
Abstract
Acoustic deterrent devices (pingers) that are designed to alert marine mammals to the presence of fishing gear have been successfully employed to reduce cetacean–fishery interactions. However, pinger sounds may affect fisheries target species: noise can induce short-term stress responses in fish that are reflected in increased blood cortisol concentrations and behavioural alterations, which may ultimately result in reduced catch rates. In order to test this hypothesis, the present study analyses the hormonal and behavioural stress response of European sardine (Sardina pilchardus) to a commercially available pinger—the 70 kHz dolphin pinger. The responses of wild captive fish to the pinger sounds were tested in tank experiments, measuring the stress level of fish by analysing blood plasma cortisol concentration and swimming behaviour. Mixed effect models were used for statistical analysis. During the experiments, pinger sounds caused subtle, but significant elevations in sardine plasma cortisol concentration and fish school compaction. However, when compared to the rates of increase reported for acute stress responses in similar studies, the observed increment was very small. This suggests that the sounds of the trialled pinger are not likely to significantly alter sardine behaviour and should consequently not have any negative impact on sardine catch rates.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pingers are acoustic deterrent devices (ADDs) that are designed to deter marine mammals from fishing gear by emitting aversive low intensity (<150 dB re 1 mPa @ 1 m), high-frequency (3–500 kHz) sounds in the hearing range of the animals (Reeves et al., 1996; Dawson et al., 2013). They are primarily employed as a bycatch reduction measure (Kraus et al., 1997; Trippel et al., 1999; Carlström et al., 2009; Gönener & Bilgin, 2009), but can also be effective in reducing marine mammal depredation on catch (Brotons et al., 2008; Gazo et al., 2008; Buscaino et al., 2009).
In Iberian Atlantic waters, interactions with cetaceans are particularly problematic for purse-seine, trawl and coastal gillnet fisheries (López et al., 2003; Vingada et al., 2011; Goetz et al., 2014). Bycatch rates of short-beaked common dolphin Delphinus delphis (Linnaeus, 1758), bottlenose dolphin Tursiops truncatus (Montagu, 1821) and harbour porpoise Phocoena phocoena (Linnaeus, 1758) (Aguilar, 1997; López et al., 2003; Goetz et al., 2014) are likely to be unsustainable in this area (López et al., 2003; Read et al., 2012; Lopez et al., 2013). The presence of common dolphins during purse-seining operations may cause the scattering of fish schools and consequently reduction of catch, the associated economic loss potentially being high. With the aim to reduce cetacean–fishery interactions in EU waters, pinger use became obligatory under the Council Regulation (EC) 812/2004Footnote 1 in specific fishing areas (including Atlantic waters of Spain and Portugal) for fisheries operating bottom-set gillnets, entangling nets and driftnets since 2004. There are many models of pingers available on the international market (for a review see Dawson et al., 2013). The Council Regulation defines technical specifications of pingers to be used in EU Community Fisheries: they should have a source level of 130–150 dB re 1 μPa @ 1 m, a fundamental frequency of 10–160 kHz with high-frequency harmonics, a pulse duration of 300 ms and an interpulse interval of 4–30 s. Pingers have successfully been employed to discourage common dolphins (Barlow & Cameron, 2003; Carretta & Barlow, 2011) from approaching static fishing gear in previous studies. The 70 kHz dolphin pinger (Future Oceans, formerly Fumunda Marine), one of the commercially available pinger model that meets the technical requirements specified under the Council Regulation, emits sounds within the known hearing range of common dolphins (see audiogram by Popov & Klishin, 1998: U-shaped audiogram with bandwidth up to 128 kHz at a level of 100 dB above the minimum threshold; minimum thresholds observed at frequencies of 60–70 kHz). This pinger model has been recently tested during field trials in Portuguese purse-seines fisheries, the pingers being attached with 100 m spacing on the float line of the purse-seine. The results showed significantly less interaction events during gear setting and hauling, bycatch of common dolphins being reduced from 0.045 to 0.009 bycaught animals per fishing event (Vingada et al., 2011). However, if the large-scale use of these pingers is considered as a potential management scenario, it is essential to rule out any negative effects of the devices on catch performance to ensure their acceptance by fishers (Gazo et al., 2008).
The European sardine, Sardina pilchardus, hereafter referred as sardine, is one of the main target species of purse-seine, artisanal driftnet and beach-seine fisheries off the NW Iberian Peninsula (Galician Ministry of Fisheries, 2013; Portuguese Directorate General of Natural Resources, Security and Maritime Services, 2013) and of great socio-economical importance for the local fishing communities and industries (Abaunza et al., 1995; Borges et al., 2003). However, no study has been conducted to date to assess the effect of pingers on the physiology and behaviour of sardines.
There have been a few field trials in the past to assess the effect of acoustic alarms on other fisheries target species, such as Atlantic herring Clupea harengus (Linnaeus, 1758), cod (Gadus morhua) and saithe Pollachius virens (Linnaeus, 1758) (Trippel et al., 1999; Culik et al., 2001; Cox et al., 2003; Buscaino et al., 2009; Gönener & Özdemir, 2012). In most cases no significant effects on catch rates were detected, although Kraus et al. (1997) reported lower catch rates of Atlantic herring in nets equipped with pingers compared to control nets and suggested that the herring possibly reacted to the pinger sounds by avoiding the nets. Culik et al. (2001), in contrast, found that Atlantic herring was attracted to nets equipped with low-frequency (2.7 kHz) pingers, resulting in higher catch rates. However, there are several factors in field trials, such as temporal fluctuations in local fish abundance, and changing oceanographic conditions, that may have a much bigger influence on catch rates than pinger sounds and make it difficult to isolate the effect of the pingers. Tank experiments can control for such factors. Kastelein et al. (2007) conducted tank trials with seven commercially available pinger models and found that some species such as Atlantic herring, European seabass Dicentrarchus labrax (Linnaeus, 1758) and thicklip grey mullet Chelon labrosus (Risso, 1827) showed aversive behaviour while being exposed to four of the tested pinger models.
