
Abstract
In the catfish Heteropneustes fossilis, the anterior kidney is a hemopoietic tissue which surrounds the adrenal homologues, interrenal (IR) and chromaffin tissues corresponding to the adrenal cortical and adrenal medulla of higher mammals. The IR tissue is arranged in cell cords around the posterior cardinal vein (PCV) and its tributaries and secretes corticosteroids. The chromaffin tissue is scattered singly or in nests of one or more cells around the epithelial lining of the PCV or blood capillaries within the IR tissue. They are ferric ferricyanide-positive. Leukemia-inhibitory factor (LIF)-like reactivity was noticed in the lining of the epithelium of the IR cell cords and around the wall of the PCV and blood capillaries. No staining was observed in the hemopoietic cells. IL-1β- and TNF-α-like immunoreactivity was seen in certain cells in the hemopoietic tissue but not in the IR region. Macrophages were identified with mammalian macrophage-specific MAC387 antibodies and are present in the hemopoietic mass but not in the IR tissue. Pigments accumulate in the hemopoietic mass as melano-macrophage centers (MMCs) and are PAS-, Schmorl’s- and Perls’-positive. The pigments contain melanin (black), hemosiderin (blue) and lipofuscin/ceroid (oxidized lipid, yellowish tan), as evident from the Perls’ reaction. The MMCs were TUNEL-positive as evident from FITC fluorescence, indicating their apoptotic nature. The MMCs showed significant seasonal variation with their density increasing to the peak in the postspawning phase. Melanins were characterized spectrophotometrically for the first time in fish anterior kidney. The predominant form is pheomelanin (PM), followed by eumelanin (EM) and alkali-soluble melanin (ASM). Melanins showed significant seasonal variations with the level low in the resting phase and increasing to the peak in the postspawning phase. Under in vitro conditions, lipopolysaccharide (10 µg/mL) treatment increased significantly the levels of PM and EM levels both at 16 and at 32 h and the ASM level at 32 h. On the other hand, the synthetic glucocorticoid dexamethasone (100 nM) decreased significantly the levels of EM, PM and ASM time-dependently. The results indicate that the anterior kidney is an important site of immune–endocrine interaction.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Unlike higher vertebrates, fishes lack bone marrow and lymph nodes, which are the primary and secondary immune organs, respectively. The kidney, which is divisible into an anterior pronephric head kidney (HK) and a posterior mesonephric renal kidney or trunk kidney, is an important lympho-myeloid or hemopoietic organ in fishes (Sailendri and Muthukkaruppan 1975; Ellis 1977; Fange 1986; Meseguer et al. 1994; Press and Evensen 1999; Fijan 2002; Zapata et al. 2006; Abdel-Aziz et al. 2010a). The HK monocytes/macrophages are important components of fish innate immune system and have a major phagocytic role in trapping the pathogens, foreign particles and substances in the blood and in the clearance of the debris and effete cells (Secombes and Fletcher 1992; Seljelid and Eskeland 1993; Fishelson 2006; Kemenade et al. 2009). Moreover, the macrophages release pro-inflammatory cytokines responsible for the initiation of host’s immune responses.
In the HK and other lympho-myeloid organs (trunk kidney, spleen, thymus, peripheral part of liver, etc.), macrophages are pigmented (melano-macrophages) and distributed singly or in clusters (except in salmonids), forming melano-macrophage centers (MMCs; Agius 1985; Herraez and Zapata 1991; Meseguer et al. 1994; Press and Evensen 1999; Matsche and Grizzle 1999; Wolke 1992; Agius and Roberts 2003; Vigliano et al. 2006; Jordanova et al. 2008, 2012; Balamurugan et al. 2012). The MMCs undergo changes related to breeding status (Jordanova et al. 2008; 2012; Kumar and Joy 2015). The pigmented macrophages are capable of synthesizing melanins, like melanocytes and the melanins absorb and neutralize various toxins like free radicals and cations (Zuasti et al. 1989; Vigliano et al. 2006; Ribeiro et al. 2011). The MMCs are differentially stained with Perls’ solution and contains pigments such as melanin, hemosiderin and lipofuscin/ceroid (oxidized lipid). Melanin stains black, hemosiderin stains blue and lipofuscin/ceroid gives a yellowish tan (Besseau and Faliex 1994; Meseguer et al. 1994; Blazer 2002; Lei et al. 2012; Diaz-Satizabal and Magor 2015). The nature of the pigments varies with the organ, species and functional state, water quality, stressor, etc. Melanins can be classified broadly into two major groups: eumelanin and pheomelanin (Ito et al. 2000). Eumelanin is composed of brown to black pigment, whereas pheomelanin is yellow to red in color. Melanin is considered as derived from exogenous source or may be generated within the phagocytosed cell (Franco-Belussi et al. 2013). There are different views regarding the origin of the MMCs such as necrosis, autophagy and apoptosis (Hughes and Gorospe 1991; Tilly et al. 1991; Guthrie et al. 1995; Janz and Kraak 1997; Wood and Van Der Kraak 2001; Santos et al. 2008; Ucuncu and Cakici 2009).
The HK encloses the adrenal homologue in teleosts. The adrenal homologue is located around the main postcardinal vein (PCV) and its tributaries and consists of interrenal (IR) and chromaffin tissues (Nandi 1962; Lofts and Bern 1972; Butler 1973; Chester et al. 1986; Abelli et al. 1996; Milano et al. 1997; Crivellato et al. 2006; Abdel-Aziz et al. 2010b). The IR tissue secretes corticosteroids, while the chromaffin tissue synthesizes catecholamines (Lofts and Bern 1972; Chester et al. 1986; Fabbri et al. 1998; Montpetit and Perry 2002).
The anatomical coexistence of the immune and endocrine components in the HK permits interactions between the two important communication systems, the immune system and the hypothalamus–pituitary–adrenal (HPA) axis (stress axis; Engelsma et al. 2002; Holland et al. 2002; Metz et al. 2006, Castellana et al. 2008; Kemenade et al. 2009, 2011). Monocytes/macrophages together with other lymphocytes produce cytokines like interleukin-1β (IL-1β) and tumor necrosis factor (TNF-α), which are pro-inflammatory cytokines (Tesch et al. 1997; Holland et al. 2002; Tkachenko et al. 2011). Though several studies are available on fish macrophage isolation, activation and functions in vitro (Sorensen et al. 1997; Braun-Nesje et al. 1982; Zou et al. 2000; Pelegrin et al. 2004; Sarmento et al. 2004; Datta et al. 2009; Kemenade et al. 2011), few studies have attempted the cellular localization in the HK (Romano et al. 1998; Mulero et al. 2008). Although the production of corticosteroids by the adrenal is primarily regulated by the HPA axis, several cytokines influence their production, which are available locally or come from outside sources, either by infiltrating macrophages or by the tissue itself (Tkachenko et al. 2011). Though IL-1β and TNF-α are mostly produced at the site of inflammation or tissue injuries, these are also produced in the steroid-producing adrenocortical cells and macrophages, suggesting a possible role in the activation of the HPA axis (Gonzalez-Hernandez et al. 1995). In human adrenal cortex, steroidogenesis is regulated by leukemia-inhibitory factor (LIF) and its receptor (LIF-R), inducing cortisol secretion and decreasing androgen release (Bamberger et al. 2000; Woods et al. 2008). LIF is a pleiotropic cytokine belonging to the IL-6 family that includes LIF, IL-6 and IL-11 and shares a common gp130 receptor subunit with IL-6.
Though the adrenal homologue of the catfish Heteropneustes fossilis, an air breathing freshwater species, was described previously (Joy 1984), there are no reports on the cellular details of the hemopoietic tissue. An understanding of the immune cells in the head kidney is important to study potential interaction between the endocrine and immune tissues. Keeping the above in mind, we have described the morpho-histology of the HK using histological and histochemical methods. Further, using heterologous antibodies, we describe MAC387, LIF, IL-1β and TNF-α-like immunoreactivities in the hemopoietic tissue. The MMCs were identified by histochemical procedures, and the seasonality in the MMC was determined. Apoptotic nature of the MMCs was demonstrated by TUNEL assay. The HK melanins were characterized biochemically, and assays were used to quantify melanins for the first time in fish kidney. Effects of lipopolysaccharide (immune response) and dexamethasone (endocrine response) on melanin contents were determined under in vitro conditions. Based on the results, the potential interaction(s) between the adrenal homologues and hemopoietic tissue was discussed.
