
Abstract
d-Serine has been shown to play an important role in the expression and control of a variety of brain functions by acting as the endogenous coagonist for the N-methyl-d-aspartate type glutamate receptor (NMDAR), at least, in the forebrain. To obtain further insight into the still debatable cellular localization of the d-amino acid, we have examined the effects of the selective destruction of the neuronal cell bodies by quinolinate on the tissue or extracellular d-serine concentrations in the medial prefrontal cortex of the rat. A local quinolinate infusion into the bilateral medial prefrontal cortex produced a cortical lesion with a marked (− 65%) and non-significant alteration (− 5%) in the cortical and striatal tissue d-serine concentrations, respectively, 7 days post-infusion. In vivo microdialysis experiments in the right prefrontal lesion site 9 days after the quinolinate application revealed that the basal extracellular d-serine levels were also dramatically reduced (− 64%). A prominent reduction in the tissue levels of GABA in the interneurons of the prefrontal cortex (− 78%) without significant changes in those in the striatum (+ 12%) verified that a major lesion part was confined to the cortical portion. The lack of a significant influence of the prefrontal quinolinate lesion on its dopamine concentrations in the mesocortical dopamine projections suggests that the nerve terminals and axons in the lesion site may be spared. These findings are consistent with the perikarya-selective nature of the present quinolinate-induced lesion and further support the view that neuronal cell bodies of intrinsic neurons in the prefrontal cortical region contain substantial amounts of d-serine, which may sustain the basal extracellular concentrations of d-serine.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A body of evidence has been accumulated indicating that d-serine plays a pivotal role in the physiological functioning of the N-methyl-d-aspartate type glutamate receptor (NMDAR) as the endogenous coagonist for the receptor by binding to its glycine modulatory site, at least, in the forebrain areas of mammals [1, 2]. Thus, selective depletion of d-serine by application of d-amino acid oxidase leads to a remarkable inhibition of the synaptic current via stimulation of the NMDAR in cultured hippocampal neurons [3] and hippocampal slices [4] even in the presence of normal tissue levels of glycine that is another coagonist for the NMDAR. Moreover, no significant induction of long-term potentiation or a dramatic attenuation in nitric oxide formation by activation of the NMDAR [5] has been found in mice lacking a d-serine synthesizing enzyme, i.e., serine racemase, which display a marked decrease in brain tissue and extracellular concentrations of d-serine without significant changes in those of glycine [5,6,7]. Finally, it has been demonstrated by using an in vivo microdialysis technique that the diminished basal extracellular concentrations of d-serine under the normal glycine levels result in the attenuation of the NMDAR-mediated increasing response of taurine release in the hippocampus of the mice with CaMKII-conditional depletion of serine racemase in the forebrain neurons [8] and in the rat medial prefrontal cortex locally perfused by a selective antagonist for the GABAA receptor, muscimol [9].
Since the NMDAR has been shown to be one of the key molecules that regulate the higher order brain functions, and since its dysfunction has been implicated in the pathophysiology of neuropsychiatric disorders including schizophrenia, Alzheimer’s disease, and cerebrovascular diseases, to clarify the cellular and molecular mechanisms underlying brain d-serine signaling is crucial to understand and develop a novel therapeutic approach to various pathological conditions of the brain [1]. Together with the close positive relationship between the brain regional distribution patterns of d-serine and those of the binding density of glutamate, glycine and phencyclidine sites of the NR1/NR2 heteromeric NMDARs [1, 10], the above intrinsic coagonist nature of d-serine indicates the possible existence of d-serine in synapse-related structures. However, the exact types and setups of cells containing d-serine and the associated molecular machinery are still inconclusive and await further elucidation [2, 11, 12]. In fact, the extracellular d-serine concentrations in the rodent brain have been reported to be modified by neuronal and glial activities [9, 13,14,15,16], suggesting the plausible localization of d-serine in neurons and glia.
Results obtained from immunohistochemical studies using the anti-d-serine antibody recognizing d-serine conjugated to glutaraldehyde or paraformaldehyde are disputed among investigators. Schell et al. [17] first showed a d-serine-like immunoreactivity predominantly in astroglia while the subsequent experiments [18, 19] drew the conclusion that d-serine was contained not only in the astroglia but also in the neurons. Williams et al. [20] found immunostaining of d-serine in vesicle-like compartments in the astroglia and radial glial cells and a subset of glutamate neurons. More recent examinations pointed out that the immunoreactivity of d-serine and serine racemase was observed in the neurons, but not in the astroglia [21, 22], and that, coincident with the histological data, neuron-specific, but not astroglia-specific, the deletion of serine racemase produced a significant decrease in the tissue and/or extracellular d-serine concentrations in the hippocampus [6, 18].
Despite these discrepancies, the localization of d-serine has not been verified by any method other than immunohistochemistry. To obtain further neuroanatomical information about d-serine for understanding these discrepancies, we have investigated the effects of intra-medial prefrontal infusion of the excitotoxin, quinolinate, which is well established to selectively destroy the cell bodies of neurons [23], on the tissue and extracellular concentrations of d-serine in the prefrontal cortex of the rat. We have also compared the influences among various amino acids that are related to the NMDAR and/or well known to be contained in the perikarya of the intrinsic or projecting neurons in the cortical area.
Methods
Approval for Animal Experiments
All experimental procedures in this study were performed in strict accordance with the guidance of the National Institute of Neuroscience, National Center of Neurology and Psychiatry (NCNP), Tokyo Medical and Dental University (TMDU), and Showa University and were approved by the Animal Investigation Committees of the respective institutes: Committee for Ethical Issues in Animal Experiments (NCNP); Institutional Animal Care and Use Committee (TMDU); Laboratory Animal Science Committee (Showa University). At Showa University, we conducted a statistical examination of the quantitative measurement data of amino acids and monoamines and microscopic analysis of tissue preparations, which were obtained from the animal experiments.
Animals
Male Wistar rats (ST strain, Clea Japan, Tokyo, Japan) on postnatal day 56 at the time of injection of the excitotoxin quinolinate were used. The animals were housed at 22 ± 0.5 °C and 50% humidity under a controlled dark–light cycle (light: 08.00–20.00 h) and had free access to food and water.
