
Abstract
We determined the effects of two nitrogen sources (ammonium and nitrate) and two irradiance levels (50 and 200 μmol photons m−2 s−1) on the growth rate, cell size, proximate composition, pigment content, and photosynthesis of the unicellular red alga, Porphyridium cruentum. Irradiance significantly affects growth rate, as well as carbohydrate, protein, and phycoerythrin content. Nitrogen form significantly affects cell size, total dry weight, organic dry weight, ash content, carotene content, phycocyanin content, allophycocyanin content, maximum relative electron transport rate (rETRm), and photosynthetic efficiency (α). However, the irradiance and nitrogen source had significantly interaction with the content of lipids and chlorophyll a content, relative electron transport rate (rETR), and irradiance of saturation (Ik). These findings demonstrate that irradiance and nitrogen source influence the metabolism of P. cruentum and that the combination of these two variables induces the production of chemical products for biotechnological, aquaculture, and nutraceutical industry.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The use of microalgae was documented in China 2000 years ago where they consumed the cyanobacterium Nostoc as food (Spolaore et al. 2006). In Mexico, the Aztecs built raised fields called chinampas and ate Spirulina (= Arthrospira) mixed with toasted corn tortillas and chilli sauce; this food was named “Tecuitlatl” (Paniagua-Michel et al. 1993; Pulz and Gross 2004). Biotechnological development of microalgae has increased exponentially in the past 20 years (Grewe and Pulz 2012). Several products can be obtained from microalgal cultures, including pigments, lipids, glycerol, and carbohydrates (Pulz and Gross 2004; Spolaore et al. 2006). Recently, synthetic pigment production has decreased, because such compounds cause problems for human health (Ahlström et al. 2005). Thus, the generation of natural organic pigments has become important for use in cosmetology, pharmacology, and food production (Arad and Yaron 1992; Ruiz-Ruiz et al. 2013). Several pigments that are produced by microalgae cells, such as chlorophylls, carotenoids, and phycobiliproteins, have garnered attention with regard to their industrial use (Cuellar-Bermúdez et al. 2014). Pigments have a significant function in photosynthesis by algae and perform many beneficial activities, having antioxidant, anticarcinogenic, anti-inflammatory, antiobesity, antiangiogenic, and neuroprotective effects (Romay et al. 2003; Guedes et al. 2011; Pangestuti and Kim 2011; Yen et al. 2013).
The unicellular rhodophyte Porphyridium cruentum (S.F. Gray) Nägeli is a microalgae strain with spherical cells, which lack a cell wall, and synthesize an accessory pigment (phycoerythrin) that gives them their red color (Gantt 1969, 1981). This microalga is used as a source of sulfated exopolysaccharides and superoxide dismutase (Misra and Fridovich 1977; Vonshak 1988) for application in the food and pharmaceutical industries. Porphyridium cruentum cells have a rich biomass nutrient profile, especially regarding their content of carbohydrates, eicosapentaenoic acid (EPA), minerals, energy, and proteins, and do not produce toxic or antinutritional factors when used as a feed supplement (Rebolloso-Fuentes et al. 2000; Razaghi et al. 2014; Kavitha et al. 2016). Also, P. cruentum contains phycoerythrin, which is used as a fluorescent pigment in immunoassays (Curtin 1985) and can be used as a supplement to carotenes, tocopherol, vitamins (Antia et al. 1970), and arachidonic acid (ARA) (Ahern et al. 1983). The exopolysaccharides and glycerol produced by P. cruentum can be used as osmotic eliminators that concentrate live microalgae cells during harvest (Mazzuca-Sobczuc et al. 2015).
Biomass production by microalgae cultures and their composition are influenced by several environmental variables (Muller-Feuga et al. 2007), the most significant of which are temperature (Oh et al. 2009), light (Dubinsky and Stambler 2009; Villafane et al. 2005), and nitrogen source (Gigova and Ivanova 2015). Light is the driving source of photosynthesis, and irradiance and spectral composition can alter the metabolism and biochemical composition of microalgal cultures (Falkowski and Raven 2007; Vadiveloo et al. 2017).
Nitrogen is the major nutrient required for photosynthetic organisms and is used for the synthesis of chlorophyll and other photosynthetic pigments, amino acids, nucleic acids, and coenzymes (Hu 2005; Raven and Giodano 2016). The metabolism of microalgae varies, depending on nitrogen availability and nitrogen source (Gigova and Ivanova 2015). Nitrate is the most common dissolved form of nitrogen in seawater, but many microalgae prefer to use ammonium as a nutrient (Miller and Castenholtz 2001; Lourenço et al. 2002).
We hypothesized that the light irradiance level and chemical nitrogen source that is supplied in the medium can be adjusted to alter the proximate composition, pigment content, and photosynthesis of P. cruentum for applications in biotechnology, aquaculture, and the nutraceutical industry.
Materials and methods
Growth conditions
The strain of Porphyridium cruentum (S.F. Gray) Nägeli [= P. purpureum (Bory de Saint-Vicent) K.M. Drew and R. Ross] was obtained from the Institute of Applied Microbiology (IAM), Tokyo Bunkyo-Ku University, Japan.
