
Abstract
The increase in the frequency and severity of extreme weather events affects populations of tree species. As a result, many forests around the world have suffered massive mortality due to extreme droughts. Therefore, it is important to understand the potential adjustments of dominant woody species of temperate forests to climate change. The objective was to show in seedlings inhabiting the most contrasting habitats, forest types, and precipitation regimes the relevance of species-specific and intra-specific adaptive trait variation. Ecophysiological differences between seedlings collected from distinct humid and dry provenances of Nothofagus pumilio and N. dombeyi in northern Patagonia were assessed using common garden and water stress manipulative experiments. Levels of genetic diversity and divergence were evaluated with microsatellite analysis. Seedlings from humid provenances of both species showed increased growth, and leaf traits differed significantly between N. dombeyi provenances. Studied species behaved differently under water stress, isohydric for N. dombeyi and anisohydric for N. pumilio, while no intraspecific differences were measured in the regulation of water status. Humid provenance of N. dombeyi had higher genetic diversity than the dry one, whereas those of N. pumilio were similarly diverse. Yet, low but significantly different from zero genetic divergence was measured between provenances of the latter. Our results show species-specific responses to water stress and that adaptive variation at distinct provenances has a genetic basis that is maintained despite potential gene flow. The strategy of using seedlings from "regional admixture provenancing" is suggested for long-term management of vulnerable taxa to desiccation as the mesic N. dombeyi.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It is well recognized the effect of climate on tree species’ populations (IPCC 2014). Variation in climate along environmental gradients produces significant intraspecific variation in key traits related to survival and performance. Because of the relatively limited vagility of propagules and the slow-growing nature of trees, it is relevant to understand the potential in situ adjustments of woody species to climate changes. Historical records show that plants, in general, and trees, in particular, were able to respond to past climate changes, either through migration tracking their ecological optimum or locally adapting to new conditions (e.g. Hampe and Jump 2011). Nonetheless, current climate changes are occurring with unprecedented speed and extent in recorded history that deserves studying on how species can respond to new conditions (Neukom et al. 2019).
Forests are exposed to different types of disturbances, including extreme weather events, and droughts are considered to have the greatest global effects (Reichstein et al. 2013). If water is one of the environmental factors that most influences growth rate and biomass production (Shao et al. 2008), trees will be vulnerable to water shortage. Particularly under global change, longer and more intense droughts are forecasted which can even trigger tree mortality (Choat et al. 2018). Thus, it is timely to understand the mechanisms of the stomatal aperture as they regulate tree water loss and carbon gain. Nonetheless, plant populations and species use water according to their genetic and plastic ecophysiological makeups, resulting in distinct behaviors that vary from water-conserving to risk-taking shown by isohydric and anisohydric plants, respectively, as well as from some intermediate responses between the aforesaid extremes (Attia et al. 2015). Isohydric species retain conservative water-balance management, in which the loss of water is limited by the reduction of stomatal conductance (Sade et al. 2012). In contrast, anisohydric plants maintain higher stomatal conductance by a less strict stomatal control and consequently they track environmental fluctuations (Martinez‐Vilalta et al. 2014). A downside of risk-taking stomatal behavior of anisohydric plants may result in embolism and desiccation during water stress. However, it was shown that anisohydric plants are resistant to cavitation as they may rapidly recover from water shortages, and thus they can be considered drought-resistant (Sade et al. 2012).
As soil water becomes limiting, isohydric plants are expected to reduce stomatal conductance and to survive, while under prolonged severe drought anisohydric trees are expected to die (reviewed by Moshelion et al. 2014). Yet the regulation of isohydric and anisohydric responses and their underlying physiological mechanisms are not yet fully understood (Martinez‐Vilalta et al. 2014) particularly at the intraspecific level of populations inhabiting contrasting precipitation conditions. The difference between isohydric and anisohydric behavior can be determined, relatively simple, from the interception (i) and slope (s) parameters based on the relationship between predawn (Ψpd) and midday (Ψmd) water potential of trees following Martinez‐Vilalta et al. (2014). Four different behaviors can be distinguished, sharing the same interception: strict isohydric (s = 0), partial isohydric (0 < s < 1), strict anisohydric (s = 1) and extreme anisohydric (s > 1). If Ψpd equals Ψmd the gas exchange ceases; whereas if the Ψmd decreases, hydraulic conductance loss occurs. Isohydric or anisohydric regulation of leaf water potential may partition species survival and mortality and may be an effective approach to modeling plant survival and mortality under future climate conditions (McDowell et al. 2008).
Especially important is to analyze adjustments of plant populations and species facing unusual local and regional climates. While short-term responses may involve plastic adjustments, it becomes relevant to evaluate levels of genetic variation in populations that may provide the raw material for genetically-based adaptations that guarantee long-term persistence. Thus, the analysis of traits under selection particularly growth and ecophysiological ones related to key climate parameters becomes relevant. Such characteristics can be evaluated under common gardens to disentangle the potential genetic basis of character variation observed in nature as well as experimentally through imposed climatic stress. Nonetheless, highly outcrossed tree populations and species are also subjected to selectively neutral forces as gene flow that limits among-population divergence (Hamrick 2004). Yet, under strong selection, the ‘swamping out’ effects of gene flow should be negligible (Sork 2016). Therefore, the analysis of adaptive and neutral traits may help to disentangle their relative contribution prompting among-population divergence.
Forests provide a wide range of ecosystem services and, in a context of environmental change of uncertain magnitude, management actions need to be developed to assure their abiding persistence. Tree species are genetically diverse which determines their great potential for plastic and adaptive adjustments to climate change through gene flow and natural selection (Fady et al. 2016). Yet, species-specific responses (individualistically) are expected to changing climates and environments because while some taxa may be vulnerable others will be naturally buffered and resilient to climate-influenced disturbances (Millar et al. 2007). Similarly, species consist of populations that become adjusted to distinct ecological settings which need to be considered under management. Thus, it is timely to compare potential population responses to variable environmental conditions of species with distinct autoecological characteristics particularly of dominant taxa that define communities, e.g. those that determine species’ interactions, and have a key role within the ecosystem functioning. In addition, we aim to contribute to the discussion on how evergreen and deciduous taxa handle drought. A recent analysis of seasonally dry tropical forests suggested that deciduous species are less tolerant to drought than evergreen species (Cruz de Souza et al. 2020). Also, different hydraulic strategies in terms of water use were documented for species under Mediterranean climates, with dry summers and wet winters; while evergreens showed a more conservative behavior (isohydric), deciduous ones occurring in the same region were less conservative in water use (anisohydric) (Mediavilla and Escudero 2003).
Patagonia is one of the places in the world where forests have been and are being impacted by recent climate extreme events (Allen et al. 2010) and, as a result, signals of forest decay and massive tree mortality have been documented (Suarez et al. 2004; Rodríguez-Catón et al. 2019). Predictions for Patagonia include warmer climates and increased drought, and the observed negative precipitation trend of the last century is projected to continue (Barros et al. 2014). These forecasted conditions are even stronger in northern Patagonia, Argentina, where the rain shadow effect of the Andes creates a steep West to East precipitation gradient. Thus, populations of wide-ranging forest species will find extremely variable conditions particularly at the opposite ends of such gradient. Populations at the dry-most end are probably exposed to regular droughts and therefore will consist of a set of genotypes adapted to water stress. In contrast, such water-stress resistant traits were probably removed by stabilizing selection in populations under more humid conditions which in turn can be more susceptible to increased drought under current trends. Evidence of massive tree mortality was recorded in humid populations of the mesic Nothofagus dombeyi as a result of the extreme drought event during the summer of 1998–1999 in northern Patagonia (Suarez et al. 2004). On the other hand, such decay was not observed at the dry-end populations, suggesting that they may consist of particular genotypes which may enable them to respond to current and future climatic trends (Diaz et al. 2020).
