
Abstract
Laminaria pallida and Ecklonia maxima are large, commercially valuable kelps that co-dominate inshore waters of the west coast of Southern Africa in a geographically changing pattern. In the south, E. maxima dominates and forms a canopy in shallow waters (< about 5 m deep), with L. pallida forming a sub-canopy and extending down to 20 m or more. Northward along the Southern African coast and into Namibia, E. maxima is progressively replaced by L. pallida. Corresponding with this change in dominance, L. pallida shows certain morphological changes along the south-to-north geographical gradient. To explain these phenomena, we examined a range of morphological characters in both kelps (stipe length, stipe weight, stipe outer diameter, stipe inner diameter, length of hollow section in the stipe, and frond weight), and various environmental factors (seawater temperature, seawater turbidity, cloud/fog data, daylength, and wave and wind data). Our results, based on measurements at seven sites along 1600 km of coast between Cape Town and Swakopmund (Namibia), quantified and confirmed the change in dominance and the northward increase in stipe hollowness in L. pallida. The morphology of E. maxima did not change with latitude. Water turbidity, wind speed, and wave height differed significantly along the coast. However, only turbidity showed a steady trend, increasing northward in terms of all indicators (chlorophyll a, particulate inorganic carbon, particulate organic carbon) while wind speed and wave height showed a generally decreasing trend. Furthermore, the hollowness of L. pallida was also not related to the flexibility of the stipes. Our results suggest that L. pallida sporophytes may progressively outcompete E. maxima northward, perhaps because they are more low-light tolerant, and we suggest that by developing a hollow stipe, the sporophytes may grow faster in length, potentially increasing their competitive advantage in the shallow water where they must compete with sporophytes of E. maxima.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
There are four species of Southern African kelps (Laminariales): Ecklonia maxima (Osbeck) Papenfuss, Ecklonia radiata (C.Agardh) J.Agardh, Laminaria pallida Greville and Macrocystis pyrifera (Linnaeus) C.Agardh (previously known in South Africa as Macrocystis angustifolia Bory de Saint-Vincent). Ecklonia maxima and L. pallida have by far the highest biomass in the inshore, rocky environment, and are the only species exploited economically in South Africa. These species are mainly used as feed for abalone, but the juice is also extracted to be used as a growth stimulant in the agricultural industry. The region, where these economical species occur, falls within the Benguela Current Large Marine Ecosystem which is largely characterised by intense inshore upwelling, bringing cold, nutrient-rich water (Andrews and Hutchings 1980; Shannon 1985; Shannon and Pillar 1986) which sustains these large kelps. In Southern Africa, subtidal kelp forests only occur from near Cape Agulhas in the south to Rocky Point in Northern Namibia (Stegenga et al. 1997), comprising either E. maxima or L. pallida or a mixture of both (Molloy and Bolton 1990).
Ecklonia maxima is a floating, canopy kelp and one of the largest members of the Laminariales, generally not exceeding lengths of 10 m except on rare occasions where stipes of up to 17 m have been recorded (Anderson et al. 1997). Extensive beds of E. maxima are found from just west of Cape Agulhas (although small beds appeared recently 70 km east of Agulhas, at De Hoop Nature Reserve; Bolton et al. 2012), to Hottentotspunt, 60 km north of Lüderitz in Namibia (Molloy and Bolton 1990; Stegenga et al. 1997) (see Fig. 1). This species forms extensive forests down to a depth of about 10 m (Simons and Jarman 1981; Bolton and Anderson 1987; Anderson et al. 2007) and can comprise up to 70% of the total plant biomass in the inshore zone (Velimirov and Griffiths 1979; Field et al. 1980; Rothman 2006).

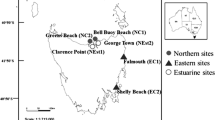
Coastline of South Africa and Namibia showing sites mentioned in the text. Inset shows the Cape Peninsula enlarged
Laminaria pallida, the other dominant kelp in South Africa and the dominant species in Namibia, is found from Danger Point (about 100 km east of Cape Town) in South Africa to Rocky Point, south of the Kunene River in northern Namibia (Molloy and Bolton 1990; Stegenga et al. 1997). Jarman and Carter (1981) estimated the standing stock of L. pallida between Cape Point and Cape Columbine to be almost 218,000 t fresh weight (no data are available for the rest of the distribution area). Along the southern west coast, L. pallida forms extensive subtidal, understorey forests below and beyond the canopy of E. maxima and can extend to depths of 20 m or more (Field et al. 1980; Molloy and Bolton 1996; Anderson et al. 2007). Northward, L. pallida is reported to become more dominant inshore, gradually replacing E. maxima (Velimirov et al. 1977; Stegenga et al. 1997), but still with extensive deeper subtidal beds.
In its southern area of distribution, L. pallida was reported to have a solid stipe that is widest near the holdfast, while in the north it was reported to have a longer, hollow stipe that is widest in the mid-region and narrows towards the distal and proximal ends (Molloy and Bolton 1996). Previously, these two forms were considered to be separate species, with the name L. pallida applied to the former (southern) entity and the name Laminaria schinzii Foslie applied to the latter. However, Stegenga et al. (1997) considered the latter to be a form of L. pallida, an opinion that has now been molecularly confirmed (Rothman et al. 2015). In the south, only occasional sporophytes are hollow-stiped (MD Rothman, personal observation), but this form is reported to be more common north of St Helena Bay on the South African west coast (Stegenga et al. 1997). Molloy and Bolton (1996) showed that stipe weight, stipe length, and stipe thickness of L. pallida are all affected by depth (between 0.25 and 2.25 m), but the greatest interaction was observed between water motion and blade thickness, with thin blades being observed in wave-exposed areas to reduce drag resistance.
Ecklonia arborea (Areschoug) M.D.Rothman, Mattio & J.J.Bolton (as Eisenia arborea Areschoug) is the only other known example of a kelp sporophyte that becomes hollower along a geographical gradient, along the NE Pacific coast (Matson and Edwards 2006). A number of studies suggest that the flexibility of kelp stipes might be related to the wave exposure, especially in shallow water (Friedland and Denny 1995; Denny et al. 1997; Gaylord and Denny 1997). These studies imply that kelp sporophytes’ strategy for survival in high water motion environments is flexibility rather than strength and resistance.