Most fish species can detect sounds between 50 Hz to approximately 1.5 kHz but there are hearing specialists, such as some species within the taxonomic order of the clupeiforms, that are able to perceive sounds of up to 5 kHz (e.g. sea herrings, sprats, sardines, pilchards) or even in the ultrasonic range >20 kHz (e.g. shads Alosa spp) (Mann et al., 2001; Popper & Schilt, 2008). These high-frequency hearing abilities are thought to be evolutionary adaptations to predation from echolocating cetaceans (Mann et al., 1997) that may have been developed particularly by shallow-water fish species (Popper et al., 2004). Sardine is an important prey species of cetaceans in coastal Iberian Atlantic waters (Silva, 1999; Pusineri et al., 2007; Santos et al., 2007; Sollmann, 2011; Fernández et al., 2012; Read et al., 2012; Santos et al., 2013). Although there is no exact audiogram of European sardine available and currently no evidence for ultrasonic hearing in this species, it is possible that sardines may have developed such hearing specializations in response to echolocation clicks of preying cetaceans in the past. Consequently, sardines should also show avoidance reactions to commercially available pingers operating in the ultrasonic range, such as the 70 kHz dolphin pinger.
Noise exposure can increase stress levels in fish being reflected in acute physiological and behavioural responses (Popper & Hastings, 2009). These responses enable the animal to compensate or adapt to a disturbance and to overcome threats, such as predation (Barton & Iwama, 1991). Physiological stress responses in fish are expressed in immediate primary hormonal responses such as the release of corticosteroids (e.g. cortisol) and catecholamines into circulation, which give rise to secondary reactions including changes in plasma and tissue ion and metabolite levels, haematological features and heat shock or stress proteins. This can finally lead to tertiary responses such as changes in growth, condition, disease resistance, reproduction and ultimate survival (Barton, 2002). As in most fish cortisol reaches highest concentration one hour after being stressed (Iwama et al., 2006), cortisol tests are a good option in acute stress experiments (Porchas et al., 2009). Increased plasma cortisol concentrations in response to sounds have been observed in fish by Wysocki et al. (2006). Behavioural responses of fish to sounds are often expressed through changes in swimming behaviour, including fish school compaction, sinking in the water column, increase in swimming speed and aversion of the sound source (Schwarz & Greer, 1984; Misund et al., 1996; Suuronen et al., 1997; Wilson & Dill, 2002; Kastelein et al., 2007). Fish with high-frequency hearing capacity, such as the American shad Alosa sapidissima (Wilson, 1811), showed a very rapid and directional response directly away from the sound source when exposed to simulated dolphin echolocation clicks (Mann et al., 1998; Plachta & Popper, 2003). Changes in fish school dynamics (e.g. schools becoming more compact or changing their relative position in the water column) are an adaptive feature for the avoidance of predators (Pitcher et al., 1996), such as cetaceans. While fish school compaction may increase catch rates, all other aversive reactions may potentially reduce fish catchability and consequently catch rates in purse-seine and driftnet fisheries.
In order to assess whether the sounds of the 70 kHz dolphin pinger cause stress in sardines which may ultimately lead to reduced catch rates in fisheries directed at this species, the aim of the present study was to quantify the hormonal stress response of captive wild sardines to the sounds of this pinger by means of plasma cortisol analysis. The behavioural responses of the fish were also recorded, although results may be more difficult to interpret, as these shoaling pelagic fish were studied in tanks, where space is limited. Our research hypotheses were as follows: if the fish are stressed by the 70 kHz dolphin pinger sounds, the blood cortisol concentration of fish exposed to active pingers should be significantly higher than that of fish exposed to placebo (i.e. non-functional) pingers and their swimming behaviour (school compaction, distance of fish school to bottom of the tank, swimming speed) should differ significantly between trials with active and placebo pingers.
Materials and methods
Behavioural and physiological stress responses of fish were assessed in two separate experiments. Fish behaviour was recorded with an underwater video camera. Plasma cortisol concentrations were derived from blood samples of experimental fish.
Live fish handling and all experiments in this study were carried out by qualified staff using a procedure in accordance with general guidelines for the ethical use of animals in research, current Spanish Regulations and respective institutional guidelines. This work required no particular permit and did not involve endangered or protected species.
Capture, transport and acclimatization of fish
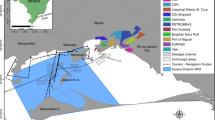
Live sardines were caught by a purse-seiner on the 11 May 2011 close to the Cies Islands (42°13′N/8°54′W) in the Ría de Vigo (Fig. 1). In order to maximize the post-capture survival rate of the fish, the captain of the vessel was thoroughly instructed about the correct catching and handling procedure and provided with all necessary materials beforehand. The catch comprised about 200 sardines that were transferred into a seawater tank (720 l) and provided with oxygen (>7 mg l−1) during the transport.

Map of the study area (Galicia, NW Spain). The fish symbol indicates the approximate location where live fish were caught
On arrival in the fishing harbour of Vigo, the transport tank was lifted onto a pick-up and brought immediately to the aquaculture facilities of ECIMAT (Estación de Ciencias Mariñas de Toralla, University of Vigo, Isla de Toralla, Vigo) (Fig. 1), where the fish were equally distributed into two cylindrical stocking tanks (\(\varnothing\) 1.36 m; 1,500 l; depth 1 m) filled with open-circuit pumped seawater (water temperature 15–17°C), supplied with air and covered with a shade net to avoid that fish jump out of the tank (Fig. 2).

Set-up of experimental tanks in the laboratory, including the square-shaped tank used for behavioural observations (on the left) and the two round stocking tanks (on the right) used for physiological experiments
Fish were acclimatized for 20 days in the stocking tanks before starting the experiments, since recovery from the acute stress of capture and transport can be expected within 2 weeks of acclimatization (Marçalo et al., 2008). The fish were kept at normal photoperiod and without any disturbance, except for water treatment, removal of dead animals and feeding twice a day. The tank water was initially treated with 200 ppm of an aqueous solution (40%) of methanal to eliminate external fish parasites and to improve the microbiological profile of the tank water and the fish. This treatment was repeated once a week. In addition, a solution of Oxitetracyclin (50 ppm), a broad-spectrum antibiotic, was added to the tank water once every day during the first week of acclimatization as a preventive treatment against possible bacterial infections (James et al., 1988). Fish adapted well to the tank conditions, started to swim in schools shortly after being introduced into the water tank and began feeding without any problems on day ten of the acclimatization period. They were initially fed with enriched Artemia spp (brine shrimp) and from day twelve on with pellets, starting with pellet size 150 μm until reaching pellet size 3.4 mm at day eighteen.
