
Abstract
The non-native aquatic fern giant salvinia, Salvinia molesta Mitchell (Salviniaceae), poses a risk to freshwater ecosystems through limiting light penetration, decreasing submerged aquatic vegetation (SAV) abundance, altering water quality, and potentially leading to changes in macroinvertebrate community structure. Here, we conducted repeated quarterly field surveys and measured light, nutrients, water quality, and aquatic macroinvertebrate community composition and energetic value to detect effects from giant salvinia invasion. Giant salvinia reduced dissolved oxygen, pH and light availability in the aquatic environment, and increased the concentration of orthophosphate and ammonium. Following initial colonization, macroinvertebrate communities in giant salvinia resembled SAV communities dominated by aquatic insects, however, richness and relative abundance in giant salvinia decreased over time, resulting in a community populated by few taxa, primarily crustaceans. Total macroinvertebrate energetic value in giant salvinia was significantly lower than SAV communities. Giant salvinia invasion changed habitat composition, triggered internal nutrient loading, and reduced macroinvertebrate abundance, diversity, and ecosystem productivity. Our findings demonstrate larger ecological impacts from giant salvinia than previously reported, including potential disruption to the transfer of energy between trophic levels.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Freshwater marshes occur worldwide, comprising 1.9% of land area globally (Lehner & Döll, 2004) and are of conservation concern (Finlayson, 2016). The introduction of non-indigenous aquatic plants impacts environmental conditions and the state of the ecosystem in freshwater marshes. Invasive free-floating aquatic plants can negatively affect native submerged aquatic vegetation (hereafter referred as SAV) abundance and ecosystem function by reducing light penetration into the water, depleting nutrients (Rommens et al., 2003), limiting gas exchange (Attermeyer et al., 2016), and altering nutrient cycling (Wang & Yan, 2017). Giant salvinia (Salvinia molesta Mitchell; Salviniaceae) is a free-floating aquatic fern native to Brazil that has become problematic globally. Giant salvinia was first observed outside its native Brazilian range in 1939 and has been identified in 61 countries (Luque et al., 2014; CABI, 2019). Giant salvinia has a rapid growth rate, doubling its biomass in 53 h (Cary & Weerts, 1983), and covering entire water surface in days (Mitchell & Tur 1975). Thus, giant salvinia is considered highly disruptive and difficult to manage in freshwater habitats (Hill et al., 2020).
Coastal freshwater marshes provide habitat for numerous species (Engle, 2011) and ecosystem services, including flood mitigation (Pattison-Williams et al., 2018), removal of pollutants and excess nutrients (Vymazal, 2007), and recreational and cultural activities (Boucek & Rehage, 2015; Vargas-Lopez et al., 2020). Submerged aquatic vegetation is important in marsh structure and function influencing water quality (Caraco et al., 2006) and nutrient cycling (Ferreira et al., 2018). The structural compounds in SAV are slow to breakdown, trapping carbon in the sediment and acting as a carbon sink (Hinson et al., 2017). In freshwater, SAV provides habitat and food resources for waterfowl (Wilson et al., 2002), macroinvertebrates and fish (Fisher et al., 2012; Schultz & Dibble, 2012). A rich diversity of SAV increases habitat complexity, offering refuge to a high number of species (Cheruvelil et al., 2002). This habitat can support a high abundance and diversity of invertebrates, which offers a prey source for small bodied and young fish (Schultz & Dibble, 2012) and attracts larger bodied predatory fish. The loss of SAV would decrease habitat abundance and complexity, altering species interactions and modifying the macroinvertebrate community.
Macroinvertebrates are critical to the function of the marsh and serve as a link between primary production and the rest of the trophic web (Covich et al., 1999; Jones & Sayer, 2003). Disturbance events, such as the introduction of an invasive species, can alter environmental conditions and habitat complexity. Changes to habitat and environmental conditions of the aquatic ecosystem would select tolerant taxa and modify species interactions, such as competition and predator–prey relationships, potentially resulting in shifts in macroinvertebrate community structure and productivity. Therefore, an altered macroinvertebrate community could be consequential for the aquatic ecosystem, yet it has not been studied as a result of a floating invasive plant within coastal freshwater marshes. The objective of this study was to examine how giant salvinia affected environmental conditions, to test whether a shift in macroinvertebrate community structure occurred, and determine whether the shift modified total energy of the macroinvertebrate community. We determined these impacts with repeated quarterly sampling of locations abundant with SAV and locations invaded with giant salvinia. Air breathing and low dissolved oxygen-tolerant insects are common in warm aquatic systems, especially in this region (Colon-Gaud et al., 2004; Kaller & Kelso, 2007; Justus et al., 2012; Kang & King, 2013; Parys et al., 2013), therefore, we expected the differences in physical structure and water chemistry to be more important (Colon-Gaud et al., 2004; Fisher & Kelso, 2007, and de Silva & Henry, 2020). Thus, we hypothesized macroinvertebrate communities in giant salvinia would support a diverse and abundant macroinvertebrate community, however, dominant taxa could be different in giant salvinia relative to SAV.
Materials and methods
Field location
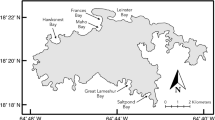
Surveys were conducted in a subtropical, coastal, freshwater marsh located in Cameron Parish (county), Louisiana, USA. This region is classified as a subtropical ecoregion, supporting both temperate and tropical organisms. The study site (29.859628, − 92.956331; ~ 22,300 ha) is privately owned and predominantly used for waterfowl hunting and recreational fishing, and is managed to control invasive species. Giant salvinia was first reported in 2000 and has since persisted in the environment, requiring continued chemical and biological control efforts. Native SAV such as Ceratophyllum demersum L., Cabomba caroliniana A. Gray, and Ruppia maritima L. are common in the marsh, and locations with one or more of these SAV species were used for reference sampling (Fig. 1), with C. caroliniana being the predominant SAV sampled. Based on monitoring by the landowner and our team over previous years, all sampling locations were dominated by SAV prior to the study. Cold temperatures during the winter of 2017–2018 reduced the occurrence giant salvinia from the area and effectively shifted the aquatic environment to SAV dominant habitat. Sampling commenced in August 2018, immediately following giant salvinia establishment and on three further occasions until May 2019, due to control efforts.

