weng ES, Zhou XH, Keough C, Beier C, Ciais P, Cramer W, Dukes JS, Emmett B, Hanson PJ, Knapp A, Linder S, Nepstad D, Rustad L (2008) Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones. Glob Chang Biol 14(9):1986–1999
Miglietta F, Hoosbeek MR, Foot J, Gigon F, Hassinen A, Heijmans M, Peressotti A, Saarinen T, van Breemen N, Wallen B (2001) Spatial and temporal performance of the miniface (free air CO2 enrichment) system on bog ecosystems in northern and central europe. Environ Monit Assess 66(2):107–127
Milchunas DG, Morgan JA, Mosier AR, LeCain DR (2005) Root dynamics and demography in shortgrass steppe under elevated CO2, and comments on minirhizotron methodology. Glob Chang Biol 11(10):1837–1855
Morgan JA, Mosier AR, Milchunas DG, LeCain DR, Nelson JA, Parton WJ (2004a) CO2 enhances productivity, alters species composition, and reduces digestibility of shortgrass steppe vegetation. Ecol Appl 14(1):208–219
Morgan JA, Pataki DE, Korner C, Clark H, Del Grosso SJ, Grunzweig JM, Knapp AK, Mosier AR, Newton PCD, Niklaus PA, Nippert JB, Nowak RS, Parton WJ, Polley HW, Shaw MR (2004b) Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 140(1):11–25
Norby RJ, Luo YQ (2004) Evaluating ecosystem responses to rising atmospheric CO2 and global warming in a multi-factor world. New Phytol 162(2):281–293
Norby RJ, Ledford J, Reilly CD, Miller NE, O’Neill EG (2004) Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. Proc Natl Acad Sci USA 101(26):9689–9693
Parton WJ, Morgan JA, Wang GM, Del Grosso S (2007) Projected ecosystem impact of the prairie heating and CO2 enrichment experiment. New Phytol 174(4):823–834
Pendall E, King JY (2007) Soil organic matter dynamics in grassland soils under elevated CO2: insights from long-term incubations and stable isotopes. Soil Biol Biochem 39(10):2628–2639
Pendall E, Del Grosso S, King JY, LeCain DR, Milchunas DG, Morgan JA, Mosier AR, Ojima DS, Parton WA, Tans PP, White JWC (2003) Elevated atmospheric CO2 effects and soil water feedbacks on soil respiration components in a colorado grassland. Glob Biogeochem Cycles 17(2)
Pendall E, Bridgham S, Hanson PJ, Hungate B, Kicklighter DW, Johnson DW, Law BE, Luo YQ, Megonigal JP, Olsrud M, Ryan MG, Wan SQ (2004) Below-ground process responses to elevated CO2 and temperature: a discussion of observations, measurement methods, and models. New Phytol 162(2):311–322
Pepper DA, Del Grosso SJ, McMurtrie RE, Parton WJ (2005) Simulated carbon sink response of shortgrass steppe, tallgrass prairie and forest ecosystems to rising [CO2], temperature and nitrogen input. Glob Biogeochem Cycles 19(1)
Pinton R, Varanini Z, Nannipieri P (2001) The rhizosphere: biochemistry and organic substances at the soil-plant interface. Marcel Dekker, New York
Rustad LE, Campbell JL, Marion GM, Norby RJ, Mitchell MJ, Hartley AE, Cornelissen JHC, Gurevitch J (2001) A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126(4):543–562
Shen WJ, Reynolds JF, Hui DF (2009) Responses of dryland soil respiration and soil carbon pool size to abrupt vs. gradual and individual vs. combined changes in soil temperature, precipitation, and atmospheric [CO2]: a simulation analysis. Glob Chang Biol 15(9):2274–2294
Sherrod SK, Belnap J, Miller ME (2002) Comparison of methods for nutrient measurement in calcareous soils: ion-exchange resin bag, capsule, membrane, and chemical extractions. Soil Sci 167(10):666–679
Shim JH, Pendall E, Morgan JA, Ojima DS (2009) Wetting and drying cycles drive variations in the stable carbon isotope ratio of respired carbon dioxide in semi-arid grassland. Oecologia 160(2):321–333
Taneva L, Gonzalez-Meler MA (2008) Decomposition kinetics of soil carbon of different age from a forest exposed to 8 years of elevated atmospheric CO2 concentration. Soil Biol Biochem 40(10):2670–2677
Townsend AR, Vitousek PM, Desmarais DJ, Tharpe A (1997) Soil carbon pool structure and temperature sensitivity inferred using CO2 and (13) CO2 incubation fluxes from five Hawaiian soils. Biogeochemistry 38(1):1–17
Wan SQ, Norby RJ, Pregitzer KS, Ledford J, O’Neill EG (2004) CO2 enrichment and warming of the atmosphere enhance both productivity and mortality of maple tree fine roots. New Phytol 162(2):437–446
Wan SQ, Hui DF, Wallace L, Luo YQ (2005) Direct and indirect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie. Glob Biogeochem Cycles 19(2)
Wedin DA, Pastor J (1993) Nitrogen mineralization dynamics in grass monocultures. Oecologia 96(2):186–192
Weintraub MN, Scott-Denton LE, Schmidt SK, Monson RK (2007) The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem. Oecologia 154(2):327–338
Acknowledgements
We thank Dan LeCain and David Smith for technical support with the field experiment and the lab of Ronald F. Follett for the soil analyses. We thank Sarah Berg, Hannah Rae Munn, Christine Rumsey, Matthew Wood and Megan Steinweg for assistance in the field and in the lab and Jennifer King for insightful review of the manuscript. This project was supported by USDA-CSREES Soil Processes Program (Grant no. 2008-35107-18655), by the US Department of Energy’s Office of Science (BER) through the Western Regional Center of the National Institute for Climatic Change Research at Northern Arizona University, NSF (DEB# 1021559) and the USDA-Agricultural Research Service.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Johan Six.
Rights and permissions
About this article
Cite this article
Carrillo, Y., Pendall, E., Dijkstra, F.A. et al. Response of soil organic matter pools to elevated CO2 and warming in a semi-arid grassland. Plant Soil 347, 339–350 (2011). https://doi.org/10.1007/s11104-011-0853-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-011-0853-4