Materials and methods
Chemical and reagents
Antibodies of LIF (M-179): sc-20087, IL-1β (C-20): sc-1250 and TNF-α (52B83): sc-52746 were purchased from Santa Cruz Biotechnology Inc, Europe. Mouse monoclonal antibody to macrophage (MAC387) was purchased from Abcam, Inc, USA. Vectastain ABC kit for immunohistochemistry was purchased from Burligame, CA, USA. TUNEL assay kit was purchased from BioVision, Milpitas, USA. Sepia melanin, lipopolysaccharide (Escherichia coli, 0127:B8), dexamethasone and (3-aminoproyl)triethoxysilane were purchased from Sigma-Aldrich, New Delhi, India. Leibovitz L-15 medium and syringe-driven filter (nylon hydrophilic membrane, pore size 0.45 µm, 30 mm diameter) were purchased from Hi Media Laboratories Pvt. Ltd (Mumbai, India). Leishman’s stain and potassium ferrocyanide were purchased from Qualigens fine chemicals, Navi Mumbai. Xylene, H2O2, sodium hydroxide pellets, sodium acetate and ethanol were purchased from Merck Specialities Private Limited, Worli, Mumbai, India. Hydroiodic acid was purchased from Spectochem Private Limited, Mumbai. 3,3-Diaminobenzidine tetrahydrochloride and tris-hydrochloride were purchased from Himedia. Sodium chloride was purchased from Ranchem (RFCL Limited, New Delhi, India). Potassium ferricyanide was purchased from Galxo Laboratories Limited India, Mumbai.
Animal
The experiments were conducted as per the guidelines of the Animal Ethics Committee of Banaras Hindu University. Care was taken to prevent cruelty of any kind. Adult female Heteropneustes fossilis (30–40 g) were collected from a local fish market in resting (December–January), preparatory (February–April), prespawning (May–June), spawning (June–July) and postspawning (September–November) phases of the annual reproductive cycle. They were maintained in the laboratory for 48 h under normal photoperiod and temperature (resting phase—10.5 L:13.5 D; preparatory phase—12.5 L:11.5 D; prespawning phase—13 L:11 D; spawning phase—13 L:11 D; postspawning—12.5 L:11.5 D and ambient temperature 22.5 ± 2 °C) to overcome stress due to transportation and fed daily with egg white and boiled liver ad libitum.
Morpho-histology of kidney
The fish were weighed and length measured. They were killed by decapitation without disturbing the head kidney. The viscera containing digestive tract, ovary, spleen, heart and fat body were carefully removed to expose the kidney. The transparent peritoneal membrane covering the posterior kidney was removed. The anterior, posterior and lateral kidneys were cleared from the adjoining tissue. Photographs of the ventral view of the fish body cavity were taken by a digital Sony camera DSC-HX10V with 18.2 megapixels. The kidneys were removed and fixed in Bouin’s fluid for 24 h and processed for paraffin embedding. Serial transverse and horizontal sections were cut at 6 µm and stained with Ehrlich’s hematoxylin–eosin and Mallory’s trichrome stain.
Cell suspension
A single cell suspension of the anterior kidney was made by pressing the anterior kidney against toothed cleaned forceps or by vigorous pipetting in phosphate-buffer saline (PBS, pH 7.4). The cell suspension was fixed in 4 % paraformaldehyde in PBS (pH 7.4) buffer for 20 min at room temperature. A smear of single cell suspension was made on clean slides. Slides were allowed to dry in an incubator at 40 °C overnight. The smear was stained with Leishman’s stain for 10 min, rinsed in distilled water and mounted. The stained smear was examined under a Leica DM 2000 microscope, and images were taken with a Leica digital camera DFC 295 with 3 megapixel.
Identification of chromaffin cells
The anterior, posterior and lateral kidneys were removed and fixed in Orth’s fluid for 4 h and embedded in paraffin. Serial sections were taken at 6 µm. The sections were deparaffinized and hydrated using different downgrades of ethanol. The hydrated sections were immersed for 10–15 min in freshly prepared ferric ferricyanide reagent for Schmorl’s reaction. Thirty milliliters of 1 % ferric chloride and 10 mL of freshly prepared 1 % potassium ferrocyanide were mixed to form the ferric ferricyanide solution (Bancroft and Gamble 2008). The sections were dehydrated in ethanol, cleared in xylene and mounted in DPX. Photographs were taken, as described above.
Identification of melano-macrophage centers
The kidneys were fixed in Bouin’s fluid for 24 h and embedded in paraffin. Serial sections were taken at 6 µm, deparaffinized and hydrated. The sections were used for Ehrlich’s hematoxylin–eosin, PAS or Perls’ staining (Bancroft and Gamble 2008). Perls’ solution was prepared by mixing 2 % potassium ferrocyanide and 2 % HCl in equal ratio. For Perls’ reaction, hydrated sections were immersed for 10–15 min in the Perls’ solution. For PAS staining, serial hydrated sections were oxidized with periodic acid for 10 min. Periodic acid solution was prepared in 0.8 % periodic acid and 0.27 % sodium acetate in 70 % ethyl alcohol. The sections were kept in 70 % alcohol for 5 min and treated with Schiff’s reagent for 10 min, followed by washing in water. The sections were differentiated with 0.5 % of sodium metabisulfite solution for 2 min and dehydrated in different grades of ethanol and mounted in DPX. Photoimages were taken, as described above.
Immunohistochemistry
The head kidneys were excised and fixed in Bouin’s fluid without acetic acid and processed for paraffin embedding. Sections were taken at 6 µm and were mounted on gelatin-coated slides. The sections were deparaffinized and hydrated using different downgrades of ethanol and then washed in phosphate-buffer saline (PBS) buffer (pH 7.4) for 15 min. Immunohistochemistry was performed according to the Vectastain ABC kit with some modifications, as mentioned below. The hydrated sections were immersed in 0.3 % H2O2 in PBS (pH 7.4) for 30 min at room temperature to remove endogenous peroxidase activity, for elimination of non-specific binding. This step was done in a dark humid chamber. The sections were rinsed with PBS three times and then blocked with blocking serum at room temperature for 1 h. The sections were washed with PBS three times and incubated with primary antibodies: MAC387 (1:50), LIF (1:200), IL-β (1:100) or TNF-α (1:50) at 4 °C overnight in a closed humid chamber. After the sections were rinsed in PBS three times, biotinylated HRP conjugated-secondary antibody was applied to the sections for 1 h at room temperature, followed by washing with PBS three times. The sections were incubated with streptavidin–biotin horseradish peroxidase complex for 30 min at room temperature. The sections were washed with PBS and treated with diaminobenzidine (DAB) substrate solution. When sufficient color intensity was developed, the sections were washed and dehydrated with various grades of alcohol. The sections were cleared in xylene and then mounted with DPX. The sections were examined under a Leica DM 2000 microscope, and images were taken.
As control, the immunostaining was done without the primary antibody or in sections over-heated.
