
Abstract
Wheat production in cooler regions like the north-western Himalayas, is significantly impeded by devastating diseases, namely stripe rust (SR) and powdery mildew (PM). Genetic resistance against SR and PM loses effectiveness over time which underscores the importance of periodic disease screening. This study aims to assess resistance to SR and PM in 81 wheat genotypes across multiple locations over three years (2019–20, 2021–22 and 2022–23); and detect candidate genes (Yr5, Yr10 and Pm24) for resistance using respective molecular markers viz., SSR/STS primers (STS7/8, Xp3000 and Xgwm337). The resistance towards SR and PM under natural epiphytotic conditions was displayed by eight and twelve genotypes respectively, across all locations. Notably, four genotypes (DH 202, HPW 368, HPW 373 and DH 114) were found resistant to both diseases. The phenotypic disease reaction for SR and PM was further validated through molecular markers. Genotypes DH 202, DH208, DH 217, CIMMYT Entry no. 23 and VL 829 emerged as high yielding disease resistant genotypes. Agrometeorological parameters specifically, precipitation and relative humidity exhibited significant positive correlations with disease incidence, leading to reduced grain yields. Genotype and genotype by environment interaction (GGE) biplot identified stable genotypes with less disease incidence over locations. Additionally, Kukumseri may serve as the optimal test site for screening wheat germplasm against SR, while Palampur and Kukumseri could be ideal for PM screening. Genotypes exhibiting combined disease resistance to both SR and PM, alongwith superior agronomic traits, hold promise for immediate deployment as wheat varieties or as potential donors for breeding resistant cultivars.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wheat, (Triticum aestivum L., 2n = 6x = 42) is an important cereal grain crop that serves as a staple food for millions of people worldwide. According to the latest data from FAO (2022), it is cultivated over 219.15 million hectares worldwide, with a production of 808.44 million tonnes. Globally, China leads wheat production with 17.04%, followed by India with 13.33%. Wheat stands as a cornerstone of global food security and contributing approximately 20% of the calories and proteins essential for human nutrition. The grains primarily consist of carbohydrates (~ 70%–80%) and proteins (~ 8%–22%) (Slafer et al. 2021). Rapidly growing global population, shrinking arable land, and exacerbating effects of climate change increases the frequency of abiotic and biotic stresses, which significantly constrain wheat productivity (Valizadeh et al. 2014).
A variety of diseases such as rusts, bunts, powdery mildew and smuts, along with abiotic stresses like heat, drought and salinity pose significant obstacles to wheat crop production (Chatrath et al. 2007). Notably, among these diseases, stripe rust (SR) and powdery mildew (PM), caused by Puccinia striiformis f. sp. tritici and Erysiphe graminis f. sp. tritici (syn. Blumeria graminis f. sp. tritici), respectively, are particularly devastating. These pathogens are well adapted to temperate areas with cool-humid weather, which are generally associated with higher elevations (Bennett 1984; Roelfs et al. 1992; Han et al. 2020; Mehta et al 2022). Specifically, SR pathogen initiates primary infection in December and January in the North hill zone (NHZ) of India, where conditions are conducive. Subsequently, winds transport the urediospores to the surrounding foothills, further leading to the spread of infection to the major wheat-producing north west plain zone (NWPZ) (Nagarajan 1977).
Globally, wheat cultivation has experienced recurring epidemics of SR, capable of causing total crop loss (Bhardwaj et al. 2019; Hovmøller et al. 2016). In severe cases, PM can cause wheat yields to drop by 50% (Morgounov et al. 2012; Xu et al. 2015). To address these challenges, plant breeders consistently seek for germplasm with a diverse array of genes/alleles to improve resilience, which involves exploring a range of potential germplasm to uncover new sources of alleles and selecting lines with greater adaptability (Verma et al. 2024). Researchers across the globe have identified various SR and PM resistance genes in wheat germplasm, among which Yr5, Yr10, Yr15, Yr24/Yr26, Yr32, YrSp (against P. striiformis) (Rani et al. 2019) and Pm1a, Pm2, Pm3/Pm8/Pm17, Pm5e, Pm21/Pm12, Pm24, Pm33, Pm41, Pm51, Pm60, Pm64, Pm69, MlZec1 and MlAB10 (against E. graminis) are still effective (Gupta et al. 2022).
Grain yield in wheat is a complex trait influenced by various genetic and environmental factors. Thus, the stability of genotypes for their performance can be best analyzed under multi-environment evaluations, providing a clear picture of their adaptability across locations (Sharma et al. 2022). In breeding programs, understanding genotype × environment interaction (GEI) is important for effectively optimizing host-plant resistance across environments and pathosystem dynamics (Sankar et al. 2021). Among various statistical methods used for GEI analysis, genotype and genotype by environment interaction (GGE) biplot analyses are widely utilized for assessing multi-environment data (Abraha et al. 2019).
So, in the present study a diverse set of wheat germplasm comprising of doubled haploids, exotic collections, landraces and popular cultivars were evaluated for adult plant response against SR and PM under natural epiphytotic field conditions at four different locations depicting diverse agroclimatic zones of the north-western Himalayan region. Further, allelic differences among wheat genotypes were characterized at genetic level by molecular markers associated with SR and PM resistance genes.