Field location was in a subtropical, coastal, freshwater marsh in Cameron Parish, Louisiana. Sampling sites contained giant salvinia and submerged aquatic vegetation (SAV). The property (~ 22,300 ha) is privately owned and predominantly used for recreational activities and is managed to control invasive species. Sampling occurred quarterly, commencing in August 2018 and concluding in May 2019
Environmental and plant quality variables
Physical and chemical variables were measured at each sampling location. Dissolved oxygen (DO), specific conductance, pH, temperature, and ammonium (NH4) were sampled with a handheld multiprobe (Pro-DS5, YSI Incorporated, Yellow Springs, OH, USA). Water samples were collected in sterile polyethylene containers for chlorophyll-a, phosphorus (orthophosphate, P-ortho) and nitrogen (nitrate, NO3, and nitrite, NO2) analyses. Samples were kept on ice during transport and refrigerated (4 °C) after returning to the laboratory. Phosphorus, nitrate, and nitrite were analyzed by Hach Method 8048 (US EPA Approved), Hach Method 8192 (Cadmium Reduction Method), and Hach Method 8507 (US EPA Approved), respectively, using a spectrophotometer (DR/2500, Hach Company, Inc., Loveland, CO, USA). Five hundred mL of water was filtered using glass fiber filters, and chlorophyll-a was measured by US EPA Method 445.0 (Arar & Collins, 1997) using a fluorometer (TD 700, Turner Designs, Inc., San Jose, CA).
Light intensity was sampled directly above the water surface, directly below the water surface (0.01 m), at 0.10 and 0.20 m, then in 0.20 m increments until the bottom using a light meter (MW700 light meter, Milwaukee Instruments, Rocky Mount, NC, USA). Giant salvinia mat thickness was determined as the distance between the top and bottom of the mat using a plastic tray (0.30 × 0.40 × 0.01, L × W × H, m). The tray was slid through the giant salvinia vertically, once through, the tray was turned horizontally against the bottom of the mat, thickness was determined with a rope, marked in 0.01 m increments, attached to the middle of the tray. Submerged frond (root) length was recorded for 10 randomly selected giant salvinia plants.
Macroinvertebrate and plant biomass sampling
Three giant salvinia invaded locations and three native SAV dominant locations were sampled quarterly over nine months (Fig. 1). At each location, three samples were collected for a total of 72 samples during the study period (72 samples = 2 macrophyte types × 3 locations × 4 quarters × 3 samples). Two samplers were used because macroinvertebrates in SAV are typically stratified vertically throughout the plant bed, whereas when giant salvinia invades, macroinvertebrates orient horizontally to the floating plants. In SAV, a suitcase sampler was used consisting of two 0.65 × 0.45 m hinged panels covered with 0.600 mm mesh. The panels were connected using a hinge and four attached rings, with rope running through, which standardized the width the sampler could open at 0.55 m. The sampling area was triangular prism shaped (0.65 × 0.65 × 0.55 × 0.45 m) that enclosed a volume of 0.07 m3 (Colon-Gaud & Kelso, 2003). Sampling was done by wading to undisturbed SAV beds then lowering the suitcase sampler into the bed. Once in place the trap would be closed and secured shut with latches. Vegetation sticking out of the edges of the sampler was trimmed away. Giant salvinia was sampled using a 0.500 mm mesh lift net (0.29 × 0.35 × 0.20 m; 0.02 m3 volume; Kaller et al., 2013). Samples were collected from the giant salvinia mat by wading to undisturbed areas. When sampling, the lift trap was placed through the giant salvinia mat with top edge first, and quickly scooped under the mat and lifted out of the water. Giant salvinia was in tertiary growth stage for all giant salvinia sites during the study. Material from the samplers were stored in a re-sealable plastic bag with water and placed in the iced cooler until processing.
Samples were processed in the laboratory following collection. Samples were processed by flushing and rinsing plant material in 75 l plastic bin filled with water. Plant matter removed from the bin was processed to determine dry biomass. Plant matter was dried in an oven for 72 h at 65 °C to measure dry mass. Macroinvertebrates rinsed from plant material and captured on a 0.500 mm sieve were then enumerated and identified to lowest taxonomic level, usually genus.
Caloric value
Caloric values from published data were calculated to determine potential energy of the macroinvertebrate community (Welsh, 1975; Fredrickson & Reid, 1988; Foley, 2015). Only macroinvertebrates with published caloric values were used in the analysis. To quantify calorific value, we multiplied taxa counts per gram dry biomass by the estimated value of each taxa, then summed the value for all taxa in the sample.
Statistical analysis
Analyses first examined differences among water quality and plant characteristics among months and habitats. Next, multivariate and community analyses examined invertebrate response among habitats and months. Finally, multivariate analyses related invertebrate responses to specific water quality and plant variables. Analyses were conducted using R statistical software version 3.4.4 (R Core Team, 2013).
Generalized linear models (GLM) compared water quality and plant variables among habitats and months. For all variables, the selected combination of link and distribution was determined by comparing candidates and fit statistics. A log link, Gaussian distribution GLM was used for temperature. Log link, gamma distribution GLMs were used for DO, pH, P-ortho, chlorophyll-a, NH4, and plant biomass. Inverse link, gamma distribution GLMs were used for specific conductivity, submerged frond length, and giant salvinia mat thickness. Logit link, binomial distribution GLMs were used for NO3 and NO2. Log link, gamma distribution GLM was used for light availability at 0.01 m depth.
For macroinvertebrate analysis, we removed taxa that had a total abundance of less than four individuals collected during the study. For comparability, we standardized species richness and abundance per gram of dry plant biomass. GLMs with log links and gamma distributions were used to examine abundance and richness between habitats and among sampling dates. Log link, gamma distribution GLM was used for caloric value difference between habitat and month. A multivariate generalized linear model (MGLM), with a negative binomial distribution and log link, was used to test for differences in macroinvertebrate community between habitats and among sampling dates (package mvabund; Warton et al., 2012). The model was adjusted for multiple comparisons using a step-down resampling procedure.
