
Abstract
Purpose
The organic cation transporters (OCTs) and multidrug and toxin extrusions (MATEs), located in the basolateral and apical membrane of proximal tubular cells respectively, are crucial determinants of renal elimination and/or toxicity of cationic drugs such as cisplatin. The purpose of this study was to discover selective OCT inhibitors over MATEs, and explore their potential to protect against cisplatin-induced nephrotoxicity that is clinically common.
Methods
The inhibition by select compounds on the uptake of the probe substrate metformin was assessed in HEK293 cells overexpressing human OCT2, OCT1, MATE1, MATE2-K, and mouse Oct2, Oct1, and Mate1. Furthermore, the effects of carvedilol on organic cation transporter-mediated cellular and renal accumulation of metformin and cisplatin, and particularly the toxicity associated with cisplatin, were investigated in HEK293 cells and mice.
Results
Five selective OCT inhibitors were identified through the screening of forty-one drugs previously reported as the inhibitors of OCTs and/or MATEs. Among them, carvedilol showed the most selectivity on OCTs over MATEs (IC50: 3.6 μM for human OCT2, 103 μM for human MATE1 and 202 μM for human MATE2-K) in the cellular assays in vitro, with the selectivity in mice as well. Moreover, carvedilol treatment could significantly decrease cisplatin accumulation and ameliorate its toxicity both in vitro in cells and in vivo in mouse kidney.
Conclusions
Our data indicate that selective inhibition of OCTs by carvedilol may protect from cisplatin-induced nephrotoxicity by restraining the cellular entry of cisplatin via OCTs, while having no impact on its elimination through MATEs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Over the past decades, it has been increasingly recognized that drug transporters play a crucial role in drug disposition, efficacy and toxicity in the kidney. Various drug transporters are located in human renal tubular cells, among which organic cation transporter 2 (OCT2) and multidrug and toxin extrusion 1 and 2-K (MATE1, MATE2-K) are critically involved in renal accumulation and elimination of cationic drugs. OCTs and MATEs together are regarded as an organic cation transport system due to their significant overlap on structurally unrelated exogenous and endogenous substrates, including tetraethyl ammonium (TEA) (1), 1-methyl-4-phenylpyridinium (MPP+) (2), metformin (3), antiretroviral drugs (4,5), anticancer drugs (6) and creatinine (7,8). While both Oct1 and Oct2 are highly expressed in rodent kidney, OCT2 is the predominant OCT isoform expressed in human kidney (9). In contrast, Mate1 is the only isoform found in rodent kidney, while MATE1 and MATE2-K are both expressed in human kidney. OCTs, which are located in the basolateral membrane, are driven by the internal negative membrane potential to uptake substrates into the tubular cells; however, MATEs, which are resided in the apical membrane, transport substrates into the urine by using the proton gradient.
Both OCTs and MATEs are important in determining renal accumulation and elimination of cisplatin, an anti-cancer drug, the use of which is dose-limited largely due to its nephrotoxicity. Renal elimination and accumulation of cisplatin were significantly reduced in Oct1/2 double knockout mice as compared to the control wild-type mice (10,11). Moreover, cancer patients who were heterozygotes for OCT2 variant 808G > T showed a significantly less nephrotoxicity than those carrying reference alleles only, as indicated by a decreased level of plasma creatinine and an improved renal histology score (11,12). In contrast, as compared to wild-type mice, Mate1 knockout mice have showed an increased accumulation of cisplatin and more severe cisplatin-induced nephrotoxicity (13). Consistently, we have also found that ondansetron can increase the renal accumulation of cisplatin and aggravate its nephrotoxicity via potent inhibition of Mate1 in mice (14). Cisplatin and oxaliplatin have been reported to exhibit more severe nephrotoxicity than other platinum-based drugs due to a higher affinity to hOCT2 (6,15). Interestingly, oxaliplatin induces less nephrotoxicity than cisplatin probably because it is a better substrate of MATE2-K for excretion into the urine (16).
In the present study, we sought to discover a selective inhibitor of OCT2 over MATE1/2-K by screening the known OCTs/MATEs inhibitors. With a relatively selective OCT2 inhibitor identified from the screening, we would further test the hypothesis that selective inhibition towards OCTs over MATEs could effectively reduce the entry of a risky substrate such as cisplatin into the renal tubular cells via OCTs and leave the elimination via MATEs unaffected, protecting from the drug-induced nephrotoxicity.
Materials and Methods
Chemicals and Reagents
Dulbecco’s Modified Eagle’s medium (DMEM), Phosphate-buffered saline (PBS), Lipofectamine 2000, and fetal bovine serum were purchased from Invitrogen. [14C]-metformin (1.0 mCi, 90 mCi/mmol) was purchased from Moravek Biochemicals and Radiochemicals (Brea, CA). All other compounds, including cisplatin, carvedilol, and unlabeled metformin, were obtained from Sigma Chemical Co. (St. Louis, MO). Cell viability was tested by cell counting kit-8 (Enzo Life Science Inc). The nitric acid used to lysate cells and the chemical standards for cisplatin measurement by inductively coupled plasma mass spectrometry (ICP-MS) were of trace ICP-MS grade and supplied by Sigma-Aldrich Corp. (St Louis, MO).