TUNEL assay
The head kidneys were fixed in 4 % paraformaldehyde (PFA) for 24 h and embedded in paraffin. Serial sections were taken at 6 µm and affixed on silane-coated slides. TUNEL assay was performed according to the Apo-BrdU DNA fragmentation assay kit (BioVision, Milpitas, USA) with minor modifications. Deparaffinized sections were hydrated in downgrades of ethanol. The hydrated sections were immersed in 0.85 % NaCl for 5 min at room temperature and washed with PBS (pH 7.4) for 5 min at room temperature. The sections were again fixed in 4 % PFA for 15 min at room temperature. The slides were washed in the PBS buffer two times for 5 min at room temperature and allowed to drain out completely the buffer. The sections were incubated with 100 µL of freshly prepared 20 µg/mL proteinase K solution (prepared in 100 mM tris–HCl, pH 8.0) for 5 min at room temperature. The slides were immersed in the PBS buffer for 5 min and again incubated with 4 % PFA for 5 min at room temperature. The sections were washed in the PBS buffer for 5 min. Excess fluid was gently removed, and the sections were incubated two times with 100 µL of the buffer and covered by a coverslip for homogenous spread of the solution over the sections for 5 min. The coverslip was removed carefully, and the sections were incubated with 50 µL of the DNA labelling solution, covered and incubated for 60 min in a 37 °C humidified chamber in dark. After the incubation, the coverslip was removed carefully and the slides were washed with the PBS buffer two times for 5 min in a Coplin jar. The sections were blot-dried around the edges with tissue paper. Ant–BrdU-FITC complex was diluted in the rinse buffer (1 part of FITC antibody and 20 part of rinse buffer), and the sections were incubated with 100 µL of the antibody solution after covering. The slides were incubated in a dark humidified incubator for 30 min at room temperature. The coverslip was removed carefully, 100 µL of propidium iodide (PI) solution was added over the sections and covered, and the sections were incubated for 30 min in the dark in a humidified incubator. The coverslip was carefully removed and the sections were washed in triple-distilled water for 5 min (two times). The sections were mounted in glycerol and kept at −20 °C overnight and observed under a Carl Zeiss LSM 780 confocal microscope (ISLS, BHU), and images were taken. The images were analyzed by Zen 2012 SP1 (black edition, 64 bits) software that controlled the scanning module, laser module and image acquisition process.
Experiments
Seasonal variations in MMC
In different reproductive phases, the acclimated fish (n = 5 per phase) were killed by decapitation and anterior kidneys were sampled, fixed in Bouin’s fixative for 24 h and embedded in paraffin. Serial sections of the whole kidney were taken at 6.0 µm, end to end. A total of eight slides were made from each tissue and numbered from the start to the end section of the anterior kidneys. Each slide contained 24–28 sections arranged in two rows in the serial order. The sections were stained with the Schiff’s reagent, as described above. MMCs were PAS-positive (pink color). The macrophage aggregates were counted in all sections of all the eight slides. An outline diagram of each section was made on a paper, and a dot was marked on the diagram for each MMC that was counted so that an accurate counting was ensured. Care was taken to avoid repetition in the counting of the MMCs. The procedure was repeated for different reproductive phases.
In vitro experiments with dexamethasone and lipopolysaccharide
Leibovitz L-15 medium was supplemented with 10 % fetal bovine serum and 100 µg/mL streptomycin. The medium was filtered in a sterilized syringe-driven filter having a nylon hydrophilic membrane of pore size 0.45 µm and 30 mm diameter. Fish were dissected, and anterior kidneys were collected in PBS (pH 7.4). The head kidney (20 mg; n = 5) in duplicate was incubated in the L-15 medium for 24 h for conditioning in a CO2 incubator at 20 °C. For experiments, 1 mg of dexamethasone (a synthetic glucocorticoid) was dissolved in 50 μL of ethanol, then diluted in propylene glycol to make up 1 mL solution and further diluted to make 100 nM working solution. After 24-h conditioning, 100 nM of dexamethasone was added to the medium containing the head kidney. The control groups were incubated in plain medium or the medium containing ethanol plus propylene glycol. After the incubation, the anterior kidneys were collected at 8 and 16 h and processed for melanin assays. Similarly, 1 mg of lipopolysaccharide (LPS) was dissolved in PBS (pH 7.4) and anterior kidneys were incubated for 8, 16 and 32 h with 10 µg/mL LPS. A longer LPS exposure was given since LPS is generally less sensitive in fish (Pelegrin et al. 2004). Control groups were incubated in plain medium in a CO2 incubator at 20 °C. After the incubation, the anterior kidneys were collected and processed for melanin assays.
Assay of eumelanin, pheomelanin and alkali-soluble melanin
Melanins were measured by the spectrophotometric methods of Ozeki et al. (1995) and Fan et al. (2010), as described earlier (Kumar and Joy 2015). Sepia melanin was used as the standard for analysis of eumelanin (EM), pheomelanin (PM) and alkali-soluble melanin (ASM). A suspension of sepia melanin in water was prepared at a concentration of 1 mg/mL by sonicating for 5 min to make a homogenous suspension.
Sample preparation
In each phase, anterior kidneys were collected and 4–6 fish HK were pooled to make samples (n = 5). Twenty milligrams of tissue was taken for melanin estimation. The samples were sonicated on ice in 500 μL of PBS at pH 7.4. The homogenate was centrifuged at 5000 rpm for 5 min at 4 °C. Supernatant was used for the assay of melanins.
Assay of eumelanin (EM)
Fifty microliters of supernatant was hydrolyzed in 450 μL of hot 30 % hydroiodic acid (HI). After the HI hydrolysis, melanin was solubilized in hot 100 μL of 1 M NaOH and 15 μL of 1.5 % H2O2. The mixture was cleared by centrifugation at 10,000 rpm for 5 min at 4 °C, and the supernatant was analyzed for absorbance at 350 nm (A 350) in a microplate reader (EPOCH, BioTek Instruments Inc, USA).
Assay of pheomelanin (PM)
One hundred microliters of supernatant was solubilized in 400 μL PBS buffer (pH 10.5) for 10 min and mixed for 10 min at room temperature. The mixture was centrifuged at 10,000 rpm for 10 min at 4 °C, and supernatant was collected. An equal volume of chloroform was added to the supernatant and mixed by vortexing. Pale yellow colored aqueous phase supernatant was collected in fresh tubes by centrifugation at 4000 rpm for 10 min at 4 °C to remove all impurities. The process was repeated again to remove all the trace impurities by centrifuging at 10,000 rpm for 10 min at 4 °C, and the supernatant containing pheomelanins was analyzed for absorbance at 400 nm (A 400) in the microplate reader.
Assay of alkali-soluble melanin (ASM)
This method is very similar to the assay of pheomelanin. Fifty microliters of supernatant was dissolved in 8 M urea/1 M NaOH solution which was prepared by dissolving 8 M urea in 1 M NaOH. Pale brown colored aqueous phase was analyzed for absorbance at 400 nm (A 400) in the microplate reader.
Calculation
Standard curves were prepared for EM, PM and ASM by taking different concentrations of sepia melanin and processed as the tissue samples. The standard curves were linear over the concentrations used. The minimum detection limit was 2 μg. Using the standard graphs, concentrations of melanins were measured in the samples and expressed in μg/mg tissue weight.
Statistical analysis
The data were expressed as mean ± SEM and were analyzed by one-way analysis of variance (ANOVA), followed by Newman–Keuls test (p < 0.05) for multiple group comparisons.
Results
Morphology of the kidney
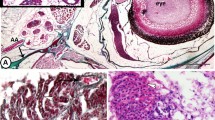
In the catfish, the kidney is divided into two major divisions, the anterior head kidney (HK) and posterior trunk kidney (TK), separated by a bony transverse septum (TS). The HK is a paired triangular mass, lodged at the angle of the T-shaped TS (Fig. 1a). The TK is an elongated paired mass, approximated together to form a single structure, extending throughout the peritoneal cavity dorsally. From the anterior end of the TK arises a pair of extension laterally, which forms the lateral kidney (LK). The three divisions of the kidney have distinct anatomical features. The HK is a hemopoietic mass enclosing the adrenal homologue (Fig. 1b) around the postcardinal vein (PCV) that runs through it. There are no renal tubules per se. The LK is composed of hemopoietic tissue and fewer renal tubules (Fig. 1c). In the hemopoietic tissue, aggregations of pigments (MMCs) can be seen (Fig. 1c). The TK consists of a large number of renal tubules and is the functional kidney (mesonephros). The renal tubules are embedded within the hemopoietic tissue that is beset with the MMCs (Fig. 1d).

