
Abstract
Elevated CO2 affects plant productivity, but also water availability and plant species composition in semi-arid grasslands, thereby potentially causing complex effects on CH4 consumption and N2O emission. We studied the effects of atmospheric CO2 concentration (400 vs 780 μL L−1), water content (15 vs 20% gravimetric soil moisture), and composition of semi-arid grassland species (perennial grasses Bouteloua gracilis, Hesperostipa comata, and Pascopyrum smithii; sub-shrub Artemisia frigida; invasive forb Linaria dalmatica grown in monoculture and all five species together) on CH4 consumption and N2O emission in a full factorial greenhouse experiment. We used a unique method where we measured microbial effects on CH4 consumption and N2O emission in isolation from effects of gas diffusivity. Microbially mediated CH4 consumption was significantly higher under elevated CO2 (by 20%), but was not affected by soil water content or plant species composition. Microbially mediated N2O emission was not significantly affected by elevated CO2, but was significantly higher with high water content (by 67%) and differed significantly among species. Treatment effects on CH4 consumption and N2O emission often could not be explained simply by differences in soil moisture, suggesting that treatment-induced changes in other soil and microbial properties played a role in causing these effects.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Earth’s atmospheric CO2 concentration has risen from ~280 µL L−1 at the start of the industrial revolution to greater than 385 µL L−1 today, and is expected to exceed 700 µL L−1 by the end of this century (Intergovernmental Panel on Climate Change 2007). This rise in atmospheric CO2 has strong direct and indirect effects on ecosystems. Semi-arid grasslands are considered to be highly responsive to rising CO2. Biological activity in these grasslands is strongly modulated by the natural seasonal variation in soil water availability (Frank and Groffman 1998; Huxman et al. 2004; Potts et al. 2006). Elevated CO2-induced increases in plant water use efficiency due to increased plant stomatal closure (Morgan et al. 2004b) would be expected to enhance that biological activity through increases in soil water content. Indeed, CO2-induced changes in soil water availability may have played an important role in the observed increases in plant productivity, shifts in plant species composition, and altered C and N cycling in semi-arid grasslands exposed to enriched CO2 concentrations (Dijkstra et al. 2008; Morgan et al. 2004b, 2007; Pendall et al. 2003).
A major concern of a global increase in atmospheric CO2 is how it will affect flux rates of other greenhouse gases such as methane (CH4) and nitrous oxide (N2O). Methane and N2O have a 25 and 298 times greater relative global warming potential than CO2 over 100 years (Intergovernmental Panel on Climate Change 2007). Semi-arid grasslands represent a significant global sink for CH4 and source for N2O (Galbally et al. 2008; Mosier et al. 1991, 1997). The long-term effects of elevated CO2 on CH4 consumption and N2O emission in these grasslands are largely unknown (Galbally et al. 2008), in part because of complicated effects of CO2 on soil water, C and N availability, as well as plant productivity and species composition.
Changes in atmospheric CO2 concentration can affect CH4 consumption and N2O emission in semi-arid grasslands through changes in soil water availability. Methane consumption often decreases in response to elevated CO2, which has been related to increased soil moisture (Ambus and Robertson 1999; Ineson et al. 1998; McLain and Ahmann 2008; McLain et al. 2002), but no change in CH4 consumption has also been observed (Kang et al. 2001; Kettunen et al. 2005; Mosier et al. 2002). Although a certain amount of soil water is required for methanotrophs to oxidize CH4, influxes of CH4 into soils may be impeded once soil water content exceeds threshold values beyond which diffusivity of CH4 becomes more limiting (Del Grosso et al. 2000; Koschorreck and Conrad 1993). On the other hand, increases in soil water content have been implicated in CO2-related increases in N2O emission (Arnone and Bohlen 1998; Kanerva et al. 2007; Kettunen et al. 2006; Robinson and Conroy 1999). Elevated CO2 may also elicit changes in CH4 and N2O exchange unrelated to soil moisture. Elevated CO2 could decrease CH4 consumption by transforming the microbial communities that are responsible for CH4 production and consumption (McLain and Ahmann 2008; Phillips et al. 2001a), while this decrease in CH4 consumption may depend on elevated CO2-induced changes in gross nitrification (Baggs and Blum 2004). Others have suggested that greater N2O emissions under elevated CO2 are a consequence of enhanced rhizodeposition, acting as an energy source for denitrification (Baggs et al. 2003a, b; Ineson et al. 1998; Kammann et al. 2008). But often N2O emissions do not respond to elevated CO2 (Ambus and Robertson 1999; Billings et al. 2002; Hungate et al. 1997a; Mosier et al. 2002). Mosier et al. (2002) suggested that a combination of greater N supply (due to increased N mineralization with increased soil moisture) and increased plant N demand under elevated CO2 maintained mineral N pools in the soil too low to alter rates of N2O emissions in a semi-arid grassland.
Elevated CO2 could also potentially affect CH4 consumption and N2O emission in semi-arid grasslands through changes in plant species composition. Elevated CO2 caused greater abundances of the C3 grass Hesperostipa comata and the sub-shrub Artemisia frigida in a semi-arid grassland in Colorado (Morgan et al. 2004a, 2007). However, little is known about how such changes in species composition will affect CH4 and N2O fluxes. Epstein et al. (1998) reported greater CH4 consumption in patches dominated by C4 grasses (mostly Bouteloua gracilis) than in patches dominated by C3 grasses (mostly Pascopyrum smithii) in a semi-arid grassland. Further, patches mixed with C3 and C4 grasses had lower N2O emissions than either the C4 or C3 dominated patches. Niklaus et al. (2006) also reported lower N2O emissions with increased diversity of temperate grassland species. In a sagebrush-steppe Norton et al. (2008) reported greater N2O emissions under the exotic grass Bromus tectorum than under the native grass P. smithii. Plant species effects on CH4 consumption and N2O emission have been associated with plant species effects on microbial community composition (Menyailo and Hungate 2003; Ullah et al. 2008), N cycling and soil NO −3 concentration (Crenshaw et al. 2008; Epstein et al. 1998; Norton et al. 2008), and on soil C:N ratios (Ambus et al. 2006; Menyailo and Huwe 1999).