Set-up and preparation of tanks
Physiological and behavioural experiments were conducted in separate tanks, as displayed in Fig. 2. The feet of all tanks were placed on polystyrene plates (thickness: 4 cm) to buffer sound propagation from the environment to the tank. Physiological experiments were conducted in the stocking tanks.
For the behavioural observations a green, square-shaped tank (2 × 2 × 1 m; 4000 l), that allowed for suitable video images and provided enough space for the fish to school and swim in circles, was selected. A reference grid (370 square cells of 15 cm × 15 cm) for the distance measurements during the experiments was taped on the walls and bottom of the tank with white adhesive tape (1.9 cm wide) (Fig. 3).

Set-up of tank for the behavioural experiments. The positions of the underwater camera and the pinger/placebo in the tank are indicated by the arrows
Technical specifications of tested pinger model (70 kHz dolphin pinger, Future Oceans)
70 kHz dolphin pingers (Fig. 4) produce tonal signals with constant pulse duration of 300 ms and an interpulse interval of 4 s and operates in the ultrasonic range with a fundamental frequency of 70 kHz at a source level of 145 dB re 1 μPa @ 1 m. The sound frequency of the pinger may alter slightly (±10% of the nominal frequency), however, all emitted sounds are in the ultrasonic range (Pers. Comm., James Turner, owner and managing director of Future Oceans). Note that the same pinger was used throughout the experiments so the sound level should have been consistent.

The 70 kHz dolphin pinger (Future Oceans)
Sampling procedure
Physiological experiment
Plasma cortisol concentrations were derived from blood samples of experimental fish that were exposed to functional and placebo 70 kHz dolphin pingers. Blood cortisol sampling was carried out in the morning, always at the same hour, leaving at least two days between each experiment in order to give the fish time to return to normal conditions.
In order to determine a control baseline plasma cortisol value of sardines for each experimental day, five fish were caught from one of the experimental tanks (“control tank”), with a handnet (all at once in <30 s) and put into a bucket (20 l) with seawater and anaesthetic (300 ppm 2-Phenoxy-1-ethanol), and air supply. While the fish were sedated, the pinger (either placebo or active pinger) was slowly introduced into the other experimental tank (“test tank”) and suspended from a rope in the middle of the tank, at about half way up the water column. It started to ping when in contact with the salt water which is indicated by a small LED that flushes while the pinger is operating. The flashing LED was neutralized by covering it with a small piece of white tape. We checked whether the LED was flashing (by removing the tape) after each session when removing the pinger from the tank. After 1 h of exposure, five fish were retrieved from the test tank and sedated as described above. As most fish species show their highest plasma increase in cortisol within about 0.5–1 h after a stressful disturbance (Barton & Iwama, 1991), an acute hormonal response in sardine should therefore be detectable after 1 h of pinger exposure. Blood samples (1 ml) were taken from completely sedated fish (after 3 min) with heparinized syringes via caudal vein puncture (Fig. 5a). As manipulation of fish can provoke an immediate elevation of plasma cortisol that can be detected within 30 s of applying an acute stressor (Gerwick et al., 1999), fish capture (<30 s), sedation (3 min) and blood withdrawal (30–60 s/fish) were carried out as fast as possible. Afterwards, samples were centrifuged (10 min at 12,000 rpm) and plasma (transparent top layer) (Fig. 5b) was frozen at −80°C. Plasma cortisol was measured in defrosted serum samples by a Cortisol Enzyme Immunoassay Kit (Cortisol EIA Kit, Cayman Chemical Company, Ann Arbor, MI, USA), following the assay protocol included in the kit. The EIA plate was read with a microplate reader (Bio Rad 550) at a wavelength of 405 nm. Cortisol concentrations were converted from pg ml−1 into μg ml−1 for subsequent statistical analysis.

a Caudal vein puncture for sardine blood withdrawal and b plasma and red blood cell layers in centrifuged blood sample
The active pinger and the placebo were tested alternately during different days, resulting in four replicates each. The two experimental tanks were used alternately as “control” and “test” tanks during the course of the experiment.
Biological data of sardines
Immediately after blood withdrawal, sardines were killed, measured (total length in cm) and weighed (in grams). Fish condition, i.e. fatness of a fish in relation to its length, was calculated following the equation for Fulton’s condition factor K as
where W = whole body wet weight in g and L = total length in cm.
Fish with condition factor lower than the mean value (\(\bar{x}\) = 0.8072) were classified as “low-fat” and fish with condition factor higher than the mean as “high-fat” fish.
Gonads were extracted from the dead fish and frozen at −80°C for subsequent microscope and histological analysis. Sex (male/female) and maturity (immature/maturing/mature) were determined by visual observation (colour, texture and size) of fresh gonads and analysis of defrosted gonad tissue and histological samples under the fluorescence microscope (Nikon Eclipse 90i), following the criteria of Simón Díaz (2009). For histological analysis, pieces of gonad were taken from every sampled specimen, fixed in Davidson´s solution (Shaw & Battle, 1957) and embedded in paraffin. Paraffin blocks were sectioned at 5 µm with a microtome. Tissue sections were deparaffinized, stained with Harris´ hematoxylin (7,11b-Dihydro-6H-indeno[2,1-c]chromene-3,4,6a,9,10-pentol) and eosin (2-(2,4,5,7-tetrabromo-3-hydroxy-6-oxoxanthen-9-yl)benzoic acid) and examined by light microscopy.
Behavioural experiment
After the acclimatization period, 30 sardines were transferred from the stocking tanks to the square-shaped tank and left there to acclimatize for another 2 days before experiments started. The 70 kHz dolphin pinger was tested during nine sessions, waiting at least 1 h between sessions in order to make sure that fish returned to normal conditions. According to Kastelein et al. (2008) an inter-trial interval of 2 min is already enough to restore active behavioural reactions (such as startle responses) of fish to acoustic stimuli. When all sessions were finished, the fish were fed and not manipulated anymore until the next day.