a A ventral view of the body cavity of female catfish H. fossilis showing anterior head kidney, bony transverse septum, lateral kidney and the posterior trunk kidney. b A section of the anterior kidney showing postcardinal vein (PCV), interrenal tissue (IR) and hemopoietic tissue (HT). c A section of the lateral kidney showing lymphoidal mass, melano-macrophage centers (MMCs) and renal tubules. d A section of the posterior kidney showing MMCs and renal tubules. HK head kidney, P1 first proximal segment of the renal tubules. Ehrlich’s hematoxylin–eosin staining
Structure of the head kidney
The HK consists of hemopoietic, interrenal and chromaffin tissues (Fig. 2a, b). The IR tissue is surrounded by the hemopoietic tissue (HT), as shown in Figs. 1b and 2a. The IR tissue is formed of cell cords around the PCV and its branches (Fig. 2a). The PCV is filled with blood cells usually round or oval with prominent spherical nuclei having homogenous eosinophilic cytoplasm. Bordering the PCV and within the IR are nests of chromaffin cells (Fig. 2b). They are present singly or in nests and have round, oval or elongated contour (Fig. 2b). These cells are chromophobic in hematoxylin–eosin preparations (Fig. 2a). The chromaffin cells were distinguished from the other cell mass with the ferric ferricyanide reagent, which produced a dark Prussian blue color (Fig. 2b). The hemopoietic tissue that surrounds the IR tissue contains a large number of erythrocytes and leukocytes. These cells are not arranged in a specific pattern but are present throughout the hemopoietic tissue. Blood sinuses can be seen extensively in the hematopoietic tissue.

Anterior kidney of female catfish H. fossilis stained with Ehrlich’s hematoxylin–eosin (a, c) showing the arrangement of the interrenal tissue (IR) and chromaffin cells (CC, arrows). The MMCs (yellow brown colored groups of cells) are present in the hemopoietic tissue (HT). b The head kidney stained with ferric ferricyanide showing CC (arrows) around the PCV and small blood capillaries. d Stained with ferric ferricyanide showing the MMCs in the hemopoietic tissue. Note the presence of many scattered dark stained melano-macrophages (arrows). e, f Stained with Perls’ solution and PAS, respectively, showing the MMCs in the HT. (Color figure online)
Melano-macrophage centers (MMCs)
The MMCs were seen within the hemopoietic tissue as discrete groups of cells and stained differentially. In the hematoxylin–eosin staining, the MMCs appeared as yellow brown pigment masses (Fig. 2c). The MMCs could be easily distinguished in Perls’ solution (Fig. 2e) having blue or greenish blue color or by PAS staining as pink magenta aggregates within the hemopoietic tissue. In the ferric ferricyanide reaction, the MMCs gave a Prussian blue color, similar to the chromaffin cells. Isolated pigmented macrophages were also seen close to the MMCs or away from them (Fig. 2d). An examination of the serial sections showed that these cells seem to be directed (migrating) toward the MMCs (Fig. 2d). The MMCs were small or large, and the larger macrophage aggregates were ovoid or irregular in shape.
Cellular composition of the hemopoietic tissue
A single cell suspension of the HK revealed different types of hemopoietic cell population after Leishman’s staining (Fig. 3). The cell types include macrophages, granulocytes, erythrocytes, neutrophils and basophils.

A smear of the single cell suspension of anterior head kidney stained with Leishman’s stain showing erythrocytes, macrophages, neutrophils, granulocytes, monocytes and basophils
Localization of macrophages
There was no staining in the control sections (Fig. 4a) when MAC387 primary antibody was omitted or the sections were over-heated prior to immunostaining. The immunostaining with the MAC387 resulted in staining of cells scattered in the hemopoietic tissue (Fig. 4b). The cells are evenly distributed in the HK and are spindle, round or irregular in morphology. Some cells have kidney-shaped nucleus. Apparently, the MMCs did not stain for the antibody. There was no immunostaining in the IR tissue or chromaffin tissue.

MAC387-like (b), IL-1β-like (d) and TNF-α-like (f) immunoreactivity in the HT cells using the respective primary antibodies. a, c, e The respective controls
Localization of IL-1β and TNF-α
IL-1β-like (Fig. 4d) and TNF-α-like (Fig. 4f) immunoreactivity was found in the cells in the hemopoietic tissue. The IL-1β-like staining was noticed in heterogenous populations like monocytes, macrophages and even granulocytes. They are round, oval or irregular in shape. The TNF-α-like cells are largely irregular, spindle or oval-shaped. The staining was not seen in the IR tissue. The control sections did not show any cell staining (Fig. 4c, e).
Localization of LIF-reactive elements
There was no staining in the control reaction (Fig. 5a). The LIF-like immunoreactivity was noticed on the cell cords in the IR tissue region (Fig. 5b, c). The immunoreactivity was found in the connective tissue, lining the IR cell cords, which is clearly shown in the Mallory staining (Fig. 5d). The IR cells per se were not immunoreactive. The hemopoietic tissue cells did not show any positive immunoreactivity (Fig. 5a, b). Strong LIF-like immunoreactivity was found in the wall of the PCV (Fig. 5e) in the IR tissue, comparable with the Mallory staining (Fig. 5f), and blood vessels in the HT (Fig. 5b).

LIF-like immunoreactivity in the head kidney. a The control staining. b LIF-like immunoreactivity within the IR region. The LIF-like activity was seen around the epithelial lining of the IR cell cords (c). d A Mallory-stained comparable section showing the IR cell cords. An intense immunolocalization was also seen around the epithelial lining of the PCV (e) and blood capillaries in the HT (b). f A comparable section stained with Mallory’s triple staining showing the wall of the PCV. IL-1β interleukin-1β, TNF-α tumor necrosis factor α, LIF leukemia-inhibitory factor
TUNEL assay
The nuclei were counterstained with propidium iodide, which gave a red fluorescence (Fig. 6a, d, g). The MMCs in the hemopoietic tissue were TUNEL-positive, giving a green to yellow FITC florescence. The MMCs were seen in aggregates (Fig. 6b, e) or scattered (Fig. 6h) in the hemopoietic mass. The interrenal (IR) cells were not TUNEL-positive (Fig. 6b, c). When merged, the TUNEL immunoreactivity was observed in the nuclei of the cells of the MMCs (Fig. 5c, f, i).

Sections of head kidney of H. fossilis stained with propidium iodide (PI, left lane), TUNEL (middle lane) and the two merged (right lane). Arrows show FITC-antibody fluorescence in melano-macrophage centers (MMCs) and scattered cells in the hemopoietic tissue (b, e, h, c, f, i). The interrenal (IR) cells did not show any reaction (b, c)
Seasonal changes in MMCs
The MMCs showed a significant seasonal variation (Fig. 7; p < 0.001, one-way ANOVA; F = 32.61). The MMCs were less in number in the resting and preparatory phases and increased, through the prespawning and spawning phases, to the peak in the postspawning phase. The MMC density did not change significantly between the resting and preparatory phases and between the prespawning and spawning phases. The MMC density in the prespawning and spawning phases was significantly higher than that of the resting and preparatory phases. In the postspawning phase, the MMC count increased significantly higher than all other phases (p < 0.05, Newman–Keuls test).

Seasonal changes in the MMCs in the head kidney of the catfish H. fossilis during different reproductive phases. Values are mean ± SEM of 5 fish each. Data were analyzed by one-way ANOVA (p < 0.001) and Newman–Keuls test (p < 0.05). Groups bearing the same letter are not significantly different, and those bearing different letters are significantly different
Seasonal changes in melanins
In the head kidney, different forms of melanins were identified by the spectrophotometric assay (Fig. 8). The major form of melanin is pheomelanins (PM), followed by eumelanins (EM) and total alkali-soluble melanins (ASM). The melanins showed significant seasonal variations (p < 0.001, one-way ANOVA; F = 32.25 for EM, F = 21.324 for PM, and F = 9.072 for ASM). The concentration of EM was low in the resting phase and increased through preparatory, prespawning and spawning phases to give the peak level in the postspawning phase. The EM levels are not significantly different in the resting, preparatory and prespawning phases. The levels in the spawning and postspawning phases are significantly different and are also different from other phases (p < 0.05, Newman–Keuls test). The pheomelanin concentration was low in the resting phase and increased steadily to the peak value in the postspawning phase. The levels in the resting, preparatory and prespawning phases are not significantly different. The spawning and postspawning levels are significantly different with each other and also from the levels in other phases. The concentration of ASM was low in the resting phase and increased in the successive phases. The levels in the preparatory, prespawning and spawning phases are not significantly different. The ASM level showed the peak increase in the postspawning phase, which is significantly different from all other values.