Chemicals
Quinolinate and (−)-bicuculline methiodide were purchased from the Wako Pure Chemical Corporation, Osaka, Japan (presently FUJIFILM Wako Pure Chemical Corporation at present) and Tocris Bioscience (USA), respectively. All other chemicals and drugs were of ultrapure quality and commercially available.
Chemical Lesion
Procedures for the quinolinate lesions were performed in principle according to the method of Schwarcz et al. [23] Quinolinate (240 nmol/2.0 µl, dissolved in 0.1 M phosphate-buffered saline, pH 7.4) or vehicle was bilaterally infused into the medial prefrontal cortex in a volume of 2 µl/side for 10 min at the following coordinate: medial prefrontal cortex, A + 3.2 mm, V + 5.2 mm, L ± 0.6 mm (the atlas of Paxinos and Watson [24]). The injection canula was left in place for a further 5 min before removal. The tissue concentrations of the amino acids were determined 7 days after the infusion. This date was chosen because quinolinate-induced lesions have been reported to reach a maximum before 7 days post-infusion and remain mostly unchanged thereafter [25,26,27].
Histological and Immunohistochemical Examination
The histological and immunohistochemical examinations of the present quinolinate-induced lesion were achieved on brain sections prepared according to our previous report [28], as described below. Under pentobarbital anesthesia (40 mg/kg, i.p.), the rats were transcardially perfused with physiological saline followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4). The whole brains were removed, post-fixed and coronally cut at 60 µm after cryoprotection by transfer in different concentrations of sucrose solution and freezing.
To evaluate the extent of the quinolinate-induce lesion, brain coronal sections obtained from frozen brains were mounted on slides and stained with cresyl violet (Chroma, Co., Ltd: Nissl staining) as already mentioned [28, 29]. Cresyl violet was dissolved in purified water to which an appropriate amount of 10% acetate was dropwise added, and used for tissue staining at a concentration of 0.1%.
For analyses of the types and distribution of damaged cells in the quinolinate-lesioned portions, we conducted immunohistochemical studies to detect various neuronal and glial cells by applying the following specific antibodies against their respective marker proteins which were purchased from Abcam (Cambridge, UK): (1) neuronal nuclei/RNA-binding Fox3 (NeuN/Rbfox3; the neuronal nucleus marker [30] (rabbit monoclonal; diluted to 1:100; ab177487), (2) glial fibrillary acidic protein (GFAP; the astroglia marker [25, 26, 31] (goat polyclonal; 1:1000; ab53554), (3) myeline basic protein (MBP: the oligodendroglia marker [32] (rabbit monoclonal; 1:5000, rabbit monoclonal; ab218021).
Immunohistochemistry was achieved on PFA-fixed frozen coronal sections embedded on microscope glasses. Following 60-min rinses (incubation) in the blocking agent, Blocking One Histo (NAKARAI TESQUE, INC. Kyoto, Japan), at room temperature, brain slices were incubated for 12 h at 4 °C in 0.01 M PBS containing 0.2% Tween-20 (PBST) and each antibody previously mentioned. After three 5-min washes in 0.01 M PBST, the brain sections were incubated for 1 h in a secondary antibody, an Alexa Fluor-conjugated goat anti-rabbit IgG ((# A-11029, Thermo Fisher Scientific Co. Ltd., Massachusetts, USA), an Alexa Fluor-conjugated goat anti-mouse IgG (Vector Labs), or a FITC (Fluoresceinisothiocyanate isomer-I)-conjugated rabbit anti-goat IgG (CiteAb, Bath, UK). The stained sections were washed 3 times with PBST and mounted onto subbed slides by using an encapsulant for fluorescent staining, Fuluoromount (DBS Diagnostic BioSystems, California, USA).
Brain Dissection
The animals were killed by cervical dislocation 7 days after the local infusion of quinolinate into the bilateral medial prefrontal cortex. The right and left medial prefrontal cortex including the lesioned area (dopamine-rich portion according to Björklund and Lindvall [33] (A + 4.2 to + 2.5 mm; the atlas of Paxinos and Watson [24]) and right anterior striatum (A + 2.4 to + 0.0 mm) were dissected out in the cold, immediately frozen on dry ice and stored at − 80 °C until quantitative measurements of the amino acids and monoamines.
In Vivo Microdialysis
An in vivo brain microdialysis method was employed to monitor the extracellular concentrations of d-serine in the medial prefrontal cortex of the freely moving rats as previously depicted [9, 34, 35]. On the 8th day of the bilateral prefrontal injection of quinolinate, the rats were anesthetized with pentobarbital (40 mg/kg, intraperitoneally) and mounted on a stereotaxic frame. Straight-shaped cellulose dialysis tubing (3.0 mm length, 0.16 mm internal diameter, molecular weight cutoff of 50,000, EICOM Co., Ltd., Japan) was then implanted into the right medial prefrontal cortex (A + 3.2 mm, V + 5.2, L ± 0.6; the atlas of Paxinos and Watson [24]).
Two days after the operation, the dialysis probe was perfused with a Ringer solution (NaCl, 147 mM; KCl, 4 mM; CaCl2, 1.3 mM; pH 7.4) at the flow rate of 2 µl/min. Following stabilization for at least 80 min, the dialysate samples were serially collected every 20 min. The first four samples were determined as the basal concentrations of D-serine and other amino acids, and the average of the amino acid measurements of the first two samples was used for the statistical analysis. After the experiments were completed, the accuracy of the position of the dialysis probe was macroscopically confirmed in each case on 150-µm-thick serial coronal sections.
We did not correct for the d-serine concentrations in the dialysate by the recovery rate of the dialysis probe because (1) our previous experiments have shown that its variability is very low when tested in vitro [13] and (2) it is not possible to determine the exact in vivo recovery rate in each animal.