Many studies have shown how environmental variables such as wave exposure, water temperature, and even substratum stability can dramatically change the morphology of a kelp sporophyte (Norton et al. 1982; Tsutsui et al. 1996; Serisawa 2002, 2003; Fowler-Walker et al. 2006; Klochkova et al. 2010; Hurd and Pildith 2011; Marins et al. 2012). While the above studies demonstrate the importance of the environment in influencing kelp morphology, relatively few have examined changes over large gradients of latitude or longitude, and most have examined the effects of wave exposure at geographically close sites. The exceptions are the studies of Wernberg et al. (2003) with E. radiata and Matson and Edwards (2006) with E. arborea. However, although E. arborea was shown to develop a hollow stipe over a latitudinal gradient, no explanation was advanced.
Although it has been descriptively reported that L. pallida gradually replaces E. maxima in inshore kelp beds along the west coast of Southern Africa and develops a hollow stipe (Molloy and Bolton 1990, 1996; Stegenga et al. 1997), it has never been confirmed quantitatively, nor has this transition been explained. The present study set out to measure this ecological transition by quantifying the morphological patterns (especially stipe morphology) in this species and its co-dominant, E. maxima, in shallow water along the coast. This study focuses on mature plants (with a stipe length of more than 50 cm), shallow-water sporophytes, in < 5 m depth, where these two species’ vertical distribution ranges overlap, and they may be expected to compete for space and other resources.
We therefore set out to test the hypothesis that, along the approximately 1600 km of the Southern African west coast, the changes in dominance of the two kelps and the change in the morphology of L. pallida have a clear pattern which may be related to a gradient in one or more environmental factors.
Materials and methods
Study area
This study was conducted along the Southern African west coast from False Bay to as far north as was logistically possible in the L. pallida distribution (Swakopmund, Namibia) (see Fig. 1). We measured a number of morphological characters in L. pallida and Ecklonia maxima, and as many environmental factors as possible, at a number of sites along the Southern African west coast.
Measurement of biological factors
Morphometrics and biomass
Between 2009 and 2010, E. maxima and L. pallida sporophytes were collected from eight sheltered to semi-exposed sites—five sites in South Africa (Bordjiesrif (34° 18′ 50.58″ S, 18° 27′ 49.43″ E), Kommetjie (34° 8′ 30.24″ S, 18° 19′ 8.10″ E), Jacobs Bay (32° 58′ 33.03″ S, 17° 52′ 50.69″ E), Hondeklip Bay (30° 18′ 26.64″ S, 17° 16′ 7.52″ E), Kleinzee (29° 42′ 32.10″ S, 17° 3′ 15.80″ E))—and in 2012 from one site in Namibia, Swakopmund (22° 42′ 5.03″ S, 14° 31′ 12.44″ E), where only L. pallida is found (Fig. 1).
At all sites except Swakopmund, shallow-water kelp sporophytes were collected, using SCUBA, along a transect perpendicular to the shore, starting from the outer edge of the surface-reaching kelp and working towards the shore. Three depth classes were selected: deep (3.1–5 m), intermediate (1.1–3 m), and shallow (0–1 m). All sampling was done at LWS (low water spring). This depth range ensured that the densest part of the kelp beds was sampled, and also allowed shore-based sampling in remote locations where a boat was unavailable.
Quadrats were formed by placing two, 1-m-long metal rods at right angles to demarcate a 1-m2 area, and all the plants with stipes longer than 50 cm were harvested at the base of the stipe. A minimum of ten quadrats (between 3 and 5 m apart depending on the width of the bed) were sampled at each site (see later), so three to four quadrats were placed in each depth class. Because of the low density of L. pallida in shallow water at Kommetjie, only one sporophyte was recorded in the quadrats. At Swakopmund, where SCUBA equipment was unavailable, 30 shallow-water sporophytes were haphazardly collected along a transect between 0.5- and 2.5-m depths at LWS.
The following measurements were taken on each sporophyte.
-
Stipe length: measured from the holdfast to the junction between the primary blade and the stipe, with a tape measure.
-
Stipe hollowness: the stipe was cut from both ends until the hollow section was reached, and air could easily be blown through that section of stipe. The length of the hollow section was measured and expressed as a percentage of the total stipe length.
-
Stipe weight: the fresh weight of the entire stipe was measured with a spring balance, to the nearest 25 g.
-
Frond weight: the entire primary and secondary frond mass, from junction of stipe and blade, was weighed with a spring balance, to the nearest 25 g.
All the above variables were analysed using the multivariate analysis package Primer 6 (Clarke and Gorley 2006). Data were normalised before principal component analysis (PCA) and non-metric multi-dimensional scaling (MDS) were used to illustrate similarity between samples (MDS showed extremely similar patterns to PCA and thus is not shown here). In addition, regressions, box plots, and ANOVA were used to analyse in more detail the major contributing factors as shown by the vectors from the PCA.
Kelp flexibility
Thirty L. pallida sporophytes (at each depth) were collected haphazardly from Kommetjie (where sporophytes almost all have solid stipes) and Doring Bay (where sporophytes almost all have hollow stipes). Sporophytes were collected from shallow (< 4 m) and deep water (between 10 and 15 m).
Each kelp stipe was separated from the holdfast and primary blade and its basal 20 cm held in a rigid and immovable clamp (see Online Resource 1; Fig. S1). The stipes were held parallel to the ground and bent horizontally to a 45° and then a 90° angle. The angles were measured relative to a bar that pivoted at the clamp, and were effectively the angles between clamp and the point of attachment of the pulling force, which was applied at two distances from the clamp: 50 and 70 cm. The tension required to reach each angle was measured with a spring balance to the nearest 25 g. Gravity and frictional forces were mitigated by resting the stipe on the edge of a ruler that acted as a track over which the stipe could be pulled. Each stipe was then sectioned to confirm whether it was hollow or solid. A one-way ANOVA was performed to test significance (sites were not separated).