Nonmetric multidimensional scaling (NMDS) of a Bray–Curtis similarity matrix of taxa (k = 4) was used to visualize the similarity between habitat types and among months (Clarke, 1993). Comparisons of exploratory ordinations (e.g., principal component analysis and detrended correspondence analysis) indicated that NMDS would be the appropriate analysis (package vegan, Oksanen et al., 2019). Analysis of variance using distance matrices (ADONIS) was used to test the solution from NMDS analysis (package Vegan, Oksanen et al., 2019). Multivariate dispersion test showed that giant salvinia (0.13) and SAV (0.12) were similar in multivariate distance, thus, the analysis was appropriate for these data (Anderson & Walsh, 2013). Association with habitat (Ordiselect, package Vegan, Oksanen et al., 2019) and community dissimilarity between habitats (similarity percentage, SIMPER, package Vegan, Oksanen et al., 2019) also were examined.
A redundancy analysis (RDA) examined how variation in macroinvertebrate community composition was explained by environmental gradients described by water quality variables and plant characteristics, following comparisons to find the method most appropriate to the data (package Vegan, Oksanen et al., 2019). Variables selected for the RDA were initially screened by linear discriminant analysis (LDA), for appropriateness for RDA, and MGLM, described previously, to indicate which environmental variables significantly varied with habitat and month (LDA, package MASS, Chang, 2015), and these variables were used in the RDA.
Results
Environmental variables and plant quality
Water quality measurements showed variation between habitat and among months (Table 1). Dissolved oxygen (t-value = 4.74, P < 0.01) and pH (t-value = 4.56, P < 0.01) were reduced in giant salvinia. Dissolved oxygen was 2.48 times higher in SAV [5.57 mg L−1 (± 3.12; n = 12)] compared with giant salvinia [2.24 mg L−1 (± 2.59; n = 12)]. pH was 1.08 times higher in SAV [6.48 (± 0.32; n = 12)] relative to giant salvinia [6.97 (± 0.30; n = 12)]. Specific conductance (t-value = 2.73, P = 0.01), P-Ortho (t-value = − 5.62, P < 0.01), and NH4 (t-value = -4.51, P < 0.01) were elevated in giant salvinia, relative to SAV. Specific conductivity was 1.56 times higher in giant salvinia [1,493 µS (± 1,445; n = 12)] relative to SAV [955 µS (± 597; n = 12)]. Phosphate was 4.84 times higher in giant salvinia [0.28 mg L−1 (± 0.34; n = 12)] compared to SAV [0.06 mg L−1 (± 0.04; n = 12)], and NH4 was 3.31 times higher in giant salvinia [0.12 mg L−1 (± 0.10; n = 12)] relative to SAV [0.04 mg L−1 (± 0.03; n = 12)]. Dry biomass was 2.98 times higher in giant salvinia [31.70 g (± 10.36; n = 36)] compared with SAV [10.65 g (± 6.39; n = 36); t-value = -9.42, P < 0.01]. Mean submerged root length was 0.13 m (± 0.04; n = 120) and mean mat thickness was 0.6 m (± 0.02; n = 12), neither significantly varied among sampling months.
In the SAV, light availability declined gradually with light reaching the bottom of the waterbody (Table 2). However, when giant salvinia was present, light availability steeply declined and was 7.77 times lower immediately below the water surface in giant salvinia [4,664 lux (± 7,989; n = 12)], compared to SAV [36,250 lux (± 18,773; n = 12; t-value = 41.68, P < 0.01)].
Macroinvertebrate community
A total of 22,812 macroinvertebrate specimens (12,412 from SAV and 10,400 from giant salvinia), in 50 lowest practical taxonomic units (LPTs) and 27 families within 12 orders, were identified (Supplementary Material Table S1). Fifteen of 50 LPTs occurred less than four times, thus were removed prior to analysis. Standardized macroinvertebrate abundance per gram was 3.23 times greater in SAV [34.35 (± 22.61)] compared with in giant salvinia [10.64 (± 7.57); t-value = 2.68, P = 0.01], and richness per gram was five times greater in SAV [1.50 (± 0.96)] than giant salvinia [0.30 (± 0.14); t-value = 14.29, P < 0.01].
During initial sampling in August, macroinvertebrate communities in SAV were Chironomidae dominant and Chironomidae and Hyalella sp. were co-dominant in giant salvinia (Fig. 2), subsequently though, communities in giant salvinia became dominated by amphipods, specifically Hyalella sp. In giant salvinia, Chironomidae abundance decreased each month from a mean of 5.92 per gram [± 2.92 (± SD)] in August to 0.5 per gram (± 0.96) in May. Aside from these two groups, all other LPTs decreased from 24% of the giant salvinia community in August to 4% in December then < 9% in February and May (Fig. 2). In SAV, the macroinvertebrate community was composed of Chironomidae (42%), Hyalella (34%), and all other LPTs represented 24% of LPTs throughout the study.

Mean abundance of macroinvertebrate taxa per gram of dry mass (± SE) in giant salvinia and submerged aquatic vegetation (SAV) habitat over four sampling months. Only taxa with ≥ 5% relative abundance were labeled with percent, taxa under this threshold were grouped as all others
Habitat, month, and interaction between the two factors described macroinvertebrate LPT abundance. Significant differences in abundance were most explained by habitat (14 of 36 LPTs; Deviance = 370.6, P < 0.01) followed by month (10 of 36 LPTs; Deviance = 521.5, P < 0.01) and interaction (5 of 36 LPTs; Deviance = 223.1, P < 0.01; Table 3; Supplementary Material Fig. S1). Of the 14 LPT differing between habitat, nine were more abundant in SAV.
Macroinvertebrate community structure between habitats was different. Ordination of macroinvertebrate community composition found distinct groups between habitat types (Fig. 3 and Table 4). Residual STRESS was 0.126, which is within ranges for interpretability (Clarke 1993). The first axis presumably explained community differences based on habitat type. The second axis presumably related to temporal changes within giant salvinia. Habitat (ADONIS F = 4.86, R2 = 0.061, P < 0.01) and month (ADONIS F = 2.58, R2 = 0.097, P = 0.01) explained variation in the macroinvertebrate communities. The dispersion test showed that giant salvinia (0.13) and SAV (0.12) were similar in multivariate distance, thus, the analysis was appropriate for these data (Anderson & Walsh, 2013). Habitat association analyses identified Gammarus sp., Collembola, Scirtes sp., and Dolichopodidae as most related to giant salvinia ordination group, while Neocorixa sp., Hydrachnidia, Parapoynx sp., and Oxyethira sp. as most related to SAV ordination group (Table 4). Similarity percentages (SIMPER) identified 61.4% dissimilarity in community composition between habitat, with Chironomidae and Hyalella sp. as contributing the most to community turnover between habitat type.

