
Abstract
To ward off pathogens, plants establish innate immunity system in which salicylic acid (SA) plays an important role. The downy mildew of foxtail millet (Setaria italica) is an oomycete disease caused by biotrophic pathogen Sclerospora graminicola, which seriously affects the yield and quality of foxtail millet. However, it is not clear how SA regulating the defense responses of foxtail millet. In this study, we found that spraying exogenous SA of low concentration (1 mM and 3 mM) had good effect on enhancing the resistance while high concentration (6 mM and 9 mM) did not significantly affect the defense of foxtail millet. Excess SA inhibited the growth and development of plants and we found spraying exogenous SA reduced the plant height, fresh weight and dry weight of foxtail millet. The low concentration of exogenous SA promoted the accumulation of Chlorophyll, soluble sugar and proline, promoted the activity of Phenylalanine Ammonia-Lyase and suppressed the accumulation of Malondialdehyde. After the treatment with 1 mM or 3 mM of SA, the expressions of genes related to disease resistance were increased at the early stage after treatment. The results indicated exogenous SA could enhance the resistance of foxtail millet against the pathogen and provided a basis for improving the control methods of downy mildew.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Foxtail millet (Setaria italica (L.) P. Beauv.), a kind of cereal crops used for food and feed, is originated from China and has a long cultivation history (Zhang et al. 2012). Meanwhile, foxtail millet is a highly light-efficient C4 plant with strong environmental adaptability which is a suitable crop to ensure food and nutritional security in the face of a quickly growing world population, particularly in arid and barren area. In addition, foxtail millet is a self-pollinated diploid plant with a small genome (~ 490 Mb) and its genome has been sequenced (Peng and Zhang 2020). These characteristics are promoting it as a new C4 model plant (Doust et al. 2009). Recently a mini foxtail millet with an Arabidopsis-like life cycle was constructed, which laid a foundation for the exploration of special developmental mechanism in C4 plants (Yang et al. 2020). In recent years, foxtail millet with high nutritional value is favored by human beings because its grains contain higher content of amino acids and vitamins compared with other cereal crops (Hou et al. 2022).
The yield and quality of foxtail millet are seriously affected by the diseases caused by various pathogens. Downy mildew disease causing serious yield loss is prevalent in main production area of foxtail millet including China, India, Japan and Russia. The downy mildew is caused by Sclerospora graminicola, a kind of oomycete whose genome sequencing was completed (Kobayashi et al. 2017). Sclerospora graminicola causes downy mildew disease in Poaceae plants including foxtail millet and pearl millet. After infected by Sclerospora graminicola, rapid and increased activation of mitogen-activated protein kinase (MAPK) of pearl millet was observed (Melvin et al. 2014). The data of transcriptome analysis showed pathways for phenylpropanoid metabolism, transcripts of genes encoding R proteins, HR induced proteins and plant hormonal signaling transduction proteins were up-regulated in genotype resistant to Sclerospora graminicola (Kulkarni et al. 2016). In foxtail millet, the secretion of virulence effectors from Sclerospora graminicola contribute to pathogenicity by suppressing plant defence responses (Kobayashi et al. 2022). This pathogen may also affect signaling of phytohormones, including inhibited the growth and development of the host by disturbing the gibberellin signal transduction (Li et al. 2020).
Phytohormone regulates not only plant development but also plant resistance to pathogens. For a long time, salicylic acid (SA) has been used as medicine and was found as chemical messengers in regulating biological processes at low concentration. Thus, SA was introduced as the ‘sixth’ principal phytohormone (Raskin 1992). More and more studies demonstrate that SA is a key plant hormone required for establishing resistance to many pathogens (Ding and Ding 2020). In the long-term co-evolution process with pathogens, plants have gradually formed system for resisting external microbial stimuli is called natural plant immune system, which is divided into two layers (Dangl et al. 2013). The first layer of defense system is called pathogen-associated molecular pattern-triggered immunity (PTI). Recognition of pathogen-associated molecular patterns (PAMPs) through pattern recognition receptors (PRRs) can rapidly trigger basic immunity which can effectively inhibit the growth of pathogenic bacteria and control the disease (Bigeard et al. 2015). Plants begin to accumulate SA between 3 and 6 h after treated with PAMPs. SA contributes to PTI against Pseudomonas syringae and that disruption of SA signaling affects expression of PAMP-regulated genes (Tsuda et al. 2008). Pathogens use their effectors to subvert PTI, to facilitate nutrient acquisition, and to contribute to dispersal. Plants have evolved R genes encode polymorphic superfamily of intracellular nucleotide-binding leucine-rich repeat (NLR) receptors, which function intracellularly and anchor the second tier of the plant immune system (effector-triggered immunity, ETI). ETI is more stronger than PTI which involved pathogen-induced hypersensitive response (HR). In Arabidopsis thaliana, hrl1 mutants which accumulates higher level of endogenous SA shows reduced HR associated ion leakage (Devadas and Raina 2002). Furthermore, SA-deficient mutants of Arabidopsis display enhanced immune-associated ion leakage (Radojičić et al. 2018). Therefore, SA and its related metabolites are also necessary in ETI especially in regulating HR and cell death.