Quantitative Analysis of Amino Acids
The simultaneous determination of the free amino acid enantiomers and non-chiral amino acids in the brain tissues and dialysates was accomplished by high performance liquid chromatography (HPLC) with fluorometric detection as previously described [9, 36]. Briefly, the brain tissue sample was homogenized in 10 volumes of 4% trichloroacetic acid (TCA) after adding d-homocysteic acid as the internal standard, and the homogenate was centrifuged at 14,500×g for 20 min at 4 °C. The supernatant and the collected dialysates following the addition of the internal standard were stored at − 80 °C until derivatization. When conducting an assay of the samples, each tissue supernatant or an aliquot of each dialysate was derivatized with N-tert-butyloxycarbonyl-l-cystein (Boc-l-Cys) and o-phthaldialdehyde for 2 min at room temperature. The amino acid derivatives were subsequently applied to the HPLC system and separated on a 4-µm Nova-Pak C18 column (300 × 3.9 mm, I.D., Waters, Japan) using the following mobile phases at the constant flow rate of 0.9 ml/min at 30 °C. Mobile phase A was 0.1 M acetate buffer (pH 6.0), including 12% acetonitrile, and mobile phase B was the acetate buffer with 20% acetonitrile. The separation procedure was accomplished with a linear gradient from mobile phase A to B in 50 min, and the isolated fluorescent derivatives were detected by an FP-2025 spectrofluorometer (Jasco International Co., Ltd., Japan). The excitation and emission wavelengths were 344 and 443 nm, respectively.
Quantitative Analysis of Monoamines
Dopamine in the brain tissues were determined by reverse-phase HPLC with electrochemical detection, as previously described [37]. Briefly, after the addition of 3,4-dihydroxyhydro-cinnamic acid (DHHCA) as an internal standard, the cortical tissues were homogenized in 0.1 M perchloric acid containing 2 mM EDTA and 4 mM sodium pyrosulfate, and the homogenates were spun at 8800×g for 20 min at 4 °C. The pH of the separated supernatant was optimized by 1 N sodium acetate and stored at − 80 °C until use. Dopamine and other monoamines and their metabolites were separated on a reverse-phase column filled with 5C18 (octadesil) using 0.1 M acetate-citrate buffer (pH 4.1) containing 15% methanol, 0.7 mM octasulfonic acid and 0.1 mM EDTA at the flow rate of 0.6 ml/min. The ECD was accomplished using a carbon graphite working electrode set at + 0.7 V.
Statistics
The results represent the mean with SEM of the data. Statistical differences of the independent data between the two groups were evaluated using the unpaired two-tailed Wilcoxon rank sum test (the homogeneous and heterogeneous variances for each experimental group).
Results
In the prefrontal areas of the quinoliinate-induced lesion, we determined the tissue concentrations of the amino acids metabolically or functionally linked with D-serine and/or the NMDAR: (1) d-Serine and glycine acting at the NMDAR glycine site [1], (2) NMDAR glutamate site agonists, l-glutamate and l-aspartate, and their respective metabolites, l-glutamine and l-aspargine [36], (3) l-arginine as a precursor for nitric oxide as a NMDAR modulator [36], (4) a precursor of d-serine, l-serine, [1] and (5) inhibitory amino acids, GABA and taurine, interacting with d-serine [9]. GABA [38] and l-glutamate [39] are also well documented to be useful markers for cell bodies of cortical interneurons and frontostriatal glutamate neuronal pathways, respectively. The dopamine levels were quantified because the prefrontal dopamine is considered to be a marker for the nerve terminals and axons in the medial prefrontal cortex.
Histological Examination of Extent of Quinolinate-Induced Lesion in the Medial Prefrontal Cortex of the Rat
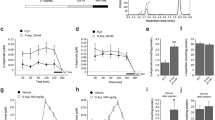
As shown in the Fig. 1a and d, our Nissl staining validated that degeneration 7 days after quinolinate injection was observed in the bilateral prelimbic, infralimbic, cingulate (Cg1) and dorsal peduncular (DP) cortices and parts of the secondary motor cortex around the injection sites, at least, at the levels from 4.2 mm to 2.5 anterior to the bregma (Paxinos and Watson 1986) [24]. There were no evident lesions in the primary motor cortex, the cortical regions lateral to the motor cortex (M1 and M2), the striatum (STR), and the white matter including the forceps minor (fmi) and the corpus callosum (ac) (Fig. 1a–c). Figure 1b depicts a photomicrograph of high-magnification image of a boundary region between a lesion and non-lesioned site, which is surrounded by the dotted line in Fig. 1a. No apparent degeneration was detected in the prefrontal section of the control animals treated with an intra-medial frontal cortex injection of vehicle (Fig. 1c).

Histological examination of the extent of quinolinate-induced lesion in the medial prefrontal cortex of the rat. Stereomicrographs and light photomicrographs display representative cresyl violet-stained brain coronal sections including quinolinate-lesioned area at the level of the medial prefrontal cortex of the rat (approximately 3.7 mm (a, b, and c) or 2.5 (d) anterior to the bregma (Paxinos and Watson 1986) [24]) 7 days after local infusion of the quinolinate (a, b and d) or vehicle (PBS: controls) (c) into the cortical region. b A high magnification image of a boundary area between the degenerated and undegenerated site, which is surrounded by the dotted line in a. e and f Schematically represent an extent of the quinolinate lesion in the medial prefrontal cortex in each of 5 animals by dotted, short-dashed, long-dashed, dot-dashed or double line (adapted from the figures at 3.72 mm (e) and 2.52 mm (f) anterior to the Bregma of the Reference [24]). The striped areas surrounded by solid line in these panels illustrate the respective prefrontal cortical tissues sampled for the amino acid quantification. ac anterior commissure; Cg1 cingulate area 1; DP dorsal peduncular cortex; fmi forceps minor; IL infralimbic cortex; M1 primary motor cortex; M2 secondary motor cortex; NA nucleus accumbens; OB orbital cortex; PL prelimbic cortex; STR striatum
Some variations in the extent of the neurotoxin-induced destruction area were observed among the 5 animals in which histological and immunohistochemical evaluation were performed (Fig. 1e and f). The tissue in the striped area depicted in Fig. 1e and f was dissected so that the smallest degradation area could be sampled.
Immunohistochemical Analysis of Quinolinate-Induced Lesion in the Medial Prefrontal Cortex of the Rat
Immunoreactivity to antibodies against the neuronal nuclear marker protein, NeuN, was verified to be dense in the neuron-predominant gray matter and sparse in the glia-predominant white matter [30] at the prefrontal coronal sections of vehicle-treated control animals (Fig. 2a). In the quinolinate-induced lesion in the cingulate, prelimbic and infralimbic cortex which was evaluated by cresyl violet staining (Fig. 1a, b and d), there was a marked decay in the intensity and density of the NeuN immunohistochemical staining (Fig. 2b and d). In contrast, the GFAP (Fig. 3b and d) and MBP (Fig. 3e and h) immunoreactivity was found to be enhanced and more densified as compared to vehicle-injected control animals (Fig. 3a and c). The medial prefrontal immunostaining of MBP in the vehicle treated animals was exclusively detected in the white matter, but not the gray matter (Fig. 3e), whereas the quinolinate-lesioned portion in the cortical gray matter clearly exhibited the MBP immunoreactivity (Fig. 3f). Incubation of the brain sections without the respective primary antibodies produced no immunostaining (data not shown).