Cell Lines and Cell Culture
Stable HEK-293 cells overexpressing human (h) OCT2, hMATE1, or hMATE2-K were previously established in our lab (14). The cells were cultured in DMEM supplemented with 10% fetal bovine serum, 100 U/mL penicillin, 100 μg/mL streptomycin, and 20 μg/mL hygromycin and incubated at 37°C in 5% CO2 in a humidified atmosphere. Transient transfection was used to overexpress human OCT1, mouse Oct1, mouse Oct2, and mouse Mate1 in HEK-293 cells by using lipofectamine 2000 (Invitrogen) according to manufacturer’s instruction. The overexpression of these transporters in HEK-293 cells was confirmed by real-time PCR and functional tests.
Inhibition of Metformin Uptake in HEK-293 Cells
Metformin was used as the substrate to probe the activities of OCT and MATE transporters. The cells were seeded to poly-D-lysine coated 24-well plates at 2.5 × 105 cells per well. After 24 h of culture, the inhibition studies were performed as described previously as for OCT2 (14), and with a minor modification as for MATEs. Briefly, as the transport of substrates by MATEs is driven by an inward proton gradient in vivo in tissues, here the transport direction in vitro was reversed by introducing H+ into the cells with an acidified K+ based buffer (140 mM KCl, 0.4 mM KH2PO4, 0.8 mM MgSO4, 1.0 mM CaCl2, 25 mM glucose, 10 mM HEPES, and 30 mM NH4Cl, KBB) for 15 min at 37°C, then for 5 min in NH4Cl-free KBB. Afterwards, the cells were immediately co-incubated with inhibitors and metformin (50 μM, containing 1/20 [14C]-labeled metformin) in KBB for 5 min. The uptake was stopped by washing with ice-cold KBB for three times. Cells were then lysed in 300 μL PBS with 1% triton X-100 by shaking for 20 min, with 250 μL cell lysates being added into 3 mL scintillation buffer (Econo-Safe™ Economical biodegradable cocktail, Research Products International Corp, Mount Prospect, IL). Radioactivity was measured by Tri-Carb 2910TR liquid scintillation analyzer (PerkinElmer, Boston, MA).
Cisplatin Uptake and Cytotoxicity in HEK-293 Cells
HEK-hOCT2 and the control cells expressing the empty vector (HEK-control) were treated with cisplatin (100 μM) for 2 h with or without a selective OCT2 inhibitor (carvedilol 10 μM). The cells were then washed by PBS, and cultured for additional 22 h in normal medium before the cell viability and intracellular cisplatin level being measured. The cell viability was determined by the cell counting kit-8 (CCK-8, Enzo Life Science Inc.) following the manufacturer’s instruction. Briefly, 200 μL of the cell counting medium (10% of CCKi-8 in the serum-containing DMEM) was added into each well. The medium was transferred to a 96-well plate and absorbance was measured at 450 nm after incubation at 37°C for 30 min. ICP-MS was used to quantitate the intracellular platinum as described (17). Briefly, cells were lysed by 200 μL of nitric oxide (67–70%, Sigma-Aldrich, St. Louis, MO), then shaken for 15 min. Thereafter, 100 μL of cell lysate along with 20 μL internal standard (Iridium, 50 μg/mL) and 1880 μL 2% nitric oxide was transferred to a 2 mL tube which was ready for quantification by ICP-MS (Agilent 7700). Protein concentrations were measured by a bicinchoninic acid (BCA) protein assay kit (Bio-Rad, Hercules, CA) and then used to normalize the cisplatin concentration values determined from ICP-MS analysis.
Animal Study
All procedures were approved by the Institutional Animal Care and Use Committee at the University of Maryland, Baltimore, and carried out in accordance with NIH guidelines for animal experimentation. Male C57BL/6J mice at 12–14 weeks of age were used. The mice were fasted for 6 h and then administrated with carvedilol (2 mg/kg) 30 min before and together with metformin (10 mg/kg, with 1/20 [14C]-metformin) or cisplatin (10 mg/kg) intraperitoneally. For metformin pharmacokinetics, the blood samples were collected before and at 5, 15, 30, 45, 60, 90, and 120 min after metformin administration. Then 15 μL of plasma were added into 2 mL scintillation buffer for radioactivity counting as described above. To determine the tissue accumulation of metformin and cisplatin, the mice were euthanized 30 min and 8 h after administration of metformin and cisplatin, respectively, with liver and kidney tissue being collected and homogenized for radioactivity counting and ICP-MS detection, respectivley. To determine cisplatin nephrotoxicity, the mice were euthanized 72 h after administration of carvedilol and/or cisplatin, with kidney tissues being collected for histology and molecular marker examination. The mRNA level of Kim1 was used as the molecular marker of nephrotoxicity as described (18). The haematoxylin and eosin (H&E) staining of kidney tissues was performed by the Pathology Department of School of Medicine, University of Maryland, Baltimore.