Nonaxenic, monospecific batch cultures were maintained in 12 Erlenmeyer flasks of 125 mL with 100 mL of “f” medium in seawater (Guillard and Ryther 1962). The nitrogen of “f” medium provided by sodium nitrate (NO3: nitrate) and used as control was replaced with ammonium chloride (NH4: ammonium), considering the atomic ratio of the active ingredient that was provided by the nitrogen source in the control medium. The concentration final of nitrogen was 17.66 × 10−4 M and the N:P atomic ratio for both nitrogen sources added to the media were 22:1. The medium was prepared with filtered seawater (1-μm pore) and sterilized at 121 °C and 1.05 kg cm−2. The culture conditions were as follows: temperature 20 ± 1 °C, salinity 33 ± 1‰, and daily manual stirring without aeration. The pH was measured daily at the initial values of 7.9 that increase throughout the culture time to reach values of 9.0. Light was provided by 40 W white fluorescent lamps (Philips F40 T12/DX) at two irradiances—50 and 200 μmol photons m−2 s−1. The two irradiances were obtained adjusting the distance of the culture to the lamp and were measured at the center of the surface of the Erlenmeyer flasks with a 4π QSL-100 quantum radiometer (Biospherical Instruments, USA). Each culture condition (nutrients and light) was maintained in triplicate Erlenmeyer flasks.
Growth rate and cell size
The cell concentration was measured daily by direct count using a hemocytometer. Clumps of cells were disaggregated by ultrasonication for 5 min at 100 kHz with an L&R Solid State/Ultrasonic model T−9B (L&R Manufacturing Company, USA). The cell concentrations in the exponential growth phase were log2-transformed to determine growth rates (μ) as per Fogg and Thake (1987).
Cell size was measured on day 7 of the culture. We randomly selected 30 cells and measured the diameter under a compound microscope using a digital camera and Image Pro-Discovery software (version 5.1).
Dry weight and proximate composition
The dry weight, proximate composition, and pigment content of the cells were measured in the exponential growth phase (day 7).
To measure the total dry weight (TDW), 25 mL of each culture was passed through washed and preweighed 47-mm Whatman GF/C glass microfiber filters (~1-μm pore size), rinsed with 15 mL ammonium formate (3%) to remove salt residues, and dried at 60 °C. To measure the ash content (ASH), the filters with cell biomass were incinerated at 450 °C for 4 h. The organic dry weight (ODW) was calculated as the difference between the total dry weight and ash content per Sorokin (1973).
To determine the proximate composition of P. cruentum triplicate, 10-mL samples were passed through washed and preweighed 25-mm Whatman GF/C glass microfiber filters and stored at − 20 °C for further analysis. Water-soluble proteins were extracted with 2 N NaOH at pH 12 and 40 °C for 60 min with manual grinding as described by Safi et al. (2014), and the content was quantified per Lowry et al. (1951). A calibration curve was generated using bovine serum albumin (98%) as the standard. Carbohydrates were extracted per Whyte (1987) and quantified by the phenol-sulfuric method (Dubois et al. 1956) using glucose (99%) as the standard. Lipids were extracted per Bligh and Dyer (1959) and Chiaverini (1972) and quantified as described by Pande et al. (1963). A calibration curve for lipids was generated using tripalmitin (99%) as standard.
Pigments and photosynthesis
Chlorophyll a and carotenoid content was measured on day 7 per Parsons et al. (1984). Phycocyanin, allophycocyanin, and phycoerytrin were extracted with sodium phosphate (10 mM, pH 7.0) and sonicated for 30 min, and the concentrations were determined per Bennet and Bogorad (1973).
The photosynthetic activity during each treatment was measured on day 7 in a 50-mL sample that was concentrated by centrifugation at 4000 rpm for 5 min at 20 °C, to obtain the same cell concentration. Photosynthetic activity was analyzed through rapid light curves using a pulse amplitude-modulation (PAM) fluorometer (Walz, Junior PAM). Relative electron transport rate (rETR) and the maximum quantum yield of photosystem II (Fv/Fm) values to each treatment were obtained (White et al. 2011). The following photosynthetic parameters were obtained from each photosynthetic curve: maximum relative electron transport rate (rETRm), photosynthetic efficiency (α), and saturating irradiance (Ik), as described by Figueroa et al. (2003).
Statistical analysis
All data were tested for homoscedasticity and normality (Sokal and Rohlf 1995). Differences in cell concentration by treatment were examined by Kruskal-Wallis nonparametric analysis of variance (one-way ANOVA). Differences to each variable analyzed were compared by two-way analysis of variance (ANOVA) considering as levels of the test irradiance and nitrogen source. When significant differences were obtained, Tukey a posteriori test was performed. The significance level was set to P < 0.05 in all cases.
Results
Growth rate and cell size
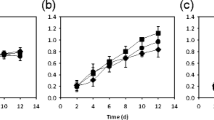
The concentrations of P. cruentum cells under the various treatments differed significantly (P < 0.013; Fig. 1), increasing rapidly during the first several days of culture, depending on the conditions. The cell density was highest with 200 μmol photons m−2 s−1 and nitrate (8.48 × 106 cells mL−1) (Fig. 1).