Dominant tree species of the family Nothofagaceae within the subgenus Nothofagus as the evergreen Nothofagus dombeyi (Mirb.) Oerst. and the deciduous Nothofagus pumilio (Poepp. et Endl) Krasser are widespread throughout low and high-elevation temperate forests, respectively, of southern Argentina and Chile. They are key components of the ecosystems where they inhabit, and, on the eastern slopes of the Andes, in northern Patagonia, they usually consist of mostly pure stands. Therefore, any impact on such species may have profound consequences on forest communities that span more than 1000 km coverage including their dependent species. Such Nothofagus consist of heterogeneous gene pools along gradients. These can be due to historical isolation that produces among-population divergence (e.g. along latitude, Premoli et al. 2012) and climate exerting differential selective pressures (e.g. at distinct elevations, Premoli 2003).
The objective of this study was to show, in widespread tree species with contrasting autoecological characteristics (low-elevation evergreen N. dombeyi and high-elevation deciduous N. pumilio), the effects of divergent selection as a result of distinct precipitation regimes due to the rainshadow effect of the Andes. We used evidence on adaptive traits measured on seedlings maintained under common gardens and manipulative experiments of water stress to analyze the impact of natural selection. Molecular information by microsatellites was used to measure levels of among-population divergence and genetic diversity in small and isolated populations towards dry-end marginal areas compared to relatively continuous ones under more humid conditions. The hypothesis tested here is that despite potential among-population gene flow, the populations maintain significant adaptive variation. We aim to contribute towards the discussion of the value of dry-edge populations, often considered marginal and of reduced ecological significance, as reservoirs of genetically-based adaptations. Although analyses on seedlings cannot be extrapolated to adults, we seek to highlight the relevance of inter- and intra-specific trait variation in response to droughts that can be used in management, conservation, and restoration practices of dominant forest trees.
Methodology
Study species
In this study were compared two sister species inhabiting contrasting habitats and elevations of the southern Andes, N. pumilio, commonly known as “lenga” and N. dombeyi, common name “coihue”. Nothofagus pumilio is a winter deciduous tree that dominates high-latitude and high-elevation environments of southern Argentina and Chile where it mostly occurs as monospecific forests. It is distributed from 36° 50′ S to 56° S latitude along the Cordillera de los Andes (Dimitri 1972; Donoso 1995). At its southern range, it can be found at sea level, although towards the north it usually forms the upper altitudinal boundary of the alpine timberline and dominates the transition between forests and subalpine vegetation where most of the precipitation falls as snow (Veblen et al. 1996). Ecophysiological studies showed a lack of daily variation in the leaf osmotic pressures of N. pumilio suggesting that water content is regulated by the opening and closing of the stomata (Alberdi 1987). Also, field and common garden gas exchange measures showed that the use of water is highly plastic (Premoli and Brewer 2007). Nothofagus dombeyi is an evergreen tree and is the largest of the South American Nothofagus that can attain 50 m height and 2 m of diameter (Veblen et al. 1996). It has a wide latitudinal distribution between 34° 37′ and 47° 30′ S latitude in Chile and 38° 30′ to 40° S in Argentina (Donoso et al. 2004). It inhabits a great variety of climates and forest types although on the eastern Andes is most commonly found under humid and relatively low-to mid-elevation forests. Nothofagus dombeyi has higher cuticular transpiration than other evergreen Nothofagus species (Alberdi 1987). Ecophysiological differences between N. dombeyi and N. pumilio include the higher wood density in the former (Bucci et al. 2012). Wood density has been related to resistance to embolism (Hacke et al. 2001), making the latter more susceptible to cavitation which in turn could be compensated by a higher leaf hydraulic efficiency (Bucci et al. 2012). Also, contrasting responses to extreme drought have been recorded; while premature leaf abscission and/or partial tree crown mortality can be observed in the deciduous N. pumilio during dry summers, the evergreen N. dombeyi suffers mortality during drought (Veblen et al. 1996).
Study area and sampling
Eastern slopes of the Andes in Argentina are under types of a Mediterranean climate with summer droughts and where most precipitation that falls in winter is in the form of snow. Soils may also vary along the precipitation gradient, at the west end forests grow predominantly on Andisols, i.e., volcanic soils (1000–3000 mm rainfall), while towards the east soils are Aridisols (from 500 to 100 mm rainfall) (Mazzarino et al. 1998). The study area of N. pumilio and N. dombeyi is located approximately at 41° S latitude where the rainshadow effect of the Andes produces an abrupt West to East precipitation gradient. Plant collection took place at the precipitation extremes between longitudes 72 and 71° W, considered hereafter humid and dry provenances, respectively. The mean annual temperature for N. pumilio and N. dombeyi at the humid provenance is 5.8 and 7.6° C with a total annual rainfall of 1100 mm; while in the dry provenance, the mean annual temperature is 5.7 and 9.1 °C with a total annual rainfall of 790 mm (Fig. S1; climate data extracted from WorldClim v.2; Fick and Hijmans 2017). Along the abrupt west–east precipitation gradient, N. pumilio and N. dombeyi forests consist of relatively continuous stands towards the humid end. At the dry end, N. pumilio stands becomes increasingly isolated to forest patches; while the presence of N. dombeyi is scarce and restricted to riparian environments as river banks where soil water is not limited at any period of the year (Donoso et al. 2004).
At different humid and dry provenances (Fig. 1 and Table 1), seedlings of both species bearing cotyledons, i.e., of the same age, that had germinated under natural conditions during the austral spring were collected directly from the field in three different years: October–December 2012 and 2013 for N. pumilio, and February 2016 for N. dombeyi. Collection sites and years reflect the fact that seed production and viability, which in turn are highly correlated, vary enormously between years, sites, and Nothofagus species (Veblen et al. 1996). All seedlings of N. pumilio were collected at the lower limit of the elevation gradient to control for adaptive differences with elevation previously reported (Premoli et al. 2007). Humid provenance of N. dombeyi corresponds to the forest stand that suffered from massive tree mortality during the extreme drought recorded during the summer of 1998–1999 (Suarez et al. 2004). Seedlings of distinct humid and dry provenances were part of previous individual studies on N. pumilio and N. dombeyi (Ignazi et al. 2020; Diaz et al. 2020).