Each session started with a 15-min period during which the placebo was suspended into the tank, followed immediately by a 15-min period during which the active pinger was placed into the tank, emitting sounds. The sequence of placebo and active pinger exposure was not randomized, because using the active pinger first might have caused a prolonged effect on fish behaviour (in case the sounds are audible for the fish) that may have biased the behavioural reactions of fish in the subsequent placebo trials. The pingers were introduced slowly into the tank and suspended from ropes about 20 cm from the pool wall, at about half way up the water column.
During each session, the fish behaviour was videotaped in continuous real-time video with a high-definition underwater camera (GoPro HD Hero 960), fixed with a bendable base at the tank wall at around 10 cm below the surface (Fig. 3). The original curved lens of the underwater camera housing was replaced by a flat methacrylate lens to increase the definition of the camera images. To maintain sufficient light for the video images, the light over the experimental tank was switched on at least 30 min before the first session started. Additional to videotaping, the behavioural reactions of the fish were examined by eye and documented by an observer.
Data analysis
Analysis of video recordings
Video footage was processed using Avidemux 2.5, a free open-source programme. A 10-min sequence with the same start and end point (minute 5–15) was cut out of each 15 min recording. A scan sampling technique was used, stopping the video sequence at 2, 4, 6, 8 and 10 min and determining the swimming parameters of fish on these screenshots.
Fish school compaction was measured as the distance between the fish closest to the bottom and the fish furthest away from it. The distance of the fish school to the bottom of the tank was measured as the distance of the snout of each fish to the bottom of the tank. For the measurement of swimming speed (in m s−1), five fish were selected from each screenshot, the video recording was run for 3 s in slow motion (one-half of original speed), and the distance swum by each fish was determined. The values for the five fish were averaged afterwards. Distances in the video images were measured using Small Measure v 1.0 (1 hour Software), a small screen ruler to determine the number of pixels between two points on the screen, and converted into cm with the help of the reference grid on the tank walls.
All measurements were averaged for each video screenshot and trial.
Statistical analysis
Statistical analysis was performed with IBM SPSS Statistics 19 and, for Modelling, with Brodgar 2.7.2.
Mixed effects models provide a powerful tool to analyse unbalanced nested data, because they allow for the inclusion of fixed and random effects as well as for correlation between observations within the sampling unit (Zuur et al., 2009)
In the present study, the data were three-way nested because several fish were sampled from two different tanks (physiological experiment)/during various video screenshots (behavioural experiment) in multiple trials (repeated measures) and therefore correlation between the response variables within each sampling unit may be expected.
It was first assessed whether random effects and a multiple variance structure (i.e. allowing for unequal variances) needed to be included into the model by visualizing the amount of variation of the fixed explanatory variables between tanks/video screenshots and between trials using conditional box plots.
A generalized least square (GLS) model was fitted, including as many fixed explanatory variables and their interactions as possible, and this model was compared with a linear mixed effects (LME) model that additionally included the nested random effects “tank/video screenshot” and “trial” using the anova function. The model structure was
-
Physiological experiment:
Control-placebo/active pinger ijk + Sex ijk + Maturity ijk + Condition Factor + Condition Factor ijk × Sex ijk + Condition Facor ijk × Maturity ijk
-
Behavioural experiment:
Placebo-active pinger ijk ,
where i = observation, j = tank/video screenshot and k = trial
Biological data were derived from dead fish and could therefore only be included into the models for physiological experiments since behavioural observations did not imply the killing of test animals.
The best model was chosen, i.e. the one with the lowest value for the Akaike Information Criterion (AIC) and it was assessed whether the inclusion of a multiple variance structure of the fixed explanatory variables could improve the model by comparing the AICs of the previous model and a model containing the variance function VarIdent.
To find the optimal model in terms of the fixed explanatory variables, Likelihood Ratio Tests were used as some factors had more than two levels. This procedure included fitting a full model, dropping all allowable terms in turn, applying Likelihood Ratio Tests of nested models, dropping the least significant term and repeating the whole process until all terms were significant.
The final model was then validated by checking if the assumptions of homogeneity and independence of residuals were met, also checking for the existence of influential data points.
Results
Biological data
The biological data for the experimental sardines are summarized in Table 1. Only 5.3% of the fish were mature, while 40.8% were maturing and 53.9% immature. The sex ratio of female to male fish was 53:47. The mortality rate of fish was 5% during transport and 17.8% during acclimatization.
Hormonal stress response of sardine to 70 kHz dolphin pinger sounds
Sardine plasma cortisol concentrations differed significantly before and after exposure to active 70 kHz dolphin pingers (GLS, meanbefore = 0.86 μg dl−1, meanafter = 1.42 μg dl−1, t = 3.61; P = 0.001), while for the placebo no significant difference between pre- and post-values was detected (Fig. 6). However, there was also a high inter-trial variability between pre- and post-exposure values (Fig. 7).

Box plots representing plasma cortisol concentrations of sardine before (pre) and after (post) exposure to placebo and active 70 kHz dolphin pingers, pooled across experimental trials. The box stretches from the 25th to the 75th percentile. The line across the box represents the median values. The ends of the vertical line indicate the minimum and maximum data values. Individual points are considered outliers

Line chart representing inter-trial differences in mean plasma cortisol concentrations of sardine before (dashed black line) and after (solid grey line) exposure to placebo (white boxes) and active (black dots) 70 kHz dolphin pingers
The biological parameters condition factor, sex and maturity had no significant effect on the plasma cortisol concentrations.
Behavioural reactions of sardines to 70 kHz dolphin pinger sounds
There were no significant differences in mean distance to the tank bottom and swimming speed of sardine between placebo and active pinger trials (Fig. 8a, b). However, the mean sardine fish school height was significantly lower for the active 70 kHz dolphin pinger than for the placebo (GLS, meanpinger = 33.85 cm, meanplacebo = 42.87 cm, t = −2.89; P = 0.005) (Fig. 8c). Differences in mean fish school height between active pingers and placebos varied greatly between trials (Fig. 9).

Box plots representing differences in behavioural reactions of sardines to active 70 kHz dolphin pingers and placebos, pooled across experimental trials: a distance of fish school to the bottom of the tank, b swimming speed and c fish school height (school compaction). The box stretches from the 25th to the 75th percentile. The line across the box represents the median values. The ends of the vertical line indicate the minimum and maximum data values. Individual points are considered outliers

Line chart representing inter-trial differences in mean fish school height during active 70 kHz pinger (solid black line) and placebo (dashed grey line) exposure
Discussion
Although some significant effects of pingers on fish plasma cortisol level and behaviour were detected, the responses of the fish to the pinger sounds were very subtle in the present experimental survey. No immediate behavioural reactions of the fish to the pinger sounds were observed during the whole course of the experiment.