Responses of CH4 consumption and N2O emission are often complex because microbial and physical factors (gas diffusivity) controlling these fluxes operate at different scales in time and space. We set up a controlled greenhouse experiment to better understand microbially mediated CH4 consumption and N2O emission. In particular, we studied how elevated CO2, soil water content, species composition of semi-arid grasslands, and their interactions affected microbially mediated CH4 consumption and N2O emission in isolation from physical processes (i.e., gas diffusivity). In two greenhouses we grew five grassland species in monoculture and all five species in competition under ambient and elevated CO2 (400 vs. 780 μL L−1), and under ‘low’ and ‘high’ soil moisture conditions (15 vs 20% gravimetric soil moisture). We used the native perennial grasses Bouteloua gracilis, Hesperostipa comata, and Pascopyrum smithii, the sub-shrub Artemisia frigida, and the invasive forb Linaria dalmatica, all common to the semi-arid grasslands of Colorado. We used a novel method to measure CH4 consumption and N2O emission in which we eliminated potential effects of gas diffusivity. In this way we were able to test the direct effects of the treatments on microbially mediated CH4 consumption and N2O production. We addressed the following questions: 1) how do elevated CO2, soil water content, and species composition affect microbially mediated CH4 consumption and N2O emission, and 2) to what extent can these effects be explained by soil moisture? Besides soil moisture, we also tested if parameters of plant activity (pot respiration, plant biomass and its N content) or inorganic soil N could explain the variation in CH4 consumption and N2O emission among treatments.
Materials and methods
Experimental design
The soil used in this experiment came from the USDA-ARS Central Plains Experimental Range in the shortgrass steppe region of north-eastern Colorado. The soil was a sandy loam of the Ascalon series (Aridic Argiustolls). The soil collected to 20-cm depth was carbonate-free, had a pH of 6.6 and 0.95% total C and 0.09% total N. The soil was homogenized by sieving (4 mm) and air-dried before use. We filled each of 96 polyvinyl chloride (PVC) pots (diam. 20 cm, height 40 cm, closed at the bottom except for an air inlet) with 14 kg of air-dried soil (Fig. 1). Before filling the pots we placed a nylon bag filled with 3 kg playground sand at the bottom of each pot to improve air circulation. Exact weights of soil and filled pots were recorded. We watered the pots to field capacity (i.e., 30% gravimetric soil moisture content). In each pot we put five plants of Artemisia frigida (sub-shrub), Linaria dalmatica (forb), Bouteloua gracilis (C4 grass), Hesperostipa comata (C3 grass), or Pascopyrum smithii (C3 grass), 16 pots for each species. All species are native to the shortgrass steppe, except for the invasive weed L. dalmatica. We also planted 16 pots with all five species combined (one plant of each species per pot). Plants were raised from seeds in peat pellets (2–5 seeds in each peat pellet). After plant emergence, plants were thinned to one plant per peat pellet and transferred to the pots.

Setup of the pots during CH4 and N2O flux measurements
We grew the plants in two greenhouses at the USDA-ARS Crops Research Laboratory in Fort Collins, Colorado. In one greenhouse we raised the atmospheric CO2 concentration to a constant level of 780 ± 50 μL L−1 (average ± standard deviation) by adding pure CO2, while in the other greenhouse the atmospheric CO2 concentration was kept near ambient level (~400 ± 40 μL L−1, note that this concentration is slightly higher than the average global CO2 concentration because our greenhouses were in an urban setting). The CO2 concentration was continuously monitored and the CO2 supply was computer-controlled (Argus Control Systems Ltd, White Rock, BCFootnote 1). The added CO2 entered the greenhouse through a ventilation system ensuring uniform distribution of the CO2 concentration inside the greenhouse. Air temperature in both greenhouses was kept between 27 and 29°C during the day and between 16 and 18°C during the night. Temperature was regulated by computer-controlled air conditioners and heaters (York International, York, PA). Both greenhouses were equipped with 600 W lights (P.L. Light Systems, Beamsville, ON) that were on during the day for 12 hrs. The light intensity in each greenhouse was ~200 W m−2 during the day. The relative humidity in each greenhouse was 24 ± 5% during the day and 30 ± 5% during the night. Half of the pots (8 pots for each species and 8 pots with all species combined) were placed in the greenhouse under elevated CO2 (total of 48 pots) and the other half of the pots were placed in the greenhouse under ambient CO2 (total of 48 pots). Once every week, the pots were swapped between the two greenhouses and the CO2 concentration switched accordingly. In this way we tried to minimize greenhouse effects and pseudoreplication at the CO2 level (Goverde and Erhardt 2003; Heijmans et al. 2002).
The pots were watered frequently (see below) to restore gravimetric soil moisture content to 15% (low water) to half of the pots, and to 20% (high water) to the other half of the pots (4 pots/replicates for each CO2 and species composition treatment), or 50 and 67% of field capacity respectively. During the first week, all pots were maintained at 30% soil moisture content, after which pots were allowed to dry down to their soil moisture target contents (15 and 20%). Once target soil moisture contents were reached (26 days after transplanting), pots were watered three times a week. Pots were weighed once every week and watered up to their target weights. The amount of water added during the other two times of the week was calculated based on previous water loss from each pot. In each greenhouse, pots were placed in four blocks, where one replicate of each of the species composition and water treatments were randomly placed in each block (each block consisting of 12 pots).