Immunohistochemical analysis of a neuronal marker protein in quinolinate-induced lesion in the medial prefrontal cortex of the rat. Photomicrographs indicate representative brain coronal sections showing the immunoreactivity of NeuN (a and b) at the level of the anterior portion of the medial prefrontal cortex (approximately 2.7–3.0 mm anterior to the bregma (Paxinos and Watson 1986) [24]) of the rat treated with local infusion of quinolinate (b and d) or vehicle (PBS: controls) (a and c) into the cortical region. c and d Display a high-magnification image of NruN immunorstaining in the prelimbic cortex medial to the forceps minor (see Fig. 1a–d). Cg1 cingulate area 1; CL clastrum; E/OV ependymal and subependymal layer/olfactory ventricle; fmi forceps minor; IL infralimbic cortex; PL prelimbic cortex. Scale bars: 500 µm in a and b; 100 µm in c and d

Immunohistochemical analysis of glial marker proteins in quinolinate-induced lesion in the medial prefrontal cortex of the rat. Photomicrographs display representative brain coronal sections exhibiting the immunoreactivity of the astroglia marker, GFAP (a–d), and the oligodendroglia marker, MBP at the level of the anterior portion of the medial prefrontal cortex of the rat treated with local infusion of quinolinate (b, d, f and h) or vehicle (PBS: controls) (a, c, e and g) into the cortical region. e–h High-magnification images of the GFAP and MBP immunorstaining in the prelimbic cortex medial to the forceps minor. j and k Schematically represent the respective photographic locations of the GFAP and MBP immunoreactivity as open squares for a, b, e and f, and as fills for c, d, g and h [adapted from the figures at 3.72 mm anterior to the Bregma of the Reference [24] (Paxinos and Watson 1986)]. i Notes the name of each brain region. Cg1 cingulate area 1; fmi forceps minor; IL infralimbic cortex; PL prelimbic cortex. Scale bars in this figure are 100 µm
Effects of Quinolinate-Induced Lesion in the Medial Prefrontal Cortex on the Tissue Concentrations of d-Serine, l-Serine, l-Glutamate, Glycine and GABA in the Cortical Portion of the Rat
Local infusion of quinolinate into the bilateral medial prefrontal cortex caused a significant decrease in the cortical tissue concentrations of d-serine (− 65%), l-glutamate (− 59%), GABA (− 78%), glycine (− 49%), l-serine (− 29%), l-aspartate (− 48%), l-aspargine (− 59%), l-glutamine (− 41%) and taurine (− 28%) to different extents (Fig. 4 and Table 1) 7 days thereafter. In contrast, there was an increase in those of l-arginine (+ 59%) (Fig. 4).

Tissue concentrations of d-serine, l-glutamate, GABA, and l-arginine in the medial prefrontal cortex of the rats 7 days after local infusion of quinolinate or vehicle into the cortical portion. The results are means with S.E.M of data obtained from 8 animals and are expressed as a percentage of the following absolute control values (µmol/g of wet weight): d-serine (d-Ser), vehicle (phosphate buffered saline)-infused controls (Veh) 0.20 ± 0.015, Quinolinate (QA) 0.069 ± 0.0072***, P = 0.00094; l-glutamate (l-Glu), Veh 13 ± 0.46, QA 5.5 ± 0.51***, P = 0.00094; GABA, Veh 1.7 ± 0.055, QA 0.38 ± 0.078***, P = 0.00094; l-arginine (l-Arg), Veh 0.15 ± 0.0073, QA 0.24 ± 0.018***, P = 0.00094. ***P < 0.001 as compared to vehicle-infused controls
Effects of Quinolinate-Induced Lesion in the Medial Prefrontal Cortex on the Tissue Concentrations of d-Serine, l-Serine, l-Glutamate, Glycine and GABA in the Anterior Striatum of the Rat
The bilateral intra-medial prefrontal cortex injection of quinolinate failed to modify the tissue concentrations of d-serine, l-serine, l-aspartate, l-asparagine, l-glutamine, l-arginine and GABA in the combined right and left anterior striatum 7 days post-injection (Fig. 5 and Table 2). In contrast, there was a small but significant diminution in the striatal tissue levels of l-glutamate (− 11%) (Fig. 5), glycine (− 17%) and taurine (Table 2).

Tissue concentrations of d-serine, l-glutamate, GABA, and l-arginine in the anterior striatum of the rats 7 days after local infusion of quinolinate or vehicle into the cortical portion. The results are means with S.E.M of data obtained from 8 animals and are expressed as a percentage of the following absolute control values (µmol/g of wet weight): d-serine (d-Ser), vehicle (phosphate buffered saline)-infused controls (Veh) 0.22 ± 0.0051, Quinolinate (QA) 0.21 ± 0.0031, P = 0.13; l-glutamate (l-Glu), Veh 12 ± 0.072, QA 10 ± 0.33*, P = 0.018; GABA, Veh 2.1 ± 0.063, QA 2.4 ± 0.13, P = 0.10; l-arginine (l-Arg), Veh 0.18 ± 0.0038, QA 0.21 ± 0.011*, P = 0.10. *P < 0.05 as compared to the vehicle-infused controls
Effects of Quinolinate-Induced Lesion in the Medial Prefrontal Cortex on the Extracellular Concentrations of d-Serine Under the Basal Condition
In vivo microdialysis experiments revealed that there was a marked reduction in the basal extracellular concentrations of d-serine (-67%) in the medial prefrontal cortex 9 days after the bilateral local administration of quinolinate into the cortical area (Fig. 6).