A slight increase in plasma cortisol concentrations after exposure to active 70 kHz dolphin pingers was found, although the mean cortisol increment (0.56 μg dl−1 per hour) and the post-stress cortisol level (1.42 μg dl−1) in the present experiment were very small when compared to the values reported by Marçalo et al. (2006) who measured a five-fold increase in mean cortisol concentrations in sardines during fishing simulation experiments, corresponding to an increase rate of 6.9 μg dl−1 per hour and a post-stress value of 8.9 μg dl−1. The measured values in the present study are also well below the characteristic cortisol elevations of fish in response to acute stressors which, according to Barton & Iwama (1991) and Wedemeyer et al. (1990), tend to range between 3 and 30 μg dl−1. Of the three behavioural parameters observed, only fish school height (i.e. level of fish school compaction) showed significant differences between active pinger and placebo trials, the relative decrease being only about 25%. This difference is relatively moderate when compared to the results of Marçalo (2009), where sardine group cohesiveness was observed to double and swimming speed showed a five-fold increase when fish were exposed to active stressors such as natural predators in an experimental tank. It is important to note that the mean plasma cortisol level of sardines increased during both, active and placebo pinger trials. This may indicate that the insertion of the device into the test tank might have caused the main effect on the fish (e.g. through associated noise and/or pressure waves that may have been detectable for the sardines). Nevertheless, even if an “insertion effect” existed, the mean rate of cortisol increase was only significant during active pinger trials. Furthermore, base cortisol levels varied significantly between trials in the present study and the pre-pinger cortisol level was lower than the pre-placebo level. This suggests that differences in cortisol concentrations may also have been caused by environmental factors, such as changes in water temperature and salinity, or by the biological characteristics (e.g. condition, developmental stage) of the fish tested (Barton, 2002). Fish were held in open-circuit pumped seawater, where slight temporal variations in water temperature and salinity do naturally occur. Although no significant effect of biological parameters on the mean base plasma cortisol concentration was detected over the whole survey, they may be influential when inter-trial variance (five fish per trial) is analysed. However, by adding random effects to the statistical model, inter-trial variance has already been taken into account. Fish handling may also introduce certain bias, since manipulation of fish, i.e. capture and blood withdrawal, provokes an immediate elevation of plasma cortisol that can be detected within 30 s of applying an acute stressor (Gerwick et al., 1999). The removal of single fish from the tank can also cause increased stress levels in the remaining fish which may increase basal cortisol concentrations in subsequent experiments (Laidley & Leatherland, 1988). As the risk for handling stress can be minimized by rapid blood withdrawal of anaesthetised fish (Pottinger et al., 1992; Olsen et al., 1995), fish were captured from the tank all at once within a few seconds, introducing them immediately into the sedation basin and taking blood samples as soon as animals were fully anaesthetised. By leaving at least 2 days between consecutive trials, the experimental fish were assumed to be able to return to their cortisol base levels, which should usually already be achieved within six hours from an acute stress (Iwama et al., 2006). The base cortisol levels measured in the present study were comparable to levels of unstressed sardines in literature (see Marçalo et al., 2006, 2008), so fish handling should not have had a significant impact on the measured cortisol levels. Nevertheless, slight handling effects cannot be completely ruled out.
Concerning behavioural reactions, the interpretation of results is more difficult. As space in the test tank was clearly limited, the schooling behaviour of sardines may have been different than in the wild. Although fish school compaction was higher for active pingers than for placebos in most trials, the opposite was observed during two trials. External factors, such as ambient noise or slight unintended differences in the procedure to introduce the device into the tank may have been influential here.
The lack of significant observable and measurable reactions of the fish to the pinger sounds in the present survey indicates that sardines do not perceive the pinger signals as a sign of imminent danger. In addition, even if sardines responded with school cohesion to the pinger sounds, this would not have a negative effect on catch rates since fish would be more concentrated and therefore probably easier to catch in a real fishing scenario. The 70 kHz dolphin pinger operates exclusively in the ultrasonic range and should therefore not be audible for most fish species that cannot detect sound above 1.5 kHz. Being a member of the family Clupeidae, sardine may, however, be able to hear sounds up to about 4 or 5 kHz, as demonstrated for scaled sardine Harengula jaguana (Poey, 1865) and round sardinella Sardinella aurita (Valenciennes, 1847), or even in the ultrasonic range just as American shad and Gulf menhaden Brevoortia patronus (Goode, 1878) (Mann et al., 2001; Wilson et al., 2009). Nevertheless, even though sardines may be able to detect high-frequency sounds, the source level (<150 dB re 1 mPa) and pulse repetition rate (maximum 15 pulses min−1) of the pinger sounds are probably not high enough to cause aversive reactions. According to Popper et al. (2004), agitated responses of clupeid fish, leading to movement away from the sound source are usually not observed until the ultrasound gets more intense (175–184 dB re 1 mPa). In addition, Wilson et al. (2011) found that allis shad Alosa alosa (Linnaeus, 1758), another clupeid fish, only reacted to ultrasound clicks with a repetition rate of at least 20 clicks s−1, suggesting that a single ultrasonic click may be detected, but not necessarily be interpreted as danger.
Recommendations for future research
An “ideal pinger” should allow for maximum deterrence of the cetacean species targeted, while exerting minimum negative impact on non-target cetaceans and fish (Reeves et al., 2001). Aversive reactions to sounds are caused by the sound pressure level, frequency spectrum and temporal aspects of sounds. Therefore, the source level of the pinger sound should be loud enough to cause aversion in cetaceans, but not excessively loud to preclude the audibility of pinger sounds by targeted fish and to avoid that cetacean species with more sensitive hearing, such as harbour porpoise (Kastelein et al., 2010), may be excluded from their habitat or even suffer hearing damage (Culik et al., 2001; Gordon & Northridge, 2002). Therefore, a high-frequency range (≥10 kHz), moderate source level (<160 dB re 1 lPa @ 1 m), low pulse repetition rate and a short pulse duration (<500 ms) are recommended features (Plachta & Popper, 2003; Kastelein et al., 2007; Wilson et al., 2011).