Statistical Analysis
GraphPad Prism version 5.01 (GraphPad Software, San Diego, CA) was used to perform the statistical analysis. All in vitro experiments were repeated for at least three times, with 3 to 6 measurements being performed in each repeat. Representative data were shown, and presented as Mean ± standard deviation (SD). For pharmacokinetics (PK), the area under the concentration-time curve (AUC) value was calculated using a linear trapezoidal method. The maximum concentration (Cmax) was the highest observed plasma concentration and Tmax was the time at which Cmax occurred. A two-tail Student’s t test was used for statistical comparison between two groups, while one-way analysis of variance (ANOVA) followed by post hoc Tukey comparison for more than two different groups. Non-parameter statistical analysis was used to compare the physicochemical parameters of the tested compounds which were obtained from www.drugbank.ca and predicted by ChemAxon. p < 0.05 was set as the threshold for the significantly statistic difference.
Results
Inhibitor Validation for hOCT2, hMATE1 and hMATE2-K
In an attempt to discover selective hOCT2 inhibitors over hMATEs, a total of 41 compounds which had been previously reported to be either hOCT2 or hMATEs inhibitors were screened for their inhibitory profile towards hOCT2, hMATE1 and hMATE2-K by using metformin as the probe substrate. The half-maximal inhibitory concentration (IC50) was calculated to reflect the inhibitory potency of each inhibitor over a series of concentrations with the highest not exceeding 500 μM. A compound would be defined as an inhibitor when at least 50% of inhibition on metformin uptake could be achieved with the concentrations examined. The relative IC50 values obtained from our study were generally comparable to previously reported ones even though a different probe substrate or different concentrations of the probe substrate were used. For instance, imatinib showed a selective inhibition on hMATEs, with the IC50 being calculated as 28.9 μM, 0.1 μM and 13.9 μM for hOCT2, hMATE1 and hMATE2-K, respectively, as compared to 5.81 μM, 0.048 μM and 0.48 μM obtained previously by others (19). Consistent with a previous report (14), more compounds showed a relatively lower IC50 for hMATE1/2-K than hOCT2. Among all the screened compounds, flutamide, fucidic acid, bufexamac, atenol and carvedilol (Table I) showed a more potent inhibitory effect on hOCT2 than hMATEs, thus being categorized as the selective hOCT2 inhibitors. With respect to inhibition towards the two hMATEs, 21 compounds had a similar IC50 for hMATE1 to that of hMATE2-K, 16 compounds showed a marginally less potent inhibitory effect on hMATE2-K than hMATE1, and four compounds which were not inhibitors of hMATE1 turned out to be inhibitors of hMATE2-K.
Notably, four compounds were grouped as hOCT2-enhancer due to their ability to increase hOCT2 activity at the low but clinically relevant concentrations (less than 10 μM) (gefitinib, cimnasizine, saquinavir, itraconazole; the last four compounds in Table I), although these four compounds could inhibit hOCT2-mediated metformin uptake at the high concentrations. In addition, they were also potent inhibitors of hMATE1, and two of them were the inhibitors of hMATE2-K as well. The physicochemical properties including water solubility, molecular weight, logP, Pka acidic and basic, physiological charge, hydrogen acceptor and donor count, polar surface area, rotatable bound count, refractivity, polarizability and number of rings were analyzed and compared with non-hOCT2-enhancers. Of all physicochemical parameters examined, the hOCT2-enhancers were found to have more rings (5.0 ± 1.4 vs. 3.2 ± 1.3) and higher refractivity (156 ± 43.6 vs. 103 ± 32.2) as compared to the non-enhancers. (Fig. 1).

Comparison of physicochemical properties between hOCT2 enhancers and non-enhancers. The physicochemical properties of compounds were obtained from www.drugbank.ca and predicted by ChemAxon. Of all physicochemical parameters examined (refer to the result section), there are significantly more rings and refractivity for the hOCT2 enhancers than those for non-hOCT2-enhancers. The parameters were compared by non-parameter statistics. *p < 0.05 compared to non- hOCT2-enhancers.
Characterization of Carvedilol as a Selective OCTs Inhibitor
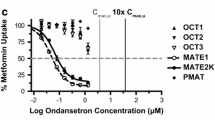
Among the five identified selective hOCT2 inhibitors, carvedilol was chosen for further investigation as it possessed the selectivity hOCT2 inhibition at clinical relevant concentrations. In addition to hOCT2, hMATE1, and hMATE2-K, the inhibition by carvedilol towards other homologs and orthologs of OCT and MATE was examined. The calculated IC50 values were 3.6 μM for hOCT2, 6.4 μM for hOCT1, 103 μM for hMATE1, and 202 μM for hMATE2-K when using metformin as the probe substrate (Fig. 2). Similar to the results obtained for the human orthologs, carvedilol also showed a selective inhibition on mouse (m) Oct1 and mOct2 over mMate1, with the IC50 values of 2.8 μM for mOct1, 23.1 μM for mOct2, and 67.0 μM for mMate1, respectively (Fig. 2).