Materials and methods
Experimental material
The study involved 81 diverse germplasm accessions, comprising 23 established doubled haploids, 11 promising landraces from the north-western Himalayas, 40 outstanding exotic collections from CIMMYT, Mexico and 7 elite Indian wheat varieties (Supplementary Table S1). The experiment was conducted at four different locations namely, CSK HP Agricultural University, Palampur; Krishi Vigyan Kendra (KVK), Sundernagar; Hill Agricultural Research and Extension Centre (HAREC), Bajaura; and HAREC Kukumseri, situated in different agroclimatic zones of the north-western Himalayan region for three years (2019–20, 2021–22 & 2022–23) (Fig. 1 and Table 1). Germplasm was sown in randomized complete block design (RCBD) with three replicates, during mid-November at all locations, except at HAREC Kukumseri, where sowing was done in April. Two rows of each entry were grown in 1 m length with inter- and intra-row distance of 20 cm and 10 cm, respectively. To ensure the uniform spread of inoculum, two infector rows of susceptible varieties like Sonalika and Agra Local were sown after every nine rows within the plot and around the experimental trial. Weather parameters for each cropping season at each location were recorded by NASA POWER DAVe (https://power.larc.nasa.gov/data-access-viewer/) (Fig. 2 and Supplementary Table S2).

a Disease incidence under field conditions, b Symptoms of Stripe rust, and c Powdery mildew on wheat plant

Variations in temperature and relative humidity during cropping seasons at different environments
Phenotyping and disease scoring
Germplasm was screened for resistant genotypes against stripe rust (SR) and powdery mildew (PM) at adult plant stage, under natural epiphytotic conditions. The disease reactions were recorded on 10 random plants at heading stage for each genotype in each replication. Disease reactions for SR was recorded in accordance to, 0–100 modified Cobb’s Scale (Peterson et al. 1948). The host responses were graded: S = susceptible (> 30), abundant mycelium development and uredia surrounded by necrotic tissues; MS = moderately susceptible (21–30), development of mycelium and small uredia surrounded by necrotic tissues; MR = moderately resistant (11–20), slight uredia surrounded by necrotic tissues; R = resistant (6–10), very slight uredia surrounded by necrosis and chlorosis; and HR = Highly resistant, slight flecking of necrotic/chlorotic spot (1–5) (Akin et al 2008). While, PM was monitored in accordance to, Saari-Prescot 0–9 scale (Bennett and Westcott 1982), where ‘1’ represents small flecks (HR), ‘2’ small chlorotic flecks (R), ‘3’ large flecks with chlorosis and necrosis (MR), ‘4–6’ mycelium and conidia barely detectable with small and moderately sized lesions (MS) and ‘7–9’ increased amount of mycelium and conidia production (S).
At each location, the data for flag leaf area (FLA) (cm2) and grain yield per plant (GY) (g) were recorded in triplicates for each genotype and in each replication five random competitive plants were chosen. To further estimate the effect of disease on agro-morphological traits, total chlorophyll content (TCC) (mg/g) was estimated using Witham et al. (1986) at Molecular Cytogenetics and Tissue culture laboratory, Department of Genetics and Plant Breeding, CSKHPKV, Palampur.
Molecular studies
Molecular studies were performed in the Molecular Cytogenetics & Tissue Culture Lab, Department of Genetics and Plant Breeding, CSKHPKV, Palampur. Genomic DNA was extracted from juvenile leaves of 10–15-day-old seedlings using the CTAB method described by Doyle and Doyle (1987), with slight modifications. DNA quantity was determined with 0.8% agarose gel and quality was checked using Eppendorf BioSpectrometer® basic.
Specific SSR/STS primers linked to SR resistance genes Yr5 (STS7/8) and Yr10 (Xpsp3000) were used (Rani et al 2019). Similarly, SSR primers associated with PM resistance genes Pm24 (Xgwm337) was employed (Cheng et al 2022) (Table 2).
DNA amplification was carried out using 12.5 µl reaction mixture consisting of 1 µl template DNA (50 ng), 1.25 µl 10X PCR buffer, 1.25 µl 25 mM MgCl2, 1.25 µl 2 mM dNTPs, 0.5 µl 10 µM each of forward and reverse primers, 0.25 µl Taq DNA polymerase and 6.5 µl sterilized doubled distilled water in Eppendorf thermal-cycler. The PCR regimen consisted of initial denaturation step at 94 °C for 5 min, followed by 35 cycles of denaturation (94 °C), annealing (X°C) and extension (72 °C) each for 45 s. The final extension step was carried out at 72 °C for 5 min. Subsequently, the PCR products were subjected to electrophoresis on a 3.5% agarose gel at 120 V for 120–150 min. The size of the amplicons was determined using a 100 bp DNA ladder and visualized with a gel documentation system (UVITEC, Cambridge).
Data analysis
The average values from the 81 genotypes for FLA, TCC, SR, PM and GY from different environments and pooled over environments were subjected to analysis of variance (ANOVA) using Microsoft Excel Data Analysis tools. Simultaneously, Bartlett’s test was applied for testing homogeneity of variance and validating ANOVA. To assess the average performance of genotypes across all environments, phenotypic data were analyzed using best linear unbiased predictions (BLUP) modeling. The BLUP values obtained were analyzed and interpreted in comparison to the best check(s) for each trait in each pooled environment. BLUP values, least significance difference (LSD) test, heritability (H2) and genetic advance as % of mean (GA) were worked out using the META-R program (Alvarado et al. 2020).