Growth curves of Porphyridium cruentum cultures maintained under two irradiances (50 and 200 μmol photons m−2 s−1) and two nitrogen sources (ammonium: NH4 and nitrate: NO3). The arrow indicates the day of collection of the samples (day 7), and values with lowercase letters indicate significant differences measured at this day between treatments by Kruskal-Wallis test, α = 0.05: a > b > c > d. Mean values ± SD, n = 3
The growth rate was significantly affected by the irradiance (P < 0.0001) but not by nitrogen source (P = 0.368) without interaction between irradiance and nitrogen source (P = 0.068) (Fig. 2 and Table 2). The higher value of growth rate was found at 200 μmol photon m−2 s−1 with nitrate (0.80 divisions day−1) (Fig. 2 and Table 2).

Growth rate and cell size of Porphyridium cruentum cultures maintained under two irradiances (50 and 200 μmol photons m−2 s−1) and two nitrogen sources (ammonium: NH4 and nitrate: NO3). Values with lowercase letters indicate significant differences between irradiances. Capital letters indicate significant differences between nitrogen sources by two-way ANOVA and Tukey a posteriori test, α = 0.05: a > b and A > B. Mean values ± SD, n = 3
Cell size was affected significantly by nitrogen source (P < 0.0001; Fig. 2 and Table 2) but not by irradiance (P = 0.125) without interaction by these variables (P = 0.211). Porphyridium cruentum cells were larger with ammonium at 200 μmol photons m−2 s−1 (13.19 μm) and 50 μmol photons m−2 s−1 (15.23 μm). Conversely, the diameters were smallest in cultures with nitrate at 200 μmol photons m−2 s−1 (11.54 μm) and 50 μmol photons m−2 s−1 (11.24 μm) (Fig. 2 and Table 2).
Dry weight and proximate composition
TDW (P = 0.578), ODW (P = 0.104), and ASH (P = 0.765) were not significantly affected by irradiance. However, TDW (P = 0.009), ODW (P = 0.017), and ASH (P = 0.016) were modified by effect of nitrogen source (Fig. 3 and Table 2). Interaction between irradiance and nitrogen source did not affect TDW (P = 0.311), ODW (P = 0.920), and ASH (P = 0.405) (Fig. 3 and Table 2). TDW (372.70 pg cell−1), ODW (158.71 pg cell−1), and ASH (213.99 pg cell−1) contents were highest in cultures that were maintained at 50 μmol photons m−2 s−1 with ammonium.

Proximate composition, total dry weight (TDW), organic dry weight (ODW), and ash content (ASH) of Porphyridium cruentum cultures maintained under two irradiances (50 and 200 μmol photons m−2 s−1) and two nitrogen sources (ammonium: NH4 and nitrate: NO3). Values with lowercase letters indicate significant differences between irradiances. Capital letters indicate significant differences between nitrogen source by two-way ANOVA and Tukey a posteriori test, α = 0.05: a > b and A > B. Mean values ± SD, n = 3
The proximate composition of P. cruentum was modified by irradiance and nitrogen source (Fig. 3 and Table 2). Protein content was significantly modified by effect of irradiance (P = 0.0008), but not by nitrogen source (P = 0.099) and with significant of these factors (P = 0.0212) (Fig. 3 and Table 2). The highest protein content was at 200 μmol photons m−2 s−1 with ammonium (10.45%). Carbohydrate content was significantly modified by irradiance (P = 0.001) (Fig. 3 and Table 2). The highest carbohydrate content was at 50 μmol photons m−2 s−1 with nitrate (37.23%). Lipid content was affected by irradiance (P = 0.004), nitrogen source (P = 0.031) with significant interaction of these variables (P = 0.002) (Fig. 3 and Table 2). The highest lipid content was at 50 μmol photons m−2 s−1 with ammonium (18.46%).
Pigments and photosynthesis
Chlorophyll a content was significantly affected by irradiance (P = 0.016) and nitrogen source (P = 0.002) with a significant interaction (P = 0.015) (Fig. 4 and Table 2). The carotenoid content was significantly affected by the nitrogen source (P = 0.002) but not by irradiance (P = 0.264) (Fig. 4 and Table 2).

Pigment concentration of Porphyridium cruentum cultures maintained under two irradiances (50 and 200 μmol photons m−2 s−1) and two nitrogen sources (ammonium: NH4 and nitrate: NO3). Values with lowercase letters indicate significant differences between irradiances. Capital letters indicate significant differences between nitrogen sources by two-way ANOVA and Tukey a posteriori test, α = 0.05: a > b and A > B. Mean values ± SD, n = 3
The phycocyanin (P = 0.008) and allophycocyanin (P = 0.021) content was only significantly influenced by the nitrogen source (Fig. 4 and Table 2), whereas the phycoerythrin content was significantly affected by irradiance level (P = 0.0015) (Fig. 4 and Table 2).