Nonmetric multidimensional scaling (NMDS) ordination plot of macroinvertebrate communities in giant salvinia and submerged aquatic vegetation (SAV) habitat. Points are individual samples and distance between points represents assemblage similarity, with most similar samples being located closest together. Color depicts habitat types, giant salvinia (black) and submerged aquatic vegetation [SAV; (gray)], and shapes identify sampling month, August (circle), December (square), February (triangle), and May (diamond)
Previous analyses indicated dry biomass, DO, specific conductance, pH, NO3−, P-ortho, NH4, and light availability at all depths significantly varied between habitat and, thus, were included in the RDA. We selected only one depth (0.01 m) of light availability for the analysis. The first two axes of the ordination explained 86% of the variation (68% in 1st axis). Monte Carlo permutation test indicated that axis one (F = 42.01, P < 0.01) and axis two (F = 11.43, P < 0.01) were significant. The first axis was positively correlated with Chironomidae-dominant communities (0.727), as well as, pH (0.716), light availability (0.608), and specific conductance (0.519), and negatively correlated with amphipod communities (− 0.805) and dry biomass (− 0.659). The second axis was positively correlated with dry biomass (0.534), and was negatively correlated with DO (− 0.828), pH (− 0.262) and light availability (− 0.386). A biplot of the first two axes showed that increased dry biomass was associated with giant salvinia habitat, while pH and light availability increased with SAV habitat (Fig. 4).

Redundancy analysis (RDA) biplot of macroinvertebrate communities and environmental variables sampled. Points represent sampling sites and arrows represent environmental gradients. Color depicts habitat types, giant salvinia (black) and submerged aquatic vegetation [SAV; (gray)], and shapes identify sampling month, August (circle), December (square), February (triangle), and May (diamond). Environmental variables include dry biomass, dissolved oxygen (DO), pH, specific conductance (conductivity), nitrate (NO3), ammonium (NH4), orthophosphate (P-ortho), and light availability at 0.01 m depth (below)
Caloric value
Based on eighteen taxa (96% relative abundance; Supplementary Material Table S1) with previously published calorific values, mean caloric value [kcal g−1 per gram dry biomass (± SD)] in SAV was 3.39 times higher [146 (± 92.2); t-value = − 2.31, P = 0.02] than giant salvinia [43.1 (± 31.2)]. In giant salvinia, mean caloric value ranged from 25.6 to 58 kcal g−1 across months, with only May being significantly lower (t = value = 2.2, P = 0.03). Mean values were 53.8 (± 17.9) and 58 (± 43.6) kcal g−1 for August and December, respectively, and 35.2 (± 21.8) kcal g−1 in February, then decreased to 25.6 (± 20.9) kcal g−1 in May. In SAV, mean caloric value across months ranged from 103 to 220 kcal g−1, with February being significantly higher (t-value = − 2.2, P = 0.03). Caloric value was 119 (± 69.3) kcal g−1 in August then 103 (± 81.3) December. Caloric value increased to 220 (± 112) kcal g−1 in February then was 144 (± 42) kcal g−1 in May.
Discussion
Our study suggests that giant salvinia invasion altered habitat availability and environmental conditions, including light, DO, and nutrients, that probably impacted macroinvertebrates and resulted in different communities relative to SAV. Our hypothesis that giant salvinia having a diverse and abundant community was not supported by the data. As infestation duration increased, giant salvinia communities became species depauperate, and lower in biomass and energy. In SAV, Chironomidae was dominant and insects were abundant in the macroinvertebrate communities, which can be expected for freshwater marshes (Bolduc & Afton, 2003, 2004; Kang & King, 2013; Mercer et al., 2017; Weller & Bossart, 2017). In giant salvinia, conversely, crustaceans, specifically amphipods, dominated the community while insects were less abundant. This could be explained by environmental filtering and habitat changes resulting from giant salvinia invasion.
Environmental conditions and presumably environmental filtering differed between SAV and giant salvinia habitats. Decreased light penetration may reduce periphyton, phytoplankton, and zooplankton abundance, which, although unmeasured, limiting feeding stratagies and functional traits of macroinvertebrates that could exist in the giant salvinia habitat (Pinto & O’Farrell, 2014). Three of the four taxa identified as positively relating to SAV were an air breather (Neocorixa sp.) and two herbivores (Parapoynx sp., and Oxyethira sp.) (Poff et al., 2016). Giant salvinia may have limited access to the surface for air breathing insects. Although external gill breathing macroinvertebrates are typically more sensitive to low DO than air breathing macroinvertebrates (Kaller & Kelso, 2007), a thin layer of higher DO exists at the giant salvinia mat and water surface interface (Frodge et al., 1990; Wahl et al., 2020), which may have permitted external gill breathing macroinvertebrates to exist within giant salvinia. In August, gill breathing macroinvertebrates were present when recorded DO was < 1 mg L−1, suggesting that taxa may have persisted by a layer of higher DO water near the water–giant salvinia interface, or these macroinvertebrates may exhibit tolerance to low DO (Justus et al. 2012; Kaller & Kelso, 2007). Differences in macroinvertebrates between other invasive free-floating with submerged macrophyte species have been observed in other systems. An increase in Lemna minuta Kunth mat thickness was negatively correlated with the number of aquatic plant species and plant coverage, as well as, a 40% decrease in macroinvertebrate richness (Ceschin et al., 2020). Even though L. minuta is a thinner matted species, the effects to macroinvertebrates, and the resulting shift in communities, were similar to the effect we observed from giant salvinia.