Apart from these technical characteristics, the choice of pinger also largely depends on the scope of application (i.e. fisheries affected and cetacean species involved) as well as on practical aspects, such as the ease of operation and price of pingers. Therefore, as a next step, fishery-specific long-term field trials should be conducted with the active co-operation of affected fisheries to assess pinger efficiency and the magnitude of possible side effects on non-target cetaceans and fish species, as well as the willingness of local fishers to accept this mitigation tool.
Notes
Council Regulation (EC) 812/2004 of 26 April 2004 laying down measures concerning incidental catches of cetaceans in fisheries and amending Regulation (EC) 88/98.
References
Abaunza, P., A. C. Fariña & P. Carrera, 1995. Geographic variations in sexual maturity of the horse mackerel, Trachurus trachurus, in the Galician and Cantabrian shelf. Scientia Marina 59: 211–222.
Aguilar, A., 1997. Inventario de los cetáceos de las águas atlánticas peninsulares: aplicación de la directiva 92/43/CEE. Memoria Final. Departamento de Biología Animal (Vert.), Facultad de Biología, Universitat de Barcelona.
Barlow, J. & G. A. Cameron, 2003. Field experiments show that acoustic pingers reduce marine mammal bycatch in the California drift gillnet fishery. Marine Mammal Science 19: 265–283.
Barton, B. A., 2002. Stress in fishes: a diversity of responses with particular reference to changes in circulating corticosteroids. Integrative and Comparative Biology 42: 517–525.
Barton, B. A. & G. K. Iwama, 1991. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annual Review of Fish Diseases 1: 3–26.
Borges, M. F., A. M. P. Santos, N. Crato, H. Mendes & B. Mota, 2003. Sardine regime shifts off Portugal: a time series analysis of catches and wind conditions. Scientia Marina 67: 235–244.
Brotons, J. M., Z. Munilla, A. M. Grau & L. Rendell, 2008. Do pingers reduce interactions between bottlenose dolphins and nets around the Balearic Islands? Endangered Species Research 5: 1–8.
Buscaino, G., G. Buffa, G. Sara, A. Bellante, A. J. Tonello Jr, F. A. S. Hardt, M. J. Cremer, A. Bonanno, A. Cuttitta & S. Mazzola, 2009. Pinger fish catch efficiency and damage to bottom gill nets related to bottlenose dolphins. Fisheries Science 75: 537–544.
Carlström, J., P. Berggren & N. J. C. Tregenza, 2009. Spatial and temporal impact of pingers on porpoises. Canadian Journal of Fisheries and Aquatic Sciences 66: 72–82.
Carretta, J. V. & J. Barlow, 2011. Long-term effectiveness, failure rates, and “dinner bell” properties of acoustic pingers in a gillnet fishery. Marine Technology Society Journal 45: 7–13.
Cox, T. M., A. J. Read, D. Swanner, K. Urian & D. Waples, 2003. Behavioral responses of bottlenose dolphins, Tursiops truncatus, to gillnets and acoustic alarms. Biological Conservation 115: 203–212.
Culik, B. M., S. Koschinski, N. Tregenza & G. M. Ellis, 2001. Reactions of harbour porpoises Phocoena phocoena and herring Clupea harengus to acoustic alarms. Marine Ecology Progress Series 211: 255–260.
Dawson, S. M., S. Northridge, D. Waples & A. J. Read, 2013. To ping or not to ping: the use of active acoustic devices in mitigating interactions between small cetaceans and gillnet fisheries. Endagered Species Research 19: 201–221.
Fernández, P. M., P. Bustamante, A. Bode, T. Chouvelon, M. Ferreira, A. López, G. J. Pierce, M. B. Santos, J. Spitz, J. Vingada & F. Caurant, 2012. Foraging ecology of five toothed whale species in the Northwest Iberian Peninsula, inferred using carbon and nitrogen isotope ratios. Journal of Experimental Marine Biology and Ecology 413: 150–158.
Galician Ministry of Fisheries [Xunta de Galicia, Consellería do Mar], 2013. Online statistics. http://www.pescadegalicia.com.
Gazo, M., J. Gonzalvo & A. Aguilar, 2008. Pingers as deterrents of bottlenose dolphins interacting with trammel nets. Fisheries Research 92: 70–75.
Gerwick, L., N. E. Demers & C. J. Bayne, 1999. Modulation of stress hormones in rainbow trout by means of anaesthesia, sensory deprivation and receptor blockade. Comparative Biochemistry and Physiology 124: 329–334.
Goetz, S., F. L. Read, M. B. Santos, C. Pita & G. J. Pierce, 2014. Cetacean-fishery interactions in Galicia (NW Spain): results and management implications of a face-to-face interview survey of local fishers. ICES Journal of Marine Science 71: 604–617.
Gönener, S. & S. Bilgin, 2009. The effect of pingers on harbour porpoise Phocoena phocoena bycatch and fishing effort in the turbot gill net fishery in the Turkish Black Sea coast. Turkish Journal of Fisheries and Aquatic Sciences 9: 151–157.
Gönener, S. & S. Özdemir, 2012. Investigation of the interaction between bottom gillnet fishery (Sinop, Black Sea) and bottlenose dolphins (Tursiops truncatus) in terms of economy. Turkish Journal of Fisheries and Aquatic Sciences 12: 115–126.
Gordon, J., & S. Northridge, 2002. Potential impacts of acoustic deterrent devices on Scottish Marine Wildlife. Scottish Natural Heritage Commissioned Report F01AA404.
Iwama, G. K., L. O. B. Afonso & M. M. Vijayan, 2006. Stress in fishes. In Evans, D. H. & J. B. Claiborne (eds), The Physiology of fishes, 3rd ed. Taylor and Francis, Boca Raton, FL: 319–342.
James, A. G., L. Hutchings, C. L. Brownell & D. A. Horstman, 1988. Methods of capture and transfer to the laboratory of wild pelagic fish. South African Journal of Marine Science 6: 17–21.
Kastelein, R. A., S. van der Heul, J. van der Veen, W. C. Verboom, N. Jennings, D. de Haan & P. J. H. Reijnders, 2007. Effects of acoustic alarms, designed to reduce small cetacean bycatch in gillnet fisheries, on the behaviour of North Sea fish species in a large tank. Marine Environmental Research 64: 160–180.