Seasonal changes in different types of melanins in the head kidney of the catfish H. fossilis during different reproductive phases. Values are mean ± SEM of 5 fish each. Data were analyzed by one-way ANOVA (p < 0.001) and Newman–Keuls test (p < 0.05). Groups bearing the same letter or number are not significantly different, and those bearing different letters or numbers are significantly different. EM eumelanin, PM pheomelanin, ASM alkali-soluble melanin
In vitro effects of LPS and dexamethasone on melanins
The incubation of the HK with 10 µg/mL of LPS increased significantly the concentrations of EM and PM (Fig. 9; p < 0.001, one-way ANOVA; p < 0.05, Newman–Keuls test) at both 16 and 32 h. The ASM level showed a significant increase only at 32 h. Dexamethasone decreased significantly the concentrations of EM, PM and ASM (Fig. 10; p < 0.001, one-way ANOVA; p < 0.05, Newman–Keuls test) at both 8 and 16 h.

In vitro effects of lipopolysaccharide (LPS) on melanin levels in the head kidney of the catfish H. fossilis. Values are mean ± SEM of 5 fish each. Data were analyzed by one-way ANOVA (p < 0.001) and Newman–Keuls test (p < 0.05). Groups bearing the same letters or numbers are not significantly different, and those bearing different letters or numbers are significantly different

In vitro effects of dexamethasone on melanin levels in the head kidney of the catfish H. fossilis. Values are mean ± SEM of 5 fish each. Data were analyzed by one-way ANOVA (p < 0.001) and Newman–Keuls test (p < 0.05). Groups bearing the same letters or numbers are not significantly different, and those bearing different letters or numbers are significantly different
Discussion
In the catfish Heteropneustes fossilis, the kidney consists of the main renal posterior kidney in the trunk region with paired elongate lateral lobes behind the transverse septum and a pair of triangular head kidney anterior to the transverse septum that contains the adrenal homologues. In an earlier study in the African catfish Clarias gariepinus, Vermeulen et al. (1995) also recognized the three regions. However, the authors named the lateral lobe as head kidney and the anterior paired lobes as ‘adrenal-like organ.’ In a majority of fish, the anterior structure has been described as the head kidney (Nandi 1962). In H. fossilis, the anterior lobes contain the adrenal homologue surrounded by the hemopoietic tissue, the lateral lobes contain fewer renal tubules and hemopoietic tissue, and the posterior kidney contains a large number of renal tubules embedded within the hemopoietic tissue.
The adrenal homologues
The IR and chromaffin tissues were described in a large number of teleosts (Nandi 1962) and the arrangement of the tissues deviates greatly from their mammalian counterparts (Chester et al. 1986). The IR tissues were described in the catfish C. batrachus (Joy and Sathyanesan 1978), H. fossilis (Joy 1984) and C. gariepinus (Vermeulen et al. 1995). The steroid-producing property of the IR tissue has been demonstrated histochemically, histoenzymologically, ultrastructurally and biochemically in the catfishes (reference as above) and in other teleosts (Milano et al. 1997; Civinini et al. 2001; Rocha et al. 2001; Abdel-Aziz et al. 2010b). In the African catfish, cortisol, 5α- and 5β-androstanedione and androstenedione are the abundant steroids secreted by the IR tissue (Vermeulen et al. 1995). In H. fossilis, cortisol is secreted by the head kidney (unpublished data).
In the catfish, the chromaffin cells occur singly or in groups around the wall of the PCV or around the blood vessels among the IR cell cords. The chromaffin cells react positively to ferric ferricyanide reagent (Schmorl’s reaction), dichromate (adrenaline and noradrenaline cells), iodate (noradrenaline cells) and ammonium silver nitrate (noradrenaline cells; Joy and Sathyanesan 1978; Joy 1984). Two types of chromaffin cells, noradrenaline and adrenaline cells, were identified ultrastructurally and immunohistochemically in the HK and opisthonephros in sea bass Dicentrarchus labrax (Abelli et al. 1996); stickleback Gasterosteus aculeatus (Gallo et al. 1993) and Salmo gairdneri (Mastrolia et al. 1984). The chromaffin cells synthesize, store and release catecholamines, adrenaline and noradrenaline (Reid et al. 1998; Montpetit and Perry 2002).
The IR and chromaffin cells in the catfish did not show any positive reaction for MAC387, IL-1β and TNF-α antibodies. This is in contrast to the situation in the adrenal cortex of higher vertebrates. Macrophages were described in the zona fasciculata and zona reticularis, which secrete cytokines such as IL-1, IL-6, TNF-α. The cytokines, in turn, stimulate the secretion of corticosteroids locally or by activating the HPA axis (Bornstein et al. 2004; Guzman et al. 2010). However, both LPS and IL-1β are shown to stimulate cortisol secretion in rainbow trout (Holland et al. 2002). It is likely that cytokines present in the hemopoietic tissue of the HK may be involved in the control of cortisol production.
On the other hand, the adrenal homologue showed strong immunoreactivity for LIF antibody. This is the first report on the demonstration of LIF-like immunoreactivity in the head kidney of a fish. The reactivity was found in the connective tissue elements in the PCV and blood vessels and around the IR cell cords. The blood vessels in the hemopoietic tissue also showed strong immunoreactivity. The connective tissue nature of the LIF staining was indicated by the Mallory triple staining. A cDNA for LIF was isolated in zebrafish that codes for a 215 amino acid protein (Abe et al. 2007). LIF is a pleiotropic cytokine and a member of IL-6 subfamily and maintains stem cells in an undifferentiated state. LIF and LIF-receptor expression was reported in different cells of the pituitary including corticotropes (Auernhammer and Melmed 2000). It stimulates ACTH secretion. In the human and bovine adrenal, LIF was localized in the cytoplasm of the adrenocortical cells in all the zones and LIF-receptor was localized in both cytoplasm and membrane (Bamberger et al. 2000; Woods et al. 2008). LIF stimulates both cortisol and aldosterone in cell cultures. LIF modulates the HPA axis at the level of the both pituitary and adrenal in a paracrine/autocrine manner in response to inflammation (Chesnokova and Melmed 2000). The LIF-like staining around the wall of the blood vessels suggests that it may be involved in hematopoiesis. The LIF-like staining in the connective tissue wall of the IR cords may suggest a paracrine role for LIF in the control of steroidogenesis.
Hemopoietic tissue
As reported previously, the HK hemopoietic tissue contained erythrocytes, macrophages, monocytes, neutrophils and basophils. Although not quantified, macrophages are a major population of the hemopoietic tissue and were identified by the MAC387 antibody which recognizes both macrophages and monocytes. There are limited studies on the demonstration of macrophages in the HK (Romano et al. 1998; Mulero et al. 2008). Both IL-1β- and TNF-α-like immunoreactivity was observed in cells, suggesting that these cells may secrete cytokines, as has been reported previously (Balm et al. 1995; Zou et al. 2000; Qin et al. 2001; Pelegrin et al. 2004; Fast et al. 2007; Nascimento et al. 2007). The cells that immunostained for MAC387 and TNF-α antibodies were similar in distribution and morphology, suggesting they may be mainly the macrophages/monocytes. However, with the IL-1β antibody, heterogenous cells were immunostained. In gilthead seabream (Sparus aurata), not macrophages but some acidophilic granulocytes (neutrophils) were immunostained for the IL-1β antibody after infection (Mulero et al. 2008). It is held that pro IL-1β processing depends on caspase 1 and is rapidly released, while the processing of pro IL-1β in the neutrophils is caspase 1-independent, which is slower. Alternatively, handling stress can be a stimulator of the cytokines and hence the immunoreactivity (Fast et al. 2007). The results of the immunocytochemical study in the present are to be viewed with some caution since we used heterologous antibodies of mammalian origin. Future studies using fish homologous antibodies or in situ hybridization for mRNA transcripts are required to confirm the results. It is interesting to note that the MMCs did not give any reaction with the heterologous antibodies. The accumulated pigments might have masked the reaction or the MMC cells are at the final stage of apoptosis or death, and are functionally modified.