Giant salvinia habitat also may not offer the spatial extent and diversity of habitat as SAV, with its more complex architecture (Fisher & Kelso, 2007; Thomaz et al., 2008). The giant salvinia mat was limited to submerged and emergent fronds, and suspended giant salvinia detritus, and presented a homogenous habitat over the infested area. Conversely, SAV can span the entire water column, offering increased surface area for habitat and predator avoidance (Fisher & Kelso, 2007), and increased habitat heterogeneity and species richness (Thomaz et al., 2008). Loss of habitat niches and spatial extent likely contributed to some taxonomic differences, possibly through increased competition (Calizza et al., 2017). Variation of macroinvertebrate community structure within SAV habitat over the study duration suggests high seasonal variation, therefore, differences in communities from giant salvinia may be explained by habitat alterations and by seasonality. We believe the divergence of macroinvertebrate community structure between macrophytes is consistent with the effect of macrophyte habitat change (Schultz & Dibble, 2012; Habib & Yousuf, 2015). However, we cannot differentiate between variation caused by habitat changes and variation contributed by seasonality, we acknowledge that it is difficult to ascertain the precise effect of giant salvinia on macroinvertebrate communities from our dataset.
It was not possible to determine if macroinvertebrate communities in giant salvinia and SAV locations were similar prior to initial sampling. Following initial invasion, communities between habitat types contained similar taxa, however, changes in communities were evident (Figs. 2 and 3), indicating that giant salvinia may have started to impact macroinvertebrates prior to our sampling or initial communities were different, although the RDA results (Fig. 4) suggest that communities in giant salvinia during August were similar to SAV communities, providing some evidence of a common starting taxonomic pool. Thus, lacking data on the macroinvertebrate communities of these freshwater marshes prior to giant salvinia invasion, we cannot determine the magnitude of change, specific taxon loss, or attribute some changes to inherent natural spatial and temporal variation as would have been possible with such data, and unfortunately, data from the literature describing communities prior to giant salvinia are not directly comparable (i.e., different taxonomic classifications; Bolduc & Afton, 2003, 2004). The inability to determine pre-invaded community structure and decipher seasonal variation in giant salvinia locations is inherent to the design of our study. More frequent sampling before and during invasion could make it possible to precisely determine the effect of giant salvinia on macroinvertebrate communities.
The growing giant salvinia mat likely created internal nutrient loading that further reinforced the invasion through nutrient release from the sediment. Reduction in DO following giant salvinia invasion has been well documented (Oliver, 1993; Flores & Carlson, 2006; Tipping et al., 2008) and can cause phosphorus and ammonium to release into the water column from the sediment (Søndergaard 2003; Zhang et al., 2014), creating more favorable conditions to sustain the invasion (Strange et al., 2019). Pontederia crassipes (Mart.) Solms tissue decomposition released phosphorus into the water column, with the concentration of release increasing when DO was < 1 mg L−1 (Masifwa et al., 2004); and in this study, locations with giant salvinia in August had a mean DO concentration < 1 mg L−1 and the highest phosphorus and ammonium concentrations. Nutrient availability is a limiting factor for giant salvinia, but internal loading, caused by a reduction in DO and increase in nutrients can reinforce giant salvinia invasion. Human intervention through chemical, mechanical, or biological control is typically required to shift back to SAV dominance (Tipping et al., 2008; Motitsoe et al., 2020). This shift back, however, is typically temporary with giant salvinia recolonizing within months after control (Oliver, 1993). Peeters (2016) found invasive free-floating plants had a higher probability of recolonizing a location which was previously controlled because of lingering phosphorus concentrations. Therefore, although anthropogenic control restored SAV dominance at this site, the high likelihood of giant salvinia reestablishment in systems like these with altered nutrients could have long-term impacts to local macroinvertebrate richness and diversity.
Reoccurring invasion at a location could act as a sink for a number of macroinvertebrate taxa. Heterogenous SAV-dominant habitat allows a greater richness of macroinvertebrates to exist (Thomaz & Cunha, 2010; Lucena-Moya & Duggan, 2011), and macroinvertebrates with adult flying stage will disperse from this habitat to colonize other locations (Wissinger, 1999). If they colonize an area frequently covered with giant salvinia during a time with little or low giant salvinia coverage, they may not be able to complete their life cycle before the environment shifts to dominance by giant salvinia. If they cannot develop before invasion, they would be subject to changes in environmental filtering and increased competition. Additionally, the mat of vegetation on the water surface may limit adult aquatic insect emergence or egg laying. If adult emergence is impacted, energy transfer from the aquatic to the terrestrial ecosystem, in the form of predation on emerging insect, would be diminished.
The flow of energy though the aquatic and adjacent terrestrial ecosystems could be altered by giant salvinia. Previous research has identified giant salvinia energy flow alterations related to decreased light penetration (Rommens et al., 2003; McFarland et al., 2004). Our results indicate a reduction in caloric value in the giant salvinia macroinvertebrate community, which could have bottom-up ramifications for the trophic web. Fish and other aquatic predators surviving in the low DO environment under giant salvinia would have lower quality and lower diversity of prey items. Toft et al. (2003) found that invertebrate communities and fish diets were altered by the introduction of P. crassipes, shifting from the highly abundant native amphipod, Hyalella azteca Saussure, as fish prey to the amphipod, Crangonyx floridanus Bousfield, that was not consumed by fish. Chironomidae are a common forage item, especially for larval, juvenile, and smaller-bodied fish (Armitage 1995). Therefore, a shift away from Chironomidae dominant communities may hinder trophic interaction and energy transfer. Even when invasive plants are controlled, it may take weeks or years for macroinvertebrate communities to resemble pre-infested communities (Wallace, 1990). Waterfowl, especially diving ducks, rely on macroinvertebrates during migration and in wintering habitat (Afton et al., 1991), often selecting habitat abundant with macroinvertebrates (Schummer et al., 2008). Similarly, many wading birds are indirectly dependent on macroinvertebrates, through fish (Gawlik, 2002). The reduction in total energy of the macroinvertebrate community could result in altered habitat use and decreased populations of waterfowl and wading birds. A collapse of overwintering diving duck populations is suspected to be caused, in part, by a decrease in macroinvertebrate abundance and total biomass (Tománková et al., 2013). Several species of wading birds have been shown to utilize habitat based on prey, fish availability, and decreased fish abundance, through loss of total energy from macroinvertebrates, could lead to reduced wading bird abundance (Gawlik, 2002).