Extracellular concentrations of d-serine in the right medial prefrontal cortex of the rat under perfusion of a Ringer solution 7 days after bilateral local infusion of quinolinate or vehicle into the cortical portion. The results are means with S.E.M of data obtained from 4 animals and are expressed as a percentage of the following absolute control values (µM): d-serine (d-Ser), vehicle (phosphate buffered saline)-infused controls (Veh) 0.46 ± 0.038, Quinolinate (QA) 0.15 ± 0.025, P = 0.029. *P < 0.05 as compared to the vehicle-infused controls
Effects of Quinolinate-Induced Lesion in the Medial Prefrontal Cortex on the Tissue Concentrations of Dopamine in the Cortical Portion of the Rat
To confirm the survival of nerve terminals and fibers in the quinolinate-lesioned medial prefrontal cortex, dopamine concentrations were measured at the lesion area where these parts of the mesocortical dopamine neurons were present. No significant changes in the cortical tissue dopamine concentrations were detected in the lesioned area (Fig. 7).

Tissue concentrations of dopamine in the medial prefrontal cortex of the rats 7 days after local infusion of quinolinate or vehicle into the cortical portion. The results are means with S.E.M of data obtained from 7–8 animals and are expressed as a percentage of the following absolute control values (nmol/g of wet weight): dopamine, vehicle (phosphate buffered saline)-infused controls (Veh) 0.38 ± 0.038, Quinolinate (QA) 0.41 ± 0.056. No significant changes were detected as compared to the vehicle-infused controls (P = 0.54)
Discussion
This study is the first to reveal that the excitotoxin-induced lesion in the medial prefrontal cortex, which is rich in d-serine, yields a drastic reduction in the tissue d-serine concentrations in the lesioned area of mammalian brains. In the prefrontal portion, the magnitude of the reduction (− 65%) is approximately twice that of l-serine (− 29%), and is slightly higher and lower than those of l-glutamate (− 59%) and GABA (− 78%), respectively. The extracellular d-serine levels also markedly declined by the lesion in the cortical portion (− 64%). The different percentages of decreases in the tissue levels of the respective amino acids and the increased l-arginine concentrations (Fig. 4) are likely to exclude the possibility that the remarkable down-regulation of the tissue d-serine concentrations of could be solely due to a non-specific effect of the quinolinate lesion.
The present histological examination using cresyl violet staining indicated that the quinolinate-induced lesion was confined in neuron-rich gray matter, but not in the glia-rich white matter, of the medial prefrontal and anterior cingulate cortical regions. Furthermore, the immunocyochemical analyses revealed a dramatic loss of the immunostaining of the neuronal nucleus protein marker, NeuN, and augmented immunoreactivity of the marker proteins for astroglia (GFAP) and oligodendroglia (MBP) in the neurotoxin lesion area.
These data confirm the previous immunohistochemical observations of neuronal [30] and astroglial [25, 26, 31] marker proteins in the quinolinate-induced lesion. Although there is so far no experiment to evaluate the influences of the quinolinate lesion on the MBP, mature oligodendroglia expressing this protein has been shown to be resistant to another excitotoxin kainite [32]. Moreover, our findings do not contradict the report of oligodendroglia proliferation at the quinolinic acid-injected brain site [40]. Taken together, our histological and immunohistochemical results validate the concept that the present quinolinate infusion caused the selective destruction of neurons without any obvious loss of the astroglia and oligodendroglia.
As previously described [23, 41,42,43], the neuron-preferred nature of the quinolinate-produced brain tissue destruction is also supported by the biochemical data that the neurotoxin application caused a prominent decrease in the prefrontal levels of GABA (Fig. 4) that has been to shown to be predominantly present in the cortical interneurons [41]. The plausibility that the neurotoxin-induced destruction extends to a large brain region other than the prefrontal region can be denied by the fact that the concentrations of GABA in the intrinsic neurons [44] were not altered in the striatum (Fig. 4). The neurotoxin has been well established to mainly degenerate the perikarya with sparing axons and terminals in the neurons [23]. This feature seems to be affirmed by no significant changes in the prefrontal tissue concentrations of dopamine (Fig. 4) that is present in the fibers and terminals of the ascending dopaminergic neurons projecting from the midbrain to the medial prefrontal cortex [45]. These data are consistent with the concept that d-serine may be substantially contained in the neurons in the mammalian cerebral cortex.
Consequently, the quinolinate-induced salient decline in the tissue d-serine concentrations in the medial prefrontal cortex indicates that d-serine may, at least, be located in the neuronal cell bodies of the brain region. These observations are in line with the previous detection of immunoreactivity of d-serine [18, 19, 21, 22] and serine racemase [46] in the somas of the neocortical neurons. Also, the experiments showing that serine racemase may be exported from the neuronal cell nucleus to the cytosol [47] fits with the concept that the excitotoxin could deplete the prefrontal d-serine by destroying these portions of the neuronal somas for d-serine production and storage.
In terms of astrogliosis occurring in and around the quinolinate injection site [25, 48] and an increase in the d-serine levels in such reactive astroglia following controlled cortical impact injury [43], the present cortical quinolinate lesion is expected to augment the d-serine concentrations in the medial prefrontal cortex. However, conversely, the resulting decrease (Fig. 4 and Table 1) is considered to be due to the large loss of nerve cell bodies containing d-serine synthase, i.e., serine racemase [21, 22, 46]. In fact, many neuronal cell bodies persisted in tissues after controlled cortical impact injury, where an increase in the astroglia and d-serine levels was observed [49]. Alternatively, differences in the subpopulations of the reactive astroglia depending on the types of lesions may explain the above discrepancies as Complement-3a-positive A1-type reactive astroglia was documented to synthesize d-serine during the cell culture preparation [50].
The striking reduction in the d-serine concentrations in the extracellular fluid (Fig. 6) is parallel to that seen in the tissue in the prefrontal quinolinate lesion site. The close correlation between the extracellular and tissue levels of d-serine in the brain is similarly noted under physiological conditions [6, 13]. Therefore, the tissue and extracellular alterations could be commonly ascribed to disruption of the neuronal cell bodies that include serine racemase [46].
The unchanged striatal concentrations of d-serine (Fig. 5) after the removal of the prefrontal perikarya by quinolinate substantiate that the putative prefrontal d-serine containing neurons do not send axons to the anterior striatum and may be interneurons. This idea is based upon the following results: (1) The degeneration procedure replicated the previously established decreasing effects of the prefrontal ablation or chemical destruction [39, 51, 52] on the striatal tissue glutamate amounts to corroborate the fronto-striatal projections of glutamate neurons, and (2) the striatal tissue levels of GABA entirely localized in the interneurons in the prefrontal portion [38] were unaffected by the prefrontal quinolinate lesion.