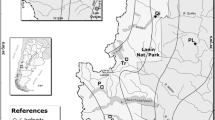
Map of northern Patagonia showing the sampling sites for Nothofagus pumilio (triangles) and N. dombeyi (circles). Seedling and leaf samples for common garden and molecular analyzes were collected at three humid provenances of N. pumilio at BAY, Cerro Bayo, OTO, Cerro Otto, and TRO, Cerro Tronador; and three dry provenances at FOR, Fortin, CHB, Valle del Challhuaco and NIR, Ñirihuau. In N. dombeyi, humid provenances corresponded to A, Arroyo Melgarejo and B, Casa del Lago; and dry provenances to C and D, Estancia Chacabuco Zone 1 and Zone 2, respectively. Isolines represent precipitation (mm year−1)
Common garden measurements
Seedling pools of N. pumilio and N. dombeyi were cultivated in common gardens during a total period of three and two years, respectively, since collection from the field. Potentially genetically-based differences between seedlings from different provenances were monitored under optimal growing conditions for both species. Seedlings were placed in individual pots, properly tagged and identified by collection site, with a substrate consisting of a mixture of their native forest soil which was kept to the minimum amount (c. 10%) to avoid root damage during manipulation of seedlings and topsoil (c. 90%) rich in organic matter collected nearby the greenhouse. Seedlings were grown with a randomized arrangement in a naturally lighted greenhouse of the Instituto de Investigaciones en Biodiversidad y Medio Ambiente in Bariloche (INIBIOMA- Universidad Nacional del Comahue), located at an intermediate elevation than seedling provenances (874 m a.s.l.). Throughout the experiment, seedlings were planted out to larger pots twice, reaching a final volume of 1 and 6 L for N. pumilio and N. dombeyi, respectively, to ensure the water availability required for the water stress experiment. Smaller plots were used for N. pumilio because seedlings were smaller than those of N. dombeyi at the time the experiment was performed. Pots were periodically rotated to avoid distinct microclimates within the common garden.
Plant mortality was recorded at the end of the common garden experiment as the number of plants that died from the total of seedlings collected in the field (N = 371 from N. pumilio and N = 517 from N. dombeyi). The common garden trial consisted of measuring distinct plant traits during cultivation. These included: total plant height (cm), basal diameter (mm), number of branches, and the total number of leaves, that were measured in 73 seedlings of N. pumilio during two consecutive years in October of 2015 and 2016, and 32 seedlings of N. dombeyi in February of 2016 and November of 2017. Leaf morphological features were measured at the end of the common garden experiment on five and 16 seedlings of N. pumilio and N. dombeyi, respectively, that were randomly selected from each seedling pool of dry and humid provenances. Five randomly chosen leaves per individual were scanned to measure the leaf area, perimeter, length, width, and roundness using the program Digimizer v 4.6.1 (MedCalc Software 2015). To quantify the dry biomass, leaves were dried at 65 °C, until a constant mass was obtained, and weighed using a digital balance (Mettler AJ150) with an accuracy of 0.0001 g. Finally, the stomatal density was measured by counting the number of stomata on the abaxial surface of five randomly selected leaves from 10 plants of each provenance using transparent enamel following Premoli and Brewer (2007).
Manipulative experiment of water deficit
Seedlings of similar size of each species were selected from the seedling pools to conduct a water deficit experiment over three weeks according to Varela et al. (2010). The experiments took place during early November 2016 and 2017 on 103 N. pumilio seedlings (75 from dry and 28 from humid provenances) and 32 N. dombeyi seedlings (16 from each dry and humid provenances), respectively. Sample size varied according to the availability and plants’ health status for each species.
The experiment consisted of two watering treatments, water deficit (near permanent wilting point; PWP) and control (near filed capacity; FC). To determine FC and PWP, a soil water retention curve (− 0.03, − 0.1, − 0.15, − 0.7, and − 1.5 MPa) was developed in the Soil Laboratory of the Instituto Nacional de Tecnología Agropecuaria in Bariloche (INTA-EEA Bariloche) to ensure the watering condition of the different treatments. Soil moisture was determined as the volumetric water content (VWC = % vol./vol.) and was recorded every two or three days, using a digital TDR (Time Domain Reflectometry, ProCheck from Decagon Devices, Inc.). We maintained the water deficit treatment near 13.8% VWC and the control treatment near 21.5% VWC throughout the experiment (Fig. S2). At the end of the water shortage treatment, the maximum water potentials (at predawn, from 5:00 to 8:00 am) and minimum water potentials (at midday, from 12:00 to 2:00 pm) were recorded in five randomly selected branches of each plant by treatment/provenance using a pressure chamber (PMS 1000, PMS Instruments, Corvallis, Oregon, USA).
Climatic variables
Based on 19 bioclimatic variables obtained from WorldClim, we determined those that best explained studied species’ ranges using MAXENT v3.4.1 (Phillips et al. 2021) (data not shown). A correlation matrix was made between the 19 bioclimatic variables using STATISTICA v.7.0 StatSoft, Inc. (2004), and weakly correlated variables (< 75%) were selected for further analysis (Fig. S3). These were: Annual Mean Temperature (Bio 1), Maximum Temperature of Warmest Month (Bio 5), Minimum Temperature of Coldest Month (Bio 6), Mean Temperature of Warmest Quarter (Bio 10), Precipitation of Wettest Quarter (Bio 16), and Precipitation of Driest Quarter (Bio 17).
Molecular genetic characteristics
Neutral genetic differences between provenances were analyzed at the molecular level using microsatellite markers. DNA was extracted from a total of 51 adult N. pumilio individuals (25 from dry and 26 from humid provenances) following the method with ATMAB described in Dumolin et al. (1995) from isozyme extracts (Arbetman and Premoli 2011). In the case of N. dombeyi, DNA extraction was performed on a total of 20 seedlings, 10 from each dry and humid provenance, based on the protocol of Novaes et al. (2009) with modifications (Diaz et al. 2020). We amplified eight loci that were previously used in N. pumilio (Mathiasen and Premoli 2013) and N. macrocarpa (Mathiasen et al. 2020). These were: Nerv111 and NnBio37 from Marchelli et al. (2008); and ncutas4, ncutas8, ncutas9, ncutas12, ncutas21, and ncutas22 from Jones et al. (2004), five and six of which were successfully amplified in N. pumilio (Ignazi 2019) and N. dombeyi (Diaz 2018), respectively.
The polymerase chain reaction (PCR) mixture contained 1.5 μl of DNA, 1 μl of buffer 10X (Invitrogen), 2 μM of MgCl2 (Invitrogen), 0.2 mM of each dNTP (Promega), 0.5 μM of each primer (Alpha DNA), and 1 U of Taq DNA polymerase (Invitrogen) in a total volume of 10 μL. The tubes were incubated in a thermocycler (Applied Biosystems) with the following PCR amplification conditions of DNA fragments: initial denaturation of 5 min at 94 °C; 35 cycles of 30 s at 94 °C denaturation, 30 s at the primer annealing temperature (50 °C for ncutas12 and ncutas9; 55 °C for ncutas8, ncutas21, and NnBio37; 57 °C for Nerv111 and ncutas4; 58 °C for ncutas22), and 90 s at 72 °C extension; and finally 30 min of final extension at 72 °C. The amplified fragments were sent to Macrogen (Seoul, Korea) to be sequenced using the capillary electrophoresis technique. The different genotypes were recorded with the program Peak Scanner v 1.0 (Guichoux et al. 2011) assigning to each allele the size of the amplified fragment obtained by comparison with a size standard marker (GS500LIZ).
Data analysis
Mortality between provenances of seedlings of each species was compared by t-tests. The variables of the common garden measurements were evaluated by general linear models (GLM) of repeated measures at two-time stages for each species and then Tukey's HSD post-hoc tests were used to compare between provenances. Stomatal densities and morphological leaf traits of populations from different provenances were compared by one-way ANOVA for each species. To test for potential effects of the initial size at plant collection (Hi) on total shoot length of the last measurements made in October 2016 and November 2017 (Hf) we calculated the relative growth rate as RGR = ln (Hf/Hi) and regressed this value against ln (Hi) for N. pumilio and N. dombeyi plants, respectively, following Premoli et al. (2007). To analyze the differential dependence of RGR to initial shoot length for humid and dry provenances during the greenhouse experiment, we compared regression slopes by using the two-slopes comparison test (Zar 1996) with the STATISTICA program. The slope model is used when one or more continuous predictor variables (covariates: ln (Hi) and RGR) have different effects at different levels of one or more categorical independent variables (factor: humid and dry).