As SA plays important roles in immunity system, exogenous application of SA can improve the plant resistance to biotic stresses. For example, SA can enhance tomato plant resistance by modulating the activity of resistance-related enzymes and inducing the expression of pathogenesis-related genes to produce systemic acquired resistance (Li et al. 2019). Exogenous SA significantly induce resistance of soybean against Fusarium solani infection and wheat variety against aphids (Bawa et al. 2019; Feng et al. 2021). However, previous studies on the functions of SA in immunity system of foxtail millet have been rarely reported. In the present study, we investigated whether exogenously spraying SA affected foxtail millet resistance to Sclerospora graminicola, and the effect of SA on plant development, the activation of resistance-related enzymes and the expression pattern of resistance-related genes. This study provides a theoretical basis for the prevention and treatment of foxtail millet downy mildew with SA as an inducing factor.
Materials and methods
Plant Material
Three foxtail millet varieties xiaomi, GBS and Jingu 21 were used in this study. Foxtail millet were sown in experimental plot located at 37 ° N, 112 ° E or in a phytotron (25℃, humidity is 50%, photoperiod of 16 h light/ 8 h darkness).
The downy mildew fungus: the oospores of downy mildew fungus were collected from the white hair and hedgehog-like panicle of diseased plants in the experimental field of Shanxi Agricultural University (harvested in 2020 and stored at -20 °C).
Pathogen infection
Mix evenly according to the mass ratio of oospores : seeds = 1 : 5, and the seed was planted in the seedling tray.
SA Treatment
The SA concentration was set at 5 levels of 0, 1, 3, 6 and 9 mmol/L, and two treatments of exogenous spraying and seed soaking were carried out respectively.
Exogenous spraying treatment: When the plants grew to the three-leaf stage, SA was sprayed with different concentrations to make the seedlings fully sprayed. Three biological replicates were performed for each treatment.
The incidence of disease was calculated at elongation stage and mature period. Since SA treatment can rapidly induce plant defense responses, in this study, samples were taken at 0 h before SA spraying and at 3, 6, 9, 12, and 24 h after treatment to determine the contents of chlorophyll, MDA, PRO, soluble sugar and PAL activity in seedlings. And the relative expression of PR1, PAL1, PAL2, SOD, POD1 and POD2 genes. Samples can be stored at -80 °C for later use.
Disease Incidence Survey
Diseased plants included foxtail millet in different growth period shown “grey back”, “white tip”, “white hair” or “hedgehog-like panicle”.
The incidence of disease (%) = Number of diseased plants / Total number of plants.
Determination of growth index
The disease-resistant variety GBS and the susceptible variety Jingu 21 were selected and cultured in Hoagland solution. The plant height, dry weight and fresh weight of foxtail millet were measured 5 days after SA treatment with different concentrations. Plant height: from the base of the stem to the growing point (cm); fresh weight: weighing with an electronic balance (g); dry weight: the foxtail millet seedlings after weighing fresh weight, fixation at 110℃ for 0.5 h, and then dried at 80℃ oven for 48 h until constant weight was measured with an electronic balance (g). Each indicator was measured with 10 biological replicates.
Determination of physiological and biochemical indexes inxiaomiseedling stage.
Chlorophyll and carotenoid were extracted from leaves with 80% acetone for 24 h. Then the absorption values of the extraction at 663, 646 and 470 nm wavelengths were measured. The method of calculations was described previously (Porra 2002). The contents of MDA, Proline, soluble sugar and PAL activity were detected according to the instructions of kits. Article Numbers of these kit used in this study are following.
MDA content kit: AKFA013M BOXBIO (Beijing).
Proline content kit: AKAM003M BOXBIO (Beijing).
Soluble content kit: AKPL008M BOXBIO (Beijing).
PAL activity kit: AKAM012M BOXBIO (Beijing).
qRT-PCR verified differential gene expression inxiaomiseedlings.
The expression levels of PR1 (Seita.2G024600), PAL1 (Seita.7G168700), PAL2 (Seita.1G240200), SOD (Seita.9G123900), POD1 (Seita.3G052400) and POD2 (Seita.5G462500) in seedlings were detected. With SiACTIN (Seita.5G464000) as the internal reference gene, the relative gene expression was calculated according to the method of 2−ΔCT Gene/ACTIN, and each sample was repeated three times. The total RNA of plants was extracted by the text kit of Sangon Biotech CO., LTD. cDNA synthesis was used the text kit of TINGEN BIOTECH (BEIJING) CO., LTD. TB Green Premix Ex TaqTM (Tli RNaseH Plus) used the text kit of TaKaRa BIO. Primers (Supplemental Table 1) were synthesized by Sangon (Shanghai) Biotech CO., LTD.