Inhibition by carvedilol on metformin uptake mediated by human OCT2, OCT1, MATE1 and MATE2-K (A) and mouse Oct2, Oct1, Mate1 (B). HEK293 cells overexpressed with human OCT2, MATE1 and MATE2-K (A) and mouse Oct2, Oct1, Mate1 (B) were incubated with metformin (50 μM, with 1/20 [14C]-labeled metformin) in the absence and presence of different concentrations of carvedilol under 37°C. The time of uptake was set for 5 min for MATEs, and 10 min for OCTs. The curves were obtained by nonlinear regression fitting by GraphPad. Mean ± SD of three measurements is shown.
Carvedilol Inhibited Cationic Drug Transporters In Vivo
To explore whether the selective inhibition of carvedilol on OCTs over MATEs in vitro could be translated to an altered drug disposition in vivo, we studied the effect of carvedilol on the accumulation of metformin, a classical substrate of organic cation transporters, in mouse kidney and liver, where both Octs and Mates are highly expressed. The mice were given two doses of carvedilol (2 mg/kg and 20 mg/kg) along with metformin. Carvedilol (2 mg/kg) significantly decreased the accumulation of metformin in the kidney and liver (Fig. 3a), implicating a more potent inhibition on mouse Octs than Mate1. The accumulation of metformin in the kidney and liver with 2 mg/kg carvedilol were 17.6 ± 2.8 ng/mg and 8.1 ± 0.2 ng/mg compared to 26.2 ± 5.3 ng/mg (p = 0.005) and 10.4 ± 1.4 ng/mg (p = 0.002) of vehicle control group, respectively. In contrast, carvedilol at the higher dose (20 mg/kg) increased the accumulation of metformin in the kidney (41.0 ± 7.2 ng/mg with carvedilol vs. 18.2 ± 0.9 ng/mg with vehicle, p = 0.001) and liver (19.0 ± 1.9 ng/mg with carvedilol vs. 9.3 ± 0.9 ng/mg with vehicle, p < 0.001), suggesting an inhibitory effect on mMate1 in addition to mOCTs (Fig. 3b). Hence, the lower dose of carvedilol (2 mg/kg) was chosen in the following PK studies due to its selective inhibition on mOcts in vivo. Carvedilol significantly increased plasma concentrations of metformin at 30, 45 and 60 min compared to vehicle control group (p < 0.05, Fig. 4). The area under curve (AUC0-120min) of metformin was significantly higher in carvedilol treatment group than control group (260 ± 44.7 μg·min/mL vs. 207 ± 40.7 μg·min/mL, p < 0.05) (Table II). There was also a trend of increase regarding Cmax and half-life in carvedilol treatment group versus vehicle group (Cmax, 4.25 ± 0.58 μg/mL vs. 3.77 ± 0.48 μg/mL, p = 0.09; and T1/2, 21.5 ± 5.4 min vs. 22.2 ± 4.7 min, p = 0.06), though no statistical significance was achieved. Overall, the data suggested that carvedilol at a low dose (2 mg/kg) could reduce the accumulation of metformin in mouse liver and kidney, and increase its systemic exposure probably due to its selective inhibition on mOcts.

Effects of carvedilol administration on the accumulation of metformin in mouse kidney and liver. A clinically relevant dose (2 mg/kg) of carvedilol (a) decreased while a relatively high dose (20 mg/kg) (b) increased the accumulation of metformin in mouse kidney and liver. C57BL/6J mice were injected intraperitoneally with carvedilol (2 mg/kg or 20 mg/kg) 30 min pre- and along with metformin (10 mg/kg, with 1/20 [14C]-labeled metformin) or saline vehicle. The mice were euthanized 30 min after metformin administration, with liver and kidney tissues being collected, weighted and homogenized for determination of radioactivity. Mean ± SD from 4 to 6 mice/group is shown. *p < 0.05 compared to the vehicle group.

Carvedilol administration increased the plasma exposure of metformin. C57BL/6J mice were injected intraperitoneally with carvedilol (2 mg/kg) 30 min pre- and along with metformin (10 mg/kg, with 1/20 [14C]-labeled metformin) or saline vehicle. The blood samples were collected before and 5, 15, 30, 45, 60, 90, 120 min after the injection of metformin. Mean ± SD from eight mice/group is shown. *p < 0.05 compared to vehicle administration.