Conclusion
This study indicates that the invasion of giant salvinia reduced habitat availability and complexity through changes in environmental conditions, and contributed to shifts in macroinvertebrate communities. Due to its widespread distribution and now documented reduction of biodiversity and energy transfer, invasion of giant salvinia could have larger implications for freshwater ecosystems than previously understood. Research is needed to understand how giant salvinia alters the flow of energy in aquatic ecosystems, specifically how adjacent trophic levels are impacted by alterations to their prey community and the timeframes for macroinvertebrate communities to recover.
Data availability
Data will be available on the Dryad Digital Repository.
References
Afton, A. D., R. H. Hier & L. Paulus, 1991. Lesser Scaup diets during migration and winter in the Mississippi flyway. Canadian Journal of Zoology 69: 328–333.
Anderson, M. J. & D. C. I. Walsh, 2013. PERMANOVA, ANOSIM, and the Mantel Test in the face of heterogenous dispersions: What null hypothesis are you testing. Ecological Monographs 83: 557–574.
Arar, E. J. & G. B. Collins, 1997. Method 445.0 In vitro determination of chlorophyll a and pheophytin in marine and freshwater algae by fluorescence. U.S. Environmental Protection Agency, Washington, DC.
Armitage, P. D., 1995. Chironomidae as food. In: Armitage, P. D., P. S Cranston, L.C.V. Pinder, (eds), The chironomidae: biology and ecology of non-biting midges. Springer, Dodrect: 423–435.
Attermeyer, K., S. Flury, R. Jayakumar, P. Fiener, K. Steger, V. Arya, F. Wilken, R. van Geldern & K. Premke, 2016. Invasive floating macrophytes reduce greenhouse gas emissions from a small tropical lake. Scientific Reports 6: 20424.
Bolduc, F. & A. D. Afton, 2004. Relationships between wintering waterbirds and invertebrates, sediments and hydrology of coastal marsh ponds. Waterbirds 27: 333–341.
Bolduc, F. & A. D. Afton, 2003. Effects of structural marsh management and salinity on invertebrate prey of waterbirds in marsh ponds during winter on the Gulf Coast Chenier Plain. Wetlands 23: 897–910.
Boucek, R. E. & J. S. Rehage, 2015. A tale of two fishes: Using recreational records to examine the link between floodplain connections in a subtropical coastal river. Estuaries and Coasts 38: 124–135.
CABI, 2019. Salvinia molesta. [original text by Parys K and Mikulyuk A]. In: Invasive Species Compendium. CAB International, Wallingford, UK.
Calizza, E., M. L. Costantini, G. Careddu & L. Rossi, 2017. Effect of habitat degradation on competition, carrying capacity, and species assemblage stability. Ecology and Evolution 7: 5784–5796.
Caraco, N., J. Cole, S. Findlay & C. Wigand, 2006. Vascular plants as engineers of oxygen in aquatic systems. Bioscience 56: 219–225.
Cary, P. R. & P. G. J. Weerts, 1983. Growth of Salvinia molesta as affected by water temperature and nutrition I. Effects of nitrogen level and nitrogen compounds. Aquatic Botany 16: 163–172.
Ceschin, S., G. Ferrante, F. Mariani, L. Traversetti & N. T. W. Ellwood, 2020. Habitat change and alteration of plant and invertebrate communities in waterbodies dominated by the invasive alien macrophyte Lemna minuta Kunth. Biological Invasions 22: 1325–1337.
Chang, J., 2015. lda: Collapsed gibbs sampling methods for topic models. R package version 1(4): 2.
Cheruvelil, K. S., P. A. Soranno, J. D. Madsen & M. J. Roberson, 2002. Plant architecture and epiphytic macroinvertebrate communities: The role of an exotic dissected macrophyte. Journal of North American Benthological Society 21: 261–277.
Clarke, K. R., 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18: 117–143.
Colon-Gaud, J. C. & W. E. Kelso, 2003. A suitcase trap for sampling macroinvertebrates in dense submerged aquatic vegetation. Journal of the Kansas Entomological Society 76: 667–671.
Colon-Gaud, J. C., W. E. Kelso & D. A. Rutherford, 2004. Spatial distribution of macroinvertebrates inhabiting Hydrilla and Coontail Beds in Atchafalaya Basin, Louisiana. Journal of Aquatic Plant Management 42: 85–91.
Covich, A. P., M. A. Palmer & T. A. Crowl, 1999. The role of benthic invertebrate species in freshwater ecosystems: Zoobenthic species influence energy flows and nutrient cycling. Bioscience 49: 119–127.
da Silva, C. V. & R. Henry, 2020. Aquatic macroinvertebrate assemblages associated with two floating macrophyte species of contrasting root systems in a tropical wetland. Limnology 21: 107–118.
Engle, V. D., 2011. Estimating the provision of ecosystem services by Gulf of Mexico coastal wetlands. Wetlands 31: 179–193.
Ferreira, T. F., L. O. Crossetti, D. M. L. Motta Marques, L. Cardoso, C. R. Fragoso Jr. & E. H. van Nes, 2018. The structuring role of submerged macrophytes in a large subtropical shallow lake: Clear effects on water chemistry and phytoplankton structure community along a vegetated-pelagic gradient. Limnologica 69: 142–154.
Finlayson, C. M., 2016. Freshwater marshes and swamps. In Finlayson, C., G. Milton, R. Prentice & N. Davidson (eds), The Wetland Book. Springer, Dordrecht.
Fisher, J. C. & W. E. Kelso, 2007. Effectiveness of artificial plants in subsurface exclosures as a substrate for Hydrilla-dwelling macroinvertebrate communities. Journal of Freshwater Ecology 22: 33–39.
Fisher, J. C., W. E. Kelso & D. A. Rutherford, 2012. Macrophyte mediated predation on hydrilla-dwelling macrophytes. Fundamental and Applied Limnology 181: 25–38.
Flores, D. & J. W. Carlson, 2006. Biological control of giant salvinia in East Texas waterways and the impact on dissolved oxygen levels. Journal of Aquatic Plant Management 44: 115–121.