We plotted the relationship between early predawn (Ψpd) and midday (Ψmd) water potentials and the fitted model was used to estimate the value of the intercept (i) and slope (s) for each species and provenance. In particular, the intercept of the relationship between predawn and midday water potential depicts the maximum transpiration rate per unit of hydraulic transport capacity, while the slope measures the relative sensitivity of the transpiration rate and plant hydraulic conductance to declining water availability (Martinez-Vilalta et al. 2014). We used general linear regression models to study the relationships between intercept and slope parameters at the species and provenance levels with climatic variables using STATISTICA.
Standard genetic diversity indices were estimated for microsatellites of each dry and humid provenance for N. pumilio and N. dombeyi. These were: average number of alleles per locus (A), the effective number of alleles (Ae), allelic richness by the rarefaction method that corrects by sample size (Ar), total number of alleles (At), number of private alleles (AP), percentage of polymorphic loci under the sensu stricto criterion (presence of at least two alleles regardless of their frequency) (%P), and the observed and expected heterozygosities under the Hardy Weinberg equilibrium condition (Ho and He, respectively) using the programs GenAlEx v 6.2 (Peakall and Smouse 2006) and FSTAT v.2.9.3 (Goudet 2001). To assess the existence of significant by-locus differences between humid and dry provenances paired t-tests were run for all genetic diversity parameters using STATISTICA. The analysis of molecular variance (AMOVA test) was also performed to explore within- and between-provenance differences for each species using GenAlEx. The degree of between-provenance divergence was estimated for each species by FST using FSTAT that also provides confidence intervals by bootstrapping over loci and thus a measure of the departure from zero values. Values close to zero indicate little between-provenance divergence and thus relatively high gene flow.
Results
Common garden measurements
Mortality was relatively high for both species during the cultivation in the greenhouse. The humid provenance of N. pumilio and dry provenance of N. dombeyi showed significantly higher mortality (P < 0.05) at the end of the experiment (Fig. 2). Seedlings’ growth traits differ significantly between dry and humid provenances of each species, except for the number of branches of N. pumilio. Total plant growth, basal diameter, and the number of leaves attained greater values in plants from the humid provenance of each species (P < 0.05; Fig. 3). Similar RGR was obtained for both provenances of each species (P > 0.05; Fig. 4).

Percentage of mortality recorded at three stages during cultivation in the common garden for a Nothofagus pumilio and b N. dombeyi seedlings from different provenances. Different letters indicate significant differences P < 0.05, t-test

Temporal variation of the architectural and leaf variables of plants measured in the common garden of Nothofagus pumilio and N. dombeyi in two growing periods: a, b height, c, d number of leaves, and e, f number of branches and g, h basal diameter. Lowercase letters indicate statistical differences within each provenance. Capital letters show statistical differences between the provenances. Results are presented as means ± standard error

Relationship between the relative growth rate (RGR) and the initial plant height ln(Hi) of a Nothofagus pumilio seedlings from humid (filled triangle) and dry (empty triangle) provenances and b N. dombeyi seedlings from humid (filled circles) and dry (empty circles) provenances. The slope test was non-significant P > 0.05 within each species
Leaf morphology differed significantly between dry and humid provenances of N. dombeyi including dry biomass, area, length, width, perimeter, and stomatal density (P < 0.05; Table 2) attaining greater values in seedlings from the humid provenance. Other leaf traits measured as roundness and specific leaf area (SLA), did not vary between provenances (P > 0.05; Table 2). In contrast, non-significant differences between the dry and humid provenances of N. pumilio were obtained for all leaf traits (P > 0.05; Table 2).
Manipulative experiment of water deficit
Both species and provenances differed in their stomatal regulation of water status as evidenced by the slope value of the relation between predawn and midday water potentials: s = 1.2 and 1.0 for humid and dry N. pumilio, and s = 0.3 and − 0.2 for humid and dry N. dombeyi, respectively (Table S1). Linear regression models that were used to test for differences in the relationship between the predawn and midday water potentials showed a good fit for N. pumilio provenances, with a significant decline in the water potential at midday; while provenances of N. dombeyi had no variation (Fig. 5a; Table S1). Intraspecific comparisons of slopes yielded non-significant results whereas significant differences were observed between the slope of dry N. pumilio and both N. dombeyi provenances (Table S2). The intercept and slope ratios of each species and provenances resulted in a highly significant correlation (r = 0.98; P = 0.014), and N. pumilio attained greater slope values and less negative midday water potentials than N. dombeyi (Fig. 5b). In turn, these parameters showed a high association with the different temperature-related climatic variables that separated both species, with both provenances of N. pumilio occupying lower temperature habitats and reaching greater slopes and less negative midday water potentials than N. dombeyi (Fig. 6). On the other hand, precipitation-related variables yielded a nearly linear relationship between provenances and species, except for dry N. pumilio that separated from the rest, which resulted in an overall non-significant relationship (Fig. S4).

a Relationship between predawn (Ψpd) and midday (Ψmd) leaf water potentials measured for seedlings of Nothofagus pumilio and N. dombeyi from different provenances (See Table S1 and S2). b Relationship between the estimated values of parameters slope (s) and intercept (i) for each species and provenance

Relationship between the estimated values of parameters intercept (i) and slope (s) with temperature-related climatic variables. All correlations were significant (P > 0.05)
Molecular genetic characteristics
Studied species differed in the degree of microsatellite genetic diversity and divergence between provenances. Humid and dry N. pumilio were similarly diverse although distinct private alleles, nine and one respectively, were present at each location (Table 3). The degree of divergence between humid and dry provenances of N. pumilio was c. 2% (FST value of 0.019, 99% confidence interval = − 0.009–0.009) thus it was not significantly different from zero. Nonetheless, AMOVA analysis yielded a low but significant between-provenance divergence of 6% (Table 4). Individuals of N. dombeyi from the humid provenance were more polymorphic and had significantly greater microsatellite diversity as yielded by distinct parameters, including the presence of more private alleles than the dry provenance, seven and two, respectively (Table 3). Yet humid and dry provenances were genetically alike as shown by non-significant different from zero FST values (FST = − 0.015, 99% confidence interval = − 0.058–0.047) and the non-significant AMOVA analysis (ΦST = 3%; P = 0,22).
Discussion
Climate exerts significant selective pressures on populations of dominant Nothofagus species. Our data show significant differences between climatically-extreme provenances at the whole plant, leaf, and function, yet those were species-specific. Architectural features of plants from benign, i.e. humid, environments outgrew those under stress, i.e. dry, as shown by greater total height, the total number of leaves, and basal diameter which was consistent for both species. Given that those differences were measured under cultivation in the common garden, they can be considered genetically-based on these morphological adaptations. Declining plant growth, in dry provenances, could be an early response to the water deficit (Xu and Zhou 2005). Yet leaf traits and responses to the experimental drought were species-dependent that reflected their distinct ecological characteristics. Seedlings of N. pumilio yielded an anisohydric stomatal regulation of their water status consisting of slope values of the relationship between predawn and midday water potentials near one, whereas those of N. dombeyi seedlings tended to be isohydric with slopes close to zero. Similar leaf traits were found in N. pumilio from extreme precipitation regimes in contrast to significantly smaller leaves and lower stomatal densities measured in dry N. dombeyi that reflect distinct strategies to drought.