Carvedilol Reduced Cellular and Renal Accumulation of Cisplatin and Ameliorated Cisplatin-Induced Nephrotoxicity via Inhibition on OCT Transporters
The above in vitro and in vivo data, and particularly the selectivity of carvedilol as an inhibitor of OCTs over MATEs, prompted us to determine if carvedilol could inhibit cellular OCT-mediated uptake of cisplatin and protect from cisplatin-induced nephrotoxicity. We first performed in vitro studies in HEK-hOCT2 and control cells. As expected, carvedilol significantly decreased the accumulation of cisplatin in HEK-hOCT2 stable cells (3.77 ± 0.02 ppm/mg protein vs. 4.17 ± 0.09 ppm/mg protein, p = 0.002) but not in control cells (3.87 ± 0.06 ppm/mg protein with carvedilol vs. 3.88 ± 0.07 ppm/mg protein, p > 0.05, Fig. 5a). Consistently, there was a decrease of cell viability for the HEK-hOCT2 cells exposed to cisplatin treatment, which could be significantly rescued by co-treatment of carvedilol (Fig. 5b). We subsequently studied the effect of carvedilol on renal cisplatin accumulation and cisplatin-induced nephrotoxicity in mice. Two injections of carvedilol (2 mg/kg), 30 min pre- and along with cisplatin (10 mg/kg) injection, were given to the mice. The accumulation of cisplatin in the kidney was significantly lower in carvedilol treatment group (6.36 ± 0.44 ppm/g tissue) than vehicle treatment group (9.12 ± 0.63 ppm/g tissue, p < 0.001, Fig. 6a). A dose of 10 mg/kg of cisplatin was used to induce renal damage in mice as reflected by the dramatic increase of Kim-1 gene transcripts (20), which was almost be brought down to the normal base by co-administration of carvedilol (Fig. 6b). This was consistent with the changes in kidney histology. No visible morphological change was present in the vehicle and carvedilol treatment-only groups, while a single dose of cisplatin (10 mg/kg) could significantly cause renal damage including tubular dilatation, tubular cell necrosis with intratubular cast formation, which could be substantially ameliorated by co-administration of carvedilol (Fig. 6c).

Carvedilol decreased the accumulation of cisplatin in HEK293-hOCT2 cells (a), and rescued these cells from cisplatin-induced cytotoxicity (b). The control cells (a) and HEK293-hOCT2 cells (a & b) were treated with carvedilol (10 μM) and cisplatin (100 μM) for 2 h, with cell viability and accumulation of cisplatin being tested after incubation in normal media for another 22 h. Mean ± SD of three measurements is shown. *p < 0.05 compared with absence of carvedilol.

Carvedilol alleviated cisplatin-induced nephrotoxicity by decreasing renal accumulation of cisplatin in mice. (a) C57BL/6J mice were injected intraperitoneally with carvedilol (2 mg/kg) 30 min pre- and along with cisplatin (10 mg/kg) (A/B/C) or vehicle control. The mice were then euthanized 8 h after cisplatin administration, with kidney tissues being collected and digested for cisplatin quantification by ICP-MS. *P < 0.05 compared to vehicle group. (b & c) The mice were euthanized 72 h after cisplatin administration with kidney sample being collected for mRNA isolation (b) and histology analysis (c). *p < 0.05 compared with cisplatin treatment only (b). Carvedilol and cisplatin co-treatment group showed less tubular necrosis as indicated by a decrease of pink tubular (c). The tubular cell necrosis with intratubular cast formation were indicated by black arrows.
Discussion
The OCTs and MATEs, expressed in the basolateral and apical membrane of proximal tubule epithelial cells, respectively, are crucial in renal elimination of certain cationic drugs; thus, inhibition of these transporters posts a risk of drug-drug interaction and drug-induced nephrotoxicity. Previously, hOCT2-mediated drug accumulation in renal cells was believed to be an important factor associated with drug-induced nephrotoxicity (11). Later on, following the discovery of MATE1/2-K, the importance of these transporters in drug-induced renal toxicity has been increasingly recognized as well (15,16).
Although cisplatin is a very effective anti-cancer drug, its usage is dose-limited by the associated nephrotoxicity (21). Cisplatin may enter and accumulate in renal tubular cells via a process mediated by transporters such as OCT2, leading to nephrotoxicity (22). Bearing the idea of alleviating cisplatin-induced nephrotoxicity by reducing its renal accumulation, efforts have been made to use a hOCT2 inhibitor. For example, cimetidine was thought to have such a therapeutic potential since it is a potent inhibitor of hOCT2. However, contradictory results have been reported with respect to its effectiveness in protection of cisplatin-induced nephrotoxicity ever since (23,24,25,26,27), which might be partially, if not all, explained by the fact that cimetidine is actually a more potent inhibitor towards MATE1 than OCT2 at clinically relevant concentrations (28). Our previous study also showed that ondansetron could increase the accumulation of cisplatin in the kidney because it is a more potent inhibitor towards MATEs than OCTs (14). Herein, the main purpose of this study was to identify selective inhibitors on hOCT2 over hMATE1/2-K and explore their potential to protect cisplatin-induced nephrotoxicity. A total of 41 drugs, which have been previously characterized as the inhibitors of either OCTs, MATEs, or both, were selected and re-screened for their inhibitory effects towards hOCT2, hMATE1 and hMATE2-K. Consistent with previous studies (29,30), approximate half of the compounds examined showed comparable inhibition between hOCT2 and hMATE1/2-K. Of importance, we were able to identify five compounds with selectivity on hOCT2 over hMATEs, of which carvedilol has been chosen for the following studies due to its highest selectivity for hOCT2 over hMATEs. Notably, the selective inhibition of carvedilol was also observed between mouse Oct1/2 and Mate1, though not as dramatic as that between human orthologs, indicating mouse as a suitable in vivo model to examine the effectiveness of selective inhibition by carvedilol in protection of cisplatin-induced nephrotoxicity.