Foley, C. C., 2015. Wading bird food availability in rice fields and crawfish ponds of the Chenier plain of southwest Louisiana and southeast Texas. LSU Master’s Thesis 19.
Fredrickson, L. H. & F. A. Reid, 1988. Waterfowl management handbook: Nutritional values of waterfowl foods. Fish and Wildlife Leaflet 13. United States Department of Interior, Fish and Wildlife Service, Washington, DC.
Frodge, J. D., G. L. Thomas & G. B. Pauley, 1990. Effects of canopy formation by floating and submergent aquatic macrophytes on the water quality of two shallow Pacific Northwest lakes. Aquatic Botany 38: 231–248.
Gawlik, D. E., 2002. The effects of prey availability on the numerical response of wading birds. Ecological Monographs 72: 329–346.
Hill, M. P., J. A. Coetzee, G. D. Martin, R. Smith & E. F. Strange, 2020. Invasive alien aquatic plants in South African freshwater ecosystems. In van Wilgen, B. W., J. Measey, D. M. Richardson, J. R. Wilson & T. A. Zengeya (eds), Biological invasions in South Africa. Springer, Cham: 97–114.
Hinson, A. L., R. A. Feagin, M. Eriksson, R. G. Najjar, M. Herrmann, T. S. Bianchi, M. Kemp, J. A. Hutchings, S. Crooks & T. Boutton, 2017. The spatial distribution of soil organic carbon in tidal wetland soils of the on the continental United States. Global Change Biology 23: 5468–5480.
Jones, J. I. & C. D. Sayer, 2003. Does the fish-invertebrate-periphyton cascade precipitate plant loss in shallow lakes? Ecology 84: 2155–2167.
Justus, B. G., S. V. Mize & D. Kroes, 2012. Invertebrate and fish assemblage relations to dissolved oxygen minima in lowland streams of southwestern Louisiana. River Research and Applications 30: 11–28.
Kaller, M. D. & W. E. Kelso, 2007. Association of macroinvertebrate assemblages with dissolved oxygen concentration and wood surface area in selected subtropical streams of the southeastern USA. Aquatic Ecology 41: 95–110.
Kaller, M. D., W. E. Kelso & J. C. Trexler, 2013. Wetland fish monitoring and assessment. In Anderson, J. & C. Davis (eds), Wetland Techniques. Springer, Dordrecht: 197–265.
Kang, S. R. & S. L. King, 2013. Effects of hydrologic connectivity on aquatic macroinvertebrate assemblages in different marsh types. Aquatic Biology 18: 149–160.
Kantrud, H. A., G. L. Krapu & G. A. Swanson, 1989. Prairie basin wetlands of North Dakota: A community profile. Biological report 85(7.28). U.S. Department of the Interior, Fish and Wildlife Service, Research and Development, Washington, DC.
Lehner, B. & P. Döll, 2004. Development and validation of a global database of lakes, reservoirs and wetlands. Journal of Hydrology 296: 1–22.
Lucena-Moya, P. & I. C. Duggan, 2011. Macrophyte architecture affects the abundance and diversity of littoral microfauna. Aquatic Ecology 45: 279–287.
Luque, G. M., C. Bellard, C. Bertelsmeier, E. Bonnaud, P. Genovesi, D. Simberloff & F. Courchamp, 2014. The 100th of the world’s worst invasive alien species. Biological Invasion 16: 981–985.
Masifwa, W. F., W. Okello, H. Ochieng & E. Ganda, 2004. Phosphorus release from decomposing water hyacinth and effects of decomposition on water quality. Uganda Journal of Agricultural Sciences 9: 389–395.
McFarland, D. G., L. S. Nelson, M. J. Grodwitz, R. M. Smart & C. S. Owens, 2004. Salvinia molesta D.S. Mitchell giant salvinia in the United States: A review of species ecology and approaches to management. U.S. Army Engineer Research and Development Center, ERDC/EL SR-04-2
Mercer, N., M. D. Kaller & M. J. Stout, 2017. Diversity of arthropods in farmed wetlands in the Gulf of Mexico Coastal Plain and effects of detrital subsidies. Journal of Freshwater Ecology 32: 163–178.
Mitchell, D. S. & N. M. Tur, 1975. The rate of growth of salvinia molesta (S. Auriculata Auct). in laboratory and natural conditions. Journal of Applied Ecology 12: 213–225.
Motitsoe, S. N., J. A. Coetzee, J. M. Hill & M. P. Hill, 2020. Biological control of Salvinia molesta DS Mitchell drives aquatic ecosystem recovery. Diversity 12: 204.
Oksanen, J., F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens, E. Szoecs & H. Wagner, 2019. Package ‘vegan,’.
Oliver, J. D., 1993. A review of the biology of giant salvinia Salvinia molesta Mitchell. Journal of Aquatic Plant Management 31: 227–231.
Parys, K. A., M. L. Gimmel & S. J. Johnson, 2013. Checklist of insects associated with Salvinia minima Baker in Louisiana, USA. Check List 9: 1488–1495.
Pattison-Williams, J. K., J. W. Pomeroy, P. Badiou & S. Gaborc, 2018. Wetlands, flood control and ecosystem services in the Smith Creek drainage basin: A case study in Saskatchewan, Canada. Ecological Economics 147: 36–47.
Peeters, E. T. H. M., 2016. Competition between free-floating plants strongly driven by previously experienced phosphorus concentrations in the water column. PLoS ONE 11:
Pinto, P. T. & I. O’Farrell, 2014. Regime shifts between free-floating plants and phytoplankton: A review. Hydrobiologia 740: 13–24.
Poff, N. L., J. D. Olden, N. K. M. Vieira, D. S. Finn, M. P. Simmons & B. C. Kondratieff, 2016. Functional trait niches of North American lotic insects: traits-based ecological applications in light of phylogenetic relationships. Journal of the North American Benthlogical Society 25: 730–755.
R Core Team, 2013. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rommens, W., J. Maes, N. Dekeza, P. Inghelbrecht, T. Nhiwatiwa, E. Holsters, F. Ollevier, B. Marshall & L. Brendonck, 2003. The impact of water hyacinth Eichhornia crassipes in a eutrophic subtropical impoundment Lake Chivero, Zimbabwe. I. Water quality. Fundamental and Applied Limnology 158: 373–388.