Measurement of environmental factors
Turbidity
Monthly composite Aqua MODIS 4-km-resolution ocean colour images were downloaded from the NASA Ocean Colour website (http://modis.gsfc.nasa.gov), for a 5-year period (2008–2013; i.e. 180 data points for each site, 60 per variable), and manually analysed. Colours from the images were matched by eye to the key to extract turbidity values at the eight sites where morphometric data were collected (a similar method to that used by Smale and Wernberg (2009) to generate a temperature dataset in Western Australia). Parameters used as proxies for turbidity were chlorophyll a concentrations (Chl a), particulate organic carbon (POC), and particulate inorganic carbon (PIC). Images from the MODIS sensors on the Terra and Aqua satellites have previously been shown to reliably measure water turbidity (Chen et al. 2007; Doxaran et al. 2009; Petus et al. 2010). A one-way ANOVA and Tukey post hoc tests were used to determine significant differences in turbidity between sites.
Cloud/fog data
Cloud data were visually extracted from MODIS Aqua and Terra 1-km-resolution images that were downloaded from the NASA website for 2011. Presence/absence of clouds was recorded for the same sites where morphometric data were collected, for every day during 2011. The satellite passed overhead twice daily, once in the morning and once in the afternoon. Data were analysed using a one-way ANOVA and a Tukey post hoc test to establish significance.
Temperature data
Seawater temperature data were obtained from the South African Weather Service for the following sites: Kommetjie, Paternoster (20 km north of Jacobs Bay), Lamberts Bay (30 km south of Doring Bay), Doring Bay, Hondeklip Bay, and Port Nolloth. The temperatures were measured daily by thermometer. For each site, temperatures were for periods of 15 to 30 years (1981–2011). Bordjiesrif temperature data were obtained from the Starmon Mini underwater temperature recorders (Star Oddi, Iceland) of the Department of Agriculture, Forestry and Fisheries, placed at 8 m depth. The Swakopmund temperature data are from NASA Sea Surface Temperature data, corrected to reflect true coastal seawater temperatures (see Smit et al. 2013). Results were plotted to show maximum and minimum monthly means, and annual means of the entire data set at each site.
Wave and wind data
Windguru (by permission Hornik (2014), http://www.windguru.cz/int/) data were downloaded for the following variables: wave speed, wave period, wave direction, wind speed, and wind direction. Data were measured 3-hourly from February 2013 till July 2014. All the above variables were analysed using the multivariate analysis package Primer 6. Data were normalised before principal component analysis (PCA) and multi-dimensional scaling (MDS) were used to investigate similarity between sites. In addition, regressions, box plots, ANOVA, and post hoc comparisons were used to analyse the major contributing factors that were shown by vectors for the PCA. Wind and wave directions were analysed and plotted using the freeware WRPLOT (Lakes Environmental Software: http://www.weblakes.com) to generate the plots to compare the different sites. The roses indicate direction as well as wave height (in the case of swell) and wind speed (in the case of wind). Regression analyses were performed (1) to indicate the relationship between the average wind speed and wave height versus the distance of the site from Cape Point; (2) to indicate the average maximum wave height versus the distance of the site from Cape Point.
Results
Kelp biological data
Kelp dominance
There was a gradual changeover in the dominance of the canopy kelps, with L. pallida replacing E. maxima from south to north (Fig. 2a). In inshore shallow water, all of the sporophytes at Bordjiesrif were E. maxima while at Swakopmund in Namibia (about 1600 km from Bordjiesrif), all were L. pallida. At Hondeklip Bay (about 600 km from Bordjiesrif), there was a 42/58 percentage split between the two species.

a Relative occurrence of E. maxima (grey) and L. pallida (black) along the Southern African West Coast. Data was collected by benthic quadrats. b Biomass of E. maxima (grey) compared to L. pallida (black) along the South African west coast. Bars indicate 95% confidence limits
Kelp biomass
Ecklonia maxima biomass per square metre (Fig. 2b) increased from Bordjiesrif in False Bay to Kommetjie (26 km further) on the Atlantic side of the Cape Peninsula. The subsequent declining trend in biomass, northward to Kleinzee (700 km from Bordjiesrif), was not significant, but the lowest average biomass was observed at Kleinzee. In contrast, the highest average biomass of L. pallida was found at Kleinzee and only a single plant was sampled at Kommetjie and none at Bordjiesrif. Hondeklip Bay represented a transition zone for dominance between the two species, with approximately equal biomass of each species. Mean plant weight of E. maxima was lowest at Bordjiesrif and highest at Kommetjie, east of which this parameter showed little variation (Online Resource 1; Fig. S2). Laminaria pallida sporophytes, however, had the highest average plant weight at Kleinzee while sporophytes from Swakopmund had similar average plant weights to sporophytes from Jacobs Bay and Hondeklip Bay (Online Resource 1; Fig. S2) (there is insufficient data for Kommetjie—only one sporophyte was found in the quadrats).
Hollowness and stipe length
Ecklonia maxima stipes were always hollow with no significant difference between sites (with relative hollowness between 86 and 90%) (Fig. 3a; r 2 = 0.6487, ANOVA, F(4, 361) = 1.3435, p = 0.2533). However, the regression (with a polynomial fit) shows that there was a clear gradient of increasing stipe hollowness in L. pallida from south to north, with relative hollowness ranging between solid at Kommetjie and 87% hollow at Swakopmund (Fig. 3a; r 2 = 0.5291, ANOVA, F(4, 174) = 54.6345, p < 0.001). At Kommetjie, only a single sporophyte was collected in the transects, but a separate study at the same site, that sought out only large, older L. pallida sporophytes, showed that 10% of them had some degree of hollowness in the stipe (n = 40). The results here, however, only use the transect data, in order to ensure methodological consistency of comparisons. E. maxima showed no correlation between stipe length and hollowness; all plants had the same degree of hollowness (Fig. 3b) although the stipe length was variable (n = 366; p = 0.2533; r 2 = 0.6487). By contrast, in L. pallida there was a positive linear correlation between stipe length and hollowness (n = 153; p < 0.05; r 2 = 0.5291) (Fig. 3b).