Measurements and analyses
We measured CH4 consumption and N2O emission 48, 69, and 83 days after transplanting, using a modified method developed for respiration in pots in the presence of plants (Cheng 1996). We also measured respiration at the same time, and used respiration as a covariate to explain variation in CH4 consumption and N2O emission among treatments (see below). On each measurement day we placed chambers (diam. 20 cm, height 45 cm) fitted with a septum and air outlet on top of the pots (Fig. 1). We circulated air inside the pot/chamber by inserting an aquarium pump (Apollo AM-3, Apollo Enterprises, Ventura, CA, flow rate 2.8 L min−1) in-line between the air inlet at the bottom of the pot and the air outlet at the top of the chamber. All pots/chambers were air-circulated for two hours to completely mix the air inside the chamber and pot. During this time CO2 inside the pot/chamber was removed by an in-line CO2 scrubber (PVC tube, diam. 3.5 cm, height 36 cm, filled with sodalime). We then removed the scrubber and pulled a 30 ml gas sample from each chamber (time 0) and another 30 ml gas sample two hours later while air still circulated inside the pot/chamber. Chambers and pumps were removed after the second gas sample was taken. Gas samples were analyzed for CH4, N2O, and CO2 on a gas chromatograph (Varian 3800, Palo Alto, CA). All species remained in a vegetative stage until the end of the experiment, except for B. gracilis and L. dalmatica that started to flower during the last measurement. Under field conditions, flowering of the perennials B. gracilis and L. dalmatica occurs throughout the growing season that does not stop their growth (indeterminate growth). Also in our experiment, B gracilis and L. dalmatica kept growing after flowering.
The purpose of circulating air through the pot/chamber was to establish well-mixed and uniform concentrations of CH4 and N2O inside the pot/chamber thereby eliminating effects of gas diffusivity on CH4 consumption and N2O emission. Thus, the rate of CH4 consumption and N2O emission that we measured indicates the activity of the microbial community. The CH4 consumption (μg C pot−1 hr−1), N2O emission (μg N pot−1 hr−1), and respiration (mg C pot−1 hr−1) were calculated based on the difference between the concentrations of the two samples and the air volume of the pot/chamber system. We assumed that the decrease in CH4 concentration and increase in N2O and CO2 concentration with time did not affect their respective consumption and production rates. For N2O and CO2 production, this is a reasonable assumption, since N2O and CO2 concentrations in the soil tend to be much higher than in the atmosphere. However, for CH4 consumption, this may not be true. The decrease in CH4 concentration inside the pot/chamber may have reduced the rate of CH4 consumption. However, the average CH4 concentrations at 2 and 4 hrs after chambers were placed on the pots were 1.52 and 1.09 μl L−1 respectively. When we assume a CH4 concentration at the time when chambers were placed on the pots of ~1.9 μl L−1 (i.e., ambient CH4 concentration measured in the greenhouse after termination of the experiment), then our assumption of a linear decrease in CH4 concentration with time inside the pot/chamber is reasonable. We also calculated a first order rate constant k to describe the rate of CH4 consumption (\( - k{\text{t}} = \ln \left( {{{{\text{C}}{{\text{H}}_{4,{\text{t}}1}}} \mathord{\left/{\vphantom {{{\text{C}}{{\text{H}}_{4,{\text{t}}1}}} {{\text{C}}{{\text{H}}_{4,{\text{t}}0}}}}} \right.} {{\text{C}}{{\text{H}}_{4,{\text{t}}0}}}}} \right) \), Hütsch et al. 1994), and treatment effects on k were very similar to treatment effects on CH4 consumption calculated with the linear approach (data not shown). We feel confident that treatment effects on our gas flux would not change if we added more data points in time. It would be more accurate to refer to net CH4 consumption and net N2O production, because of simultaneous CH4 production (by methanogens) and consumption (by methanotrophs) and N2O production (through nitrification and denitrification) and consumption (complete denitrification). However, because of the relatively low soil moisture content and high oxygen supply during our measurements we assumed CH4 production and complete denitrification to N2 to be negligible. We did not attempt to characterize the different microbial groups responsible for CH4 and N2O consumption and production. We are aware that our artificial way of measuring CH4 and N2O fluxes in a greenhouse pot study does not allow us to extrapolate our results quantitatively to field situations. However, we want to emphasize that the purpose of this study was to better understand microbially mediated CH4 consumption and N2O emission that are not confounded by gas diffusivity effects.
We harvested all plants after the final CH4 consumption and N2O emission measurement (85 days after transplanting). Plants were separated into aboveground biomass, crowns, and roots. Plant biomass was dried (60°C) and weighed. Plant biomass was analyzed for N concentration on a mass spectrometer (20–20 Stable Isotope Analyzer, Europa Scientific, Chesire, UK). The soil from each pot was thoroughly homogenized before taking a sub-sample for analysis. We added 60 ml of 2 M KCl to 25 g moist soil. Samples were shaken for 1 h and filtered through pre-leached (with 2 M KCl) Whatman No. 1 filter paper. Extracts were frozen until analyses for NH +4 and NO −3 on a flow injection analyzer (QuickChem FIA + , Lachat Instruments, Milwaukee, WI). The NH +4 and NO −3 concentrations were expressed on oven-dry soil weight (105°C) basis.
We used repeated measures ANOVA to test for main effects of CO2 (two levels: ambient and elevated), water (two levels: low and high water), species composition (six levels: five species grown in monoculture and all five species grown in competition), and their interactions on CH4 consumption and N2O emission. We also used repeated measures ANOVA to test for main effects of CO2, water, and species number (two levels: 1 and 5 species), and their interactions on CH4 consumption and N2O emission. The repeated measures ANOVA included random effects of date (48, 69, and 83 days after transplanting) and block. We also used ANOVA to test for main effects of CO2, water, species composition, and their interactions (or for main effects of CO2, water, species number, and their interactions) on CH4 consumption and N2O emission at each date. We then tested for the same effects after adjusting for effects caused by soil moisture (covariate) using ANCOVA. Treatment effects on CH4 consumption and N2O emission could be mediated by treatment effects on plant growth, inorganic N uptake, soil NH +4 and NO −3 , and rhizodeposition. We therefore also used plant biomass, plant biomass N content (an integrated measure of plant N uptake), soil NH +4 and NO −3 measured at the end of the experiment, and respiration (which includes rhizodeposition) as a covariate. We realize that soil NH +4 and NO −3 measured at the end of the experiment may not reflect dynamics of their availability during the experiment. Also, respiration may be a poor indicator of rhizodeposition. Unfortunately, we have no direct measurements of soil NH +4 and NO −3 and rhizodeposition during the experiment. Plant biomass, plant biomass N content, and soil NH +4 and NO −3 were only used as a covariate for the analyses of the last date of CH4 and N2O measurements. We used linear regressions to relate CH4 consumption and N2O emission to soil moisture, respiration, plant biomass, plant biomass N content, or NH +4 and NO −3 . When necessary, data were log transformed to reduce heteroscedasticity. All statistical analyses were done with JMP (version 4.0.4; SAS Institute, Cary, North Carolina, USA).