Schultz, R. & E. Dibble, 2012. Effects of invasive macrophytes on freshwater fish and macroinvertebrate communities: the role of invasive plant traits. Hydrobiologia 684: 1–14.
Schummer, M. L., S. A. Petrie & R. C. Bailey, 2008. Interaction between macroinvertebrate abundance and habitat use by diving ducks during winter on northeastern Lake Ontario. Journal of Great Lakes Research 34: 54–71.
Søndergaard, M., 2003. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 506: 135–145.
Strange, E. F., P. Landi, J. M. Hill & J. A. Coetzee, 2019. Modelling top-down and bottom-up drivers of a regime shift in invasive aquatic plant stable states. Frontiers in Plant Science 10: 889.
Thomaz, S. M., E. D. Dibble, L. R. Evangelista, J. Higuti & L. M. Bini, 2008. Influences of aquatic macrophyte habitat complexity on invertebrate abundance and richness in tropical lagoons. Freshwater Biology 53: 358–367.
Thomaz, S. M. & E. R. Cunha, 2010. The role of macrophytes in habitat structuring in aquatic ecosystems: methods of measurement, cause and consequences on animal assemblages’ composition and biodiversity. Acta Limnologica Brasiliensia 22: 218–236.
Tipping, P. W., M. R. Martin, T. D. Center & T. M. Davern, 2008. Suppression of Salvinia molesta Mitchell in Texas and Louisiana by Cyrtobagous salviniae Calder and Sands. Aquatic Botany 88: 196–202.
Toft, J. D., C. A. Simenstad, J. R. Cordell & L. F. Grimaldo, 2003. The effects of introduced water hyacinth on habitat structure, macroinvertebrate assemblages, and fish diets. Estuaries 26: 746–758.
Tománková, I., C. Harrod, A. D. Fox & N. Reid, 2013. Chlorophyll-a concentrations and macroinvertebrate declines coincide with the collapse of overwintering diving duck populations in a large eutrophic lake. Freshwater Biology 59: 249–256.
Turner, K., 1991. Economics and wetland management. Environmental. Economics 20: 59–63.
Vargas-Lopez, I. A., W. E. Kelso, C. P. Bonvillain, R. F. Keim & M. D. Kaller, 2020. Influence of water quality, local knowledge and river-floodplain connectivity on commercial wild crayfish harvesting in the Atchafalaya River Basin. Fisheries Management and Ecology 27: 417–428.
Vymazal, J., 2007. Removal of nutrients in various types of constructed wetlands. Science of the Total Environment 380: 48–65.
Wallace, J. B., 1990. Recovery of lotic macroinvertebrate communities from disturbance. Environmental Management 14: 605–620.
Welsh, B. L., 1975. The role of grass shrimp Palaemonetes pugio, in a tidal marsh ecosystem. Ecology 56: 513–530.
Wahl, C. F., R. Diaz & J. Ortiz-Zayas, 2020. Assessing Salvinia molesta impact on environmental conditions in an urban lake: case study of Lago Las Curias, Puerto Rico. Journal of Aquatic Invasions 15.
Wang, Z. & S. Yan, 2017. Impact of water hyacinth on aquatic environment in phytoremediation of eutrophic lakes. In Yan, S. & J. Guo (eds), water hyacinth. Press-Taylor and Francis, London: 204–250.
Warton, D. I., S. T. Wright & Y. Wang, 2012. Distance-based multivariate analyses confound location and dispersion effects. Methods in Ecology and Evolution 3: 89–101.
Weller, M. O. & J. L. Bossart, 2017. Insect community diversity tracks degradation and recovery of a waste water assimilation marsh in southeast Louisiana. Wetlands 37: 661–673.
Wilson, B. C., C. A. Manlove & C. G. Esslinger, 2002. North American Waterfowl Management Plan, Gulf Coast Joint Venture: Mississippi River Coastal Wetlands Initiative. North American Waterfowl Management Plan, Albuquerque, New Mexico.
Wissinger, S. A., 1999. Ecology of wetland invertebrates: synthesis and application for conservation and management. In Batzer, D. P., R. B. Rader & S. A. Wissinger (eds), Invertebrates in freshwater wetlands of North America: Ecology and management. Wiley, Hoboken: 1043–1086.
Zhang, L., S. Wang & Z. Wu, 2014. Coupling effect of pH and dissolved oxygen in water column on nitrogen release at water–sediment interface of Erhai Lake, China. Estuarine, Coastal and Shelf Science 149: 178–186.
Acknowledgements
We would like to thank the following people for field and laboratory assistance, Louisiana State University AgCenter employees, William Kelso, Debra Kelly, Fatima Nguyen, Keyla Pruett, Emily Passman; US Army Corps. Engineers employee, Christopher Mudge; and Miami Corporation employees, Darren Richard, Chad Courville. Thanks to Louisiana Department of Wildlife and Fisheries, Delta Waterfowl, and Mid-South Aquatic Plant Management Society for providing funds. This study was supported in part by the USDA National Institute of Food and Agriculture, Hatch project Accession Number 1013004 and McIntire-Stennis Cooperative Forestry Program as Project Number LAB-94335.
Funding
Department of Wildlife and Fisheries, Delta Waterfowl, and Mid-South Aquatic Plant Management Society provided partial funding. This study was supported in part by the USDA National Institute of Food and Agriculture, Hatch project Accession Number 1013004 and McIntire-Stennis Cooperative Forestry Program as Project Number LAB-94335.
Author information
Authors and Affiliations
Contributions
CFW, MK, and RD conceived the ideas and designed methodology; CFW collected the data; CFW and MK analyzed the data; CFW led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
There are no conflict of interest.
Additional information
Handling editor: Maria de los Angeles Gonzalez Sagrario
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Wahl, C., Kaller, M. & Diaz, R. Invasion of floating fern alters freshwater macroinvertebrate community structure with implications for bottom-up processes. Hydrobiologia 848, 2523–2537 (2021). https://doi.org/10.1007/s10750-021-04571-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-021-04571-4