The rETR curves were significantly affected by the nitrogen source (P = 0.010; Fig. 5), with higher values occurring with nitrate. The photosynthetic parameters also differed between irradiance levels and nitrogen sources (Tables 1 and 2). The rETRm (P = 0.0002) and photosynthetic efficiency (α) (P = 0.0001) were affected by nitrogen source (Tables 1 and 2). The saturating irradiance (Ik) and Fv/Fm were unaltered by the irradiance (P > 0.05) or nitrogen source (P > 0.05) (Tables 1 and 2).

Photosynthetic curves measured as relative electron transport (rETR) of Porphyridium cruentum cultures maintained under two irradiances (50 and 200 μmol photons m−2 s−1) and two nitrogen sources (ammonium: NH4 and nitrate: NO3). Values with lowercase letters indicate significant differences between irradiances. Capital letters indicate significant differences between nitrogen sources by two-way ANOVA and Tukey a posteriori test, α = 0.05: a > b and A > B. Mean values ± SD, n = 3
Discussion
In this work, changes in the culture conditions affected the cell concentration, growth rate, and size of P. cruentum cells. The cell concentrations of P. cruentum in our treatments were higher compared with other reports on this strain (You and Barnett 2004; Razaghi et al. 2014). In our study, duration of the exponential growth phase in the P. cruentum cultures was shorter than those reported by other groups (12 days) (Adda et al. 1986; Nuutila et al. 1997; Razaghi et al. 2014), implying a reduction in the time to reach maximum biomass production. All growth rates in our work were higher than other values reported for P. cruentum with varying availability of nitrogen—from 0.25 to 0.37 divisions day−1 (Gigova and Ivanova 2015). Nevertheless, during heterotrophic growth and under various light:dark cycles, Gigova and Ivanova (2015) recorded a higher growth rate of P. cruentum (from 1.0 to 1.8 divisions day−1) than we did.
The P. cruentum cells in our study were larger with ammonium at the two irradiance levels, and these values were higher when compared with the same strain maintained in cultures at various N:P ratios using sodium nitrate as nitrogen source. The cell size correlated positively (from 2 to 5 μm) with the increase of N:P ratio (from 1.6 to 50) (Razaghi et al. 2014). The N:P ratio in our work was similar to N:P ratio used by Razaghi et al. (2014) in cultures of P. cruentum maintained at irradiance of 98 μmol photons m−2 s−1 with an 18:6 h light:dark cycle. Based on our results, a lower irradiance did not produce smaller cells, but nitrate decreased the size of P. cruentum cells, which nevertheless exceeded the values reported by other groups (Thepenier and Gudin 1985; Adda et al. 1986; Razaghi et al. 2014).
The total dry weight (TDW), organic dry weight (ODW), and ash content (ASH) of P. cruentum were modified by nitrogen sources. TDW values were 83 to 108% higher than those during maximum cell production by the same strain at an N:P ratio of 50 (1.2 g L−1) with nitrate at 98 μmol photons m−2 s−1 (Razaghi et al. 2014). The ODW of P. cruentum at 50 μmol photons m−2 s−1 with ammonium was 0.82 g L−1, similar to that by the same strain with “f” medium (0.95 g L−1) and lacking nitrogen (0.84 g L−1) when cultured at 28 °C and 132 μmol photons m−2 s−1 (Gigova and Ivanova 2015).
The lipid content of P. cruentum with ammonium was similar to other studies (2 to 19%) of the same strain on day 25 of culture with a 12:12 light:dark cycle at 30 °C, 10–25 μmol photons m−2 s−1, and 125 rpm (Oh et al. 2009). This rise was faster in our study (day 7), which decreased the time to reach the cellular components of interest, such as lipids, lowering the costs of maintenance and light supply. The lipid content of P. cruentum was 6.7% when cultured with Hemerick’s medium at 20 °C (Bermejo-Román et al. 2002). In this study, the lipid content of P. cruentum cultures increased at a lower irradiance and ammonium supply—other studies found that the accumulation of lipids can be induced in several microalgae strains by nitrogen-limiting conditions in the medium (Sánchez-Saavedra and Voltolina 2005; Muller-Feuga et al. 2007). Inducing lipid production through nitrogen limitation slows or halts biomass production; thus, our findings raise the possibility of increasing lipid content without affecting biomass production. We found that a combination of factors, such as low irradiance level (50 μmol photons m−2 s−1) and ammonium, could stimulate lipid production in a short time (7 days). Because lipid content was measured in the exponential growth phase, possibly the lipid content in P. cruentum might be enhanced if the cultures are harvested in the stationary growth phase.
Protein synthesis in P. cruentum was stimulated at the high-irradiance levels (200 μmol photons m−2 s−1). Our values of protein obtained were lower than others under different culture conditions (28 to 39%), measured as percentage of dry weight (Becker 2007). Generally, the hydro-soluble protein content increases during exponential growth; during stationary growth, nitrogen becomes limited, decreasing protein synthesis and thus reducing the amount of soluble protein that is available for metabolic processes (Matos-Moura et al. 2007).