The Pearson’s correlation coefficient (r) between agro-morphological traits and disease scores was determined across locations and combined. These results are graphically represented by using R version 4.0.5 (R Core Team 2021; Peterson and Carl 2020). Pooled correlation was also worked out with agrometeorological data including temperature, precipitation, relative humidity (RH%) and root zone soil wetness (RZSW%). The GGE Biplot analysis for all environments was performed using the package GGE Biplots of R studio (R Core Team 2021).
Results
Mean performance and analysis of variance
ANOVA was performed for all the traits per individual environments and pooled over years for all the locations, which indicated that the mean sum of squares of genotypes were significantly different for FLA, TCC, SR, PM and GY across all the four locations over three years, suggesting ample genetic variability for the studied traits in the germplasm (Supplementary Table S3). Bartlett’s test confirmed that variances were uniform for studied traits. Consequently, pooled ANOVA for four locations combined over three years revealed that the mean squares attributed to genotypes and genotype × year interaction differed significantly; suggesting morpho-genetic variability among treatments throughout the experiments (Table 3).
BLUP values for FLA ranged from 17.34–31.98 cm2 with an average of 24.29 cm2. GY ranged from 5.56–8.83 g with an average of 7.05 g. Five genotypes namely, DH 202, DH 210, DH 217, CIMMYT Entry no. 23 and Chamba landrace 3, showed significantly higher yields over their respective check varieties (VL829, HPW373 and DH114) across all the environments. Additionally, the average GY is highest in Sundernagar and lowest in Kukumseri.
High heritability (95%) coupled with moderate genetic advance (23.63%) was observed for FLA, while moderate heritability (72%) combined with low genetic advance (19.34%) was observed for GY (Table 4, Fig. 3, Supplementary Table S4 and S5).


PCR amplification profile of marker a STS7/8, associated with SR resistance gene Yr5, and b Xpsp3000 linked with SR resistance gene Yr10
Disease screening under field conditions
Eight genotypes (9.88%) viz., DH 202, DH 208, DH 217, CIMMYT Entry No. 133, CIMMYT Entry No. 278, HPW 368, HPW 373 and DH 114 showed consistent SR resistant reactions over years at all the locations. For powdery mildew 12 genotypes (14.81%) viz., DH 195, DH 198, DH 202, DH 219, CIMMYT Entry no.23, CIMMYT Entry no.242, CIMMYT line 30–16, Chamba landrace 1. HPW 368, HPW 373, VL 829 and DH 114 demonstrated consistent PM resistant reactions over years at all the locations.
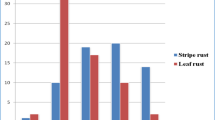
Moreover, 11 and 28 genotypes were found resistant to SR and PM, respectively in pooled over 12 environments (Table 5 and Supplementary Table S6). Overall, the disease intensity for SR is highest at Kukumseri, while for PM, it is higher at Kukumseri and Palampur than rest of the locations.
Screening at molecular level
The STS marker, STS7/8 (resistant amplicon size 500 bp), used in the study is the dominant type marker, indicating the presence of the SR resistance gene Yr5, amplified in 71 lines (87.65%), while the remaining 8 wheat genotypes failed to amplify indicating the absence of the gene. On the other hand, the SSR marker Xpsp3000 (resistant amplicon size 260 bp) was detected in 18 genotypes (22.22%), indicating the presence of SR resistance Yr10 gene, while it was absent in the remaining 62 entries tested (Fig. 3). Eleven genotypes, namely DH 198, DH 200, DH 207, DH 208, DH 216, CIMMYT Entry no.278, CIMMYT line 60-35, CIMMYT line 30-7, Chamba landrace 16, Chamba landrace 17 and DH 114 each possessing both SR resistance genes (Yr5 and Yr10) demonstrated resistant reactions in fields. However, seven genotypes, DH 202, DH 217, CIMMYT line 60-44, CIMMYT line 30-10, HPW 360, HPW 368 and HPW 373 exhibited resistance to SR in the fields without amplifying either or both of the genes (Yr5 and Yr10), whereas two genotypes (DH196 and CIMMYT line 30-5) displayed moderately susceptible reactions despite possessing both genes (Fig. 3 and Table 6).
PM resistance gene Pm24 was not found at all in the whole germplasm as maker Xgwm337 failed to amplify.
Correlation studies
Correlation studies conducted across all locations (Fig. S1), pooled results showed a notable positive correlation between GY and TCC, conversely both morpho-biochemical traits were negatively associated with SR and PM occurrences (Fig. 4a). Additionally, the correlation between the studied traits, disease prevalence and agrometeorological highlighted significant positive correlations between precipitation and RH% with SR and PM diseases, while showing negative correlations with GY (Fig. 4b).

Heatmap displaying Pearson’s correlation coefficient pooled across locations among a studied traits with disease scores, and b studied traits and disease scores with agrometeorological data (* P < 0.05) (Temp-Temperature(ºC), Ppt- Precipitation, RH%- Relative Humidity, RZSW- Root zone soil wetness)
Mega-environment analysis of genotypes
The GGE biplot revealed that PC1 (disease score) and PC2 (resistance stability) accounted for 58.66% & 16.73% and 40.42% & 20.33% of the total variation for SR and PM, respectively (Figs. 6).