Melano-macrophage centers
In the catfish kidney, the MMCs were identified as yellow brown in hematoxylin–eosin, blue green in the Perls’ and ferric ferricyanide reaction and pink magenta in the PAS, implying that they contain melanin, hemosiderin and lipofuscin/ceroid pigments. The MMCs are present in all the divisions of the kidney. The MMCs apart, we found free pigmented macrophages in the kidney. The presence of both free pigmented macrophages and MMCs indicate that the former eventually aggregate forming a new MMC or are added to the existing ones and increasing their size (Meseguer et al. 1994). The MMCs are TUNEL-positive implying that they are formed by apoptotic cells. Apart from the MMCs, we found free and scattered TUNEL-positive cells in the head kidney. The TUNEL-positive bodies are confined to the hemopoietic tissue and not found in the adrenal homologues. The MMCs not only contain lipofuscins but also degraded erythrocytes indicating the presence of apoptotic cells which lead to an increase in the MMCs in the goldfish Carassius auratus (Agius and Roberts, 2003). The results of our study indicate that macrophages undergo apoptosis during the process of phagocytosis and these aggregate to form centers. Judging from the properties (density, size, nature of pigment, etc.), the MMCs may be used as bioindicators of fish health and environmental stress (Agius 1985; Wolke 1992; Fournie et al. 2001).
In the catfish, the MMCs showed significant seasonal variations in their number during the annual reproductive cycle with a low frequency in the gonad resting phase, increasing through the gonad recrudescence to the peak density in the postspawning phase. Such variations were described during the breeding cycle of female ohrid trout (Salmo letnica; Jordanova et al. 2012). In this species, the amount of pigmented macrophages increased significantly in the kidney from previtellogenic to late vitellogenic phase and decreased thereafter in the spawning and postspawning phases. The authors suggested that sex steroid may influence the seasonal variability. In the liver of the same species, Jordanova et al. (2008) reported that hemosiderin-laden pigmented macrophage volume showed a striking increase after spawning but the melano-pigmented macrophages did not. While the cause of the seasonal variations in the head kidney is not clear, we reported seasonal variations in the pigmented atretic follicles in the ovary of the catfish and a steady rise in the melanin contents (Kumar and Joy 2015). Follicular atresia is a well-known process in the ovary and occurs at any stage of development. The postspawning spurge in atretic follicles is a postovulatory event. There is a close parallelism in the seasonal pattern of the ovarian melanin contents and the density of the MMCs and melanin contents in the head kidney. Therefore, the processes that occur in the kidney and ovary may be similar due to phagocytic activity. There are seasonal variations in phagocytosis in seasonally breeding fishes. The postspawning season appears to be the time of heightened cleaning, sanitizing and remodelling activities after breeding, and this allows the fish to prepare for the next reproductive cycle.
In the present study, we characterized the HK melanin pigments biochemically for the first time. In the head kidney, pheomelanin (PM; yellow to reddish brown) is the predominant form, followed by eumelanin (EM; brown black) and alkali-soluble melanin (ASM). In contrast in the ovary, the predominant form is the eumelanin, followed by pheomelanin and ASM (Kumar and Joy 2015). This may be due to differences in the formation of melanin pigments in the tissues. The head kidney melanin pigments showed significant seasonal variations with the content low in the resting phase, increasing through the recrudescence phase to the peak levels in the postspawning phase. There is a close parallelism in the density of the MMCs and melanin contents. Hence, the melanin assay may be useful to quantify MMC activity in the lymphoid organs.
The basis for physiological changes in the melanin content was examined using in vitro head kidney culture experiments. LPS is a pro-inflammatory endotoxin used as a standard macrophage activating agent, augmenting macrophage functions like phagocytosis, activation of HPA axis and cytokine production (Balm et al. 1995; Zou et al. 2000; Qin et al. 2001; Holland et al. 2002). The LPS treatment increased significantly PM and EM levels at both 16 and 32 h and the ASM level at 32 h. The LPS-induced increase in the melanin contents might be due to stress that led to increased MMC activity in the HK. LPS that was observed not effective to increase the melanin levels at 8 h may be due to its insensitivity or fish resistance, as reported earlier (Iliev et al. 2005). On the other hand, dexamethasone, the synthetic glucocorticoid and immunosuppressant decreased the melanin contents at 8 and 16 h. The steroid acted early unlike LPS and may suppress phagocytosis and MMC formation. Dexamethasone is known to inhibit pro-inflammatory responses in animals. In Eupemphix nattereri (Anura), testicular pigmentation increased early (2, 6, 12 h), while the liver pigmentation increased later (24, 48 h) of the LPS treatment (Franco-Belussi et al. 2013). The changes were attributed to bactericidal action of melanin, which neutralizes the effect of LPS (Franco-Belussi and de Oliveira 2011). It is to be seen whether the steroid treatment suppresses the LPS-induced increase in the melanin contents.
In conclusion, various cell types in the hemopoietic tissue of the head kidney were identified. The cells displayed MAC387-, IL-1β- and TNF-α-like immunoreactivity. LIF-like staining was observed in the around cell cords in the IR region and the PCV and blood vessels in the hemopoietic tissue. The MMCs are TUNEL-, ferric ferricyanide-, Perls’- and PAS-positive and show seasonal variation with peak density in the postspawning phase. Head kidney melanins consist of PM, EM and ASM in the decreasing order of concentration. Melanins showed seasonal variations and peaked in the postspawning phase. The LPS treatment increased, and dexamethasone treatment decreased the melanin levels in vitro.
References
Abdel-Aziz EH, Abdu SBS, Ali TE, Fouad HF (2010a) Haemopoiesis in the head kidney of tilapia, Oreochromis niloticus (Teleostei: Cichlidae): a morphological (optical and ultra structural) study. Fish Physiol Biochem 36:323–336
Abdel-Aziz EH, Ali TE, Abdu SBS, Fouad HP (2010b) Chromaffin cells and interrenal tissue in the head kidney of the grouper, Epinephilus tauvina (Teleostei, Serranidae): a morphological (optical and ultra structural) study. J Appl Ichthyol 26:522–527
Abe T, Mikekado T, Haga S, Kisara Y, Watanabe K, Kurokawa T, Suzuki T (2007) Identification, cDNA cloning, and mRNA localization of a zebrafish ortholog of leukemia inhibitory factor. Comp Biochem Physiol B 147:38–44
Abelli L, Gallo VP, Civinini A, Mastrolia L (1996) Immunohistochemical and ultrastructural evidence of adrenal chromaffin cell subtypes in Sea Bass Dicentrarchus labrax (L.). Gen Comp Endocrinol 102:113–122
Agius C (1985) The melano-macrophage centers of fish: a review. In: Manning MJ, Tatner MF (eds) Fish Immunology. Academic Press, London, pp 85–104
Agius C, Roberts RJ (2003) Melano-macrophage centres and their role in fish pathology. J Fish Dis 26:499–509
Auernhammer CJ, Melmed S (2000) Leukemia-inhibitory factor- neuroimmune modulator of endocrine function. Endocr Rev 21:313–345