The samples of P. cruentum cultures were collected in the exponential growth phase, and the lower protein content between culture conditions could be attributed to the protein extraction method as was mentioned by Safi et al. (2014), whom isolated hydro-soluble proteins with 2 N NaOH at pH 12. We considered that ultrasonication could have increased the cell disruption and thus the protein extraction. The protein content in future studies should also be quantified based on total nitrogen per the Kjelndahl method (APHA 1971). Protein extraction by manual grinding affects the amount of proteins that is quantified and has disparate yields (Safi et al. 2014). In other work with P. cruentum, the protein content ranges from 24.8 to 90%, and the content depends on the extraction method. The efficiency of protein extraction improves from manual grinding, followed by ultrasonication, chemical grinding, and high-pressure homogenization (Safi et al. 2014).
The differences in growth rate and biochemical composition of P. cruentum using various chemical forms of nitrogen are attributed to the finding that microalgae prefer ammonia over nitrate to conserve the energy of the eight electrons that are needed to reduce nitrate (South and Whittick 1987; Miller and Castenholtz 2001). Also, concentrations of ammonia over 0.5 to 1.0 μM can inhibit nitrate absorption (Darley 1987). The growth rate and proximal content of P. cruentum were similar to those of several microalgae strains that are used widely in aquaculture (Brown 2002; Muller-Feuga et al. 2007). The high carbohydrate and lipid content of P. cruentum allow it to be used in aquaculture to feed several species of marine organisms under acclimatization to increase gonad growth, such as postlarval spiny lobster, mussels, and oysters, because these stages require an abundance of carbohydrates and lipids in food (Flaak and Epifanio 1978; Widdows et al. 1979; Williams 2007).
The pigment content of P. cruentum was influenced by the culture conditions and demonstrates that the nitrogen was used for protein synthesis and consequently increased pigment-protein complexes in P. cruentum. The rise in phycopigments in P. cruentum correlates with the increase in carbohydrate content at low irradiance with nitrate. Phycocyanin (0.89 pg cell−1) and allophycocyanin (0.61 pg cell−1) levels were higher at the low irradiance (50 μmol photons m−2 s−1) with ammonium. Conversely, phycoerythrin production rates were greater when the cells were maintained at high irradiance (200 μmol photons m−2 s−1) with ammonium (6.35 mg L−1 day−1 = 0.98 pg cell−1 × 6.48 106 cells mL−1 × 1000 mL) and nitrate (2.55 mg L−1 day−1 = 1.03 pg cell−1 × 8.48 106 cells mL−1 × 1000 mL). Kathiresan et al. (2007) optimized the growth and phycoerythrin production by P. purpureum by response surface analysis, reporting phycoerythrin production of 29.62, 6.11, 1.59, and 0.076 g L−1 day−1, depending on the concentration of chloride, nitrate, sulfate, and phosphate. Phycoerythrin production in P. cruentum is 25 mg L−1 day−1 at pH 7.6 (Lee and Tan 1988), peaking at 49 mg L−1 day−1 for small-scale semicontinuous indoor cultures (Fábregas et al. 1998). Our phycoerythrin production rates were lower compared to these other groups, perhaps due to differences in culture conditions, such as steady-state continuous chemostat culture, medium (nutrient composition and concentration), temperature, irradiance, and agitation.
The rETR curves and photosynthetic activity were higher with nitrate at both irradiance levels, whereas the rETRm and Ik were unaffected by irradiance levels. In our study, there were no differences in photosynthetic efficiency (α) among the irradiance levels. The Fv/Fm value may be used to study microalgae cultures, and the values of this parameter varies from 0.6 to 0.7 for unlimited cultures (Kromkamp and Peene 1999). In our study values of Fv/Fm were around 0.30 and no significant differences were obtained. The values of Fv/Fm obtained in our work were lower compared to other studies performed in Chaetoceros debilis (0.58), Dicrateria inornata (0.45), or Platymonas subcordiformis (0.5) (Qi et al. 2013). In red algae, the presence of phycobilisomes in the light-harvesting system may lead to obtain lower values of Fv/Fm than those obtained in green or brown algae (Figueroa et al. 2013). However, the chemical form of nitrogen—ammonium and nitrate—altered photosynthetic activity. The size of P. cruentum cells was lower under the same culture conditions that increased the photosynthetic efficiency (nitrate at both irradiances levels). In these treatments, the rETR value was higher and the chlorophyll content in lower irradiance was higher than in treatments with high irradiance. It is possible that higher cell density, in treatments with nitrate and low irradiance, elicited a self-shading effect due to the increment in cell concentration—i.e., the package effect—resulting in a direct effect on light absorption due to a combination of photosynthetic pigments and cell size (Geider and Osborne 1992; Kirk 1994). Conversely, in treatment with high irradiance and nitrate as nitrogen source, the rETR value and photosynthetic efficiency was higher, but the chlorophyll content was lower. This response can be due to an adaptation of P. cruentum cells to high availability of light. Therefore, there is no need to produce high concentrations of chlorophyll. The photosynthetic apparatus can adapt to irradiance and photoperiod (including light-shade adaptation) (Falkowski and Owens 1980; Raven and Geider 2003). Villafañe et al. (2005) found that P. cruentum is affected by solar ultraviolet radiation (280–400 nm and 315–400 nm) in the short term but can acclimate quickly to high irradiances, adjusting their photosynthesis but not growth rate.