a Relative hollowness of E. maxima (grey) and L. pallida (black) with increased distance from Bordjiesrif. b Relative hollowness of E. maxima (grey) and L. pallida (black) with increase in stipe length
Laminaria stipe flexibility
Tukey post hoc analysis showed that hollowness did not affect the flexibility of L. pallida stipes (ANOVA, F(12,185.49) = 4.0554, p > 0.001) (Online Resource 1; Fig. S3 and Table S1), irrespective of whether stipes were bent to 45° or 90°.
Multivariate analysis of morphological variables
The PCA of the E. maxima morphological data (Online Resource 1; Fig. S4), indicated that none of the sites (except some of the individuals at Kommetjie) separated out, indicating no effect of site on morphology (vectors from the PCA analysis indicated no significance). However, in the L. pallida morphological data, the PCA (Online Resource 1; Fig. S5) showed some separation of three groups of sites. The first group comprised the northernmost sites (Kleinzee and Swakopmund) with the percent hollowness being the major forcing vector for this separation. The second group comprised mostly the intermediate Hondeklip Bay site, with some overlap between Jacobs Bay, Hondeklip Bay, Kleinzee, Swakopmund, and the only sporophyte from Kommetjie. The third group was from Jacobs Bay with little overlap from Hondeklip Bay (there was no L. pallida sampled at Bordjiesrif).
Environmental variables
Turbidity
The values of all three variables that were used as proxies for turbidity increased from south to north (Fig. 4a–c). Moving northwards from Bordjiesrif, all three variables showed a progressive increase, but Jacobs Bay, Hondeklip Bay, and Kleinzee all had similar levels of Chl a, PIC, and POC. The highest levels for all three variables were observed for Swakopmund in Namibia. The regressions (with a polynomial fit) showed that there were very good direct positive relationships between Chl a (r 2 = 0.7778; ANOVA, F(7, 137) = 10.1799, p < 0.001), PIC (r 2 = 0.8931; ANOVA, F(7, 137) = 12.0156, p < 0.001), and POC (r 2 = 0.8854; ANOVA, F(7, 137) = 13.9381, p < 0.001).

Three variables indicating turbidity of seawater, at various distances from Bordjiesrif along the Southern African west coast. a Chlorophyll a concentration. b Particulate organic carbon. c Particulate inorganic carbon. Bars indicate 95% confidence intervals
Fog or cloud cover
The relative fog or cloud cover (Online Resource 1; Fig. S6; Table S2) showed no significant variation over the South African portion of the study area for the year 2010, with values ranging from 28.5 to 35.6% (ANOVA, F(1,6) = 1.241, p = 0.2818). However, the relative cloud/fog cover at Swakopmund (46%) was significantly higher than at the South African sites (p < 0.001).
Daylength
Summer daylength in Cape Town is 18 min longer than at Port Nolloth and 49 min longer than at Swakopmund. In winter, Swakopmund (the northern site) has days 29 min longer than Port Nolloth and 46 min longer than Cape Town.
Temperature data
After a sudden decrease from 15.5 to 13.3 °C, between Bordjiesrif and Kommetjie, the mean annual seawater temperature decreased slightly northwards to 12.3 °C at Port Nolloth, after which, over the next 880 km, it rose again to 14.7 °C at Swakopmund (Fig. 5). The extreme ends of the area under investigation, Bordjiesrif in the south and Swakopmund in the north, showed the highest mean annual seawater temperatures of 15.5 and 14.7 °C respectively. Maximum and minimum mean monthly temperatures followed the same general trends as the mean temperatures. Again, the sites at the extremes of the study area (Bordjiesrif and Swakopmund) had the highest minimum and maximum monthly mean temperatures. Swakopmund showed the biggest range between mean monthly maximum and minimum (8 °C).

Average seawater temperatures for the South African west coast for a period of about 30 years (1981–2011). The blue line indicates the mean annual temperatures while the green and brown lines indicate the maximum and minimum monthly mean temperatures respectively. Data from the South African Weather Services. Bordjiesrif data are from a temperature recorder at 8 m depth from the Department of Agriculture Forestry and Fisheries. Swakopmund data are from NASSA SST corrected by AJ Smit
Wind and waves
For almost all sites, average wind direction (Online Resource 1; Fig. S7 a) was South-South Westerly between 200° and 210°, except for Lüderitz where it was South (177°). Average wind speeds (Fig. S7 b) were highest at the most southerly sites (Cape Point and Kommetjie), and otherwise rather variable between 4 and 6 knots. Average and maximum wave height (Online Resource 1; Fig. S7 b) showed a trend of highest values at the southernmost sites (Cape Point and Kommetjie) and lowest at the most northern site (Swakopmund), with intermediate values at the intervening sites.
Multivariate analysis
The PCA of environmental factors (Fig. 6) partly separated out three groups of sites. The first group included Cape Point and Kommetjie (the most southerly sites), the second group included Lüderitz and Swakopmund (the most northerly sites), and the final group comprised the sites that lie in between (Jacobs Bay, Shelly Point, Elands Bay, and Doring Bay). The direction of the arrows revealed that the most southerly sites had high values for wind speed, and low values for those factors linked with water turbidity (Chl a, PIC, POC).

PCA plot of environmental factors at different sites. Environmental factors: temp temperature, wi spd wind speed, wi dir wind direction, w dir wave direction, w hgt wave height, w per wave period, cloud relative cloud cover, chl a chlorophyll a, PInC particulate inorganic carbon, POC particulate organic carbon
Discussion
This study investigated the morphology of shallow-water (0–5 m) E. maxima and L. pallida sporophytes over most of their distribution ranges and attempted to pinpoint the environmental factors that might be responsible for any morphological patterns. The change in dominance between the two kelps E. maxima and L. pallida along a latitudinal gradient in kelp beds on the west coast of Southern Africa confirms previous anecdotal observations (e.g. Stegenga et al. 1997). Both relative occurrence (Fig. 2a) and biomass data (Fig. 2b) showed a clear transition from the south, where inshore beds are dominated by E. maxima, to the north, where beds are dominated by L. pallida.