Anisohydric behavior of N. pumilio seedlings suggests that they can be considered drought tolerant since they may continue gas exchange by keeping their stomata open for longer periods. This result has also been observed in other deciduous species of Nothofagus inhabiting north, i.e. warm, Patagonian forests such as N. nervosa and N. obliqua (Varela et al. 2010). Although this is at risk of experiencing potentially lethal phenomena of drought-induced cavitation, N. pumilio is deciduous and has other mechanisms to avoid extreme droughts such as facultative dehiscence that may occur during the summer (Veblen et al. 1996) and an overall reduced stomatal density of less than half of that quantified using the same metrics in its mesic congener N. dombeyi. On the other hand, the sister deciduous species N. antarctica was more responsive to water stress compared with N. pumilio by higher stomatal closure and also lower SLA values which are related to a greater concentration of structural constituents (abundant sclerenchyma and bigger vascular bundles) and a thicker cuticle (Peri et al. 2009). Yet leaf traits of distinct provenances of N. dombeyi measured in cultivation clearly showed genetic differences and thus can be considered distinct ecotypes (Diaz et al. 2020).
Individuals of N. dombeyi inhabiting the drier extreme consisted of smaller stomata and less density as expected in isohydric plants (Attia et al. 2015) which in the case of the dry-most provenance can be considered extreme isohydric (slope < 0). Isohydric plants are often identified as drought avoiders and they may respond through a faster stomatal closure to evade the potentially dangerous effects of an excessive reduction in water potential due to drought (Moran et al. 2017). However, in a comparative study of the survival of the evergreen species of Nothofagus conducted by Piper et al. (2007), they found that N. dombeyi seedlings showed greater drought tolerance than those of N. nitida which in turn occupies more humid environments than the former.
It should be noted that plant responses to water deficits can be explained by various mechanisms, such as photosynthetic rate, stomatal conductance, efficiency in water use, among others. The low number of samples used for measurements and reduced duration of treatment under stress could affect the results. Similarly, other important aspects must be considered, such as the different responses during the ontogeny of plants, to transfer the knowledge of experiments with seedlings in a common garden to the forests (Niinemets 2010). Plants at distinct life stages may differ in their response to drought as documented in adults of Pinus nigra ssp. laricio that followed an isohydric strategy, whereas seedlings in a glasshouse experiment did not (Cinnirella et al. 2002). Similar results were obtained by a previous study that compared ecophysiological traits of adults under field conditions and common garden-grown seedlings to disentangle potential genetically-based differences between N. pumilio plants from contrasting elevations (Premoli and Brewer 2007). While significantly distinct ecophysiological variables related to the use of water were measured under field conditions, no such differences were found in seedlings, which was interpreted as a result of environmentally-driven controls on such traits (Premoli and Brewer 2007). A reverse situation is documented here where most plant traits analyzed on seedlings of distinct species from dry and humid provenances that were grown under common environmental settings yielded significant differences. We may speculate that such differences will probably be amplified under field conditions, and thus maintained in adults. Also, it would be important to know how populations are re-conditioned following the stress event, as distinct Nothofagus differed in their response after drought in New Zealand (Sun et al. 1995).
Mesic N. dombeyi is mostly associated with humid climates and therefore its isohydric behavior tends to narrow soil-leaf water potential gradient and hence the rate of water loss to the atmosphere, such that it matches the capacity of the soil–plant hydraulic system to supply water to leaves (Attia et al. 2015). While humid populations of N. dombeyi consisting of genotypes with larger leaves and greater stomatal densities along with drought avoidance mechanisms can well respond to their local climate, they seem to be deprived of adaptations to extreme droughts such as those recorded in the last 20 years in northern Patagonia. This has resulted in a greater susceptibility to hydraulic failure that caused massive (Suarez et al. 2004) genetically-driven mortality of humid N. dombeyi (Fasanella et al. 2021). Hence, the evolution of an isohydric behavior in N. dombeyi has resulted from its adaptation to water surplus. Yet it seems that dry N. dombeyi populations were able to persist locally through time because of divergent selection for genotypes adapted to recurrent droughts which were purged by stabilized selection towards the humid extreme (Diaz et al. 2020). Thus, while N. dombeyi can sustain viable populations even at the eastern-most extreme, those at the humid end may be vulnerable under current and future forecasted increased drought trends.
The adaptive divergence of N. dombeyi inhabiting precipitation extremes occurs in the face of continuous gene flow as reflected by non-significant than zero genetic divergence measures at neutral markers as FST and AMOVA. Also, the lack of microsatellite divergence may reflect a unique ancient gene pool that was fragmented once the precipitation gradient was established. Sequences of the chloroplast that provide a historical signal due to the maternal nature of its hereditary transmission showed that although humid and dry provenances belong to the same haplogroup (Acosta et al. 2014; Premoli et al. 2012), yet they consist of unique haplotypes (Diaz 2018). This evidence, together with the fact that humid and dry provenances presented seven and two private microsatellite alleles, suggests that populations have differentiated sometime in the past and that these differences have been maintained over time due to selection at contrasting precipitation regimes. Besides, the two humid and dry provenances exhibit significant differences in their levels of neutral diversity with greater microsatellite diversity in the former. This may be a reflection of smaller and more naturally fragmented dry populations, which suffer from the effects of genetic drift and isolation that tend to erode genetic variation. This was also found in Nothofagus species in Australia, with little divergence in microsatellite markers between the extremes of genetically based morphological and physiological altitude clines, suggesting that adaptive differentiation is strongly driven by selection because it is likely to occur in the presence of gene flow (Duncan et al. 2016).
On the other hand, populations of N. pumilio maintain significant within-population microsatellite diversity and divergence between humid and dry provenances. Long-term persistence throughout the range under distinct environmental settings has produced the accumulation of genetic differences as depicted by the low yet significant between-provenance divergence yielded by AMOVA and the existence of private microsatellite alleles at both precipitation extremes. Given that N. pumilio is a mountain species, it is found on the Andes to the west, i.e. humid, and on mountain ranges that run parallel towards the east, and no individual populations are found in the lowlands. Thus, genetic divergence results from isolation between such allopatric populations inhabiting extreme conditions. Furthermore, the analysis of 37 and 33 populations of N. pumilio from humid and dry provenances, respectively, using DNA sequences of the chloroplast yielded four new variants for the species (Ignazi 2019) compared to previous studies (Mathiasen and Premoli 2010; Premoli et al. 2012). One and three of these new haplotypes were exclusive to the humid and dry provenances, respectively, and one out of the last three ones was private to the entire Nothofagus subgenus (Ignazi 2019). This highlights the relevance of dry-most N. pumilio populations not only in relation to the presence of adaptive variants with the ability to respond to drought but also as reservoirs of genetic diversity and thus as evolutionary novelties. Nonetheless, equally diverse N. pumilio populations suggest that genetically viable populations were maintained even under very distinct climates fostered by an overall greater tolerance to desiccation, i.e. anisohydric behavior. However, it has been suggested that N. pumilio forests currently located in relatively mesic provenances will become more susceptible to decline (Rodríguez-Catón et al. 2016). Although anisohydric behavior would be beneficial under conditions of minimal to moderate stress, it would not necessarily be so under prolonged drought (Sade et al. 2012). More frequent and intense extreme weather events, such as droughts, are forecasted for northern Patagonia which most probably will increase water deficit in eastern-most N. pumilio stands (Rodríguez-Catón et al. 2016). Thus, experiments with distinct levels of imposed drought would improve our understanding of the potential responses of tree species under predicted increased desiccation.