Experimental design and data analysis
Each experiment was performed with at least three biological replicates. Microsoft Excel 2019 was used to organize data, Origin 2021 and GraphPad Prism 8 were used for draw graphs, ANOVA (http://vassarstats.net/anova1u.html) was used to perform significant test (p < 0.05 was considered significant difference).
Results
Sclerospora graminicolainhibited the growth and disrupted panicle development of foxtail millet.
Downy mildew occurs in the whole growth period of foxtail millet and the symptoms from the germination to heading stage are different and special. Seeds of foxtail millet infected by Sclerospora graminicola may fail to germinate or the seedlings died before emergence. The abaxial surface of infected leaves displayed grey mildew which is called “grey back” (Fig. 1A). In addition, Sclerospora graminicola caused abnormal development of panicles of foxtail millet. Some disease plants did not produce spike at heading time and an etiolated leaf grew at the top plants instead. This kind of leaves was called “white tip” (Fig. 1B). With the senility of host plants, the“white tip” leaves turned brown and split longitudinally which looked like hair (“white hair”, Fig. 1 C). Some disease plants produced hedgehog-like panicles which was the result of inflorescences was replaced by leaves (Fig. 1D).

Disease symptoms of foxtail millet infected bySclerospora graminicola
(A) “Grey back”: grey mildew on the abaxial surface of infected leaves; (B) “white tip”: etiolated leaf grew at the top plants; (C) “white hair”: senescent leaves and split longitudinally (D) hedgehog-like panicles
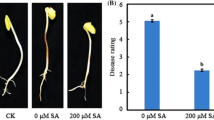
Low concentration of exogenous SA enhanced the disease resistance of foxtail millet
SA can improve the resistance to pathogens of some plants, we hypothesized that application of exogenous SA may enhance the resistance of foxtail millet to downy mildew. To prove this, we planted seeds of xiaomi (an Arabidopsis-like mini foxtail millet) mixed with oospores of Sclerospora graminicola. Three-leaf stage seedlings were sprayed with different concentrations of SA were used to spray the leaves to explore the effect of different SA concentrations on the downy mildew of foxtail millet, and the incidence of disease was investigated after heading stage. The ratio of plants displayed disease phenotype including grey back, white tip, white hair and hedgehog-like panicles were counted. The result showed that with the increase of SA concentration, the incidence of foxtail millet after spraying treatment showed a decreasing-increasing-decreasing trend (Fig. 2). The incidence of foxtail millet treated with 1 mM to 3 mM SA spray was lower than the control (0 mM SA). The incidence of disease under 1 mM SA spray treatment was significantly lower than other treatments, and the incidence rate decreased by 24.26% compared with 0 mM SA spray treatment. On the contrary, high concentration SA treatment did not reduce disease incidence, but made foxtail millet more susceptible to downy mildew.

Disease incidence survey of foxtail millet treated by exogenous SA
All plants displayed “grey back”, “white tip”, “white hair” or “hedgehog-like panicle” were counted as number of diseased plants. The incidence of disease (%) = Number of diseased plants / Total number of plants. Different lower-case letters indicate statistically significant differences (P < 0.05; ANOVA, 3 biological repeating groups and every group contains 50 plants)
Exogenous SA inhibits the growth and development of foxtail millet
Exogenous SA reduced the disease incidence, we tested whether SA affect the growth and development of Jingu 21 (JG21) and GBS, two different varieties of foxtail millet. JG21, a cultivar with the largest planting area in China, is very susceptible to downy mildew. GBS was another variety with high resistance to downy mildew. Through incidence survey in our experimental field, we found the average incidence in last 3 years of JG21 was 10 times higher than GBS (Supplemental Table 2).
In this study, the resistant variety GBS and susceptible variety JG21 were cultured in Hoagland Solution, and different concentrations of SA were sprayed at the three-leaf stage. It was found that SA treatment had inhibition on the growth of foxtail millet. With the increase of SA concentration, the overall trend of plant height, fresh weight and dry weight of both JG21 and GBS were downward (Supplemental Table 3). To compare the differences between the two varieties, we set the control group at 100% and analyzed the rate of growth change after SA treatments. The results showed that SA with low concentration (1 mM and 3 mM) reduced the growth plant height, fresh weight and dry weight of of foxtail millet, and the change rate of susceptible variety JG21 was higher than resistant variety GBS. SA with high concentration (6 mM and 9 mM) seriously suppressed the growth of foxtail millet, however, there was not significant different of the change rate between the two varieties (Fig. 3).