Kastelein, R. A., S. van der Heul, W. C. Verboom, N. Jennings, J. van der Veen & D. de Haan, 2008. Startle response of captive North Sea fish species to underwater tones between 0.1 and 64 kHz. Marine Environmental Research 65: 369–377.
Kastelein, R. A., L. Hoek, C. A. F. de Jong & P. J. Wensveen, 2010. The effect of signal duration on the underwater detection thresholds of a harbor porpoise (Phocoena phocoena) for single frequency-modulated tonal signals between 0.25 and 160 kHz. Journal of the Acoustic Society of America 128: 3211–3222.
Kraus, S. D., A. J. Read, E. Anderson, K. Baldwin, A. Solow, T. Spradlin & J. Williamson, 1997. Acoustic alarms reduce porpoise mortality. Nature 388: 525.
Laidley, C. W. & J. F. Leatherland, 1988. Cohort sampling, anaesthesia and stocking density effects on plasma cortisol, thyroid hormone, metabolite and ion levels in rainbow trout, Salmo gairdneri Richardson. Journal of Fish Biology 33: 73–88.
López, A., G. J. Pierce, M. B. Santos, J. Gracia & A. Guerra, 2003. Fishery by-catches of marine mammals in Galician waters: results from on-board observations and an interview survey of fishermen. Biological Conservation 111: 25–40.
López, A., J. A. Vázquez, J. A. M. Cedeira, A. Cañadas, E. Marcos, I. Maestre, A. Ruano, L. Larias, A. Llavona, C. MacLeod & P. Evans, 2013. Abundance estimates for harbour porpoise (Phocoena phocoena) in the Spanish area of the Iberian Peninsula Management Unit. IWC Scientific Committee Annual Meeting, 4–15 June 2013, Jeju Island, Republic of Korea. SC/65a/SM20.
Mann, D. A., Z. Lu & A. N. Popper, 1997. A clupeid fish can detect ultrasound. Nature 389: 341.
Mann, D. A., Z. Lu, M. C. Hastings & A. N. Popper, 1998. Detection of ultrasonic tones and simulated dolphin echolocation clicks by a teleost fish, the American shad (Alosa sapidissima). The Journal of the Acoustical Society of America 104: 562–568.
Mann, D. A., D. M. Higgs, W. N. Tavolga, M. J. Souza & A. N. Popper, 2001. Ultrasound detection by clupeiform fishes. The Journal of the Acoustical Society of America 109: 3048–3054.
Marçalo, A. L., 2009. Sardine (Sardina pilchardus) delayed mortality associated with purse seine slipping: contributing stressors and responses. PhD thesis. Faculdade de Ciências e tecnologia, University of the Algarve, Portugal.
Marçalo, A., L. Mateus, J. H. D. Correia, P. Serra, R. Fryer & Y. Stratoudakis, 2006. Sardine (Sardina pilchardus) stress reactions to purse seine fishing. Marine Biology 149: 1509–1518.
Marçalo, A., P. Pousão Ferreira, L. Mateus, J. H. Duarte Correia & Y. Stratoudakis, 2008. Sardine early survival, physical condition and stress after introduction to captivity. Journal of Fish Biology 72: 103–120.
Misund, O. A., J. T. Ovredal & M. T. Hafsteinsson, 1996. Reactions of herring schools to the sound field of a survey vessel. Aquatic Living Resources 9: 5–11.
Olsen, Y. A., I. E. Einursdouir & K. J. Nilsson, 1995. Metomidate anaesthesia in Atlantic salmon, Salmo salar, prevents plasma cortisol increase during stress. Aquaculture 134: 155–168.
Pitcher, T., O. Misund, A. Ferno, B. Totland & V. Melle, 1996. Adaptive behaviour of herring schools in the Norwegian Sea as revealed by high-resolution sonar. ICES Journal of Marine Science 53: 449–452.
Plachta, D. T. T. & A. N. Popper, 2003. Evasive responses of American shad (Alosa sapidissima) to ultrasonic stimuli. Acoustics Research Letters Online 4: 25–30.
Popov, V. V. & V. O. Klishin, 1998. EEG study of hearing in the common dolphin, Delphinus delphis. Aquatic Mammals 24: 13–20.
Popper, A. N. & M. C. Hastings, 2009. Effects of anthropogenic sources of sound on fishes. Journal of Fish Biology 75: 455–498.
Popper, A. N. & C. R. Schilt, 2008. Hearing and acoustic behavior (basic and applied). In Webb, J. F., R. R. Fay & A. N. Popper (eds), Fish Bioacoustics. Springer, New York: 17–48.
Popper, A. N., D. T. T. Plachta, D. A. Mann & D. Higgs, 2004. The response of clupeid fishes to ultrasound: a review. ICES Journal of Marine Science 61: 1057–1061.
Porchas, M. M., L. R. M. Córdova & R. R. Enriquez, 2009. Cortisol and glucose: reliable indicators of fish stress? Pan-American Journal of Aquatic Sciences 4: 158–178.
Portuguese Directorate General of Natural Resources, Security and Maritime Services [Direção-Geral de Recursos Naturais, Segurança e Serviços Marítimos], 2013. Online statistics. http://www.dgrm.min-agricultura.pt/xportal/xmain?xpid=dgrm.
Pottinger, T. G., T. A. Moran & P. A. Cranwell, 1992. The biliary accumulation of corticosteroids in rainbow trout, Oncorhynchus mykiss, during acute and chronic stress. Fish Physiology and Biochemistry 10: 55–66.
Pusineri, C., V. Magnin, L. Meynier, J. Spitz, S. Hassani & V. Ridoux, 2007. Food and feeding ecology of the common dolphin (Delphinus delphis) in the oceanic Northeast Atlantic and comparison with its diet in neritic areas. Marine Mammal Science 23: 30–47.
Read, F. L., M. B. Santos, A. F. González, A. López, M. Ferreira, J. Vingada & G. J. Pierce, 2012. Understanding harbour porpoise (Phocoena phocoena) and fishery interactions in the north-west Iberian Peninsula. Final report to ASCOBANS (SSFA/ASCOBANS/2010/4).