Descriptors as the slope and the intercept of the relationship between predawn and midday leaf water potentials were significantly associated with temperature variables of the climates inhabited by both species. However, only populations of the mesic N. dombeyi inhabiting drier areas yielded pressure drops under drier climates while that of N. pumilio remain fairly constant as expected in strict anisohydric plants. It was recently suggested that the water potential is determined not only by stomatal responses but also by the vulnerability of the plant hydraulic system which can be portrayed by the slope of the relationship between the water potential in leaves and soil using midday and predawn leaf water potentials as proxies of each of them, respectively. Thus, the smaller the slope values the more vulnerable the population and/or species is to suffer from xylem embolism (Martinez-Vilalta et al. 2014) as is the case of humid N. dombeyi. On the contrary, in dry N. dombeyi this vulnerability may be compensated by greater elasticity of the cell-wall (Diaz et al. 2020). Cell walls with higher elasticity will be more sensitive to small changes in water-content differences, resulting in faster closure of the stomata and thus avoiding cell dehydration (Sade et al. 2012). In a recent genomic study, we found that distinct genotypes for adaptive single nucleotide polymorphisms, i.e. SNPs, were present in N. dombeyi seedlings from humid and dry provenances which in turn can be associated with genes involved in the extensibility of the cell wall (Diaz et al. 2020). Also, although no osmotic adjustments were measured under water deficit treatments, a substantial change in cell wall properties resulted in an elastic adjustment, i.e. more rigid cell walls, as evidenced in humid and dry seedlings of N. pumilio under water deficit (Ignazi et al. 2020). This is consistent with studies of distinct Nothofagus species where populations from the driest extreme appeared less vulnerable than those grown in the wettest sites, but there were species- and population-specific compensatory mechanisms such as higher water storage and hydraulic efficiency that reduced the water potential drop and thus the probability of embolism formation in stems (Bucci et al. 2012).
In the face of global climate change, ecological protection and management of dry habitats are of paramount importance, reducing for example other extrinsic pressures (Moritz et al. 2012), to preserve existing adaptive diversity (Kapeller et al. 2016). This knowledge could also be used for restoration purposes, minimizing the effects of climate change through the introduction of individuals adapted from marginal, i.e. dry-most populations, to promote the faster adaptation of the general population (Thiel et al. 2014). While local sources were advocated as the best choice for restoration, this measure should be revisited under climate change given that in the case of humid provenances as of N. dombeyi local sources will probably have lower performance, i.e. reduced acclimation potential, under forecasted increased drought. Under this scenario, it was suggested that ‘regional admixture provenancing’ may be the hint for long-term persistence as a compromise strategy where seeds are collected from multiple populations within the same region (Bucharova et al. 2018). In the particular case of N. dombeyi, geographically structured chloroplast DNA lineages can be used to define regional sources (e.g. Premoli et al. 2012), and localities can be ascribed concerning adaptive responses to local climates and genetically divergent populations. Therefore, we propose that actions to manage and restore species and populations susceptible to abiotic stress, such as N. dombeyi at the humid end, should be carried out by mixing propagule sources considering both local and regional levels (Fig. 7).

Diagram representing climate-driven adaptive management and restoration recommendations. For explanation see text. Pictures at the bottom represent pure Nothofagus dombeyi (left), landscape view of the altitudinal zonation of low-elevation N. dombeyi and high-elevation N. pumilio (center), and pure N. pumilio (right)
Our results are in line with previous studies of tree species growing under Mediterranean climates with summer droughts (Mediavilla and Escudero 2003) such that a conservative use of water, i.e. isohydric, avoiding desiccation characterize the evergreen N. dombeyi, whereas the deciduous N. pumilio tend to be more drought-resistant, i.e. anisohydric. This information seeks to contribute to our understanding of species-specific tree strategies under ongoing climate change. Thus, deciduous anisohydric species may have the ability to cope with drought in addition to other mechanisms such as early partial crown defoliation during the summer. In contrast, isohydric species that have evolved in mesic climates may suffer from growth reduction that may affect tree survival under water shortage. “Forest decline” and “forest dieback” are processes that are globally affecting forests’ health and, in some increasingly frequent cases, inducing massive tree mortality (Allen et al. 2010). Thus, we need studies on different forest types, deciduous and evergreen, that analyze responses to increased drought around the globe.
Conclusions
Our results show the genetic basis of character variation and that adjustments of plant species and populations to distinct ecological settings facing unusual local and regional climates need to be considered under management. While under water stress some taxa may be vulnerable, i.e. as the isohydric N. dombeyi, others will be naturally buffered and resilient to climate-influenced disturbances as the anisohydric N. pumilio. Climate projections for Patagonia include frequent and more extreme precipitation shortages which have shown to produce massive N. dombeyi decay in humid areas. Therefore, active actions should be taken, in the short term, to restore populations of N. dombeyi affected by extreme drought. Moreover, if climate trends remain as predicted, in the long term, even anisohydric plants as N. pumilio will be vulnerable to prolonged droughts.
References
Acosta MC, Mathiasen P, Premoli AC (2014) Retracing the evolutionary history of Nothofagus in its geo-climatic context: new developments in the emerging field of phylogeology. Geobiology 12(6):497–510. https://doi.org/10.1111/gbi.12098
Alberdi M (1987) Ecofisiología de especies chilenas del género Nothofagus. Bosque 8(2):77–84. https://doi.org/10.4206/bosque.1987.v8n2-02
Allen CD, Macalady AK, Chenchouni H, Bachelet D et al (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259(4):660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Arbetman MP, Premoli AC (2011) Oldies (but goldies!): extracting DNA from cryopreserved allozyme homogenates. J Hered 102(6):764–769. https://doi.org/10.1093/jhered/esr096
Attia Z, Domec JC, Oren R, Way DA, Moshelion M (2015) Growth and physiological responses of isohydric and anisohydric poplars to drought. J Exp Bot 66(14):4373–4381. https://doi.org/10.1093/jxb/erv195
Barros VR, Boninsegna JA, Camilloni IA, Chidiak M, Magrín GO, Rusticucci M (2014) Climate change in Argentina: trends, projections, impacts and adaptation. WIREs Clim Change 6(2):151–169. https://doi.org/10.1002/wcc.316