Interestingly, the chemical destruction of the prefrontal neuronal cell bodies produced distinct influences on another coagonist for the NMDAR, glycine, in the prefrontal and striatal regions. Its tissue concentrations were lessened in both the prefrontal lesion site and anterior striatum (Figs. 4 and 5), which indicated the possible presence of the unidentified prefronto-striatal glycinergic neurons and/or of prefrontal modification of the striatal glycine levels. These observations may be related to the differences in the mode of neuroanatomical localization and/or regulation of the two NMDAR coagonists, d-serine and glycine, in the brain [1, 12]. In addition, the newly observed reduction in the tissue concentrations of taurine in the two brain regions after the prefrontal neuronal perikaryal lesion (Tables 1 and 2) suggests the potential taurine-containing pathways from the prefrontal area to the anterior striatum and/or indirect tuning by the prefrontal cortex of the striatal taurine dynamics. d-Serine could participate in the regulation of the taurine metabolism by the NMDAR through the interaction with the GABAergic system [9].
In conclusion, the present findings from the experiments using the neurotoxin quinolinate provide further evidence for the view that d-serine is, at least, present in the cell bodies of the intrinsic neurons in the medial frontal cortex of the rat. These putative d-serine-containing neurons appear to be involved in the basal-evel maintenance of the extracellular d-serine concentrations for physiological activation of the NMDAR.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author by reasonable request.
References
Nishikawa T (2011) Analysis of free d-serine in mammals and its biological relevance. J Chromatogr B 879:3169–3183. https://doi.org/10.1016/j.jchromb.2011.08.030
Wolosker H, Balu DT, Coyle JT (2016) The rise and fall of the d-serine-mediated gliotransmission hypothesis. Trends Neurosci 39:712–721. https://doi.org/10.1016/j.tins.2016.09.007
Mothet JP, Parent AT, Wolosker H, Brady RO Jr, Linden DJ, Ferris CD, Rogawski MA, Snyder SH (2000) d-Serine is an endogenous ligand for the glycine site of the N-methyl-d-aspartate receptor. Proc Natl Acad Sci USA 97:4926–4931. https://doi.org/10.1073/pnas.97.9.4926
Yang Y, Ge W, Chen Y, Zhang Z, Shen W, Wu C, Poo M, Duan S (2003) Contribution of astrocytes to hippocampal long-term potentiation through release of d-serine. Proc Natl Acad Sci USA 100:15194–15199. https://doi.org/10.1073/pnas.2431073100
Basu AC, Tsai GE, Ma CL, Ehmsen JT, Mustafa AK, Han L, Hashimoto K (2009) Targeted disruption of serine racemase affects glutamatergic neurotransmission and behavior. Mol Psychiatry 14:719–727. https://doi.org/10.1007/s10571-014-0027-z
Horio M, Kohno M, Fujita Y, Ishima T, Inoue R, Mori H et al (2011) Levels of d-serine in the brain and peripheral organs of serine racemase (Srr) knock-out mice. Neurochem Int 59:853–859. https://doi.org/10.1016/j.neuint.2011.08.017
Labrie V, Fukumura R, Rastogi A, Fick LJ, Wang W, Boutros PC, Kennedy JL, Semeralul MO, Lee FH, Baker GB, Belsham DD, Barger SW, Gondo Y, Wong AH, Roder JC (2009) Serine racemase is associated with schizophrenia susceptibility in humans and in a mouse model. Hum Mol Genet 18:3227–3243. https://doi.org/10.1093/hmg/ddp261
Ishiwata S, Umino A, Balu DT, Coyle JT, Nishikawa T (2015) Neuronal serine racemase regulates extracellular d-serine levels in the adult mouse hippocampus. J Neural Transm (Vienna) 122:1099–1103. https://doi.org/10.1007/s00702-015-1388-2
Umino A, Ishiwata S, Iwama H, Nishikawa T (2017) Evidence for tonic control by the GABAA receptor of extracellular d-serine concentrations in the medial prefrontal cortex of rodents. Front Mol Neurosci 10:240. https://doi.org/10.3389/fnmol.2017.00240
Hashimoto A, Nishikawa T, Oka T, Takahashi K (1993) Endogenous d-serine in rat brain: N-methyl-d-aspartate receptor-related distribution and aging. J Neurochem 60:783–786. https://doi.org/10.1111/j.1471-4159.1993.tb03219.x
Wolosker H, Balu DT, Coyle JT (2017) Astroglial versus neuronal d-serine: check your controls. Trends Neurosci 40:520–522. https://doi.org/10.1016/j.tins.2017.06.010
Papouin T, Henneberger C, Rusakov DA, Oliet SHR (2017) Astroglial versus neuronal d-serine: fact checking. Trends Neurosci 40(9):517–520. https://doi.org/10.1016/j.tins.2017.05.007
Hashimoto A, Oka T (1995) Extracellular concentration of endogenous free d-serine in the rat brain as revealed by in vivo microdialysis. Neuroscience 66(3):635–643. https://doi.org/10.1016/0306-4522(94)00597-x
Henneberger C, Papouin T, Oliet SH, Rusakov DA (2010) Long-term potentiation depends on release of d-serine from astrocytes. Nature 463:232–236. https://doi.org/10.1038/nature08673
Ishiwata S, Umino A, Nishikawa T (2018) Involvement of neuronal and glial activities in control of the extracellular d-serine concentrations by the AMPA glutamate receptor in the mouse medial prefrontal cortex. Neurochem Int 119:120–125. https://doi.org/10.1016/j.neuint.2017.09.009
Kanematsu S, Ishii S, Umino A, Fujihira T, Kashiwa A, Yamamoto N, Kurumaji A, Nishikawa T (2006) Evidence for involvement of glial cell activity in the control of extracellular d-serine contents in the rat brain. J Neural Transm (Vienna) 113:1717–1721. https://doi.org/10.1007/s00702-006-0517-3
Schell MJ, Molliver ME, Snyder SH (1995) d-Serine, an endogenous synaptic modulator: localization to astrocytes and glutamate-stimulated release. Proc Natl Acad Sci USA 92:3948–3952. https://doi.org/10.1073/pnas.92.9.3948