Results
Between watering events pots under ambient CO2 dried out faster than pots under elevated CO2 (Fig. 2a). Soil moisture contents also diverged among pots with different species composition after each watering event (pots with L. dalmatica drying out fastest and pots with H. comata slowest, Fig. 2b). Because of differences in soil moisture among CO2 and species composition treatments, we used soil moisture content as a covariate to test if treatment effects on CH4 consumption and N2O emission still existed after adjusting for soil moisture effects. We should note that the soil moisture content measured at the time of CH4 and N2O flux measurements may not reflect effects caused by differences in the drying-rewetting cycles that occurred throughout the experiment.

Gravimetric soil moisture concentration during the experiment for a the CO2 (aCO2: ambient CO2, eCO2: elevated CO2) and water treatments (low water: 15% soil moisture, high water: 20% soil moisture), averaged across species composition treatment and b the species composition treatment averaged across the CO2 and water treatments
We observed CH4 consumption in all pots at all three dates. Repeated measures ANOVA on all three dates showed that elevated CO2 significantly increased the CH4 consumption (on average by 20%), while there were no significant water and species composition effects (Table 1, Fig. 3). There were also no significant treatment interactions on CH4 consumption. The increase in CH4 consumption under elevated CO2 was not significant during the first date when plants were still small, but became significant during the last two dates of measurement (ANOVA, Table 1, Fig. 3). After adjusting for soil moisture effects, CH4 consumption was still higher under elevated CO2 during the last two dates of measurement (ANCOVA, Table 1). Surprisingly, we observed no significant relationship between the CH4 consumption and soil moisture content for each date or when all three dates were combined (P > 0.1, Fig. 4a). Methane consumption 83 days after transplanting was not significantly related to plant biomass, plant biomass N, or NH +4 and NO −3 concentrations in the soil (P > 0.1) and adjusting for plant biomass, plant biomass N, or NH +4 and NO −3 concentrations in the soil did not change treatment effects (data not shown). Methane consumption was significantly positively related to pot respiration, although the variability explained was small (P < 0.0001, R 2 = 0.18). When respiration was used as a covariate in the ANCOVA it did not change treatment effects (data not shown).

Methane consumption on 48, 69, and 83 days after planting affected by CO2 (aCO2: ambient CO2, eCO2: elevated CO2), averaged across water treatment and species composition treatment

a Methane consumption and b N2O emission 48, 69, and 83 days after transplanting as a function of soil moisture
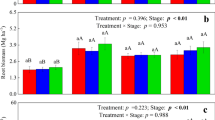
The emission of N2O at all three dates was not significantly affected by elevated CO2, but was significantly higher in the high water treatment (on average by 67%) and differed significantly among the six levels of the species composition treatment (repeated measures ANOVA, Table 1, Fig. 5). The N2O emission was highest under H. comata, particularly in the high water treatment (significant water*species composition interaction). The ANOVA results for each date show that water treatment effects became less significant by the end of the experiment, but that species composition effects remained significant throughout the experiment (mostly due to H. comata). When we used soil moisture content as a covariate, the water treatment effect disappeared 48 and 83 days after transplanting and was only marginally significant 69 days after transplanting. However, species composition effects remained significant for all dates and the water*species composition interaction remained the same after correcting for soil moisture effects (ANCOVA, Table 1). Not surprisingly, the N2O emission was significantly related to soil moisture content, although the variability explained by soil moisture was small (P < 0.0001, R 2 = 0.15 Fig. 4b). The N2O emission was not significantly related to pot respiration, and using pot respiration as a covariate in the ANCOVA did not change any of the treatment effects (data not shown). The N2O emission 83 days after transplanting was not related to plant biomass, or plant biomass N, but was significantly related to NO −3 concentrations in the soil (P < 0.0001, data not shown). The main water effect disappeared, and the species composition effect was less significant after adjusting for NO −3 concentrations in the soil (Table 1).

Nitrous oxide emission on 48, 69, and 83 days after transplanting affected by soil water content (low water: 15% soil moisture, high water: 20% soil moisture) and species composition averaged across the CO2 treatment
We observed no significant species number (1 vs. 5 species) effects on CH4 consumption (P = 0.12, repeated measures ANOVA, data not shown). We did observe a significant reduction in N2O emission with 5 species compared to 1 species (P = 0.04, repeated measures ANOVA), particularly under high water (species number*water interaction, P = 0.05). However, the species number effect and species number*water interaction disappeared during the last measurement, when plants were largest.
Discussion
Methane consumption significantly increased with elevated CO2, but was not affected by soil water content or species composition, while N2O emission was not affected by elevated CO2, but significantly increased with increased water content and showed a significant species composition effect. The CO2 effect on CH4 consumption and the species composition effect on N2O emission could not be explained by CO2 or species composition effects on soil moisture.
We utilized a new method to measure microbially mediated CH4 consumption and N2O emission by eliminating effects of gas diffusivity. For CH4, gas diffusivity may impede the influx of CH4 into the soil when soil water content reaches a certain threshold (Del Grosso et al. 2000; Koschorreck and Conrad 1993). Mosier et al. (2008) reported that this threshold was around 10–20% water-filled pore space, or around 6.2–12% gravimetric soil moisture content (using a soil bulk density of 1 g cm−3 and a solid soil density of 2.65 g cm−3), for the USDA-ARS Central Plains Experimental Range where we collected our soil for this experiment. This threshold is at the lower end of soil moisture contents we maintained in our experiment. Thus gas diffusivity most likely limited CH4 consumption in our pots during much of the time we did not circulate air through the pots, although diffusivity may have been higher in our pots than in the field due to gas exchange between the pot wall and the soil. Although we may not have completely eliminated gas diffusivity, we are confident that, during our measurements when we circulated air through the pots, we increased this threshold so that gas diffusivity was not the limiting factor for CH4 consumption (and N2O emission) anymore. If gas diffusivity played a significant role in CH4 consumption during our measurements, we would have expected lower CH4 consumption in the wetter soils under elevated CO2, but we observed the opposite. We also did not observe a negative relationship between CH4 consumption and soil moisture.