Bucci SJ, Scholz FG, Campanello PI, Montti L, Jimenez-Castillo M, Rockwell FA, La Manna L, Guerra P, Lopez Bernal P, Troncoso O, Enricci J, Holbrook MN, Goldstein G (2012) Hydraulic differences along the water transport system of South American Nothofagus species: do leaves protect the stem functionality? Tree Physiol 32(7):880–893. https://doi.org/10.1093/treephys/tps054
Bucharova A, Bossdorf O, Hölzel N, Kollmann J, Prasse R, Durka W (2018) Mix and match: regional admixture provenancing as the golden mean between seed-sourcing strategies for ecological restoration. Conserv Genet. https://doi.org/10.1007/s10592-018-1067-6
Choat B, Brodribb TJ, Brodersen CR, Duursma RA, López R, Medlyn BE (2018) Triggers of tree mortality under drought. Nature 558(7711):531–539. https://doi.org/10.1038/s41586-018-0240-x
Cinnirella S, Magnani F, Saracino A, Borghetti M (2002) Response of a mature Pinus laricio plantation to a three-year restriction of water supply: structural and functional acclimation to drought. Tree Physiol 22(1):21–30. https://doi.org/10.1093/treephys/22.1.21
Cruz de Souza B, Carvalho ECD, Oliveira RS, de Araujo FS, de Lima ALA, Rodal MJN (2020) Drought response strategies of deciduous and evergreen woody species in a seasonally dry neotropical forest. Oecologia 194(1):221–236. https://doi.org/10.1007/s00442-020-04760-3
Diaz DG (2018) Potenciales respuestas diferenciales a la sequía y su base genética en Nothofagus dombeyi. Degree thesis, Universidad Nacional del Comahue, Bariloche, Argentina
Diaz DG, Mathiasen P, Premoli AC (2020) Subtle precipitation differences yield adaptive adjustments in the mesic Nothofagus dombeyi. For Ecol Manag 461:117931. https://doi.org/10.1016/j.foreco.2020.117931
Dimitri MJ (1972) La Región de los Bosques Andino-Patagónicos. Sinopsis General. Colección Científica X. Buenos Aires: INTA
Donoso C (1995) Bosques templados de Chile y Argentina, 3ra edn. Editorial Universitaria, Santiago
Donoso C, Premoli A, Donoso P (2004) Variación en Nothofagus siempreverdes sudamericanos. In: Donoso C, Premoli AC, Gallo LA, Iliniza R (eds) Variación intraespecífica en las especies arbóreas de los bosques templados de Chile y Argentina. Editorial Universitaria, Santiago de Chile, pp 145–166
Dumolin S, Demesure B, Petit RJ (1995) Inheritance of chloroplast and mitochondrial genomes in pedunculate oak investigated with an efficient PCR method. Theoret Appl Genet 91(8):1253–1256. https://doi.org/10.1007/BF00220937
Duncan CJ, Worth JRP, Jordan GJ, Jones RC, Vaillancourt RE (2016) Genetic differentiation in spite of high gene flow in the dominant rainforest tree of southeastern Australia, Nothofagus cunninghamii. Heredity 116(1):99–106. https://doi.org/10.1038/hdy.2015.77
Fady B, Cottrell J, Ackzell L, Alía R, Muys B, Prada A, González-Martínez SC (2016) Forests and global change: what can genetics contribute to the major forest management and policy challenges of the twenty-first century? Reg Environ Change 16(4):927–939. https://doi.org/10.1007/s10113-015-0843-9
Fasanella M, Suarez MLL, Hasbun R, Premoli AC (2021) Individual-based dendrogenomic analysis of forest dieback driven by extreme droughts. Can J For Res (ja). https://doi.org/10.1139/cjfr-2020-0221
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol 37(12):4302–4315. https://doi.org/10.1002/joc.5086
Goudet J (2001) FSTAT, a program to estimate and test gene diversities and fixation indices, version 2.9. 3. http://www2.unil.ch/popgen/softwares/fstat.htm
Guichoux E, Lagache L, Wagner S, Chaumeil P, Léger P, Lepais O, Lepoittwvin C, Malausa T, Revardel E, Salin F, Petit RJ (2011) Current trends in microsatellite genotyping. Mol Ecol Resour 11(4):591–611. https://doi.org/10.1111/j.1755-0998.2011.03014.x
Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh KA (2001) Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126(4):457–461. https://doi.org/10.1007/s004420100628
Hampe A, Jump AS (2011) Climate relicts: past, present, future. Annu Rev Ecol Evol Syst 42:313–333. https://doi.org/10.1146/annurev-ecolsys-102710-145015
Hamrick JL (2004) Response of forest trees to global environmental changes. For Ecol Manag 197(1–3):323–335. https://doi.org/10.1016/j.foreco.2004.05.023
Ignazi G (2019) Variación genética en poblaciones marginales de Nothofagus pumilio, su relación con procesos ecológico-evolutivos e importancia en la conservación. PhD. Thesis, Universidad Nacional del Comahue, Bariloche, Argentina
Ignazi G, Bucci SJ, Premoli AC (2020) Stories from common gardens: water shortage differentially affects Nothofagus pumilio from contrasting precipitation regimes. For Ecol Manag 458:117796. https://doi.org/10.1016/j.foreco.2019.117796
IPCC (2014) Cambio climático 2014: impactos, adaptación y vulnerabilidad – Resumen para responsables de políticas. In: Field CB, Barros VR, Dokken DJ, Mach KJ, Mastrandrea MD, Bilir TE, Chatterjee M, Ebi KL, Estrada YO, Genova RC, Girma B, Kissel ES, Levy AN, MacCracken S, Mastrandrea PR, White LL (eds) Contribución del Grupo de trabajo II al Quinto Informe de Evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático. Organización Meteorológica Mundial, Ginebra, Suiza
Jones RC, Vaillancourt RE, Jordan GJ (2004) Microsatellites for use in Nothofagus cunninghamii (Nothofagaceae) and related species. Mol Ecol Notes 4(1):14–16. https://doi.org/10.1046/j.1471-8286.2003.00558.x
Kapeller S, Dieckmann U, Schueler S (2016) Varying selection differential throughout the climatic range of Norway spruce in Central Europe. Evol Appl 10(1):25–38. https://doi.org/10.1111/eva.12413
Marchelli P, Caron H, Azpilicueta MM, Gallo LA (2008) Primer note: a new set of highly polymorphic nuclear microsatellite markers for Nothofagus nervosa and related South American species. Silvae Genet 57(1–6):82–85. https://doi.org/10.1515/sg-2008-0013
Martínez-Vilalta J, Poyatos R, Aguadé D, Retana J, Mencuccini M (2014) A new look at water transport regulation in plants. New Phytol 204(1):105–115. https://doi.org/10.1111/nph.12912
Mathiasen P, Premoli AC (2010) Out in the cold: genetic variation of Nothofagus pumilio (Nothofagaceae) provides evidence for latitudinally distinct evolutionary histories in austral South America. Mol Ecol 19(2):371–385. https://doi.org/10.1111/j.1365-294X.2009.04456.x
Mathiasen P, Premoli AC (2013) Fine-scale genetic structure of Nothofagus pumilio (lenga) at contrasting elevations of the altitudinal gradient. Genética 141(1–3):95–105. https://doi.org/10.1007/s10709-013-9709-6
Mathiasen P, Venegas-González A, Fresia P, Premoli AC (2020) A relic of the past: current genetic patterns of the palaeoendemic tree Nothofagus macrocarpa were shaped by climatic oscillations in central Chile. Ann Bot mcaa. https://doi.org/10.1093/aob/mcaa111
Mazzarino MJ, Bertiller MB, Schlichter T, Gobbi ME (1998) Nutrient cycling in Patagonian ecosystems. Ecol Austral 8(2):167–181
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T et al (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178(4):719–739. https://doi.org/10.1111/j.1469-8137.2008.02436.x
MedCalc Software (2015) Digimizer (image analysis software) version 4.6.1. www.digimizer.com