Exogenous SA inhibits the growth of foxtail millet
Seedlings of JG21 and GBS were sprayed by exogenous SA, Plant height (A), fresh weight (B) and dry weight (C) were measured 5 days after treatment. Data were shown as mean ± SD. Asterisks indicate statistically significant differences (“*”: P < 0.05; “***”: P < 0.001 n = 10, ANOVA)
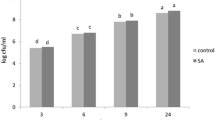
Effects of SA on chlorophyll content of foxtail millet
Chlorophyll is an important material in plant photosynthesis, whose content is closely related to the photosynthetic rate of plants. The content of chlorophyll is positively related to the net photosynthetic rate and it can also be used as one of the important index to reflect the growth status of foxtail millet seedlings. Data above indicated that the SA concentration was 1 mM to 3 mM to induce the disease resistance of foxtail millet and we wondered the effect of different concentrations of SA spraying on the chlorophyll content. We sampled at 6 time points before treating (0 h) and spraying after 3, 6, 9, 12, and 24 h, the chlorophyll content of leaves was measured and the relative content of chlorophyll was calculated (the ratio of chlorophyll content at 3, 6, 9, 12, 24 h and 0 h). The results indicated that with the application of 1 mM to 3 mM SA spraying, the chlorophyll content in foxtail millet leaves had the same trend, which showed a decrease-increase-decrease. At each time point after treatment, chlorophyll content in leaves at 3 mM SA was significantly higher than that at 1 mM SA (Fig. 4). The results showed that the appropriate concentration of SA could promote the accumulation of chlorophyll in seedlings, thereby affecting the growth of foxtail millet.

Exogenous SA affects the chlorophyll content of foxtail millet
Chlorophyll contents of xiaomi seedlings were detected at different time point after sprayed by 1 mM and 3 mM SA. Data were shown as mean ± SD. Different lower-case letters indicate statistically significant differences (P < 0.05, n = 3, ANOVA)
Exogenous SA affects the content of malondialdehyde, proline, soluble sugar and the activity of phenylpropanoid in foxtail millet.
In order to reveal the reason why SA improves disease resistance, we examined some substances related to plant resistance. Malondialdehyde (MDA) is the final product of cell membrane lipid peroxidation, which is toxic to cells, damages cell biofilms and can inhibit the activity of protective enzymes. Therefore, the degree of plant damage can be judged by MDA content. As shown in Fig. 5 A, the relative content of MDA in untreated plants showed an upward trend over time. The MDA content in foxtail millet treated with 1 mM or 3 mM SA was increased compared with 0 mM when 3–9 h after treatment. But at 24 h after treatment, the MDA content of 1 mM-3 mM SA spray treatment was significantly lower than that of 0 mM SA treatment.

Exogenous SA affects the content of substances and the enzymes activity related to resistance in foxtail millet
The content of malondialdehyde (A), soluble sugar (B), proline(C) and the activity of phenylpropanoid (D) of xiaomi seedlings were detected at different time point after sprayed by 1 mM and 3 mM SA. Data were shown as mean ± SD. Different lower-case letters indicate statistically significant differences (P < 0.05, n = 3, ANOVA)
Soluble sugar mainly participates in the respiration metabolism of organisms by providing energy for life activities. Soluble sugar can improve the osmotic regulation ability of plant cells and reduce the damage degree of plasma membrane under stresses. We found that the soluble sugar content of foxtail millet in 0 mM SA and 3 mM SA showed decreasing-increasing-decreasing trend with the processing time increases, while the change of soluble sugar content under 1mM SA was opposite. Under 3 mM SA treatment, the soluble sugar content increased significantly compared with 0mM treatment at 3 ~ 6 h and 12 ~ 24 h, and the maximum increased about 3 times (12 h). At 9 h after treatment, the soluble sugar content in foxtail millet treated with 3 mM SA was the lowest, and significantly lower than 0 mM treatment. The soluble sugar content of 1mM SA treatment increased significantly at 6 h ~ 12 h compared with 0 mM treatment, and the maximum increased about 1.6 times (Fig. 5B).
Proline (PRO) contribute to the stability of normal metabolism of plants under stress by regulating osmotic balance, scavenging reactive oxygen species and reducing the toxic substances. At the early stage of SA spraying treatment (3-6 h), the relative content of PRO was significantly increased when the SA concentration was 3 mM compared with the 0 mM treatment, while the relative content of PRO at the later stage of the treatment (9-24 h) was significantly lower than that in the 0 mM treatment; PRO relative content in the 1 mM SA treatment was significantly lower than that in the 0 mM treatment at different time points after spraying; with the prolongation of the treatment time, the change trend of the PRO content under the three SA concentration treatments was basically the same (Fig. 5 C).