The activity of methanotrophs to oxidize CH4 is limited by soil moisture content (Del Grosso et al. 2000; Stein and Hettiaratchi 2001). The increase in soil moisture under elevated CO2 may have contributed to greater CH4 consumption in pots under elevated CO2 compared to pots with the same water treatment under ambient CO2 (Fig. 2). However, the microbially mediated CH4 consumption was not solely affected by soil moisture content, since CH4 consumption under elevated CO2 remained significantly higher after correcting for soil moisture. We also observed no relationship between CH4 consumption and soil moisture. These results suggest that the sensitivity of methanotroph activity to water content is not straightforward and that factors other than soil moisture contributed to the greater methanotroph activity under elevated CO2.
High concentrations of NH +4 and NO −3 in the soil often inhibit methanotroph activity (Bédard and Knowles 1989), although increased CH4 oxidation with NH +4 fertilization in rice paddies has also been observed (Bodelier et al. 2000). Elevated CO2 did not affect soil NO −3 concentration (ANOVA, P = 0.95) but increased the NH +4 concentration by 25% (ANOVA, P = 0.02; on average NH +4 concentrations were 0.32 and 0.40 mg N kg−1 soil for ambient and elevated CO2 respectively). Although the increase in NH +4 concentration under elevated CO2 could potentially have reduced N limitation of methanotroph activity (Bodelier et al. 2000), the NH +4 (or NO −3 ) concentration in the soil was not related to CH4 consumption.
It is possible that increased labile substrates under elevated CO2 may have stimulated methanotrophs to consume CH4. It is well known that elevated CO2 can increase the input of labile C substrates into the soil (Cheng and Johnson 1998; Dijkstra et al. 2005; Hungate et al. 1997b). Evidence that methanotrophs are limited by C substrates is less clear. Conrad (1984) suggested that methanotrophs are able to use organic matter as a carbon source, and indeed several studies have suggested increased CH4 consumption with greater availability of labile C substrates (Benstead et al. 1998; Goldman et al. 1995; Jacinthe and Lal 2005). However, elevated CO2 reduced CH4 consumption in a loblolly pine plantation that could not be explained by potential increases in labile C substrates (Phillips et al. 2001b). We did not measure availability of labile C substrates in our experiment, but found a significant positive relationship between CH4 consumption and respiration. Nevertheless, respiration could not explain the CO2 treatment effect on CH4 consumption, possibly because our respiration measurements may not have been a strong indicator of labile C substrates into the soil. A potentially greater availability of labile C substrates (as well as a greater soil moisture content) under elevated CO2 may also have increased methanogen activity (i.e., production of CH4). Although methanogen activity can occur in aerobic conditions (Khalil and Baggs 2005), soil moisture content in our experiment was most likely too small to cause a significant effect on CH4 production (Von Fischer and Hedin 2007), and increased methanogen activity would have countered the CO2 treatment effect on net CH4 consumption.
In field studies a decrease in CH4 consumption in response to elevated CO2 has been associated with greater soil moisture content (Ambus and Robertson 1999; Ineson et al. 1998; McLain et al. 2002; McLain and Ahmann 2008). Rather than affecting methanotroph activity, it was suggested in these studies that soil moisture reduced CH4 diffusivity into the soil thereby limiting CH4 consumption or that soil moisture increased CH4 production by methanogens. However, by eliminating the effects of CH4 diffusivity on CH4 consumption we were able to observe a significant increase in methanotroph activity under elevated CO2, possibly due to an increase in soil moisture, and perhaps due to other mechanisms that are unclear. Our results indicate that elevated CO2 effects on microbially mediated CH4 consumption may be particularly important for ecosystems that are dry during much of the year and where CH4 consumption is not generally constrained by gas diffusivity (Del Grosso et al. 2000). Indeed, CH4 consumption in a semi-arid grassland in Colorado tended to be greater under elevated CO2 than under ambient CO2, despite an increase in soil moisture (Mosier et al. 2002). Field measurements from a free air CO2 enrichment experiment in a semi-arid grassland in Wyoming showed greater CH4 consumption under elevated CO2 at times when soils were dry (i.e., during much of the growing season, Dijkstra et al., unpublished results).
Not surprisingly, soil moisture explained much of the higher N2O emission in the high water treatment (i.e., the water treatment effect disappeared or was strongly reduced when soil moisture was used as a covariate in the ANCOVA, Table 1). On the other hand, pots under elevated CO2 had greater soil moisture contents during most of the experiment, but elevated CO2 did not increase N2O emission. Possibly, because of our frequent watering, the CO2-induced soil moisture increase may not have been large enough to cause significant increases in N2O emission that were observed in other studies (Arnone and Bohlen 1998; Kanerva et al. 2007; Kettunen et al. 2006; Robinson and Conroy 1999). It was suggested that a greater plant and microbial N demand under elevated CO2 prevented an increase in N2O emission (Hungate et al. 1997a; Mosier et al. 2002), or even reduced N2O emission (Kettunen et al. 2007; Pleijel et al. 1998). However, total plant biomass N content at the end of the experiment was not significantly affected by the CO2 treatment (P > 0.1, ANOVA, data not shown), while we do not have information about microbial N immobilization. Elevated CO2 could also increase N2O emission because of increased labile C input into the soil fueling the denitrifying community (Baggs et al. 2003a, b; Ineson et al. 1998; Kammann et al. 2008). However, nitrifiers that are an important contributor of N2O emission in semi-arid systems (Mosier et al. 2008), do not require labile C as an energy source. Nitrification may have dominated the production of N2O in our study, and thus N2O emission may not have increased under elevated CO2 despite potentially greater inputs of labile C substrates. Others have suggested that the lack of response of N2O emission to elevated CO2 was due to limited potential for denitrification (Ambus and Robertson 1999; Billings et al. 2002). The large variability in N2O emission in response to elevated CO2 among studies indicates a complex number of factors and interactions involved, which requires further investigation.