Balamurugan S, Deivasigamani B, Kumaran S, Sakthivel M, Rajsekar T, Priyadharsini P (2012) Melanomacrophage centres aggregation in P. lineatus spleen as bio-indicator of environmental change. Asian Pac J Trop Dis 12:1–4
Balm PHM, Lieshout E, Lokate J, Bonga SEW (1995) Bacterial lipopolysaccharide (LPS) and interleukin 1 (IL-1) exert multiple physiological effects in the tilapia Oreochromis mossambicus (Teleostei). J Comp Physiol 165:85–92
Bamberger AM, Schulte HM, Wullbrand A, Jung R, Beil FU, Bamberger CM (2000) Expression of leukemia inhibitory factor (LIF) and LIF receptor (LIF-R) in the human adrenal cortex: implications for steroidogenesis. Mol Cell Endocrinol 162:145–149
Bancroft JD, Gamble M (2008) Theory and practice of histological techniques, 6th edn. Churchill Livingstone, London
Besseau L, Faliex E (1994) Resorption of unemitted gametes in Lithognathus mormyrus (Sparidae: Teleostei): a possible synergic action of somatic and immune cells. Cell Tissue Res 276:123–132
Blazer VS (2002) Histopathological assessment of gonadal tissue in wild fishes. Fish Physiol Biochem 26:85–110
Bornstein SR, Rutkowski H, Vrezas I (2004) Cytokines and steroidogenesis. Mol Cell Endocrinol 215:135–141
Braun-Nesje R, Kaplan G, Seljelid R (1982) Rainbow trout macrophages in vitro: morphology and phagocytic activity. Dev Comp Immunol 6:281–291
Butler DG (1973) Structure and function of the adrenal gland of fishes. Am Zool 13:839–879
Castellana B, Iliev DB, Sepulcre MP, Mackenzie S, Goetz FW, Mulero V, Planas JV (2008) Molecular characterization of interleukin-6 in the gilthead seabream (Sparus aurata). Mol Immunol 45:3363–3370
Chesnokova V, Melmed S (2000) Leukemia Inhibitory factor mediates the hypothalamic pituitary adrenal axis response to inflammation. Endocrinology 141:4032–4040
Chester J, Ingleton PM, Phillips JG (1986) Fundamentals of comparative vertebrate endocrinology. Plenum Press, New York, pp 95–120
Civinini A, Padula D, Gallo VP (2001) Ultrastructure and histochemical study on the interrenal cells of the male stickleback, Gasterosteus aculeatus teleostea, in relation to the reproductive annual cycle. J Anat 199:303–316
Crivellato E, Civinini A, Gallo VP (2006) Chromaffin cells in the adrenal Homolog of Aphanius fasciatus (Teleost fish) express Piecemeal degranulation in response to osmotic stress: a hint for a conservative evolutionary process. Anat Rec A Discov Mol Cell Evol Biol 288:1077–1086
Datta S, Ghosh D, Saha DR, Bhattacharaya S, Mazumder S (2009) Chronic exposure to low concentration of arsenic is immunotoxic to fish: role of head kidney macrophage as biomarkers of arsenic toxicity to Clarias batrachus. Aquat Toxicol 92:86–94
Diaz-Satizabal L, Magor BG (2015) Isolation and cytochemical characterization of melanomacrophages and melanomacrophage clusters from goldfish (Carassius auratus L). Dev Comp Immunol 48:221–228
Ellis AE (1977) The leucocytes of fish: a review. J Fish Biol 11:453–491
Engelsma MY, Huising MO, Van Muiswinkel WB, Flik G, Kwang J, Savelkoul HFJ, Verburg-van Kemenade LBM (2002) Neuroendocrine-immune interactions in fish: a role for interleukin-1. Vet Immunol Immunopathol 87:467–479
Fabbri E, Capuzzo A, Moon TW (1998) The role of circulating catecholamines in the regulation of fish metabolism: a overview. Comp Biochem Physiol C 120:177–192
Fan R, Yang G, Dong C (2010) Study of hair melanins in various hair color Alpaca (Lama pacos). Asian-Aust J Anim Sci 23:444–449
Fange R (1986) Physiology of haemopoiesis. In: Nilsson S, Holmgren S (eds) Fish physiology: recent advance. Groom Helm, London, pp 1–23
Fast MD, Johnson SC, Jones SRM (2007) Differential expression of the pro-inflammatory cytokines IL-1b-1, TNFa-1 and IL-8 in vaccinated pink (Oncorhynchus gorbuscha) and chum (Oncorhynchus keta) salmon juveniles. Fish Shellfish Immunol 22:403–407
Fijan N (2002) Composition of main haematopoietic compartments in normal and bled channel catfish. J Fish Biol 60:1142–1154
Fishelson L (2006) Cytomorphological alterations of the thymus, spleen, head-kidney, and liver in cardinal fish (Apogonidae, Teleostei) as bioindicators of stress. J Morphol 267:57–69
Fournie JW, Summers JK, Courtney LA, Engle VD (2001) Utility of splenic macrophage aggregates as an indicator of fish exposure to degraded environments. J Aquat Anim Health 13:105–116
Franco-Belussi L, de Oliveira C (2011) Lipopolysaccharides induce changes in the visceral pigmentation of Eupemphix nattereri (Anura: Leiuperidae). Zoology 114:298–305
Franco-Belussi L, de Lauro Castrucci AM, de Oliveira C (2013) Response of melanocytes and melanomacrophages of Eupemphix nattereri (Anura: Leiuperidae) to Nle4, D-Phe7-α-melanocyte stimulating hormone and lipopolysaccharides. Zoology 116:316–324
Gallo VP, Civinini A, Mastrolia L, Leitner G, Porta S (1993) Cytological and biochemical studies on chromaffin cells in the head kidney of Gasterosteus aculeatus (Teleostei, Gasterosteidae). Gen Comp Endocrinol 92:133–142
Gonzalez-Hernandez JA, Bornstein SR, Ehrhart-Bornstein M, Gschwend JE (1995) IL-1 is expressed in human adrenal gland in vivo. Possible role in a local immune-adrenal axis. Clin Exp Immunol 99:137–141
Guthrie HD, Grimes RW, Cooper BS, Hammond JM (1995) Follicular atresia in pigs: measurement and physiology. J Anim Sci 73:2834–2844
Guzman C, Hernandez-Bello R, Morales-Montor J (2010) Regulation of steroidogenesis in reproductive, adrenal and neural tissues by cytokines. Open Neuroendocrinol J 3:161–169
Herraez MP, Zapata AG (1991) Structural characterization of the melano-macrophage centres (MMC) of goldfish Carassius auratus. Eur J Morphol 29:89–102
Holland JW, Pottinger TG, Secombes CJ (2002) Recombinant interleukin-1 activates the hypothalamic–pituitary–interrenal axis in rainbow trout, Oncorhynchus mykiss. J Endocrinol 175:261–267
Hughes FM Jr, Gorospe WC (1991) Biochemical identification of apoptosis (programmed cell death) in granulosa cells: evidence for a potential mechanism underlying follicular atresia. Endocrinology 129:2415–2422
Iliev DB, Roach JC, Mackenzie S, Planas JV, Goetz FW (2005) Endotoxin recognition: in fish or not in fish? FEBS Lett 579:6519
Ito S, Wakamatsu K, Ozeki H (2000) Chemical analysis of melanins and its application to the study of the regulation of melanogenesis. Pigment Cell Res 13:103–109
Janz DM, Kraak GVD (1997) Suppression of apoptosis by gonadotropin, 17β-estradiol, and epidermal growth factor in rainbow trout preovulatory ovarian follicles. Gen Comp Endocrinol 105:186–193
Jordanova M, Miteva N, Rocha E (2008) A qualitative and quantitative study of the hepatic pigmented macrophage aggregates during the breeding cycle of Ohrid Trout, Salmo letnica Kar. (Teloestei, Salmonidae). Microsc Res Tech 71:822–830
Jordanova M, Rocha MJ, Rebok K, Rocha E (2012) Changes in the amount of kidney pigmented macrophage aggregates throughout the breeding cycle of female Ohrid trout, Salmo letnica Kar. (Teleostei, Salmonidae). Microsc Res Tech 75:176–181
Kemenade BMLV, Stolte EH, Metz JR, Chadzinska M (2009) Neuroendocrine-immune interactions in teleost fish. Fish Neuroendocronol 28:313–365
Kemenade BMLV, Ribeiro CMS, Chadzinska M (2011) Neuroendocrine–immune interaction in fish: differential regulation of phagocyte activity by neuroendocrine factors. Gen Comp Endocrinol 172:31–38
Kumar R, Joy KP (2015) Melanins as biomarkers of ovarian follicular atresia in the catfish Heteropneustes fossilis: biochemical and histochemical characterization, seasonal variation and hormone effects. Fish Physiol Biochem 41:761–772
Lei L, Tzekov R, Tang S, Kaushal S (2012) Accumulation and autofluorescence of phagocytized rod outer segment material in macrophages and microglial cells. Mol Vis 18:103–113