Reeves, R. R., R. J. Hofman, G. K. Silber & D. Wilkinson, 1996. Acoustic deterrence of harmful marine mammal-fishery interactions: proceedings of a workshop held in Seattle, Washington, 20-22 March 1996. NOAA Tech. Memo. NMFS-OPR-10.
Reeves, R. R., A. J. Read & G. N. di Sciara, 2001. Report of the workshop on interactions between dolphins and fisheries in the Mediterranean: evaluation of mitigation alternatives. Instituto Centrale per la Ricerca Applicata al Mare, Rome, Italy.
Santos, M. B., R. Fernández, A. López, J. A. Martínez & G. J. Pierce, 2007. Variability in the diet of bottlenose dolphin, Tursiops truncatus (Montagu), in Galician waters, NW Spain, 1990–2005. Journal of the Marine Biological Association of the UK 87: 231–242.
Santos, M. B., I. German, D. Correia, F. L. Read, J. M. Cedeira, M. Caldas, A. López, F. Velasco & G. J. Pierce, 2013. Long-term variation in common dolphin diet in relation to prey abundance. Marine Ecology Progress Series 481: 245–268.
Schwarz, A. L. & G. L. Greer, 1984. Responses of Pacific herring, Clupea harengus pallasi, to some underwater sounds. Canadian Journal of Fisheries and Aquatic Sciences 4: 1183–1192.
Shaw, B. L. & H. I. Battle, 1957. The gross and microscopic anatomy of the digestive tract of the oyster Crassostrea virginica (Gmelin). Canadian Journal of Zoology 35: 325–347.
Silva, M. A., 1999. Diet of common dolphins, Delphinus delphis, off the Portuguese continental coast. Journal of the Marine Biological Association of the UK 79: 531–540.
Simón Díaz, M., 2009. Maduración gonadal e inducción al desove de la sardina del Pacífico Sardinops sagax caeruleus. M.Sc. Thesis. Centro de Investigación Científica y de Educación Superior de Ensenada, Baja California, Mexico.
Sollmann, K., 2011. Diet and distribution of the bottlenose dolphin (Tursiops truncatus) in Galicia, north-west Spain. M.Sc. dissertation, Julius-Maximilians-Universität Würzburg, Germany.
Suuronen, P., E. Lehtonen & J. Wallace, 1997. Avoidance and escape behaviour of herring encountering midwater trawls. Fisheries Research 29: 13–24.
Trippel, E. A., M. B. Strong, J. M. Terhune & J. D. Conway, 1999. Mitigation of harbour porpoise (Phocoena phocoena) by-catch in the gillnet fishery in the lower Bay of Fundy. Canadian Journal of Fisheries and Aquatic Sciences 56: 113–123.
Vingada, J., M. Ferreira, A. Marçalo, J. Santos, H. Araújo, I. Oliveira, S. Monteiro, S. Nicolau, P. Gomes, C. Tavares & C. Eira, 2011. SafeSea—Manual de Apoio para a Promoção de uma Pesca Mais Sustentável e de um mar seguro para cetáceos; Programa EEAGrants—EEA Financial Mechanism 2004–2009 (Projecto 0039). Braga, Portugal.
Wedemeyer, G. A., B. A. Barton & D. J. McLeay, 1990. Stress and acclimation. In Schreck, C. B. & P. B. Moyle (eds), Methods for Fish Biology. American Fisheries Society, Bethesda: 491–527.
Wilson, B. & L. M. Dill, 2002. Pacific herring respond to simulated odontocete echolocation sounds. Canadian Journal of Fisheries and Aquatic Sciences 59: 542–553.
Wilson, M., E. W. Montie, K. A. Mann & D. A. Mann, 2009. Ultrasound detection in the Gulf menhaden requires gas-filled bullae and an intact lateral line. Journal of Experimental Biology 212: 3422–3427.
Wilson, M., H. B. Schack, P. T. Madsen, A. Surlykke & M. Wahlberg, 2011. Directional escape behavior and energy detection in allis shad (Alosa alosa) exposed to ultrasonic clicks mimicking an approaching toothed whale. Journal of Experimental Biology 214: 22–29.
Wysocki, L. E., J. P. Dittami & F. Ladich, 2006. Ship noise and cortisol secretion in European freshwater fishes. Biological Conservation 128: 501–508.
Zuur, A. F., E. N. Ieon, N. J. Walker, A. A. Saveliev & G. M. Smith, 2009. Mixed effects modelling and extensions in ecology with R. Statistics for Biology and Health. Springer, New York.
Acknowledgments
The study was funded by the Portuguese Ministry of Science (Fundação para a Ciência e Tecnologia– FCT) through a PhD Grant of SG (SFRH/BD/47931/2008). We would like to thank the captain of the purse-seiner (José Manuel Saveedra) and his crew for facilitating the capture and transport of live fish. Moreover, we want to thank Ana Marçalo for suggestions on the experimental design, Manuel Garci for technical advice on underwater video recordings and James Turner from the company Future Oceans for providing technical details on the 70 kHz dolphin pingers. We would also like to acknowledge the scientific advice of Dr. José Iglesias and the technical and logistic support for the preparation of the laboratory and the materials for tank experiments by Enrique Martínez González, Ricardo Pazó and other staff at the aquaculture facilities of the Spanish Institute for Oceanography (IEO) and the Marine Sciences Station of Toralla (ECIMAT) in Vigo. Furthermore, we are grateful to Francisco de la Granda Grandoso for his practical assistance during the fish tank experiments and to Juan Santos Blanco for helping with statistical analysis. Finally, we would like to thank Pilar Riobó Agula, Amelia Fernandez Villamarin, José Franco Soler, José Luis Muñoz, Angela Benedetti, Marcos Antonio Lopez Patiño and Marta Conde Sieira for scientific advice and practical support with cortisol analysis and Rosana Rodríguez for preparing histological samples for us.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Vasilis Valavanis
Rights and permissions
About this article

Cite this article
Goetz, S., Santos, M.B., Vingada, J. et al. Do pingers cause stress in fish? An experimental tank study with European sardine, Sardina pilchardus (Walbaum, 1792) (Actinopterygii, Clupeidae), exposed to a 70 kHz dolphin pinger. Hydrobiologia 749, 83–96 (2015). https://doi.org/10.1007/s10750-014-2147-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-014-2147-3