Carvedilol is an beta adrenergic receptor blocker used to treat congestive heart failure and hypertension (31). Carvedilol has a low oral bioavailability (24%) due to extensive metabolism in the liver mainly by CYP2D6 and CYP2C9. The peak plasma concentration is generally achieved 1–2 h after oral administration and the terminal half-life is 6–10 h (32). The plasma binding of carvedilol is approximately 95% and carvedilol in the plasma is mostly bound to the albumin. The concentration of unbound carvedilol in the plasma are expected to be still much lower than the IC50 values of OCTs. However, carvedilol is prone to accumulate in certain organs, in particular kidney and liver (33). Therefore, a clinical relevant dose of carvedilol may be able to locally inhibit OCTs and/or MATEs in these tissues/organs. Two doses of carvedilol (2 and 20 mg/kg) were tested in mice during our current study to investigate whether carvedilol could selectively inhibit Oct1/2 over Mate1 by using metformin as the probe substrate. Notably, the lower one (2 mg/kg) is clinical relevant based on the dose extrapolation between mice and humans by body surface area (BSA) normalization with the consideration of human oral carvedilol pharmacokinetics. We found that the lower dose could decrease the accumulation of metformin in the kidney and liver, while the higher one could increase the accumulation instead. It thus likely that the drug concentration of the lower dose was not high enough to inhibit Mate1, while sufficient to inhibit Oct1/2 and achieve its selectivity in the kidney and liver. In contrast, the concentration of the higher dose could inhibit both Oct1/2 and Mate1. It is worthy to mention that the blocking of Oct1/2 would not always result in a reduced metformin accumulation in the kidney. Metformin could apparently enter renal and liver cells via other routes, such as passive diffusion or other transporters like OCTNs (34,35). For instance, Higgins and coworkers found that the enhanced plasma exposure of metformin could cancel out the double knockout effect of Oct1/2, leading to unchanged accumulation in the liver and kidney (36). In contrast, blocking of Mate1 could dramatically increase the plasma exposure and tissue accumulation of metformin (14,37). Therefore, with respect to our study, the higher dose of carvedilol could block the Oct1/2 and Mate1 in mice kidney and resulted in an increased exposure in the plasma, which could in turn lead to an increase in its accumulation both in the liver and in the kidney via those other routes. Further studies with genetic mouse models of Oct and Mate-deficiency are necessary to clarify the role of these transporters in mediating the effects of carvedilol on renal and liver disposition of cationic drugs.
In the present study, carvedilol (2 mg/kg) was further investigated regarding its potential to protect against cisplatin-induced nephrotoxicity in mice by selective inhibition of mOcts. The function of OCTs and MATEs is not a major determinant of cisplatin systemic pharmacokinetics. However, it is critical to the renal accumulation of cisplatin (11). A significant decrease of cisplatin accumulation was observed in the kidney by carvedilol co-treatment, along with dramatic decrease of Kim-1 expression and substantial amelioration of histological damage in the kidney. Interestingly, carvedilol has been previously shown to alleviate cisplatin-induced nephrotoxicity probably by reducing oxidative stress, apoptosis and mitochondrial toxicity (38,39,40). The mice or rats were treated with carvedilol up to three days following cisplatin injection in these studies. In our studies, the mice only received carvedilol treatment 30 min before and together with injection of cisplatin, which led to a significant decrease in renal cisplatin accumulation. Whereas carvedilol may protect cisplatin-induced nephrotoxicity via additional mechanisms, our findings have strongly supported the hypothesis that a selective OCT inhibitor may alleviate cisplatin-induced nephrotoxicity by reducing cisplatin accumulation in the kidney. In particular, carvedilol is usually very safe, well tolerated, and widely approved in many countries for the treatment of high blood pressure and heart failure (41). There is a great potential of carvedilol being developed as a clinical adjuvant therapy for cisplatin-induced nephrotoxicity.
Unexpectedly, we found that several drugs were able to intriguingly enhance hOCT2-mediated uptake of metformin at their clinically relevant concentrations. Similar phenomenon has been reported previously (42), with the mechanism being elusive. Since these compounds were the inhibitors of MATEs (Table I), their use may particularly increase the accumulation of OCTs/MATEs substrate in the kidney, thus an increased risk of drug-induced renal toxicity. The underlying mechanism and clinical implication of these enhancers of OCT activities deserve further investigation and is one of our current interests.
In summary, we have screened a variety of known OCTs/MATEs inhibitors and identified carvedilol as a selective inhibitor of OCTs over MATEs. For the first time, we demonstrated that carvedilol could protect against cisplatin-induced nephrotoxicity at least partially by decreasing renal accumulation of cisplatin. Futures studies are warranted to assess the potential clinical benefits of selective hOCT2 inhibitors in alleviation of the nephrotoxicity associated with cisplatin chemotherapy.