The irradiance level and chemical nitrogen form increased the content of several chemical products in P. cruentum, such as chlorophyll a, carotenoids, phycocyanin, phycoerythrin, and allophycocyanin. Thus, our experimental culture conditions can be used to improve biomass production in P. cruentum and synthesize pigments, carbohydrates, and lipids for use in cosmetology, pharmacology, and feed production (Arad and Yaron 1992; Spolaore et al. 2006; Ruiz-Ruiz et al. 2013; Razaghi et al. 2014).
In conclusion, irradiance level and chemical nitrogen form modify the growth and proximate composition of P. cruentum cultures. For aquaculture applications, P. cruentum can be cultured with ammonium and irradiance of 50 μmol photons m−2 s−1 due to this growth condition increases the nutritional value—lipid and carotenoid content—of P. cruentum when used as feed. On the other hand, if the interest is to obtain phycoerythrin—the most coveted pigment in Rhodophyta—for applications in biotechnology, food, and cosmetology industries, it is recommended to use nitrate and irradiance of 200 μmol photons m−2 s−1 because this growth condition elevates the content of this pigment in P. cruentum.
References
Adda M, Merchu JC, Arad SM (1986) Effect of nitrate on growth and production of cell-wall polysaccharide by the unicellular red alga Porphyridium. Biomass 10:131–140
Ahern TJ, Katoh S, Sada E (1983) Arachidonic acid production by the red algae Porphyridium cruentum. Biotechnol Bioeng 25:1057–1070
Ahlström LH, Eskilsson CS, Björklund E (2005) Determination of banned azo dyes in consumer goods. Trends Anal Chem 24:49–56
Antia NJ, Desai ID, Romilly MJ (1970) The tocopherol, vitamin K and related isoprenoid quinone composition of an unicellular red algae (Porphyridium cruentum). J Phycol 6:305–312
APHA (American Public Health Association (APHA) (1971) Standard methods for the examination of water and wastewater, 13th edn. APHA, Washington, D.C, p 110
Arad A, Yaron A (1992) Natural pigments from red microalgae for use in foods and cosmetics. Trends Food Sci Technol 3:92–97
Becker EW (2007) Micro-algae as a source of protein. Biotechnol Adv 25:207–210
Bennet A, Bogorad L (1973) Complimentary chromatic adaptation in a filamentous blue-green alga. J Cell Biol 58:419–435
Bermejo-Román R, Alvárez-Pez JM, Acién-Fernández FG, Molina-Grima E (2002) Recovery of pure B-phycoerythrin from the microalgae Porphyridium cruentum. J Biotechnol 93(1):73–85
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Brown MR (2002) Nutritional value of microalgae for aquaculture. In: Cruz-Suárez LE, Ricque-Marie D, Tapia-Salazar M, Gaxiola-Cortez MG, Simoes N (eds) Avances en Nutrición Acuícola VI Simposium Internacional de Nutrición Acuícola. 3–6 de septiembre, Cancún, Quintana Roo, pp 281–292
Chiaverini J (1972) Techniques d’extraction et d’analyse des lipids. Université de Paris et Marie Curie, Paris. Station Zoologique Villefranche-Sur-Mer. Notes de Travail 12, p 12
Cuellar-Bermúdez SP, Aguilar-Hernández I, Cárdenas-Chávez DL, Ornelas-Soto N, Romero-Ogawa MA, Parra-Saldívar R (2014) Extraction and purification of high-value metabolites from microalgae: essential lipids, astaxanthin and phycobiliproteins. Microbioal Biotechnol 8:190–209
Curtin ME (1985) Chemicals from the sea. Biotechnology 3:34–37
Darley MW (1987) Biología de las Algas: Enfoque Fisiológico. In: México, DF (ed) Limusa SA de CV. p 236
Dubinsky Z, Stambler N (2009) Photoacclimation processes in phytoplankton mechanism, consequences, and applications. Aquat Microb Ecol 56:163–176
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356
Fábregas J, García D, Morales E, Domínguez A, Otero A (1998) Renewal rate of semicontinuous cultures of the microalga Porphyridium cruentum modifies phycoerythrin, exopolysaccharides and fatty acid productivity. J Ferment Bioeng 86:477–481
Falkowski PG, Owens TG (1980) Light-shade adaptation: two strategies in marine phytoplankton. Plant Physiol 66:592–595
Falkowski PG, Raven JA (2007) Aquatic photosynthesis. Princeton University Press, Princenton, p 484
Figueroa FL, Conde-Álvarez R, Gómez I (2003) Relation between electron transport rates determined by pulse amplitude modulated chlorophyll fluorescence and oxygen evolution on macroalgae under different light conditions. Photosynth Res 75:259–275
Figueroa FL, Jerez CG, Korbee N (2013) Use of in vivo chlorophyll fluorescence to estimate photosynthetic activity and biomass productivity in microalgae grown in different culture systems. Lat Am J Aquat Res 41(5):801–819