Lofts B, Bern HA (1972) Functional morphology of steroidogenic tissue. Idler DR (Edn) Steroids in non-mammalian vertebrates. Academic Press, New York, pp 37–125
Mastrolia L, Gallo VP, La Marca A (1984) The adrenal chromaffin cells of Salmo gairdneri Richardson (Teleostei, Salmonidae). J Anat 138:503–511
Matsche MA, Grizzle JM (1999) Early changes in pigmented macrophages in head kidney of channel catfish infected with Aeromonas hydrophila. J Aquat Anim Health 11:253–261
Meseguer J, Lopez-Ruiz A, Esteban MA (1994) Melano-macrophages of the seawater teleosts, sea bass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata): morphology, formation and possible function. Cell Tissue Res 277:1–10
Metz JR, Huising MO, Leon K, Vernurg-van Kemenade BML, Flik G (2006) Central and peripheral interleukin-1β and interleukin-1 receptor expression and their role in the stress response of common carp, Cyprinus carpio L. J Endocrinol 191:25–35
Milano EG, Basari F, Chimenti C (1997) Adrenocortical and adrenomedullary homologs in eight species of adult and developing teleosts: morphology, histology and immunohistochemistry. Gen Comp Endocrinol 108:483–496
Montpetit CJ, Perry SF (2002) Adrenergic regulation of catecholamine secretion from trout (Oncorhynchus mykiss) chromaffin cells. J Endocrinol 173:187–197
Mulero I, Sepulcre MP, Roca FJ, Meseguer J, Garcia-Ayala A, Mulero V (2008) Characterization of macrophages from the bony fish gilthead seabream using an antibody against the macrophage colony-stimulating factor receptor. Dev Comp Immunol 32:1151–1159
Nandi J (1962) The structure of the interrenal gland in teleost fishes. Univ Calif Publ Zool 65:129–212
Nascimento DS, Pereira PJB, Reis MIR, Vale A, Zou J, Silva MT, Secombes CJ, Santos NMS (2007) Molecular cloning and expression analysis of sea bass (Dicentrarchus labrax L.) tumor necrosis factor-α (TNF-α). Fish Shellfish Immunol 23:701–710
Ozeki H, Ito S, Wakamatsu K, Hirobe T (1995) Chemical characterization of hair melanins in various coat-color mutants of mice. J Invest Dermatol 105:361–366
Pelegrin P, Chaves-Pozo E, Mulero V, Meseguer J (2004) Production and mechanism of secretion of interleukin-1β from the marine fish gilthead seabream. Dev Comp Immunol 28:229–237
Press CM, Evensen O (1999) The morphology of the immune system in teleost fishes. Fish Shellfish Immunol 9:309–318
Qin QW, Ototake M, Noguchi K, Soma G, Yokomizo Y, Nakanishi T (2001) Tumor necrosis factor alpha (TNFα)-like factor produced by macrophages in rainbow trout, Oncorhynchus mykiss. Fish Shellfish Immunol 11:245–256
Reid SG, Bernier NJ, Perry SF (1998) The adrenergic stress response in fish: control of catecholamine storage and release. Comp Biochem Physiol C 120:1–27
Ribeiro HJ, Procopio MS, Gomes JMM, Vieira FO, Russo RC, Balzuweit K, Chiarini- Garcia H, Casrro ACS, Rizzo E, Correa JD (2011) Functional dissimilarity of melano-macrophage centres in the liver and spleen from females of the teleost fish Prochilodus argenteus. Cell Tissue Res 346:417–425
Rocha RM, Santes HS, Vecentini CA, Cruz C (2001) Structural and ultrastructural characteristics of interrenal gland and chromaffin cell of matrinxa. Brycon cephalus Gunther 1869 (Teleostei-Characidae). Anat Histol Embryol 30:351–355
Romano N, Picchietti S, Taverne-Thiele JJ, Taverne N, Abelli L, Mastrolia L, Kemenade BMLV, Rombout JHWM (1998) Distribution of macrophages during fish development: an immunohistochemical study in carp (Cyprinus carpio L.). Anat Embryol 198:31–41
Sailendri K, Muthukkaruppan VR (1975) Morphology of lymphoid organ in a cichlid teleost, Tilapia mossambica (Peters). J Morphol 147:109–122
Santos HB, Thome RG, Arantes FP, Sato Y, Bazzoli N, Rizzo E (2008) Ovarian follicular atresia is mediated by heterophagy, autophagy, and apoptosis in Prochilodus argenteus and Leporinus taeniatus (Teleostei: Characiformes). Theriogenology 70:1449–1460
Sarmento A, Marques F, Ellis AE, Afonso A (2004) Modulation of the activity of sea bass (Dicentrarchus labrax) head-kidney macrophages by macrophage activating factor (s) and lipopolysaccharide. Fish Shellfish Immunol 16:79–92
Secombes CJ, Fletcher TC (1992) The role of phagocytes in the protective mechanisms of fish. Ann Rev Fish Dis 2:53–71
Seljelid R, Eskeland T (1993) The biology of macrophages: I general principles and properties. Eur J Haematol 51:267–275
Sorensen KK, Sveinbjornsson B, Dalmo RA, Smedsrod B, Bertheussen K (1997) Isolation, cultivation and characterization of head kidney macrophages from Atlantic cod, Gadus morhua L. J Fish Dis 20:93–107
Tesch GH, Yang N, Yu H, Lan HY, Foti R, Chadban SJ, Atkins RC, Nikolic- Paterson DJ (1997) Intrinsic renal cells are the major source of interleukins-1β synthesis in normal and diseased rat kidney. Nephrol Dial Transplant 12:1109–1115
Tilly JL, Kowalski KI, Johnson AL, Hsueh AJW (1991) Involvement of apoptosis in ovarian follicular atresia and postovulatory regression. Endocrinology 129:2799–2801
Tkachenko IV, Jaaskelainen T, Jaaskelainen J, Palvimo JJ, Voutilainen R (2011) Interleukins 1α and 1β as regulators of steroidogenesis in human NCI-H295R adrenocortical cells. Steroids 76:1103–1115
Ucuncu SI, Cakıcı O (2009) Atresia and apoptosis in preovulatory follicles in the ovary of Danio rerio (zebrafish). Turk J Fish Aquat Sci 9:215–221
Vermeulen GJ, Lambert JGD, Teitsma CA, Zandbergen MA, Goos HJT (1995) Adrenal tissue in the male African catfish, Clarias gariepinus: localization and steroid hormone secretion. Cell Tissue Res 280:653–657
Vigliano FA, Bermudez R, Quiroga IM, Nieto JM (2006) Evidence for melano-macrophage centres of teleost as evolutionary precursors of germinal centres of higher vertebrates: an immunohistochemical study. Fish Shellfish Immunol 21:467–471
Wolke RE (1992) Piscine macrophage aggregate, a review. Ann Rev Fish Dis 2:91–108
Wood AW, Van Der Kraak GJ (2001) Apoptosis and ovarian function: novel perspectives from the teleosts. Biol Reprod 64:264–271
Woods AM, McIlmoil CJ, Rankin EN, Packer AA, Stevens JC, Macievic JA, Brown AB, Porter JP, Judd AM (2008) Leukemia inhibitory factor protein and receptors are expressed in the bovine adrenal cortex and increase cortisol and decrease adrenal androgen release. Domest Anim Endocrinol 35:217–230
Zapata A, Diez B, Cejalvo T, Gutiérrez-de Frías C, Cortés A (2006) Ontogeny of the immune system of fish. Fish Shellfish Immunol 20:126–136
Zou J, Holland J, Pleguezuelos O, Cunningham C, Secombes CJ (2000) Factors influencing the expression of interleukin-1β in cultured rainbow trout (Oncorhynchus mykiss) leucocytes. Dev Comp Immunol 2:575–582
Zuasti A, Jara JR, Ferrer C, Solano F (1989) Occurrence of melanin granules and melanosynthesis in the kidney of Sparus auratus. Pigment Cell Res 2:93–99
Acknowledgments
R.K. is a recipient as senior research fellow of Rajiv Gandhi National Fellowship of UGC, New Delhi, which is gratefully acknowledged. This work was partly supported by the UGC-CAS Programme (No. 4197) to Department of Zoology, BHU. Our thanks are due to Prof. B. Lal, Department of Zoology, BHU, for the photomicrography facility.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kumar, R., Joy, K.P. & Singh, S.M. Morpho-histology of head kidney of female catfish Heteropneustes fossilis: seasonal variations in melano-macrophage centers, melanin contents and effects of lipopolysaccharide and dexamethasone on melanins. Fish Physiol Biochem 42, 1287–1306 (2016). https://doi.org/10.1007/s10695-016-0218-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-016-0218-2