Abbreviations
- AUC:
-
Area under curve
- Cmax :
-
The maximum concentration
- DDI:
-
Drug-drug interaction
- H&E staining:
-
Haematoxylin and Eosin staining
- IC50 :
-
The half maximal inhibitory concentration
- MATE:
-
Multidrug and toxin extrusions
- MPP+ :
-
1-methyl-4-phenylpyridinium
- OCT:
-
Organic cation transporters
- T1/2 :
-
Half life
- TEA:
-
Tetraethyl ammonium
- Tmax :
-
Time of the maxium concentration
References
Gorboulev V, et al. Cloning and characterization of two human polyspecific organic cation transporters. DNA Cell Biol. 1997;16(7):871–81.
Grundemann D, et al. Selective substrates for non-neuronal monoamine transporters. Mol Pharmacol. 1999;56(1):1–10.
Kimura N, et al. Metformin is a superior substrate for renal organic cation transporter OCT2 rather than hepatic OCT1. Drug Metab Pharmacokinet. 2005;20(5):379–86.
Muller F, et al. Role of organic cation transporter OCT2 and multidrug and toxin extrusion proteins MATE1 and MATE2-K for transport and drug interactions of the antiviral lamivudine. Biochem Pharmacol. 2013;86(6):808–15.
Jung N, et al. Organic cation transporters OCT1 and OCT2 determine the accumulation of lamivudine in CD4 cells of HIV-infected patients. Infection. 2013;41(2):379–85.
Burger H, et al. Differential transport of platinum compounds by the human organic cation transporter hOCT2 (hSLC22A2). Br J Pharmacol. 2010;159(4):898–908.
Urakami Y, et al. Creatinine transport by basolateral organic cation transporter hOCT2 in the human kidney. Pharm Res. 2004;21(6):976–81.
Zhang Y, et al. Impact on creatinine renal clearance by the interplay of multiple renal transporters: a case study with INCB039110. Drug Metab Dispos. 2015;43(4):485–9.
Motohashi H, et al. Gene expression levels and immunolocalization of organic ion transporters in the human kidney. J Am Soc Nephrol. 2002;13(4):866–74.
Jonker JW, et al. Deficiency in the organic cation transporters 1 and 2 (Oct1/Oct2 [Slc22a1/Slc22a2]) in mice abolishes renal secretion of organic cations. Mol Cell Biol. 2003;23(21):7902–8.
Filipski KK, et al. Contribution of organic cation transporter 2 (OCT2) to cisplatin-induced nephrotoxicity. Clin Pharmacol Ther. 2009;86(4):396–402.
Iwata K, et al. Effects of genetic variants in SLC22A2 organic cation transporter 2 and SLC47A1 multidrug and toxin extrusion 1 transporter on cisplatin-induced adverse events. Clin Exp Nephrol. 2012;16(6):843–51.
Nakamura T, et al. Disruption of multidrug and toxin extrusion MATE1 potentiates cisplatin-induced nephrotoxicity. Biochem Pharmacol. 2010;80(11):1762–7.
Li Q, et al. Ondansetron can enhance cisplatin-induced nephrotoxicity via inhibition of multiple toxin and extrusion proteins (MATEs). Toxicol Appl Pharmacol. 2013;273(1):100–9.
Yonezawa A, et al. Cisplatin and oxaliplatin, but not carboplatin and nedaplatin, are substrates for human organic cation transporters (SLC22A1-3 and multidrug and toxin extrusion family). J Pharmacol Exp Ther. 2006;319(2):879–86.
Yokoo S, et al. Differential contribution of organic cation transporters, OCT2 and MATE1, in platinum agent-induced nephrotoxicity. Biochem Pharmacol. 2007;74(3):477–87.
Minami T, et al. Accumulation of platinum in the intervertebral discs and vertebrae of ovarian tumor-bearing patients treated with cisplatin. Biol Trace Elem Res. 1994;42(3):253–7.
Wang EJ, et al. Validation of putative genomic biomarkers of nephrotoxicity in rats. Toxicology. 2008;246(2–3):91–100.
Minematsu T, Giacomini KM. Interactions of tyrosine kinase inhibitors with organic cation transporters and multidrug and toxic compound extrusion proteins. Mol Cancer Ther. 2011;10(3):531–9.
Vaidya VS, et al. Kidney injury molecule-1 outperforms traditional biomarkers of kidney injury in preclinical biomarker qualification studies. Nat Biotechnol. 2010;28(5):478–85.
Madias NE, Harrington JT. Platinum nephrotoxicity. Am J Med. 1978;65(2):307–14.
Yonezawa A, et al. Association between tubular toxicity of cisplatin and expression of organic cation transporter rOCT2 (Slc22a2) in the rat. Biochem Pharmacol. 2005;70(12):1823–31.
Sleijfer DT, et al. The protective potential of the combination of verapamil and cimetidine on cisplatin-induced nephrotoxicity in man. Cancer. 1987;60(11):2823–8.
Katsuda H, et al. Protecting cisplatin-induced nephrotoxicity with cimetidine does not affect antitumor activity. Biol Pharm Bull. 2010;33(11):1867–71.