Along this latitudinal gradient, there was no change in the morphology of E. maxima sporophytes. However, for L. pallida there was a distinct gradient in relative hollowness, quantitatively confirming previous observations that northern plants have hollow stipes (e.g. Stegenga et al. 1997).
Saccharina latissima (as Laminaria agardhii Kjellman) and Saccharina longicruris (as Laminaria longicruris) (now a single species) showed decreasing stipe length and hollowness as wave exposure increased at three sites in Nova Scotia (Chapman 1973). Another example of a hollow-stiped kelp is the annual Nereocystis luetkeana (Denny et al. 1997). However, the only other comparable study is that of Matson and Edwards (2006) who showed a similar gradual change in stipe hollowness of Ecklonia arborea (as Eisenia arborea) northwards along the west coast of North America. However, they offered no explanation for this phenomenon.
Having established clear latitudinal patterns in relative dominance of L. pallida and E. maxima, and in relative hollowness of L. pallida stipes, the question arises—if the causes are environmental, what factors could be involved?
Multivariate analyses
Because the morphology of E. maxima in shallow water was unaffected by geographical location (e.g. Online Resource 1; Fig. S4, PCA), clearly this species is morphologically unaffected by whatever environmental factors affect the morphology of L. pallida.
In L. pallida, the PCA showed a latitudinal trend related most strongly to relative hollowness of the stipes (Online Resource 1; Fig. S5) with the other major contributing factors being percent hollowness, stipe weight, and stipe length, as indicated by the direction and length of the vectors. However, this does not answer the question: what might cause this hollowness pattern along a geographical gradient, in shallow water?
Temperature
Most of the environmental factors that we investigated showed no relationship with the clear latitudinal patterns in Laminaria dominance and hollowness. Water temperatures are very similar at Swakopmund and Bordjiesrif and markedly higher than at the intermediate sites, but Swakopmund has only L. pallida and Bordjiesrif only E. maxima in shallow water. Despite the importance of temperature in setting seaweed species’ biogeographical limits (Bolton and Anderson 1987; Lüning 1990; Anderson et al. 2007), factors other than temperature must drive the northward dominance and increasing hollowness of L. pallida.
Nutrients can also be discounted, because of the close relationship (negative correlation) between temperature and nutrients on this coast (Dieckmann 1980; Waldron and Probyn 1992). Furthermore, all sites lie within the Benguela Marine Province, with relatively high nutrient levels that are associated with upwelling.
We do not consider wave height to be a possible explanation. Although there was a slight trend of decreasing mean wave height along this coast, the differences are small in real terms: a maximum of 3 m at Cape Point versus 2 m at Swakopmund. Furthermore, although there were no data for Bordjiesrif, it is the most sheltered site (it lies within False Bay and faces away from the prevailing SW swells), yet had no L. pallida, whereas Swakopmund, the next most sheltered site, had only that species. In previous studies of wave exposure and kelp morphology, very sheltered sites were contrasted with very exposed sites (e.g. Norton et al. 1982; Serisawa et al. 2002, 2003; Hurd and Pilditch 2011), whereas all our sites are relatively wave exposed in this context.
Light/turbidity
Among the environmental variables measured here, those that affect underwater light showed the most convincing latitudinal gradients. Underwater light could be affected by water turbidity, cloud cover or fog, daylength, and depth (Shirley 1929; Burgess 2009).
Depth can be discounted as a latitudinal factor, since all sites were sampled in the same depth range. Daylength is unlikely to be the cause of the geographic variation in L. pallida. Although seasonal changes in daylength affect growth rate in L. pallida (Dieckmann 1980), the maximum difference in daylength between even Swakopmund and Bordjiesrif is relatively slight (about 49 min). Photoperiodic daylength effects on algal life histories/growth take the form of on/off switches (Lüning and Dring 1972; Izquierdo et al. 2002; Muñoz et al. 2004; Oppliger et al. 2012) and would not explain the gradient observed in the Laminaria morphological changes.
Clouds and fog cause scattering of incident light and reduce the amount of light that reaches the earth (Blinn 1982). Despite anecdotal reports of increasing fog northwards up the west coast, the satellite image data for 2011 showed no significant trends except for significantly more cloud/fog cover at Swakopmund. This however, cannot account for differences in the L. pallida at the other sites, and we are left with water turbidity as the only factor that shows a convincing latitudinal gradient.
Data from NASA satellite imagery showed a northward increase in turbidity for all three variables that were measured (Fig. 4), corroborating our visual impressions from decades of working on this coast. Although it was logistically impossible to measure underwater light over a long period (and at many sites), we believe that there is a clear, decreasing trend northwards. On this coast, inshore water clarity is strongly affected by upwelling, which brings in cold, clear, nutrient-rich water, and upwelling is directly related to the strength and duration of SW-SE winds (Shannon 1985; Monteiro and Largier 1999; Berger and Wefer 2002). Wind speeds (Fig. S7 b) were highest at the southern sites, where upwelling is highest, and turbidity lowest. In contrast, the most northern site, Swakopmund, had a significantly lower wind speed, and there is weak and sporadic upwelling (Nelson and Hutchings 1983).
On the west coast, a gradual decrease in underwater light corresponds well with the gradual increase in dominance of L. pallida in the nearshore environment (and the increasing prevalence of hollow stipes in this species). From this, it is hypothesised that L. pallida may be adapted to lower light compared to E. maxima.
This seems reasonable, also because in the south L. pallida grows under the canopy of E. maxima, and all along the coast L. pallida forms extensive beds in deeper water down to 20 m and more. We believe that where there is good light in shallows (e.g. Cape Peninsula, down to about 5 m depth), E. maxima outcompetes L. pallida. While E. maxima may well be depth-limited by how long its stipe can grow (because it seldom occurs deeper than about 12 m), it might also be depth-limited at more than 12 m because even when the water is clear, at that depth light would become limiting. Using the Jerlov (1977) system of optical water types, Lüning (1990) showed that water clarity can have an enormous effect on light penetration: the deepest Laminaria in Helgoland is found at 8 m depth, but in the Mediterranean at 95 m. The quantity of light at the former site is drastically reduced by turbidity. It is possible that on the Southern African west coast, the northward gradient of increasing dominance of L. pallida in shallow water is a result of the gradient in water turbidity and hence light: as inshore light decreases, L. pallida may be able to outcompete E. maxima.