It is unclear what caused the large effect of species composition on N2O emission. These effects could not be explained simply by species composition effects on soil moisture. When soil NO −3 concentration was used as a covariate then species composition effects on N2O emission were smaller, but remained significant (Table 1). The N2O emission was largest under H. comata. This species also had the highest NO −3 concentration in the soil at the end of the experiment (1.10 mg N kg−1 compared to 0.67, 0.43, 0.74, 0.77, and 0.62 mg N kg−1 for A. frigida, L. dalmatica, B. gracilis, P. smithii, and all species combined respectively). These results suggest that besides variation in soil moisture, variation in NO −3 concentrations in the soil may have contributed to the variation in N2O emission. However, other factors, such as root exudates and other root-derived available soil C (Baggs et al. 2003a, b; Ineson et al. 1998), differences in root morphology affecting root-microbial interactions (Brown et al. 1997), or differences in the composition of the nitrifying and denitrifying microbial community (Patra et al. 2006; Priha et al. 1999) may also have caused species composition effects on N2O loss.
Increasing the plant species number from 1 to 5 did not affect CH4 consumption and only reduced N2O emission early in the experiment. Virtually nothing is known about plant species number or plant diversity effects on CH4 fluxes and very little on N2O fluxes. Reduced N2O emission with increased plant species or plant functional diversity has been associated with more efficient plant N uptake (Epstein et al. 1998; Niklaus et al. 2006). More efficient N uptake early on in our experiment may also have contributed to the reduced N2O emission in the mixed pots, which included species of diverse root morphologies.
Conclusion
Semi-arid grasslands have been shown to be very responsive to elevated CO2 with large shifts in species composition and changes in C and N cycling (Dijkstra et al. 2008; Morgan et al. 2007; Pendall et al. 2003). Changes in species composition and in C and N cycling in response to elevated CO2 likely also affect exchange of the greenhouse gases CH4 and N2O. However, the mechanisms involved in the microbially-mediated exchanges of CH4 and N2O between soils and the surrounding atmosphere have been obscured in previous experiments due to confounding effects of soil water content on microbial activity and diffusivity of gas through the soil matrix. We used a unique method that reduces soil matrix gas diffusional resistance to evaluate how elevated CO2, soil water content, and plant species composition directly affect microbially mediated consumption of CH4 and emission of N2O. Elevated CO2 significantly increased CH4 consumption, while N2O emission significantly differed among plant species and significantly increased with greater soil moisture content. Although soil moisture is a key factor controlling biological processes in semi-arid grasslands, treatment effects on CH4 consumption and N2O emission often remained significant after correcting for treatment effects on soil moisture. This suggests that treatment-induced changes in soil and microbial properties such as availability of plant-derived labile C substrates, inorganic N, and microbial community composition are also important. More research is needed to better understand the interactions of environmental factors and different microbial groups contributing to CH4 and N2O exchange between the soil and atmosphere.
Notes
Trade and company names are given for the reader’s benefit and do not imply endorsement or preferential treatment of any product by the USDA.
References
Ambus P, Robertson GP (1999) Fluxes of CH4 and N2O in aspen stands grown under ambient and twice-ambient CO2. Plant Soil 209:1–8
Ambus P, Zechmeister-Boltenstern S, Butterbach-Bahl K (2006) Sources of nitrous oxide emitted from European forest soils. Biogeosciences 3:135–145
Arnone JA III, Bohlen PJ (1998) Stimulated N2O flux from intact grassland monoliths after two growing seasons under elevated atmospheric CO2. Oecologia 116:331–335
Baggs EM, Blum H (2004) CH4 oxidation and emissions of CH4 and N2O from Lolium perenne swards under elevated atmospheric CO2. Soil Biol Biochem 36:713–723
Baggs EM, Richter M, Cadisch G, Hartwig UA (2003a) Denitrification in grass swards is increased under elevated atmospheric CO2. Soil Biol Biochem 35:729–732
Baggs EM, Richter M, Hartwig UA, Cadisch G (2003b) Nitrous oxide emissions from grass swards during the eighth year of elevated atmospheric pCO2 (Swiss FACE). Glob Change Biol 9:1214–1222
Bédard C, Knowles R (1989) Physiology, biochemistry, and specific inhibitors of CH4, NH +4 , and CO oxidation by methanotrophs and nitrifiers. Microbiol Rev 53:68–84
Benstead J, King GM, Williams HG (1998) Methanol promotes atmospheric methane oxidation by methanotrophic cultures in soils. Appl Environ Microbiol 64:1091–1098
Billings SA, Schaeffer SM, Evans RD (2002) Trace N gas losses and N mineralization in Mojave desert soils exposed to elevated CO2. Soil Biol Biochem 34:1777–1784
Bodelier PLE, Hahn AP, Arth IR, Frenzel P (2000) Effects of ammonium-based fertilisation on microbial processes involved in methane emission from soils planted with rice. Biogeochemistry 3:225–257
Brown TN, Kulasiri D, Gaunt RE (1997) A root-morphology based simulation for plant/soil microbial ecosystem modeling. Ecol Modell 99:275–287