Flaak AR, Epifanio CE (1978) Dietary protein levels and growth of the oyster Crassostrea virginica. Mar Biol 45:157–163
Fogg GE, Thake BJ (1987) Algal cultures and phytoplankton ecology. University of Wisconsin Press, London, p 269
Gantt E (1969) Properties and ultrastructure of phycoerythrin from Porphyridium cruentum. Plant Physiol 44:1629–1638
Gantt E (1981) Phycobilisomes. Annu Rev Plant Physiol 32:327–347
Geider RJ, Osborne BA (1992) Algal photosynthesis: the measurement of algal gas exchange. Chapman and Hall Press, New York, p 256
Gigova LG, Ivanova NJ (2015) Microalgae respond differently to nitrogen availability during culturing. J Biosci 40:365–374
Grewe CB, Pulz O (2012) The biotechnology of cyanobacteria. In: Whitton BA (ed) Ecology of Cyanobacteria II. Springer, Netherlands, pp 707–739
Guedes AC, Amaro HM, Malcata FX (2011) Microalgae as sources of carotenoids. Mar Drugs 9:625–644
Guillard RLL, Ryther JH (1962) Studies on marine planktonic diatoms I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can J Microbiol 8:229–239
Hu Q (2005) Environmental effects on cell composition. In: Richmond A (ed) Handbook of microalgal culture. Biotechnology and Applied Phycology. Blackwell Publishing, Oxford, pp 83–93
Kathiresan S, Sarada R, Bhattacharya S, Ravishankar GA (2007) Culture media optimization for growth and phycoerythrin production from Porphyridium purpureum. Biotechnol Bioeng 96:456–463
Kavitha MD, Seema Shree MH, Vidyashankar S, Sarada R (2016) Acute and subchronic safety assessment of Porphyridium purpureum biomass in the rat model. J Appl Phycol 28:1071–1083
Kirk JTO (1994) Light and photosynthesis in aquatic ecosystems. Cambridge University Press, Cambridge, p 401
Kromkamp J, Peene J (1999) Estimation of phytoplankton photosynthesis and nutrient limitation in the Eastern Scheldt estuary using variable fluorescence. Aquat Ecol 33:101–104
Lee YK, Tan HM (1988) Effect of temperature, light intensity and dilution rate on the cellular composition of red alga in light-limited chemostat cultures. MIRCEN J Appl Microbiol Biotechnol 4:231–237
Lourenço SO, Barbarino E, Mancini-Filho J, Schinke KP, Aidar E (2002) Effects of different nitrogen sources on the growth and biochemical profile of 10 marine microalgae in batch culture: an evaluation for aquaculture. Phycologia 41:158–168
Lowry OH, Rosebrough HJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin-phenol reagent. J Biol Chem 193:265–275
Matos-Moura A, Bezerra-Neto E, Koening ML, Leca EE (2007) Chemical composition of three microalgae species for possible use in mariculture. Braz Arch Biol Technol 50(3):461–467
Mazzuca-Sobczuc T, Ibañez-González MJ, Molina-Grima E, Urrutia-Martínez T, Yusuf Ch (2015) Concentración de cultivos de microalgas por un proceso de eliminación osmótica del medio utilizandodisoluciones de glicerol. Spanish Patent 2 545 829
Miller SR, Castenholtz RW (2001) Ecological physiology of Synechococcus sp. strain SH-94-5, a naturally occurring cyanobacterium deficient in nitrate assimilation. Appl Environ Microbiol 67:3002–3009
Misra HP, Fridovich I (1977) Purification and properties of superoxide dismutase from a red alga, Porphyridium cruentum. J Biol Chem 252:6421–6423
Muller-Feuga A, Moaly J, Kaas R (2007) The microalgae of aquaculture. In: Støttrup JG, McEvoy LA (eds) Live feeds in marine aquaculture. Blackwell Science, Oxford, pp 207–252
Nuutila AM, Aura AM, Kiesvaara M, Kauppinen V (1997) The effect of salinity, nitrate concentration, pH and temperature on eicosapentaenoic acid (EPA) production by the red unicellular alga Porphyridium purpureum. J Biotechnol 55:55–63
Oh SH, Han JG, Kim Y, Ha JH, Kim SS, Jeong MH, Jeong HS, Kim NY, Cho JS, Yoon WB, Lee SY, Kang DH, Lee SY (2009) Lipid production in Porphyridium cruentum grown under different culture conditions. J Biosci Bioeng 108:429–434
Pande SV, Khan RP, Venkitasubramanian TA (1963) Microdetermination of lipids and serum total acids. Anal Biochem 6:415–423
Pangestuti R, Kim S-K (2011) Biological activities and health benefit effects of natural pigments derived from marine algae. J Funct Foods 3:255–266
Paniagua-Michel J, Dujardin E, Sironval C (1993) Le tecuitlatl, concentré de spirulines source de protéines comestibles chez les Aztèques. Cah Agric 2(4):283–287
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis. Pergamon Press, Oxford, p 173
Pulz O, Gross W (2004) Valuable products from biotechnology of microalgae. Microbiol Biotechnol 65:635–648