Hollowness
This study demonstrated a gradual change in turbidity along the distribution range of L. pallida. Similarly, a gradual increase in the dominance of L. pallida northwards was observed, which corresponded with a gradual increase in the degree of hollowness of L. pallida stipes. Although the degree of hollowness in E. maxima does not change along the west coast, the degree of hollowness in L. pallida does vary, and this begs the question: why does L. pallida have a hollow stipe in shallow water on the northern parts of the coastline? Two reasons are considered.
The first possibility, apart from the obvious reason of creating buoyancy to keep the plant upright, is that because L. pallida is growing in a much more wave-stressed (shallower) environment, the hollowness may make it more flexible and better able to survive (see Friedland and Denny 1995 and Denny et al. 1997). The flexibility of kelp stipes is an important property that allows these plants to become large under conditions of very high wave action (Denny et al. 1997). The hydrodynamic forces imposed on plants are reduced by their flexibility and their ability to ‘go with the flow’ (Koehl 1986; Denny et al. 1997). Several studies have shown the importance of flexibility of seaweeds in the intertidal (Friedland and Denny 1995; Utter and Denny 1996; Denny et al. 1997). However, other mechanical properties of the tissues also influence the survivorship of individuals. Demes et al. (2013) showed that Egregia menziesii had a higher survival rate if the fronds are weak because the fronds self-prune, decreasing hydrodynamic drag forces and decreasing mortality. Both E. maxima (e.g. Mann et al. 1979) and L. pallida (Dieckmann 1980) lose material from the ends of their fronds, effectively ‘self-pruning’ and limiting the drag force exerted on the sporophytes. It appears that self-pruning, together with the overall flexibility of the sporophytes, is sufficient to allow them to survive in the extremely turbulent shallow water of the Southern African west coast. But most importantly, in L. pallida, increasing hollowness did not make the stipes more flexible, and could therefore not be an adaptation to shallower, more turbulent water: we suggest that hollowness has other adaptive advantages. Another adaptive advantage of a hollow stipe might be that there is less investment in energy, compared to building a solid medulla. This would enable the sporophytes to grow more quickly towards the surface, where more light is available. Although a solid cylinder is stronger than a hollow cylinder in absolute terms, the ratio of strength to weight is higher in a hollow cylinder of the same diameter (for a mathematical explanation, see Bhandari 2010). By becoming hollow, a stipe will lose some strength, but lose a disproportionately greater amount of weight (i.e. medullary tissue). If a slight loss of strength is less important than a relatively large saving in energy (in the form of the non-photosynthetic medullary tissue that would have been necessary to produce a solid stipe), these plants will have an advantage, because they should be able to grow faster than solid plants. L. pallida may be exploiting this relationship to grow faster in more turbid water, leading to a northward increase in hollowness. This hollowness may therefore simply be an energy-saving adaptation, since the present results show that the force needed to bend a L. pallida stipe to a predetermined position does not depend on the hollowness of the stipe (Online Resource 1; Fig. S3, Table S1). This suggests that L. pallida sporophytes are not hollow to mitigate the higher wave action that prevails in shallow water. Also, at Swakopmund, where L. pallida had the highest percentage hollowness, wave height was lowest among all the sites measured (Online Resource 1; Fig. S7 b), and all indicators of turbidity were highest (Fig. 3).
The PCA model separated out three groups and showed that the main driving vectors for this separation were wind speed and turbidity, acting in opposite directions. It is established that upwelling is wind-induced (Andrews and Hutchings 1980; Shannon 1985) and that during periods of upwelling, water is clearer and underwater irradiance is considerably higher in shallow water (Anderson and Bolton 1985). This inverse relationship between wind speed (Online Resource 1; Fig. S7 b) and water turbidity (Fig. 4) is evident for various sites along the west coast, and supports the idea that turbidity is an important environmental factor that must influence kelps along this coast. Whether there is a genetic influence on the morphology of L. pallida (e.g. incipient speciation: see Roberson and Coyer 2004) could only be investigated by a study of population genetics of this kelp.
Conclusion
Laminaria pallida sporophytes gradually replace E. maxima northwards in shallow water on the Southern African west coast until they completely replace E. maxima in central Namibia. Concurrently, the stipes of L. pallida become more hollow moving northwards. The results discount a number of environmental variables that could be responsible for the distribution pattern of L. pallida and the hollowness of its stipe. Seawater temperature, and by inference nutrient levels, do not correlate with these patterns, but there is a strong geographic gradient in water turbidity that must directly affect underwater light. Cloud/fog cover was shown to be potentially important in reducing inshore light levels only at the northern-most site, Swakopmund. However, we would expect that other sites in northern Namibia (for which we do not have data), where the desert coastline impinges on cool upwelled water, are also likely to have high cloud/fog cover. Dust storms from deserts can also increase turbidity in the adjacent ocean, and although Southern Africa deserts are not a major source of dust (Washington et al. 2003), offshore transport of particulates from the Namib desert may contribute to turbidity in these waters and could explain why E. maxima is absent from central and northern Namibia. From these results, we conclude that there is strong evidence that L. pallida progressively outcompetes E. maxima northward because it is more low-light tolerant and that by developing a hollow stipe it may grow faster, allowing it to reach the surface quicker.