Cheng W (1996) Measurement of rhizosphere respiration and organic matter decomposition using natural 13C. Plant Soil 183:263–268
Cheng W, Johnson DW (1998) Elevated CO2, rhizosphere processes, and soil organic matter decomposition. Plant Soil 202:167–174
Conrad R (1984) Capacity of aerobic microogransims to utilize and grow on atmospheric trace gases H2, CO2, CH4. In: Klug MJ, Reddy CA (eds) Current perspectives in microbial ecology. American Society for Microbiology, Washington, D.C. pp 461–467
Crenshaw CL, Lauber C, Sinsabaugh RL (2008) Fungal control of nitrous oxide production in semiarid grassland. Biogeochemistry 87:17–27
Del Grosso SJ, Parton WJ, Mosier AR, Ojima DS, Potter CS, Borken W, Brumme R, Butterbach-Bahl K, Crill PM, Dobbie K, Smith KA (2000) General CH4 oxidation model and comparisons of CH4 oxidation in natural and managed systems. Glob Biogeochem Cycles 14:999–1019
Dijkstra FA, Hobbie SE, Reich PB, Knops JMH (2005) Divergent effects of elevated CO2, N fertilization, and plant diversity on soil C and N dynamics in a grassland field experiment. Plant Soil 272:41–52
Dijkstra FA, Pendall E, Mosier AR, King JY, Milchunas DG, Morgan JA (2008) Long-term enhancement of N availability and plant growth under elevated CO2 in a semi-arid grassland. Funct Ecol 22:975–982
Epstein HE, Burke IC, Mosier AR, Hutchinson GL (1998) Plant functional type effects on trace gas fluxes in the shortgrass steppe. Biogeochemistry 42:145–168
Frank DA, Groffman PM (1998) Denitrification in a semi-arid grazing ecosystem. Oecologia 117:564–569
Galbally IE, Kirstine WV, Meyer CP, Wang YP (2008) Soil-atmosphere trace gas exchange in semiarid and arid zones. J Environ Qual 37:599–607
Goldman MB, Groffman PM, Pouyat RV, McDonnell MJ, Pickett STA (1995) CH4 uptake and N availability in forest soils along an urban to rural gradient. Soil Biol Biochem 27:281–286
Goverde M, Erhardt A (2003) Effects of elevated CO2 on development and larval food-plant preference in the butterfly Coenonympha pamphilus (Lepidoptera, Satyridae). Glob Change Biol 9:74–83
Heijmans MMPD, Klees H, De Visser W, Berendse F (2002) Response of a Sphagnum bog plant community to elevated CO2 and N supply. Plant Ecol 162:123–134
Hungate BA, Lund CP, Pearson HL, Chapin FS III (1997a) Elevated CO2 and nutrient addition alter soil N cycling and N trace gas fluxes with early season wet-up in a California annual grassland. Biogeochemistry 37:89–109
Hungate BA, Holland EA, Jackson RB, Chapin FS III, Mooney HA, Field CB (1997b) The fate of carbon in grassland under carbon dioxide enrichment. Nature 388:576–579
Hütsch BW, Webster CP, Powlson DS (1994) Methane oxidation in soil as affected by land use, soil pH and N fertilization. Soil Biol Biochem 26:1613–1622
Huxman TE, Smith MD, Fay PA, Knapp AK, Shaw MR, Lolk ME, Smith SD, Tissue DT, Zak JC, Weltzin JF, Pockman WT, Sala OE, Haddad BM, Harte J, Koch GW, Schwinning S, Small EE, Williams DG (2004) Convergence across biomes to a common rain-use efficiency. Nature 429:651–654
Ineson P, Coward PA, Hartwig UA (1998) Soil gas fluxes of N2O, CH4 and CO2 beneath Lolium perenne under elevated CO2: The Swiss free air carbon dioxide enrichment experiment. Plant Soil 198:89–95
Intergovernmental Panel on Climate Change (2007) Working Group I Report. The Physical Science Basis. Technical Summary. Available online. http://www.ipcc.ch. Accessed 7 May 2009
Jacinthe PA, Lal R (2005) Labile carbon and methane uptake as affected by tillage intensity in a Mollisol. Soil Till Res 80:35–45
Kammann C, Müller C, Grünhage L, Jäger H-J (2008) Elevated CO2 stimulates N2O emissions in permanent grassland. Soil Biol Biochem 40:2194–2205
Kanerva T, Regina K, Rämö K, Ojanperä K, Manninen S (2007) Fluxes of N2O, CH4 and CO2 in a meadow ecosystem exposed to elevated ozone and carbon dioxide for three years. Environ Poll 145:818–828
Kang H, Freeman C, Ashendon TW (2001) Effects of elevated CO2 on fen peat biogeochemistry. Sci Total Environ 279:45–50
Kettunen R, Saarnio S, Martikainen PJ, Silvola J (2005) Elevated CO2 concentration and nitrogen fertilisation effects on N2O and CH4 fluxes and biomass production of Phleum pratense on farmed peat soil (2005) Soil Biol Biochem 37:739–750
Kettunen R, Saarnio S, Martikainen PJ, Silvola J (2006) Increase of N2O fluxes in agricultural peat and sandy soil under elevated CO2 concentration: concomitant changes in soil moisture, groundwater table and biomass production of Phleum pratense. Nutr Cycl Agroecosyst 74:175–189
Kettunen R, Saarnio S, Martikainen PJ, Silvola J (2007) Can a mixed stand of N2-fixing and non-fixing plants restrict N2O emissions with increasing CO2 concentration? Soil Biol Biochem 39:2538–2546
Khalil MI, Baggs EM (2005) CH4 oxidation and N2O emission at varied soil water-filled pore spaces and headspace CH4 concentrations. Soil Biol Biochem 37:1785–1794
Koschorreck M, Conrad R (1993) Oxidation of atmospheric methane in soil: measurements in the field, in soil cores and in soil samples. Glob Biogeochem Cycles 7:109–122
McLain JET, Ahmann D (2008) Increased moisture and methanogenesis contribute to reduced methane oxidation in elevated CO2 soils. Biol Fert Soils 44:623–631
McLain JET, Kepler TB, Ahmann DM (2002) Belowground factors mediating changes in methane consumption in a forest soil under elevated CO2. Glob Biogeochem Cycles 16:1050. doi:10.1029/2001GB001439