Zhang J, Zhou W. Ameliorative effects of SLC22A2 gene polymorphism 808 G/T and cimetidine on cisplatin-induced nephrotoxicity in Chinese cancer patients. Food Chem Toxicol. 2012;50(7):2289–93.
Sprowl JA, et al. Conjunctive therapy of cisplatin with the OCT2 inhibitor cimetidine: influence on antitumor efficacy and systemic clearance. Clin Pharmacol Ther. 2013;94(5):585–92.
Dorr RT, Soble MJ. Cimetidine enhances cisplatin toxicity in mice. J Cancer Res Clin Oncol. 1988;114(1):1–2.
Ito S, et al. Competitive inhibition of the luminal efflux by multidrug and toxin extrusions, but not basolateral uptake by organic cation transporter 2, is the likely mechanism underlying the pharmacokinetic drug-drug interactions caused by cimetidine in the kidney. J Pharmacol Exp Ther. 2012;340(2):393–403.
Wittwer MB, et al. Discovery of potent, selective multidrug and toxin extrusion transporter 1 (MATE1, SLC47A1) inhibitors through prescription drug profiling and computational modeling. J Med Chem. 2013;56(3):781–95.
Masuda S, et al. Identification and functional characterization of a new human kidney-specific H+/organic cation antiporter, kidney-specific multidrug and toxin extrusion 2. J Am Soc Nephrol. 2006;17(8):2127–35.
Ruffolo RR, Feuerstein GZ. Carvedilol case history: the discovery and development of the first beta-blocker for the treatment of congestive heart failure. Expert Opin Drug Discovery. 2006;1(1):85–9.
Morgan T. Clinical pharmacokinetics and pharmacodynamics of carvedilol. Clin Pharmacokinet. 1994;26(5):335–46.
Stahl E, et al. Carvedilol stereopharmacokinetics in rats: affinities to blood constituents and tissues. Arch Pharm (Weinheim). 1993;326(9):529–33.
Nakamichi N, et al. Involvement of carnitine/organic cation transporter OCTN1/SLC22A4 in gastrointestinal absorption of metformin. J Pharm Sci. 2013;102(9):3407–17.
Wu X, et al. Structural and functional characteristics and tissue distribution pattern of rat OCTN1, an organic cation transporter, cloned from placenta. Biochim Biophys Acta. 2000;1466(1–2):315–27.
Higgins JW, Bedwell DW, Zamek-Gliszczynski MJ. Ablation of both organic cation transporter (OCT)1 and OCT2 alters metformin pharmacokinetics but has no effect on tissue drug exposure and pharmacodynamics. Drug Metab Dispos. 2012;40(6):1170–7.
Li Q, et al. Deficiency of multidrug and toxin extrusion 1 enhances renal accumulation of paraquat and deteriorates kidney injury in mice. Mol Pharm. 2011;8(6):2476–83.
Carvalho Rodrigues MA, et al. Carvedilol efficiently protects kidneys without affecting the antitumor efficacy of cisplatin in mice. Chem Biol Interact. 2013;206(1):90–9.
Carvalho Rodrigues MA, et al. Carvedilol protects against apoptotic cell death induced by cisplatin in renal tubular epithelial cells. J Toxic Environ Health A. 2012;75(16–17):981–90.
Rodrigues MA, et al. Carvedilol protects against cisplatin-induced oxidative stress, redox state unbalance and apoptosis in rat kidney mitochondria. Chem Biol Interact. 2011;189(1–2):45–51.
Ruffolo RR Jr, Feuerstein GZ. Pharmacology of carvedilol: rationale for use in hypertension, coronary artery disease, and congestive heart failure. Cardiovasc Drugs Ther. 1997;11(Suppl 1):247–56.
Bachmakov I, et al. Interaction of beta-blockers with the renal uptake transporter OCT2. Diabetes Obes Metab. 2009;11(11):1080–3.
Acknowledgments and Disclosures
The present study was supported by the National Institute of General Medical Sciences of the US National Institutes of Health (NIH) [R01GM099742] and by the US Food and Drug Administration (FDA) [U01FD004320]. Dr. Dong Guo is an M-CERSI Scholar (FDA 1U01FD005946). Dr. Yan Shu is a co-founder for and owns equity in Optivia Biotechnology.
Author information
Authors and Affiliations
Contributions
Designed Research: Yan Shu, James E. polli, Dong Guo.
Performed Research: Dong Guo, Hong Yang, Qing Li, Hyo Jung Bae, Sujuan Zeng, Tong Su.
Analyzed Data: Dong Guo, Hong Yang, Qing Li, Hyo Jung Bae, Yan Shu.
Wrote Manuscript: Dong Guo, Yan Shu.
Corresponding author
Rights and permissions
About this article

Cite this article
Guo, D., Yang, H., Li, Q. et al. Selective Inhibition on Organic Cation Transporters by Carvedilol Protects Mice from Cisplatin-Induced Nephrotoxicity. Pharm Res 35, 204 (2018). https://doi.org/10.1007/s11095-018-2486-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11095-018-2486-2