The ‘which-won-where’ perspective of the GGE biplot demonstrated that genotypes i.e. G-5 (DH 198), G-9 (DH 202), G-15 (DH 208), G-19 (DH 216), G-20 (DH 217), G-36 (CIMMYT Entry no.133), G-40 (CIMMYT Entry no.240), G-41 (CIMMYT Entry no.242), G-43 (CIMMYT Entry no.278), G-49 (CIMMYT line 60-44), G55 (CIMMYT line 30-24), G-58 (CIMMYT line 30-10), G-75 (HPW 360), G-76 (HPW 368), G-77 (HPW 373), G-79 (VL892) and G-81 (DH 114) had low levels of SR intensity by being the farthest to the right side of the origin of biplot. While Genotypes, G-13 (DH 206), G-21 (DH 218), G-29 (CIMMYT Entry no.92), G-30 (CIMMYT Entry no. 95), G-31 (CIMMYT Entry no.98), G-32 (CIMMYT Entry no.101), G-45 (CIMMYT line 60-24), G-68 (Chamba landrace 2), G-70 (Chamba landrace 13) and G-72 (Chamba landrace 15) constantly showed higher disease score for SR and were located outermost to the left side of the origin of the biplot (Fig. 5a).

Which-won-where view of GGE biplot based on a SR, and b PM disease scores on 81 genotypes of wheat under 12 environments Green numbers correspond to genotypes as listed in (Supplementary Table S1)
The genotypes present at the left side of the hull showed more PM susceptibility and those on the right side had stable resistance across the environments. Genotypes i.e. G-2 (DH 195), G-4 (DH 198), G-5 (DH 198), G-29 (CIMMYT Entry no.92), G-41 (CIMMYT Entry no.242), G-56 (CIMMYT line 30-16), G-64 (Chamba landrace 1), G-70 (Chamba landrace 13), G-76 (HPW 368), G-77 (HPW 373), G-78 (VL 829) and G-81 (DH 114) had low levels of PM intensity by being farthest to the left side of the origin of the biplot (Fig. 5b). Genotypes, viz., G-13 (DH 206), G-14 (DH 207), G-15 (DH 208), G-30 (CIMMYT Entry no.95), G-33 (CIMMYT Entry no.105), G-34 (CIMMYT Entry no.106), G-35 (CIMMYT Entry no.107), G-37 (CIMMYT Entry no.164), G-45 (CIMMYT line 60-24) and G-59 (CIMMYT line 30-7) consistently showed high level of disease score and were located outermost to the right side of the origin of the biplot (Fig. 5b).
The polygon view partitioned the biplot into several sectors, aiding in the clustering of environments into mega-environments (ME). All the environments were categorized into three MEs for SR and two MEs for PM. Mega environments I (ME-I) for SR includes environments E-1 (Palampur 2019–20), E-2 (Palampur 2021–22), E-5 (Sundernagar 2021–22), E-7 (Bajaura 2019–20), E-8 (Bajaura 2021–22) and E-9 (Bajaura 2022–23), ME-II comprised environments E-3 (Palampur 2022–23), E-4 (Sundernagar 2019–20) and E-6 (Sundernagar 2022–23) and ME-III comprised E-10 Kukumseri 2019), E-11 (Kukumseri 2021) and, E-12 (Kukumseri 2022) (Fig. 5a). While, for PM, ME-I include E-1 (Palampur 2019–20), E-2 (Palampur 2021–22), E-3 (Palampur 2022–23), E-9 (Bajaura 2022–23), E-10 (Kukumseri 2019), E-11 (Kukumseri 2021) and E-12 (Kukumseri 2022) and ME-II comprised environments E-4 (Sundernagar 2019–20), E-5 (Sundernagar 2021–22), E-6 (Sundernagar 2022–23), E-7 (Bajaura 2019–20) and E-8 (Bajaura 2021–22) (Fig. 5b).
Mean vs. Stability
The GGE biplot of “Mean vs. Stability” view ranked genotypes based on their average performance across 12 environments for SR and PM. The single arrowed line indicated the AEC (average environment coordinate) abscissa, pointing towards higher disease intensity (Yan and Tinker 2006) (Figs. 7). The GGE biplot revealed that, in terms of the least disease occurrence for SR, the overall best performing genotypes with wider adaptability were G-9 (DH 202), G-15 (DH 208), G-19 (DH 216), G-20 (DH 217), G-36 (CIMMYT Entry no.133), G-43 (CIMMYT Entry no.278), G-49 (CIMMYT line 60-44), G-58 (CIMMYT line 30-10), G-75 (HPW 360), G-76 (HPW 368), G-77 (HPW 373), G-79 (VL892) and G-81 (DH 114) (Fig. 6a). In case of PM, i.e., G-2 (DH 195), G-4 (DH 197), G-5 (DH 198), G-76 (HPW 368), G-77 (HPW 373), G-78 (VL 829) and G-81 (DH 114) were the overall best genotypes (Fig. 6b).

Mean vs stability view of GGE biplot based on, a SR, and b PM disease scores on 81 genotypes of wheat under 12 environments

Discriminativeness vs. representativeness view of GGE biplot based on, a SR, and b PM disease scores on 81 genotypes of wheat under 12 environments
Evaluating test environments: discrimination ability and representativeness
The ‘discrimitiveness vs. representativeness’ view of GGE biplot for test environments explained that E-10 (Kukumseri 2019) and E-11 (Kukumseri 2021) had greater vector length for SR, while E-3 (Palampur 2022–23) exhibited a greater vector length for PM compared to other environments, suggesting the higher capability of these environments for discriminating and distinguishing genotypes based on the respective disease (Fig. 7).