Yasuda E, Ma N, Semba R (2001) Immunohistochemical evidences for localization and production of d-serine in some neurons in the rat brain. Neurosci Lett 299:162–164. https://doi.org/10.1016/s0304-3940(01)01502-6
Kartvelishvily E, Shleper M, Balan L, Dumin E (2006) Neuron-derived d-serine release provides a novel means to activate N-methyl-d-aspartate receptors. J Biol Chem 281(20):14151–14162. https://doi.org/10.1074/jbc.M512927200
Williams SM, Diaz CM, Macnab LT, Sullivan RK, Pow DV (2006) Immunohistochemical analysis of d-serine distribution in the mammalian brain reveals novel anatomical compartmentalizations in glia and neurons. Glia 53:401–411. https://doi.org/10.1002/glia.20300(2006)
Balu DT, Takagi S, Puhl MD, Benneyworth MA, Coyle JT (2014) d-Serine and serine racemase are localized to neurons in the adult mouse and human forebrain. Cell Mol Neurobiol 34:419–435. https://doi.org/10.1007/s10571-014-0027-z
Benneyworth MA, Li Y, Basu AC, Bolshakov VY, Coyle JT (2012) Cell selective conditional null mutations of serine racemase demonstrate a predominate localization in cortical glutamatergic neurons. Cell Mol Neurobiol 32:613–624. https://doi.org/10.1007/s10571-012-9808-4
Schwarcz R, Whetsell WO Jr, Mangano RM (1983) Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science 219:316–318. https://doi.org/10.1126/science.6849138
Paxinos G, Watson C (1986) The rat brain in stereotaxic coordinates, 2nd edn. Academic Press, San Diego
Dihné M, Block F, Korr H, Töpper R (2001) Time course of glial proliferation and glial apoptosis following excitotoxic CNS injury. Brain Res 902:178–189. https://doi.org/10.1016/s0006-8993(01)02378-2
Haas SJ, Ahrens A, Petrov S, Schmitt O, Wree A (2004) Quinolinic acid lesions of the caudate putamen in the rat lead to a local increase of ciliary neurotrophic factor. J Anat 204:271–281. https://doi.org/10.1111/j.0021-8782.2004.00279.x
Slawik H, Volk B, Fiebich B, Hüll M (2004) Microglial expression of prostaglandin EP3 receptor in excitotoxic lesions in the rat striatum. Neurochem Int 45:653–660. https://doi.org/10.1016/j.neuint.2004.04.007
Umino A, Nishikawa T, Takahashi K (1995) Methamphetamine-induced nuclear c-Fos in rat brain regions. Neurochem Int 26(1):85–90. https://doi.org/10.1016/0197-0186(94)00096-d
Nishikawa T, Scatton B (1985) Inhibitory influence of GABA on central serotonergic transmission. Involvement of the habenulo-raphé pathways in the GABAergic inhibition of ascending cerebral serotonergic neurons. Brain Res 331:81–90. https://doi.org/10.1016/0006-8993(85)90717-6
Senatorov VV, Ren M, Kanai H, Wei H, Chuang DM (2004) Short-term lithium treatment promotes neuronal survival and proliferation in rat striatum infused with quinolinic acid, an excitotoxic model of Huntington’s disease. Mol Psychiatry 9:371–385. https://doi.org/10.1038/sj.mp.4001463
Brickell KL, Nicholson LF, Waldvogel HJ, Faull RL (1999) Chemical and anatomical changes in the striatum and substantia nigra following quinolinic acid lesions in the striatum of the rat: a detailed time course of the cellular and GABA(A) receptor changes. J Chem Neuroanat 17:75–97. https://doi.org/10.1016/s0891-0618(99)00029-0
Rosenberg PA, Dai W, Gan XD, Ali S, Fu J, Back SA, Sanchez RM, Segal MM, Follett PL, Jensen FE, Volpe JJ (2003) Mature myelin basic protein-expressing oligodendrocytes are insensitive to kainate toxicity. J Neurosci Res 71:237–245. https://doi.org/10.1002/jnr.10472
Björklund A, Lindvall O (1984) Dopamine-containing systems in the CNS. In: Björklund A, Hökfelt T (eds) Handbook of chemical neuroanatomy, vol 2. Classical Transmitters in the CNS, Part2. Elsevier, Amsterdam, pp 55–122
Ishiwata S, Umino A, Umino M, Yorita K, Fukui K, Nishikawa T (2013) Modulation of extracellular d-serine content by calcium permeable AMPA receptors in rat medial prefrontal cortex as revealed by in vivo microdialysis. Int J Neuropsychopharmacol 16:1395–1406. https://doi.org/10.1017/S1461145712001484
Nishijima K, Kashiwa A, Hashimoto A, Iwama H, Umino A, Nishikawa T (1996) Differential effects of phencyclidine and methamphetamine on dopamine metabolism in rat frontal cortex and striatum as revealed by in vivo dialysis. Synapse 22:304–312. https://doi.org/10.1002/(SICI)1098-2396(199604)22:4%3c304::AID-SYN2%3e3.0.CO;2-F
Sakurai S, Ishii S, Umino A, Shimazu D, Yamamoto N, Nishikawa T (2004) Effects of psychotomimetic and antipsychotic agents on neocortical and striatal concentrations of various amino acids in the rat. J Neurochem 90:1378–1388. https://doi.org/10.1111/j.1471-4159.2004.02592.x
Umino A, Takahashi K, Nishikawa T (1998) Characterization of the phencyclidine-induced increase in prefrontal cortical dopamine metabolism in the rat. Br J Pharmacol 124:377–385. https://doi.org/10.1038/sj.bjp.0701830
Fonnum F (1987) Biochemistry, anatomy and pharmacology of GABA neurons. In: Meltzer HY (ed) Psychopharmacology; the third generation of progress. Raven Press, New York, pp 173–182
Cotman CW, Monaghan DT (1987) Chemistry and anatomy of excitatory amino acid systems. In: Meltzer HY (ed) Psychopharmacology; the third generation of progress. Raven Press, New York, pp 197–210
Gunčová I, Látr I, Mazurová Y (2011) The neurodegenerative process in a neurotoxic rat model and in patients with Huntington’s disease: histopathological parallels and differences. Acta Histochem 113:783–792. https://doi.org/10.1016/j.acthis.2010.11.007
Beal MF, Swartz KJ, Finn SF, Mazurek MF, Kowall NW (1991) Neurochemical characterization of excitotoxin lesions in the cerebral cortex. J Neurosci 11:147–158. https://doi.org/10.1523/JNEUROSCI.11-01-00147.1991