References
Anderson RJ, Bolton JJ (1985) Suitability of the agarophyte Suhria vittata (L.) J. Ag. (Rhodophyta: Gelidiaceae) for mariculture: geographical distribution, reproductive phenology and growth of sporelings in culture in relation to light and temperature. S Afr J Mar Sci 3:169–178
Anderson RJ, Carrick P, Levitt G, Share A (1997) Holdfasts of adult kelp Ecklonia maxima provide refuges from grazing for recruitment of juvenile kelps. Mar Ecol Prog Ser 159:265–273
Anderson RJ, Rand A, Rothman MD, Share A, Bolton JJ (2007) Mapping and quantifying the South African kelp resource. Afr J Mar Sci 29:369–378
Andrews W, Hutchings L (1980) Upwelling in the southern Benguela Current. Prog Oceanogr 9:1–81
Berger W, Wefer G (2002) On the reconstruction of upwelling history: Namibia upwelling in context. Mar Geol 180:3–28
Bhandari V (2010) Design of machine elements. Tata McGraw-Hill Education Private Limited, Delhi, 930 pp
Blinn JF (1982) Light reflection functions for simulation of clouds and dusty surfaces. ACM SIGGRAPH Comp Graph 16(3):21–29
Bolton JJ, Anderson RJ (1987) Temperature tolerances of two southern African Ecklonia species (Alariaceae: Laminariales) and of hybrids between them. Mar Biol 96:293–297
Bolton JJ, Anderson RJ, Smit AJ, Rothman MD (2012) South African kelp moving eastwards: the discovery of Ecklonia maxima (Osbeck) Papenfuss at De Hoop Nature Reserve on the south coast of South Africa. Afr J Mar Sci 34:147–151
Burgess P (2009) Variation in light intensity at different latitudes and seasons, effects of cloud cover, and the amount of direct and diffused light Available at: http://ccfg.org.uk/conferences/downloads/P_Burgess.pdf. Accessed 17 Oct 2016
Chapman ARO (1973) Phenetic variability of stipe morphology in relation to season, exposure, and depth in the non-digitate complex of Laminaria Lamour (Phaeophyta, Laminariales) in Nova Scotia. Phycologia 12:53–57
Chen Z, Hu C, Muller-Karger F (2007) Monitoring turbidity in Tampa Bay using MODIS/Aqua 250-m imagery. Remote Sens Environ 109:207–220
Clarke K, Gorley R (2006) User manual/tutorial. Primer-E Ltd., Plymouth, 93 pp
Demes KW, Pruitt JN, Harley CD, Carrington E (2013) Survival of the weakest: increased frond mechanical strength in a wave-swept kelp inhibits self-pruning and increases whole-plant mortality. Funct Ecol 27:439–445
Denny M, Gaylord B, Cowen E (1997) Flow and flexibility. II. The roles of size and shape in determining wave forces on the bull kelp Nereocystis luetkeana. J Exp Biol 200:3165–3183
Dieckmann GS (1980) Aspects of the ecology of Laminaria pallida (Grev.) J. Ag. off the Cape Peninsula (South Africa). Bot Mar 23:579–585
Doxaran D, Froidefond J, Castaing P, Babin M (2009) Dynamics of the turbidity maximum zone in a macrotidal estuary (the Gironde, France): observations from field and MODIS satellite data. Estuar Coast Shelf Sci 81:321–332
Field JG, Griffiths CL, Griffiths RJ, Jarman N, Zoutendyk NP, Velimirov B, Bowes A (1980) Variation in structure and biomass of kelp communities along the South-west Cape coast. Trans Roy Soc S Afr 44:145–203
Fowler-Walker MJ, Wernberg T, Connell SD (2006) Differences in kelp morphology between wave sheltered and exposed localities: morphologically plastic or fixed traits? Mar Biol 148:755–767
Friedland MT, Denny MW (1995) Surviving hydrodynamic forces in a wave-swept environment: consequences of morphology in the feather boa kelp, Egregia menziesii (Turner). J Exp Mar Biol Ecol 190:109–133
Gaylord B, Denny M (1997) Flow and flexibility. I. Effects of size, shape and stiffness in determining wave forces on the stipitate kelps Eisenia arborea and Pterygophora californica. J Exp Biol 200:3141–3164
Hornik V (2014) Windguru http://www.windguru.cz/int/
Hurd CL, Pilditch CA (2011) Flow-induced morphological variation affect diffusion boundary-layer thickness of Macrocystis pyrifera (Heterokontophyta, Laminariales). J Phycol 47:341–351
Izquierdo J, Pérez-Ruzafa IM, Gallardo T (2002) Effect of temperature and photon fluence rate on gametophytes and young sporophytes of Laminaria ochroleuca Pylaie. Helgol Mar Res 55:285–292
Jarman NT, Carter R (1981) The primary producers of the inshore regions of the Benguela. Trans Roy Soc S Afr 44:321–326
Jerlov N (1977) Classification of sea water in terms of quanta irradiance. J Conseil 37(3):281–287
Klochkova TA, Kim GH, Lee KM, Choi H, Belij MN, Klochkova NG (2010) Brown algae (Phaeophyceae) from Russian far eastern seas: re-evaluation of Laminaria multiplicata Petrov et Suchovejeva. Algae 25:77–87
Koehl M (1986) Seaweeds in moving water: form and mechanical function. In: Givnish TJ (ed) On the economy of plant form and function. Cambridge Univ. Press, Cambridge, pp 603–634
Lüning K, Dring M (1972) Reproduction induced by blue light in female gametophytes of Laminaria saccharina. Planta 104:252–256
Lüning K (1990) Seaweeds: their environment, biogeography, and scophysiology. Wiley-Interscience, New York, 527pp
Marins BV, Amado-Filho GM, Barreto MB, Longo LL (2012) Taxonomy of the southwestern Atlantic endemic kelp: Laminaria abyssalis and Laminaria brasiliensis (Phaeophyceae, Laminariales) are not different species. Phycol Res 60:51–60
Mann K, Jarman N, Dieckmann G (1979) Development of a method for measuring the productivity of the kelp Ecklonia maxima (Osbeck) Papenfuss. Trans R Soc S Afr 44:27–41
Matson PG, Edwards MS (2006) Latitudinal variation in stipe hollowing in Eisenia arborea (Phaeophyceae, Laminariales). Phycologia 45:343–348