A smaller (acute) angle of test environment vector with AEC signifies stronger representativeness of the environment (Mehta et al. 2022; Das et al. 2019). Small angles of E-3 (Palampur 2022–23), E-4 (Sundernagar 2019–20) and E-6 (Sundernagar 2022–23) with AEC for SR; and E-1 (Palampur 2019–20), E-9 (Bajaura 2022–23) and E-10 (Kukumseri 2019) for PM, were indicative of stronger representativeness (Fig. 7 a and b).
Discussion
Developing elite wheat cultivars through resistant breeding programs represents a financially and environmentally sustainable strategy for disease management. However, despite developing resistant varieties against SR and PM, there remains a potential risk of resistance loss, owing to pathogen evolution and emergence of new strains. This highlights the need for regular disease screenings and the identification of diverse resistance genes.
Periodic screening of wheat germplasm over seasons and locations, against prevalent pathogen races is crucial to develop high-yielding varieties with durable resistance to multiple diseases (Singh et al. 2015). The present study was such a kind of periodic screening, conducted at four different locations in diverse agroclimatic zones of the north-western Himalayan region, well-recognized as a hotspot for SR and PM diseases. These locations provide varied habitats for both the crop and pathogens, enabling effective natural screening of resistant germplasm against SR and PM. Phenotypic evaluation of the tested genotypes for various agro-morphological traits showed significant differences across all the four locations. Five genotypes, namely DH 202, DH 210, DH 217, CIMMYT Entry no. 23 and Chamba landrace 3, demonstrated significantly higher yields compared to their respective check varieties across various locations. Sharma et al. (2022) and Jee et al. (2019) also observed significant differences among wheat genotypes they evaluated. High heritability coupled with moderate genetic advance for FLA, indicating additive and non-additive gene action in the inheritance, providing scope of improvement through selection for this trait. Conversely, moderate heritability combined with low genetic advance was observed for GY, indicating non-additive gene action. Therefore, selections based solely on GY would not be effective. The findings of this study align with those of Adhikari et al. (2018) and Singh et al. (2018). Field screening of the germplasm revealed that 12 and 28 genotypes were highly resistant to SR and PM, respectively (Table 5). Among these, four genotypes, namely DH 202, HPW 368, HPW 373 and DH 114, demonstrated resistance to both SR and PM. Notably, DH 202, DH 208 and DH 217 exhibited high yields along with resistance to SR, while DH 202, CIMMYT Entry no. 23 and VL 829 showed high yields coupled with resistance to PM. Overall, a high-yielding genotype DH 202 exhibited resistance to both SR and PM. In the study conducted by, Mishra et al. (2015) it was observed that out of 616 accessions, 197 were found to be resistant to SR. Kumar et al. (2016) screened 19,460 accessions for SR at a hotspot, Gurdaspur (Punjab) and identified 498 potential resistant accessions to multiple rusts. Similar set of wheat germplasm (19,460 accessions) were screened by Vikas et al (2020) at Wellington, a hotspot for PM, for two consecutive seasons, results indicated that 7,271 accessions were resistant.
The primary goal of plant breeders is to develop varieties that consistently resist multiple diseases and produce higher yields. This is achieved by identifying and combining diverse disease-specific resistance genes. Molecular analysis showing distinct amplification patterns in resistant and susceptible genotypes confirmed the phenotypic evaluation. Among the twelve genotypes demonstrating the presence of both SR resistance genes (Yr5 and Yr10), eleven exhibited resistance in the field across multiple locations.
The results of molecular screening for SR resistance are consistent with those reported by Rani et al. (2019), who observed the presence of STS7/8 (Yr5) in 23 genotypes out of 68 wheat genotypes and by Haider et al. (2023), where Xp3000 (Yr10) marker was amplified in ten out of 45 tested wheat accessions. Cheng et al. (2022) while assessing 332 wheat germplasms for PM resistance noted that, 16 accessions amplified Pm24 gene (Xgwm337). The resistant disease reactions observed in certain genotypes, despite lacking the studied genes associated with SR and PM resistance, may be attributed to the presence of other race-specific genes not included in the study (Kokhmetova et al. 2021; Brar and Kutcher 2016). Likewise, some genotypes amplifying the genes under consideration were found susceptible in field conditions, potentially attributed to environmental factors, the new emerging pathogen races or gene-environment interactions (Ali et al 2017; Wang and Chen 2017).
Correlation analysis indicated an inverse relationship between disease incidences with GY. These findings align with those of Sharma-Poudyal and Chen (2011) and Murray et al. (1994) for SR and Draz et al. (2019) and Cerón and Martel (2003) for PM, who concluded that the proportion of leaf area, affected by SR/PM significantly contributes to yield losses in wheat. Further, the negative correlation of disease scores with TCC is indicative of cellular damage caused by the fungal pathogen to the infected plants. These findings align with Mishra et al. (2015) and Cao et al. (2009) regarding SR and PM incidence, respectively. Correlation studies between agro-morphological traits, disease responses and weather parameters are pivotal for understanding the epidemiology of the pathogen and its impact on crop productivity (Ali et al 2023; Liu et al. 2015). The findings of correlation analysis among all the traits under consideration indicate a significant contribution of the weather parameters towards disease development, consistent with previous research by EI Jarroudi et al. 2020 in SR; Mehta et al. (2018), Singh and Pannu (2014) in PM and Mehta et al. (2022), Basandrai and Basandrai 2018 and Kumar et al. 2016 in both diseases. These studies underscore the role of low temperatures and high relative humidity in promoting PM and YR incidence.