Menyailo OV, Huwe B (1999) Activity of denitrification and dynamics of N2O release in soils under six tree species and grassland in central Siberia. J Plant Nutr Soil Sci 162:533–538
Menyailo OV, Hungate BA (2003) Interactive effects of tree species and soil moisture on methane consumption. Soil Biol Biochem 35:625–628
Morgan JA, Mosier AR, Milchunas DG, LeCain DR, Nelson JA, Parton WJ (2004a) CO2 enhances productivity, alters species composition, and reduces digestibility of shortgrass steppe vegetation. Ecol Appl 14:208–219
Morgan JA, Pataki DE, Körner C, Clark H, Del Grosso SJ, Grunzweig JM, Knapp AK, Mosier AR, Newton PCD, Niklaus PA, Nippert JB, Nowak RS, Parton WJ, Polley HW, Shaw MR (2004b) Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 140:11–25
Morgan JA, Milchunas DG, LeCain DR, West M, Mosier AR (2007) Carbon dioxide enrichment alters plant community structure and accelerates shrub growth in the shortgrass steppe. P Natl Acad Sci USA 104:14724–14729
Mosier A, Schimel D, Valentine D, Bronson K, Parton W (1991) Methane and nitrous oxide fluxes in native, fertilized and cultivated grasslands. Nature 350:330–332
Mosier AR, Parton WJ, Valentine DW, Ojima DS, Schimel DS, Heinemeyer O (1997) CH4 and N2O fluxes in the Colorado shortgrass steppe 2. Long-term impact of land use change. Glob Biogeochem Cycles 11:29–42
Mosier AR, Morgan JA, King JY, LeCain D, Milchunas DG (2002) Soil-atmosphere exchange of CH4, CO2, NOx, and N2O in the Colorado shortgrass steppe under elevated CO2. Plant Soil 240:201–211
Mosier AR, Parton WJ, Martin RE, Valentine DW, Ojima DS, Schimel DS, Burke IC, Adair EC, Del Grosso SJ (2008) Soil-atmosphere exchange of trace gases in the Colorado shortgrass steppe. In: Lauenroth WK, Burke IC (eds) Ecology of the shortgrass steppe. Oxford University Press, Oxford, pp 342–372
Niklaus PA, Wardle DA, Tate KR (2006) Effects of plant species diversity and composition on nitrogen cycling and the trace gas balance of soils. Plant Soil 282:83–98
Norton U, Mosier AR, Morgan JA, Derner JD, Ingram LJ, Stahl PD (2008) Moisture pulses, trace gas emissions and soil C and N in cheatgrass and native grass-dominated sagebrush-steppe in Wyoming, USA. Soil Biol Biochem 40:1421–1431
Patra AK, Abbadie L, Clays-Josserand A, Degrange V, Grayston SJ, Guillaumaud N, Loiseau P, Louault F, Mahmood S, Nazaret S, Philippot L, Poly F, Prosser JI, Roux XL (2006) Effects of management regime and plant species on the enzyme activity and genetic structure of N-fixing, denitrifying and nitrifying bacterial communities in grassland soils. Environ Microbiol 8:1005–1016
Pendall E, Del Grosso S, King JY, LeCain DR, Milchunas DG, Morgan JA, Mosier AR, Ojima DS, Parton WJ, Tans PP, White JWC (2003) Elevated atmospheric CO2 effects and soil water feedbacks on soil respiration components in a Colorado grassland. Global Biogeochem Cy 17:doi:10.1029/2001GB001821
Phillips RL, Whalen SC, Schlesinger WH (2001a) Influence of atmospheric CO2 enrichment on methane consumption in a temperature forest soil. Glob Change Biol 7:557–563
Phillips RL, Whalen SC, Schlesinger WH (2001b) Response of soil methanotrophic activity to carbon dioxide enrichment in a North Carolina coniferous forest. Soil Biol Biochem 33:793–800
Pleijel H, Sild J, Danielsson H, Klemedtsson L (1998) Nitrous oxide emissions from a wheat field in response to elevated carbon dioxide concentration and open-top chamber enclosure. Envion Pollut 102, S1:167–171
Potts DL, Huxman TE, Cable JM, English NB, Ignace DD, Eilts JA, Mason MJ, Weltzin JF, Williams DG (2006) Antecedent moisture and seasonal precipitation influence the response of canopy-scale carbon and water exchange to rainfall pulses in a semi-arid grassland. New Phytol 170:849–860
Priha O, Grayston SJ, Pennanen T, Smolander A (1999) Microbial activities related to C and N cycling and microbial community structure in the rhizospheres of Pinus sylvestris, Picea abies and Betula pendula seedlings in an organic and mineral soil. FEMS Microbiol Ecol 30:187–199
Robinson D, Conroy JP (1999) A possible plant-mediated feedback between elevated CO2, denitrification and the enhanced greenhouse effect. Soil Biol Biochem 31:43–53
Stein VB, Hettiaratchi JPA (2001) Methane oxidation in three Alberta soils: influence of soil parameters and methane flux rates. Environ Technol 22:101–111
Ullah S, Frasier R, King L, Picotte-Anderson N, Moore TR (2008) Potential fluxes of N2O and CH4 from soils of three forest types in Eastern Canada. Soil Biol Biochem 40:986–994
Von Fischer JC, Hedin LO (2007) Controls on soil methane fluxes: tests of biophysical mechanisms using stable isotope tracers. Glob Biogeochem Cycles 21, doi:10.1029/2006GB002687
Acknowledgements
We thank Joseph Hansen and Mary Smith for technical assistance. We thank Jean McLain, Joe von Fischer, and two anonymous reviewers for a critical review of a previous version of the manuscript. This publication is based upon work supported by the Agricultural Research Service under the ARS GRACEnet Project.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Elizabeth M. Baggs.
Rights and permissions
About this article
Cite this article
Dijkstra, F.A., Morgan, J.A., LeCain, D.R. et al. Microbially mediated CH4 consumption and N2O emission is affected by elevated CO2, soil water content, and composition of semi-arid grassland species. Plant Soil 329, 269–281 (2010). https://doi.org/10.1007/s11104-009-0152-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-009-0152-5