Phenylpropanoid metabolism is an important pathway for the metabolism of secondary metabolites in plants, and the changes in enzyme activity of this pathway are closely related to plant disease resistance. In this study, we found that the phenylalanine ammonia lyase (PAL) activity in foxtail millet showed an overall decreasing-increasing-declining trend under 1 mM and 3 mM SA treatment, while the change of PAL activity under 0 mM SA treatment was opposite. With the increase of treatment time, PAL activity under 3 mM SA treatment was significantly lower than that under 0 mM SA treatment (Fig. 5D).
Exogenous SA modulated the expression level of resistance related genes in foxtail millet
To reveal the reason why SA improves disease resistance, we also tested the expression level of some resistance related genes. POD1, POD2 and SOD encode peroxidase (POD) and superoxide dismutase (SOD) which are the key enzymes in the oxygen free radical scavenging system. The expression levels of SiPOD1 and SiPOD2 in foxtail millet seedlings treated with 1 mM to 3 mM SA tended to be consistent with the prolongation of treatment time, showing an increase-decrease-increase. The expression levels of the two genes showed a turning point at 3 and 9 h after treatment. At 3–24 h after treatment, the expression of SiPOD1 treated with 1 mM-3 mM SA was significantly up-regulated; and the expression of SiPOD2 was also significantly up-regulated after treatment with 1 mM SA, and the expression of this gene was the highest at 3 h after treatment, which was higher than that before treatment (0 h) increased by about 35.6 times (Fig. 6 A-6B). The expression of SiSOD was significantly up-regulated at 3–12 h after treatment with 1 mM-3 mM SA compared with the before treatment (0 h), especially at 9–12 h, the expression of SiSOD treated with 3 mM SA was significantly increased compared with the other two concentrations; with the prolongation of treatment time, the expression trend of SiSOD gene was consistent, which showed that it first increased and then decreased (Fig. 6 C).

Exogenous SA modulated the expression level of resistance related genes in foxtail millet
Seedlings of xiaomi were treated with 1 mM and 3 mM exogenous SA. The transcriptional accumulation of genes related to plant resistance were detected by quantitative realtime PCR. SiACTIN was used as an internal control. Different lower-case letters indicate statistically significant differences (P < 0.05, n = 3, ANOVA)
PR1 gene is a downstream gene involved in the process of plant acquired resistance, and is involved in the process of MAPK regulating plant homeostasis. Studies have shown that SA can affect the expression of PR1. In this study, SiPR1 gene expression was detected by different concentrations SA leaf spraying. The results showed that the expression of SiPR1 gene in 1mM SA treated foxtail millet seedlings showed an increase-decrease-increase-decrease trend. However, after 3mM SA treatment, the change of gene expression was opposite, showing a decreasing-increasing-decreasing trend. The gene expression level after 1 mM and 3 mM SA treatment was significantly higher than that before treatment (0 h), but decreased after 24 h treatment (Fig. 6D).
SiPAL1 and SiPAL2 are key genes of PAL pathway which is necessary for plant resistance. Treating seedlings with 1mM SA could significantly increase the expression levels of SiPAL1 and SiPAL2 at the early stage (3-6 h). While it was opposite with 3 mM SA. the expression levels of SiPAL1 and SiPAL2 were significantly decreased at 3 h after treatment (Fig. 6E F).
Discussion
When infected by pathogens, plants can activate the defense system and improve the resistance to pathogens in which SA played a necessary role. In order to protect against pathogens effectively, plants activate systemic acquired resistance (SAR). SA acts as a mobile signal molecular for activating SAR because SA accumulates in both the primary infected and systemic uninfected tissue (Malamy et al. 1990). For this reason, exogenous SA is commonly used as the activator of plant resistance to biotic stresses (Feng et al. 2021). In particular, we showed that exogenous SA with appropriate concentration (1 mM ~ 3 mM) significantly activated the resistance of foxtail millet to downy mildew. Some studies indicated that plant cells pretreated with SA reacted more rapidly and effectively to subsequent pathogen (Bernsdorff et al. 2016), and SA did not directly inhibit the growth of pathogens (Bawa et al. 2019). SA mainly acts as a kind of priming signals which enhances plant immunity system by promoting the expression of resistance related genes and the accumulation of resistance related substances (Vlot et al. 2009). Phytohormones are a kind of substances that regulate plant life activities at low concentrations. SA accumulation is associated with the onset of HR and required for the spontaneous cell death phenotype (Radojičić et al. 2018). Therefore, excess SA negatively regulates the growth and development of plant. In the present study, high concentration of SA may inhibit the resistance to Sclerospora graminicola and strongly suppressed the plant height, fresh weight and dry weight of foxtail millet which may also disrupt the activation of plant resistance responses. Our finding was similar to previous studies (Deenamo et al. 2018). Hence, pretreating plants by SA with appropriate concentration effectively enhance plant resistance. However, excessive SA may first inhibit the normal growth of plants which is not conducive to the activation of immune responses.