For effective resistance breeding programs, understanding the significance of the GEI is crucial for ensuring the stability of host genotype across different locations (Das et al. 2019; Sankar et al. 2021). The ‘which-won-where’ view of the GGE biplot aids in interpreting GEI in multi-environment trial data. The genotypes, positioned at distinct vertices of the polygon (convex hull), exhibit the lowest and highest SR and PM scores, thus can be categorized into best and worst performers, among the environments in that sector (Yan and Tinker 2006; Yan et al. 2007). Overall, the biplot revealed that five genotypes (DH 198, CIMMYT Entry No. 242, HPW 368, HPW 373 and DH 114) exhibited low levels of SR and PM intensities across various locations. As observed in current research, resistance sources for SR have been documented worldwide (Mehta et al. 2022; Sood et al. 2020; El-Orabey et al. 2020). Furthermore, sources of resistance to PM have been identified by various researchers (Mehta et al. 2022; Vikas et al. 2020; Draz et al. 2019; Gupta et al. 2016; HaiRong et al. 2011). After partitioning of the biplot, all the environments were categorized into three and two MEs for SR and PM, respectively. These results were in concise with Mehta et al. (2022) where the total environments were classified into three MEs for SR and PM. “Mean vs. Stability” view helps in inferring the most stable genotypes across different environments. The research findings were in alignment with the biplot studies conducted by earlier workers (Mehta et al. 2022; Das et al. 2019). In multi-environmental trials, the longer vector length indicates the greater discriminatory capability of the environments, whereas smaller angle formed by each vector with the abscissa signifies stronger representativeness of a mega-environment (Yan et al. 2007). In the study, Kukumseri exhibited a longer vector length for SR, whereas Palampur displayed a longer vector length for PM, indicating their enhanced capacity to discriminate between genotypes based on the respective diseases. Similar findings were reported in the research conducted by Mehta et al. (2022). The variation in disease intensity across different locations may be attributed to spatial evolution of pathogen pathotypes in the north-western Himalayan region or/and genetic variability among the genotypes (Aggarwal et al. 2018; Vikas et al. 2020).
The findings of our current investigation are quite promising and consistent with previous studies (Mehta et al. 2022; Vikas et al. 2020; Sood et al. 2020; Yang et al. 2017), demonstrating the development of wheat varieties showing dual resistant to SR and PM, while maintaining higher yields. In our research, we have identified wheat genotypes resistant to both diseases. This phenotypic screening was further confirmed by molecular studies using PCR amplification with a disease-specific gene-linked markers. Using GGE biplot analysis, stable genotypes with low levels of SR and PM intensities across various locations were identified. Kukumseri and Palampur have shown potential as optimal test sites for screening wheat germplasm against SR and PM, respectively. The high-yielding genotypes, possessing resistance to both diseases, can be released as varieties following multi-location trials. Furthermore, these genotypes can serve as valuable donors in future sustainable wheat improvement and can be used in molecular mapping studies and association analyses to identify and characterize new sources of resistance.
Data availability
No datasets were generated or analysed during the current study.
References
Abraha MT, Shimelis H, Solomon T, Hailu A (2019) Genotype-by-environment interaction and selection of elite wheat genotypes under variable rainfall conditions in Northern Ethiopia. J Crop Improv 33(6):1–17
Adhikari N, Joshi BP, Shrestha J, Bhatta NR (2018) Genetic variability, heritability, genetic advance and trait association study for yield and yield components in advanced breeding lines of wheat. Nepal J Agric Sci 17:229–238
Aggarwal R, Sharma S, Gupta S, Banrjee S, Bashyal BM, Bhardwaj SC (2018) Molecular characterization of predominant Indian wheat rust pathotypes using URP and RAPD markers. Ind J Biotech 17:327–336
Akin B, Zencirci N, Özseven İ (2008) Field resistance of wheat (Triticum aestivum L.) genotypes from different countries to leaf rust (Puccinia triticina). Turk J Agric for 32(6):479–486
Ali S, Rodriguez-Algaba J, Thach T, Sørensen CK, Hansen JG, Lassen P, Nazari K, Hodson DP, Justesen AF, Hovmøller MS (2017) Yellow rust epidemics worldwide were caused by pathogen races from divergent genetic lineages. Front Plant Sci 8:1057. https://doi.org/10.3389/fpls.2017.01057
Ali Y, Iqbal S, Aatif HM, Naveed K, Khan AA et al (2023) Predicting stripe rust severity in wheat using meteorological data with environmental response modeling. J King Saud Univ - Sci 35(4):102591. https://doi.org/10.1016/j.jksus.2023.102591