Qi H, Wang J, Wang Z (2013) A comparative study of the sensitivity of Fv/Fm to phosphorus limitation on four marine microalgae. J Ocean Univ China 12:77–84
Raven JA, Geider RJ (2003) Adaptation, acclimation and regulation in algal photosynthesis. In: Larkum AWD, Douglas SE, Raven JA (eds) Advances in photosynthesis and respiration, Photosynthesis in algae, Kluwer, Dordrecht, pp 385–412
Raven JA, Giodano M (2016) Combined nitrogen. In: Borowitzka MA, Beardall J, Raven JA (eds) The physiology of microalgae. Springer, Dordrecht, pp 143–154
Razaghi A, Godhe A, Albers E (2014) Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Cent Eur J Biol 9:156–162
Rebolloso-Fuentes MM, Acién-Fernández GG, Sánchez-Péz JA, Guil-Guerrero JL (2000) Biomass nutrient profiles of the microalga Porphyridium cruentum. Food Chem 70:345–353
Romay C, González R, Ledón N, Remirez D, Rimbao V (2003) C-Phycocyanin: a biliprotein with antioxidant, anti-inflamatory and neuroprotective effects. Curr Protein Pept Sci 4:207–216
Ruiz-Ruiz F, Benavides J, Rito-Palomares M (2013) Scaling-up of a B-phycoerythrin production and purification bioprocess involving aqueous two phase systems: practical experiences. Process Biochem 48:738–745
Safi C, Ursu AV, Laroche C, Zebib B, Merah O, Pontalier P-Y, Vaca-García C (2014) Aqueous extraction of proteins from microalgae: effect of different cell disruption methods. Algal Res 3:61–65
Sánchez-Saavedra MP, Voltolina D (2005) The growth rate, biomass production and composition of Chaetoceros sp. grown with different light sources. Aquac Eng 35:161–165
Sokal RR, Rohlf FJ (1995) Biometry. The principles and practice of statistics in biological research, 3rd edn. WH Freeman & Co., New York, p 887
Sorokin C (1973) Dry weight, packed cell volume and optical density. In: Stein JR (ed) Handbook of phycological methods and growth measured. Cambridge University Press, New York, pp 321–343
South GR, Whittick A (1987) Introduction to phycology. Blackwell Scientific Publications, London, p 341
Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006) Commercial applications of microalgae. J Biosci Bioeng 101:87–96
Thepenier C, Gudin C (1985) Studies on optimal conditions for polysaccharide production by Porphyridium cruentum. World J Microbiol Biotech 1:257–268
Vadiveloo A, Moheimani NR, Cosgrove JJ, Parlevliet D, Bahri PA (2017) Effects of different light spectra on the growth, productivity and photosynthesis of two acclimated strains of Nannochloropsis sp. J Appl Phycol 29:1765–1774
Villafañe VE, Gao K, Helbling EW (2005) Short- and long-term effects of solar ultraviolet radiation on the red algae Porphyridium cruentum (S.F. Gray) Nägeli. Photochem Photobiol 4:376–382
Vonshak A (1988) Porphyridium. In: Borowitzka MA, Borowitzka LJ (eds) Micro-algal biotechnology. Cambridge University Press, Cambridge, pp 122–134
White S, Anandraj A, Bux F (2011) PAM fluorometry as a tool to assess microalgal nutrient stress and monitor cellular neutral lipids. Bioresour Technol 102:1675–1682
Whyte JNC (1987) Biochemical composition and energy content of six species of phytoplankton used in mariculture of bivalves. Aquaculture 60:231–241
Widdows J, Fieth P, Worrall CM (1979) Relationships between seston, available food and feeding activity in the common mussel Mytilus edulis. Mar Biol 50:195–207
Williams KC (2007) Nutritional requirements and feeds development for post-larval spiny lobster: a review. Aquaculture 263(1):1–14
Yen HW, Hu IC, Chen CY, Ho SH, Lee DJ, Chang JS (2013) Microalgae-based biorefinery–from biofuels to natural products. Bioresour Technol 135:166–174
You T, Barnett SM (2004) Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem Eng J 19:251–258
Acknowledgments
Nava-Ruiz V.M., Ruiz-Güereca D.A., Villagómez-Aranda A.L., and Siqueiros-Vargas F. acknowledge their Master in Science scholarship from CONACyT. We thank F.D. López-Figueroa for his help to photosynthetic parameters calculation. English language was edited by Blue Pencil Science and C.G. Paniagua-Chávez.
Funding
This work has been funded by Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), Fund for Scientific Research and Technological Development of CICESE (Project: 623801), and CICESE (Project: 623101).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Sánchez-Saavedra, M.d.P., Castro-Ochoa, F.Y., Nava-Ruiz, V.M. et al. Effects of nitrogen source and irradiance on Porphyridium cruentum . J Appl Phycol 30, 783–792 (2018). https://doi.org/10.1007/s10811-017-1284-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-017-1284-2