To survive better in the complicated and diversified environment, the biochemical basis of induced defense in plants is very active and dynamic with intense effect on the stress-causing agents (Kawano 2003). For example, MDA is the final product of membrane lipid peroxidation, which is toxic to cells and inhibits the activity of antioxidant enzymes, therefore MDA is not beneficial to the growth of plants. Biotic and abiotic stresses induce the accumulation of reactive oxygen species (ROS) which are signaling molecules of resistance pathways. But the excessive accumulation of ROS lead to oxidative stress and the production of toxic substances like MDA. In some plants, exogenous SA application contributes to increase the activity of peroxidase (Kawano 2003). In this study, SA treatment reduced the accumulation of MDA to avoid cell damage. In addtion, PAL is considered a necessary enzyme in plant immunity because it catalyzes the biosynthesis of defense related metabolites such as phenolic compounds and lignin. Proline and soluble sugar are important to osmotic regulation and energy metabolism under stresses. Our results showed that exogenous SA promoted the accumulation of proline and soluble sugar, and improved the activity of PAL which were all beneficial to improve plant disease resistance.
Chloroplasts modulate plant defense responses as environmental sensors which become the target of pathogens (Littlejohn et al. 2021). Chloroplast-derived reactive oxygen species (cROS) induced by PTI and ETI lead to lipid peroxidation of cell membrane and HR (Dogra et al. 2019). Chlorophyll, the main pigment in chloroplast, is involved in photosynthesis, and its content can directly affect the intensity of photosynthesis. Pathogen infection can degrade chlorophyll in leaves, and exogenous SA treatment also has influence on chlorophyll content, high concentration SA treatment can inhibit chlorophyll accumulation. In this study, spraying 3 mM SA significantly enhanced the resistance of foxtail millet and improved the content of chlorophyll. These results suggest that SA may protect chloroplasts during the infection of pathogens.
SA also promoted the expression level of defense related genes. SOD and POD encode the key enzymes of antioxidant systems to protect cells against oxidative damage (Nanda et al. 2010). PAL encodes phenylalanine ammonia lyase which catalyzes the biosynthesis of defense related metabolites. The accumulation of endogenous SA was correlated with the induction of PR1 gene expression. The expression of PR1 gene was highly induced in SA treatment that correlated with increased resistance during infection of pathogen (Halder et al. 2019). The qPCR results indicated that the expression level of SiPOD1, SiPOD2, SiSOD, SiPAL1, SiPAL2, and disease resistance related genes (SiPR1) was regulated by the treatment of SA. Exogenous SA treatment can affect PR gene expression, and different concentrations of SA have different effects on it, indicated that the effective induction effect was related to the appropriate concentration of SA. This study found that the expression of SiSOD, SiPOD1 and SiPOD2 were up-regulated after exogenous SA induction, indicating that appropriate concentration of SA treatment can promote the expression of SOD and POD enzyme genes, thereby enhancing the disease resistance of foxtail millet. The change trend of SiPR1 gene was opposite after 1 mM and 3 mM SA treatment, and the expression levels of both decreased in the later stage of treatment; The changes of SiPAL1 and SiPAL2 were consistent. Compared with the control, their expression level showed a trend of decreasing-increasing-decreasing. The gene expression levels were significantly higher at 6-9 h than that of the control. Taken together, exogenous SA upregulated the expression level of these defense related genes at different stage after treatment which helped plant activate the immune system.
Conclusions
In conclusion, SA can induce resistance of foxtail millet to downy mildew. After treatment with appropriate concentration (1 mM or 3 mM) of SA, the incidence of disease was significantly reduced compared with the control. High concentration of SA treatment could not reduce the incidence but significantly reduced the plant height, fresh weight and dry weight of foxtail millet. Appropriate concentration of SA promotes the activation of PAL and the accumulation osmotic regulation substances and chlorophyll content. These enzyme or metabolites protect foxtail millet from cell damage caused by pathogen infection. The expression levels of antioxidant enzyme genes SiSOD, SiPOD1 and SiPOD2 and SiPR1 showed the same trend. This study measured the changes of physiological and biochemical levels after SA treatment, as well as the expression changes of defense related genes. This study provided a theoretical basis for the possibility of using exogenous SA as a substance instead of pesticides and it provided a theoretical basis for the possibility of using SA as a control method for the prevention of downy mildew in the production of foxtail millet.