Molloy FJ, Bolton JJ (1990) Utilized and potentially utilizable seaweeds on the Namibian coast: biogeography and accessibility. Hydrobiologia 204/205:293–299
Molloy FJ, Bolton JJ (1996) The effects of wave exposure and depth on the morphology of inshore populations of the Namibian kelp, Laminaria schinzii Foslie. Bot Mar 39:525–531
Monteiro P, Largier J (1999) Thermal stratification in Saldanha Bay (South Africa) and subtidal, density-driven exchange with the coastal waters of the Benguela upwelling system. Estuar Coast Shelf Sci 49:877–890
Muñoz V, Hernández-González MC, Buschmann AH, Graham M, Vásquez J (2004) Variability in per capita oogonia and sporophyte production from giant kelp gametophytes (Macrocystis pyrifera, Phaeophyceae). Rev Chil Hist Nat 77:639–647
National Aeronautics and Space Administration, Moderate resolution image spectroradiometer: http://modis.gsfc.nasa.gov. Accessed 3 Feb 2014
Nelson G, Hutchings L (1983) The Benguela upwelling area. Prog Oceanogr 12:333–356
Norton T, Mathieson A, Neushul M (1982) A review of some aspects of form and function in seaweeds. Bot Mar 25:501–510
Oppliger LV, Correa JA, Engelen AH, Tellier F, Vieira V, Faugeron S, Valero M, Gomez G, Destombe C (2012) Temperature effects on gametophyte life-history traits and geographic distribution of two cryptic kelp species. PLoS One 7:e39289
Petus C, Chust G, Gohin F, Doxaran D, Froidefond J, Sagarminaga Y (2010) Estimating turbidity and total suspended matter in the Adour River plume (South Bay of Biscay) using MODIS 250-m imagery. Cont Shelf Res 30:379–392
Roberson LM, Coyer JA (2004) Variation in blade morphology of the kelp Eisenia arborea: incipient speciation due to local water motion? Mar Ecol Prog Ser 282:115–128
Rothman MD (2006) Investigations into the harvesting ecology of the South African kelp Ecklonia maxima (Alariaceae, Laminariales). Dissertation, University of Cape Town, M.Sc
Rothman MD, Mattio L, Wernberg T, Anderson RJ, Uwai S, Mohring MB, Bolton JJ (2015) A molecular investigation of the genus Ecklonia (Phaeophyceae, Laminariales) with special focus on the Southern Hemisphere. J Phycol 51:236–246
Serisawa Y, Akino H, Matsuyama K, Ohno M, Tanaka J, Yokohama Y (2002) Morphometric study of Ecklonia cava (Laminariales, Phaeophyta) sporophytes in two localities with different temperature conditions. Phycol Res 50:193–199
Serisawa Y, Aoki M, Hirata T, Bellgrove A, Kurashima A, Tsuchiya Y, Toshihiko S, Ueda H, Yokohama Y (2003) Growth and survival rates of large-type sporophytes of Ecklonia cava transplanted to a growth environment with small-type sporophytes. J Appl Phycol 15:311–318
Shannon L (1985) The Benguela ecosystem. I: evolution of the Benguela physical features and processes. Oceanogr Mar Biol 23:105–182
Shannon L, Pillar S (1986) The Benguela ecosystem. Part III. Plankton. Oceanogr Mar Biol Annu Rev 24:65–170
Shirley HL (1929) The influence of light intensity and light quality upon the growth of plants. Am J Bot 16:354–390
Simons RH & Jarman NG (1981) Subcommercial harvesting of a kelp on a South African shore. In: Levring T (ed) Proceedings of the 10th International Seaweed Symposium. pp 731-736
Smale DA, Wernberg T (2009) Satellite-derived SST data as a proxy for water temperature in nearshore benthic ecology. Mar Ecol Prog Ser 387:27–37
Smit AJ, Roberts M, Anderson RJ, Dufois F, Dudley SF, Bornman TG, Olbers J, Bolton JJ (2013) A coastal seawater temperature dataset for biogeographical studies: large biases between in situ and remotely-sensed data sets around the coast of South Africa. PLoS One 8(12):e81944
Stegenga H, Bolton JJ, Anderson RJ (1997) Seaweeds of the South African west coast. Contrib Bolus Herb 18:3–637
Tsutsui I, Arai S, Terawaki T, Ohno M (1996) A morphometric comparison of Ecklonia kurome (Laminariales, Phaeophyta) from Japan. Phycol Res 44:215–222
Utter B, Denny M (1996) Wave-induced forces on the giant kelp Macrocystis pyrifera (Agardh): field test of a computational model. J Exp Biol 199:2645–2654
Velimirov B, Field JG, Griffiths CL, Zoutendyk P (1977) The ecology of kelp bed communities in the Benguela upwelling system. Helgo Meeresun 30:495–518
Velimirov B, Griffiths C (1979) Wave-induced kelp movement and its importance for community structure. Bot Mar 22:169–172
Waldron H, Probyn T (1992) Nitrate supply and potential new production in the Benguela upwelling system. S Afr J Mar Sci 12:29–39
Washington R, Todd M, Middleton NJ, Goudie AS (2003) Dust-storm source areas determined by the total ozone monitoring spectrometer and surface observations. Ann Assoc Am Geol 93:297–313
Wernberg T, Coleman M, Fairhead A, Miller S, Thomsen M (2003) Morphology of Ecklonia radiata (Phaeophyta: Laminarales) along its geographic distribution in south-western Australia and Australasia. Mar Biol 143:47–55
Acknowledgements
This study was supported by the Department of Agriculture, Forestry and Fisheries, South Africa, the University of Cape Town, and the National Research Foundation (NRF). Thank you to Cathy Boucher for the artwork and AJ Smit for the Namibian temperature data. Lastly, thank you to M. Noffke, J. Fridjhon, S. Conrad, D. Williams and S. John for assisting with field work and sample collections.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Rothman, M.D., Bolton, J.J., Stekoll, M.S. et al. Geographical variation in morphology of the two dominant kelp species, Ecklonia maxima and Laminaria pallida (Phaeophyceae, Laminariales), on the west coast of Southern Africa. J Appl Phycol 29, 2627–2639 (2017). https://doi.org/10.1007/s10811-017-1255-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-017-1255-7