Gerónimo-Olvera C, Tristán-López L, Martínez-Lazcano JC, García-Lara L, Sánchez-Mendoza A, Morales-Martínez A, Hernández-Melesio MA, Arregui L, Ríos C, Pérez-Severiano F (2019) Striatal protection in nNOS knock-out mice after quinolinic acid-induced oxidative damage. Neurochem Res 44:421–427. https://doi.org/10.1007/s11064-018-2688-3
Santamaría A, Ríos C, Pérez P, Flores A, Galván-Arzate S, Osorio-Rico L (1996) Quinolinic acid neurotoxicity: in vivo increased copper and manganese content in rat corpus striatum after quinolinate intrastriatal injection. Toxicol Lett 87:113–119. https://doi.org/10.1016/0378-4274(96)03772-1
Beal MF, Kowall NW, Ellison DW, Mazurek MF, Swartz KJ, Martin JB (1986) Replication of the neurochemical characteristics of Huntington’s disease by quinolinic acid. Nature 321:168–171. https://doi.org/10.1038/321168a0
Goes TC, Almeida Souza TH, Marchioro M, Teixeira-Silva F (2018) Excitotoxic lesion of the medial prefrontal cortex in Wistar rats: effects on trait and state anxiety. Brain Res Bull 142:313–319. https://doi.org/10.1016/j.brainresbull.2018.08.009
Miya K, Inoue R, Takata Y, Abe M, Natsume R, Sakimura K (2008) Serine racemase is predominantly localized in neurons in mouse brain. J Comp Neurol 510:641–654. https://doi.org/10.1002/cne.21822
Kolodney G, Dumin E, Safory H, Rosenberg D, Mori H, Radzishevsky I, Wolosker H (2015) Nuclear compartmentalization of serine racemase regulates d-serine production: implications for N-methyl-d-aspartate (NMDA) receptor activation. J Biol Chem 290:31037–31050. https://doi.org/10.1074/jbc.M115.699496
Ignarro RS, Vieira AS, Sartori CR, Langone F, Rogério F, Parada CA (2013) JAK2 inhibition is neuroprotective and reduces astrogliosis after quinolinic acid striatal lesion in adult mice. J Chem Neuroanat 48–49:14–22. https://doi.org/10.1016/j.jchemneu.2013.02.005
Perez EJ, Tapanes SA, Loris ZB, Balu DT, Sick TJ, Coyle JT, Liebl DJ (2017) Enhanced astrocytic d-serine underlies synaptic damage after traumatic brain injury. J Clin Invest 127:3114–3125. https://doi.org/10.1172/JCI92300
Li S, Uno Y, Rudolph U, Cobb J, Liu J, Anderson T, Levy D, Balu DT, Coyle JT (2018) Astrocytes in primary cultures express serine racemase, synthesize d-serine and acquire A1 reactive astrocyte features. Biochem Pharmacol 151:245–251. https://doi.org/10.1016/j.bcp.2017.12.023
Fonnum F, Storm-Mathisen J, Divac I (1981) Biochemical evidence for glutamate as neurotransmitter in corticostriatal and corticothalamic fibres in rat brain. Neuroscience 6:863–873. https://doi.org/10.1016/0306-4522(81)90168-8
Hassler R, Haug P, Nitsch C, Kim JS, Paik K (1982) Effect of motor and premotor cortex ablation on concentrations of amino acids, monoamines, and acetylcholine and on the ultrastructure in rat striatum. A confirmation of glutamate as the specific cortico-striatal transmitter. J Neurochem 38:1087–1098. https://doi.org/10.1111/j.1471-4159.1982.tb05352.x
Acknowledgements
We thank Ms. Koyuki Kaneda and Ms. Mika Nakai for their technical assistance. We are very grateful to Dr. Katsunobu Takahashi for his help in analyzing some of the data from the microdialysis experiments and his valuable comments on a part of the data of these experiments at the National Institute of Neuroscience. This study was partly supported by the Grant-in-Aid for Scientific Research B (Research Program No. 19390302) and C (Grant No. 18K07548), and a Grant-in-Aid for Scientific Research on Priority Areas from the Ministry of Culture, Sports, Science and Technology of Japan, a Grant from the Ministry of Health, Labor and Welfare, and the CREST (Core Research for Evolutional Science & Technology) program funded by the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Funding
Funding was provided by Ministry of Culture, Sports, Science and Technology of Japan [Grant-in-Aid for Scientific Research B (Research Program No. 19390302), Grant-in-Aid for Scientific Research C (18K07548)], Ministry of Education, Culture, Sports, Science and Technology of Japan (Grant No. CREST), and Ministry of Health, Labor and Welfare of Japan (Research Grant).
Author information
Authors and Affiliations
Contributions
AU and TN established the methods for the in vivo microdialysis technique, and the semi-automatic quantitative HPLC analysis of the chiral and non-chiral amino acids and of monoamines and their metabolites, which were used in the present study. AU and HI performed the animal experiments and HPLC measurements of the various amino acids and monoamines, wrote their protocols and parts of the draft of the manuscript, figures and a table, and undertook the statistical analysis. TN and DS performed the cresyl violet staining of the brain sections and prepared their histological specimens. MU conducted the histological analysis by macro- and microscopic observations and preparation of their photomicrographs and figures, achieved parts of statistical analysis, and wrote the corresponding portions of the draft of the manuscript. YK discussed and validated the experimental designs and data analyses, and critically reviewed the draft versions of the manuscript. TN conceived, designed and directed this project including the statistical analysis, and wrote the final version of the manuscript. All authors contributed to and have approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Umino, A., Iwama, H., Umino, M. et al. Effects of Quinolinate-Induced Lesion of the Medial Prefrontal Cortex on Prefrontal and Striatal Concentrations of d-Serine in the Rat. Neurochem Res 47, 2728–2740 (2022). https://doi.org/10.1007/s11064-022-03627-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-022-03627-8