References
Bawa G, Feng L, Yan L, Du Y, Shang J, Sun X, Wang X, Yu L, Liu C, Yang W, Du J (2019) Pre-treatment of salicylic acid enhances resistance of soybean seedlings to Fusarium solani. Plant Mol Biol 101(3):315–323. doi:https://doi.org/10.1007/s11103-019-00906-x
Bernsdorff F, Döring AC, Gruner K, Schuck S, Bräutigam A, Zeier J (2016) Pipecolic Acid Orchestrates Plant Systemic Acquired Resistance and Defense Priming via Salicylic Acid-Dependent and -Independent Pathways. Plant Cell 28(1):102–129. doi:https://doi.org/10.1105/tpc.15.00496
Bigeard J, Colcombet J, Hirt H (2015) Signaling mechanisms in pattern-triggered immunity (PTI). Mol Plant 8(4):521–539. doi:https://doi.org/10.1016/j.molp.2014.12.022
Dangl JL, Horvath DM, Staskawicz BJ (2013) Pivoting the plant immune system from dissection to deployment. Science 341(6147):746–751. doi:https://doi.org/10.1126/science.1236011
Deenamo N, Kuyyogsuy A, Khompatara K, Chanwun T, Ekchaweng K, Churngchow N (2018) Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora. Int J Mol Sci 19(7). doi:https://doi.org/10.3390/ijms19071883
Devadas SK, Raina R (2002) Preexisting systemic acquired resistance suppresses hypersensitive response-associated cell death in Arabidopsis hrl1 mutant. Plant Physiol 128(4):1234–1244. doi:https://doi.org/10.1104/pp.010941
Ding P, Ding Y (2020) Stories of Salicylic Acid: A Plant Defense Hormone. Trends Plant Sci 25(6):549–565. doi:https://doi.org/10.1016/j.tplants.2020.01.004
Dogra V, Li M, Singh S, Li M, Kim C (2019) Oxidative post-translational modification of EXECUTER1 is required for singlet oxygen sensing in plastids. Nat Commun 10(1):2834. doi:https://doi.org/10.1038/s41467-019-10760-6
Doust AN, Kellogg EA, Devos KM, Bennetzen JL (2009) Foxtail millet: a sequence-driven grass model system. Plant Physiol 149(1):137–141. doi:https://doi.org/10.1104/pp.108.129627
Feng JL, Zhang J, Yang J, Zou LP, Fang TT, Xu HL, Cai QN (2021) Exogenous salicylic acid improves resistance of aphid-susceptible wheat to the grain aphid, Sitobion avenae (F.) (Hemiptera: Aphididae). B Entomol Res 111(5):544–552. doi:https://doi.org/10.1017/s0007485321000237
Halder V, Suliman MNS, Kaschani F, Kaiser M (2019) Plant chemical genetics reveals colistin sulphate as a SA and NPR1-independent PR1 inducer functioning via a p38-like kinase pathway. Sci Rep 9(1):11196. doi:https://doi.org/10.1038/s41598-019-47526-5
Hou S, Man X, Lian B, Ma G, Sun Z, Han L, Yan L, Gao H, Du W, Wang X, Zhang Y, Li H, Han Y (2022) Folate metabolic profiling and expression of folate metabolism-related genes during panicle development in foxtail millet (Setaria italica (L.) P. Beauv). J Sci Food Agric 102(1):268–279. doi:https://doi.org/10.1002/jsfa.11355
Kawano T (2003) Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep 21(9):829–837. doi:https://doi.org/10.1007/s00299-003-0591-z
Kobayashi M, Hiraka Y, Abe A, Yaegashi H, Natsume S, Kikuchi H, Takagi H, Saitoh H, Win J, Kamoun S, Terauchi R (2017) Genome analysis of the foxtail millet pathogen Sclerospora graminicola reveals the complex effector repertoire of graminicolous downy mildews. BMC Genomics 18(1):897. doi:https://doi.org/10.1186/s12864-017-4296-z
Kobayashi M, Utsushi H, Fujisaki K, Takeda T, Yamashita T, Terauchi R (2022) A jacalin-like lectin domain-containing protein of Sclerospora graminicola acts as an apoplastic virulence effector in plant-oomycete interactions. Mol Plant Pathol. doi:https://doi.org/10.1111/mpp.13197
Kulkarni KS, Zala HN, Bosamia TC, Shukla YM, Kumar S, Fougat RS, Patel MS, Narayanan S, Joshi CG (2016) De novo Transcriptome Sequencing to Dissect Candidate Genes Associated with Pearl Millet-Downy Mildew (Sclerospora graminicola Sacc.) Interaction. Front Plant Sci 7:847–847. doi:https://doi.org/10.3389/fpls.2016.00847