Mediavilla S, Escudero A (2003) Stomatal responses to drought at a Mediterranean site: a comparative study of co-occurring woody species differing in leaf longevity. Tree Physiol 23(14):987–996. https://doi.org/10.1093/treephys/23.14.987
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17(8):2145–2151. https://doi.org/10.1890/06-1715.1
Moran E, Lauder J, Musser C, Stathos A, Shu M (2017) The genetics of drought tolerance in conifers. New Phytol 216(4):1034–1048. https://doi.org/10.1111/nph.14774
Moritz C, Langham G, Kearney M, Krockenberger A, VanDerWal J, Williams S (2012) Integrating phylogeography and physiology reveals divergence of thermal traits between central and peripheral lineages of tropical rainforest lizards. Philos Trans R Soc B: Biol Sci 367(1596):1680–1687. https://doi.org/10.1098/rstb.2012.0018
Moshelion M, Halperin O, Wallach R, Oren RAM, Way DA (2015) Role of aquaporins in determining transpiration and photosynthesis in water-stressed plants: crop water-use efficiency, growth and yield. Plant Cell Environ 38(9):1785–1793. https://doi.org/10.1111/pce.12410
Neukom R, Steiger N, Gómez-Navarro JJ, Wang J, Werner JP (2019) No evidence for globally coherent warm and cold periods over the preindustrial Common Era. Nature 571(7766):550–554. https://doi.org/10.1038/s41586-019-1401-2
Niinemets Ü (2010) Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: past stress history, stress interactions, tolerance and acclimation. For Ecol Manag 260(10):1623–1639. https://doi.org/10.1016/j.foreco.2010.07.054
Novaes RML, Rodrigues JG, Lovato MB (2009) An efficient protocol for tissue sampling and DNA isolation from the stem bark of Leguminosae trees. Genet Mol Res 8(1):86–96. https://doi.org/10.4238/vol8-1gmr542
Peakall ROD, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6(1):288–295. https://doi.org/10.1111/j.1471-8286.2005.01155.x
Peri PL, Pastur GM, Lencinas MV (2009) Photosynthetic response to different light intensities and water status of two main Nothofagus species of southern Patagonian forest Argentina. J For Sci 55(3):101–111. https://doi.org/10.17221/66/2008-JFS
Phillips SJ, Dudík M, Schapire RE (2021) Maxent software for modeling species niches and distributions (Version 3.4.1). http://biodiversityinformatics.amnh.org/open_source/maxent/. Accessed 25 Feb 2021
Piper FI, Corcuera LJ, Alberdi M, Lusk C (2007) Differential photosynthetic and survival responses to soil drought in two evergreen Nothofagus species. Ann For Sci 64(4):447–452. https://doi.org/10.1051/forest:2007022
Premoli AC (1994) Genetic, morphological, and ecophysiological variation in geographically restricted and widespread species of Nothofagus from southern South America. Ph.D. Thesis. University of Colorado, Boulder
Premoli AC (2003) Isozyme polymorphisms provide evidence of clinal variation with elevation in Nothofagus pumilio. J Hered 94(3):218–226. https://doi.org/10.1093/jhered/esg052
Premoli AC, Brewer CA (2007) Environmental v. genetically driven variation in ecophysiological traits of Nothofagus pumilio from contrasting elevations. Austral J Bot 55(6):585–591. https://doi.org/10.1071/BT06026
Premoli AC, Mathiasen P, Cristina Acosta M, Ramos VA (2012) Phylogeographically concordant chloroplast DNA divergence in sympatric Nothofagus ss How deep can it be? New Phytol 193(1):261–275. https://doi.org/10.1111/j.1469-8137.2011.03861.x
Premoli AC, Raffaele E, Mathiasen P (2007) Morphological and phenological differences in Nothofagus pumilio from contrasting elevations: evidence from a common garden. Austral Ecol 32(5):515–523. https://doi.org/10.1111/j.1442-9993.2007.01720.x
Reichstein M, Bahn M, Ciais P, Frank D, Mahecha MD, Seneviratne SI, Papale D (2013) Climate extremes and the carbon cycle. Nature 500(7462):287–295. https://doi.org/10.1038/nature12350
Rodríguez-Catón M, Villalba R, Morales M, Srur A (2016) Influence of droughts on Nothofagus pumilio forest decline across northern Patagonia, Argentina. Ecosphere 7(7):e01390. https://doi.org/10.1002/ecs2.1390
Rodríguez-Catón M, Villalba R, Srur A, Williams AP (2019) Radial growth patterns associated with tree mortality in Nothofagus pumilio forest. Forests 10(6):489. https://doi.org/10.3390/f10060489
Sade N, Gebremedhin A, Moshelion M (2012) Risk-taking plants: anisohydric behavior as a stress-resistance trait. Plant Signal Behav 7(7):767–770. https://doi.org/10.4161/psb.20505
Shao HB, Chu LY, Jaleel CA, Zhao CX (2008) Water-deficit stress-induced anatomical changes in higher plants. CR Biol 331(3):215–225. https://doi.org/10.1016/j.crvi.2008.01.002
Sork VL (2016) Gene flow and natural selection shape spatial patterns of genes in tree populations: implications for evolutionary processes and applications. Evol Appl 9(1):291–310. https://doi.org/10.1111/eva.12316
StatSoft, Inc. (2004) STATISTICA (data analysis software system), version 7. www.statsoft.com
Suarez ML, Ghermandi L, Kitzberger T (2004) Factors predisposing episodic drought-induced tree mortality in Nothofagus: site, climatic sensitivity and growth trends. J Ecol 92:954–966
Sun OJ, Sweet GB, Whitehead D, Buchan GD (1995) Physiological responses to water stress and waterlogging in Nothofagus species. Tree Physiol 15(10):629–638. https://doi.org/10.1093/treephys/15.10.629
Swofford DL, Selander RB (1989) BIOSYS-1: a computer program for the analysis of allelic variation in population genetics and biochemical systematics
Thiel D, Kreyling J, Backhaus S, Beierkuhnlein C, Buhk C, Egen K, Huber G, Konnert M, Nagy L, Jentsch A (2014) Different reactions of central and marginal provenances of Fagus sylvatica to experimental drought. Eur J Forest Res 133(2):247–260. https://doi.org/10.1007/s10342-013-0750-x
Varela SA, Gyenge JE, Fernandez ME, Schlichter T (2010) Seedling drought stress susceptibility in two deciduous Nothofagus species of NW Patagonia. Trees 24(3):443–453. https://doi.org/10.1007/s00468-010-0412-2
Veblen TT, Kitzberger T, Burns BR (1996) Natural disturbance and regeneration dynamics in Andean forests of southern Chile and Argentina. In: Armesto JJ, Arroyo MTK, Villagrán C (eds) Ecology of Southern Chilean and Argentinean Nothofagus Forests. Universidad de Chile, Santiago de Chile, pp 169–198
Xu ZZ, Zhou GS (2005) Effects of water stress and nocturnal temperature on carbon allocation in the perennial grass, Leymus chinensis. Physiol Plant 123(3):272–280. https://doi.org/10.1111/j.1399-3054.2005.00455.x
Zar JH (1996) Biostatistical analysis. Pearson Education, Bangalore
Acknowledgements
We are grateful to P. Alvear for field assistance and P. Suarez INIBIOMA-CONICET for assistance during plant cultivation in common garden. Ramiro Ripa assisted with pressure chamber. We thank Administración de Parques Nacionales for extending permits to work within protected areas. This research was funded by Agencia de Promoción Científica y Tecnológica PICT-2015-1565 from Argentina and The Rufford Foundation, United Kingdom project 29211-1.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Diaz, D.G., Ignazi, G., Mathiasen, P. et al. Climate-driven adaptive responses to drought of dominant tree species from Patagonia. New Forests 53, 57–80 (2022). https://doi.org/10.1007/s11056-021-09843-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-021-09843-4