Alvarado G, Rodríguez FM, Pacheco A, Burgueño J, Crossa J, Vargas M, Pérez-Rodríguez P, Lopez-Cruz MA (2020) META-R: a software to analyze data from multi-environment plant breeding trials. Crop J 8(5):745–756. https://doi.org/10.1016/j.cj.2020.03.010
Bariana S, Brown GN, Ahmed NU, Khatkar S, Conner RL, Wellings CR (2002) Characterization of Triticum vavilovii derived stripe rust resistance using genetic, cytogenetic and molecular analyses and its marker-assisted selection. Theor Appl Gene 104:315–320. https://doi.org/10.1007/s001220100767
Basandrai AK, Basandrai D (2018) Powdery mildew of wheat and its management. In: Singh DP (ed) Management of wheat and barley diseases. Apple Academic Press, Canada, pp 173–181
Bennett FGA (1984) Resistance to powdery mildew in wheat: a review of its use in agriculture and breeding programmes. Plant Pathol 33:279–300
Bennett FGA, Westcott B (1982) Field assessment of resistance to powdery mildew in mature wheat plants. Plant Pathol 31:261–268
Bhardwaj SC, Singh GP, Gangwar OP, Prasad P, Kumar S (2019) Status of wheat rust research and progress in rust management-Indian context. Agronomy 9:892. https://doi.org/10.3390/agronomy9120892
Brar GS, Kutcher HR (2016) Race characterization of Puccinia striiformis f. sp. tritici, the cause of wheat stripe rust, in Saskatchewan and Southern Alberta, Canada and virulence comparison with races from the United States. Plant Dis 100(8):1744–1753
Cao X, Zhou Y, Duan X, Cheng D (2009) Relationships between canopy reflectance and chlorophyll contents of wheat infected with powdery mildew in fields. Acta Phytopathol Sinica 39(3):290–296
CerónMartel MMCIS (2003) Effects of powdery mildew severity (Blumeria graminis f. sp. tritici) on breeding lines of durum wheat (Triticum aestivum L. spp. durum) yield in western Andalusia. Span J Agric Res 1:13–18
Chatrath R, Mishra B, Ortiz Ferrara G, Singh SK, Joshi AK (2007) Challenges to wheat production in south Asia. Euphytica 157:447–456
Cheng P, Guo M, Hao X, Guo X, Yao Q, Guo Q, Li Q, Wang B (2022) Evaluation of powdery mildew resistance and molecular detection of resistance genes in an international wheat collection. Crop Prot 160:106033. https://doi.org/10.1016/j.cropro.2022.106033
Das A, Parihar AK, Saxena D, Singh D, Singha KD et al (2019) Deciphering genotype-by-environment interaction for targeting test environments and rust resistant genotypes in field pea (Pisum sativum L.). Front Plant Sci 10:825
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19:11–15
Draz IS, Esmail SM, Abou-Zeid MAE, Essa TAE (2019) Powdery mildew susceptibility of spring wheat cultivars as a major constraint on grain yield. Ann Agric Sci 64(1):39–45. https://doi.org/10.1016/j.aoas.2019.05.007
EI Jarroudi M, Lahlali R, Kouadio L, Denis A, Belleflamme A, El Jarroudi M, Boulif M, Mahyou H, Tychon B (2020) Weather-based predictive modeling of wheat stripe rust infection in Morocco. Agronomy 10:280
Elkot M, Mohammed H, El-Aziz A (2016) Molecular identification of some stem rust and yellow rust resistance genes in Egyptian wheat and some exotic genotypes. Ass J Agric Sci 47:124–135
El-Orabey WM, Ashmawy MA, Shahin AA, Ahmed MI (2020) Screening of CIMMYT wheat genotypes against yellow rust in Egypt. Int J Phytopathol 9(01):51–70
FAO (2022) Food and Agriculture Organization Corporate Statistical Database (FAOSTAT). https://www.fao.org/faostat/en/#data/QCL/ visualize. Accessed 17 Apr 2024
Gupta P, Balyan H, Sharma P, Gaurav SS, Sharma S, Kumar R et al (2022) Catalogue of gene symbols for wheat: 2022 supplement. Annu Wheat Newslett 68:68–81
Gupta V, Kumar RS, Kumar S, Mishra CN, Tiwari V, Sharma I (2016) Evaluation and identification of resistance to powdery mildew in Indian wheat varieties under artificially created epiphytotic. J Appl Nat Sci 8:565–569
Haider MW, Kaur J, Bala R et al (2023) Stripe rust resistance gene(s) postulation in wheat germplasm with the help of differentials and tagged molecular markers. Sci Rep 13:9007. https://doi.org/10.1038/s41598-023-36197-y
HaiRong Z, Peng W, Ning Z, Yu ZW, Yong CQ (2011) Identification and analysis of resistance to powdery mildew and rust in some wheat germplasm resources. J Triticeae Cr 29:925–929
Han J, Liu Y, Hou C, Li J, Wang J, Zhang Q, Yang Q, Chen X, Wu J (2020) A 1Ns disomic addition from Psathyrostachys huashanica keng confers resistance to powdery mildew in wheat. Agronomy 10(2):312
Hovmøller MS, Walter S, Bayles R, Hubbard A, Flath K, Sommerfeldt N et al (2016) Replacement of the European wheat yellow rust population by new races from the centre of diversity in the near-Himalayan region. Plant Pathol 65:402–411. https://doi.org/10.1111/ppa.12433
Jee C, Pathak VN, Verma SP, Verma OP, Singh OP (2019) Association studies for grain yield and its contributing components in diverse genotypes of wheat (Triticum aestivum L. em. Thell). J Pharmacogn Phytochem 8(3):1177–1180