Li T, Huang Y, Xu ZS, Wang F, Xiong AS (2019) Salicylic acid-induced differential resistance to the Tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Bio 19(1):173. doi:https://doi.org/10.1186/s12870-019-1784-0
Li R, Han Y, Zhang Q, Chang G, Han Y, Li X, Zhang B (2020) Transcriptome Profiling Analysis Reveals Co-regulation of Hormone Pathways in Foxtail Millet during Sclerospora graminicola Infection. Int J Mol Sci 21(4). doi:https://doi.org/10.3390/ijms21041226
Littlejohn GR, Breen S, Smirnoff N, Grant M (2021) Chloroplast immunity illuminated. New Phytol 229(6):3088–3107. doi:https://doi.org/10.1111/nph.17076
Malamy J, Carr JP, Klessig DF, Raskin I (1990) Salicylic Acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science 250(4983):1002–1004. doi:https://doi.org/10.1126/science.250.4983.1002
Melvin P, Prabhu SA, Anup CP, Shailasree S, Shetty HS, Kini KR (2014) Involvement of mitogen-activated protein kinase signalling in pearl millet-downy mildew interaction. Plant Sci 214:29–37. doi:https://doi.org/10.1016/j.plantsci.2013.09.008
Nanda AK, Andrio E, Marino D, Pauly N, Dunand C (2010) Reactive oxygen species during plant-microorganism early interactions. J Integr Plant Biol 52(2):195–204. doi:https://doi.org/10.1111/j.1744-7909.2010.00933.x
Peng R, Zhang B (2020) Foxtail Millet: A New Model for C4 Plants. Trends Plant Sci 20:S1360–1385. doi:https://doi.org/10.1016/j.tplants.2020.12.003
Porra RJ (2002) The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth Res 73(1–3):149–156. doi:https://doi.org/10.1023/a:1020470224740
Radojičić A, Li X, Zhang Y (2018) Salicylic Acid: A Double-Edged Sword for Programed Cell Death in Plants. Front Plant Sci 9:1133. doi:https://doi.org/10.3389/fpls.2018.01133
Raskin I (1992) Salicylate, a new plant hormone. Plant Physiol 99(3):799–803. doi:https://doi.org/10.1104/pp.99.3.799
Tsuda K, Sato M, Glazebrook J, Cohen JD, Katagiri F (2008) Interplay between MAMP-triggered and SA-mediated defense responses. Plant J 53(5):763–775. doi:https://doi.org/10.1111/j.1365-313X.2007.03369.x
Vlot AC, Dempsey DA, Klessig DF (2009) Salicylic Acid, a multifaceted hormone to combat disease. Annu Rev Phytopathol 47:177–206. doi:https://doi.org/10.1146/annurev.phyto.050908.135202
Yang Z, Zhang H, Li X, Shen H, Gao J, Hou S, Zhang B, Mayes S, Bennett M, Ma J, Wu C, Sui Y, Han Y, Wang X (2020) A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat Plants 6(9):1167–1178. doi:https://doi.org/10.1038/s41477-020-0747-7
Zhang G, Liu X, Quan Z, Cheng S, Xu X, Pan S, Xie M, Zeng P, Yue Z, Wang W, Tao Y, Bian C, Han C, Xia Q, Peng X, Cao R, Yang X, Zhan D, Hu J, Zhang Y, Li H, Li H, Li N, Wang J, Wang C, Wang R, Guo T, Cai Y, Liu C, Xiang H, Shi Q, Huang P, Chen Q, Li Y, Wang J, Zhao Z, Wang J (2012) Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat Biotechnol 30(6):549–554. doi:https://doi.org/10.1038/nbt.2195
Funding
This work was supported by the National Natural Science Foundation of China (32101722 and 32101760), Scientific and Technological Innovation Programs of Higher Education Institutions in Shanxi/ STIP (2021L118), Natural Science Foundation of Shanxi Province (20210302123420 and 20210302123362), Fundamental Research Program of Shanxi Province (20210302123397) and Grand Science and Technology Special Project in Shanxi Province (202101140601027).
Author information
Authors and Affiliations
Contributions
Yaofei Zhao designed this study. Shupei Hou, Zhaowen Liu and Yaqiong Li performed the experiments. Mingzhen Yang and Siyu Hou participated in the data analysis. Yaofei Zhao and Shupei Hou wrote the manuscript. Yuanhuai Han and Zhaoxia Sun reviewed and edited the manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hou, S., Liu, Z., Li, Y. et al. Exogenous salicylic acid enhanced resistance of Foxtail Millet (Setaria italica) to Sclerospora graminicola. Plant Growth Regul 99, 35–44 (2023). https://doi.org/10.1007/s10725-022-00854-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-022-00854-5