Kokhmetova A, Rsaliyev A, Malysheva A, Atishova M, Kumarbayeva M, Keishilov Z (2021) Identification of stripe rust resistance genes in common wheat cultivars and breeding lines from Kazakhstan. Plants 10(11):2303. https://doi.org/10.3390/plants10112303
Kumar S, Archak S, Tyagi RK, Kumar J, Vikas VK et al (2016) Evaluation of 19,460 wheat accessions conserved in the Indian national genebank to identify new sources of resistance to rust and spot blotch diseases. PLoS ONE 11(12):e0167702
Liu N, Lei Y, Gong G, Zhang M, Wang X, Zhou Y, Qi X, Chen H, Yang J, Chang X, Liu K (2015) Temporal and spatial dynamics of wheat powdery mildew in Sichuan province, China. Crop Prot 74:150–157. https://doi.org/10.1016/j.cropro.2015.05.001
Mehta A, Basandrai AK, Banyal DK et al (2018) Effect of weather parameters on powdery mildew development of wheat at different location in Himachal Pradesh. Indian Phytopathology 71:349–353. https://doi.org/10.1007/s42360-018-0062-7
Mehta A, Basandrai D, Rana V, Dhillon HK, Basandrai AK (2022) Deciphering field resistance to powdery mildew and yellow rust among popular cultivars of wheat and set of differential lines. Indian J Genet Plant Breed 82(1):38–46. https://doi.org/10.31742/IJGPB.82.1.6
Mishra CN, Kumar S, Gupta V, Tiwari V, Sharma I (2015) Utilization of chlorophyll content index (CCI) to infer yellow rust severity in wheat (Triticum aestivum L.). J Appl Nat Sci 7(1):38–42. https://doi.org/10.31018/jans.v7i1.560
Morgounov A, Tufan HA, Sharma R, Akin B, Bagci A, Braun HJ et al (2012) Global incidence of wheat rusts and powdery mildew during 1969–2010 and durability of resistance of winter wheat variety Bezostaya. Eur J Plant Pathol 132:323–340. https://doi.org/10.1007/s10658-011-9879-y
Murphy LR, Santra D, Kidwell K, Yan G, Chen X, Campbell KG (2009) A lLinkage maps of wheat stripe rust resistance genes Yr5 and Yr15 for use in marker-assisted selection. Crop Sci 49:1786–1790
Murray GM, Ellison PJ, Watson A, Cullis BR (1994) The relationship between wheat yield and stripe rust as affected by length of epidemic and temperature at the grain development stage of crop growth. Plant Pathol 43(2):397–405
Nagarajan S (1977) Meteorology and forecasting of epidemics of diseases. In: Symposium on basic sciences and agriculture, INSA, New Delhi, 136–143
Peterson BG, Carl P (2020) Performance analytics: econometric tools for performance and risk analysis. R package version 2, Available at: https://CRAN.R-project.org/package=PerformanceAnalytics
Peterson RF, Campbell AB, Hannah AE (1948) A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can J Res 26:496–500
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/
Rani R, Singh R, Yadav NR (2019) Evaluating stripe rust resistance in Indian wheat genotypes and breeding lines using molecular markers. CR Biol 342(5–6):154–174. https://doi.org/10.1016/j.crvi.2019.04.002
Roelfs AP, Singh RP, Saari EE (1992) Rust diseases of wheat: concepts and methods of disease management. CIMMYT, Mexico D.F, p 81
Sankar SM, Singh SP, Prakash G, Satyavathi CT, Soumya SL, Yadav Y, Sharma LD, Rao AR, Singh N, Srivastava RK (2021) Deciphering genotype-by- environment interaction for target environmental delineation and identification of stable resistant sources against foliar blast disease of pearl millet. Front Plant Sci 12:656158
Sharma S, Rana V, Verma S, Gupta C, Priyanka, et al (2022) Genetic variability studies in bread wheat (Triticum aestivum L.) under multi-environment trials in northern hills zone. Biol Forum–int J 14(2):307–313
Sharma-Poudyal D, Chen XM (2011) Models for predicting potential yield loss of wheat caused by stripe rust in the US Pacific Northwest. Phytopathology 101(5):544–554
Singh G, Kumar P, Kumar R, Gangwar LK (2018) Genetic diversity analysis for various morphological and quality traits in bread wheat (Triticum aestivum L.). J Appl Nat Sci 10(1):24–29
Singh RP, Hodson DP, Jin Y, Lagudah ES, Ayliffe MA, Bhavani S, Rouse MN, Pretorius ZA, Szabo LJ, Huerta-Espino J, Basnet BR (2015) Emergence and spread of new races of wheat stem rust fungus: continued threat to food security and prospects of genetic control. Phytopathology 105(7):872–884
Singh S, Pannu PPS (2014) Influence of weather factors on occurrence and progress of powdery mildew of wheat in the screen house. Plant Dis Res 30:50–55
Slafer GA, Savin R, Pinochet D, Calderini DF (2021) Wheat. In: Calderini D, Sadras V (eds) Crop physiology case histories for major crops. Academic Press, Burlington USA, pp 98–163
Sood T, Basandrai D, Basandrai AK, Sohu VS, Rana V, Mehta A, Sharma BK, Mavi GS, Kaur J, Bains NS (2020) Stable sources of resistance to yellow rust and powdery mildew in Indian and exotic wheat germplasm. J Cereal Res 12(1):23–28
Valizadeh J, Ziaei SM, Mazloumzadeh SM (2014) Assessing climate change impacts on wheat production (a case study). J Saudi Soc Agric Sci 13(2):107–115. https://doi.org/10.1016/j.jssas.2013.02.002
Verma S, Chaudhary HK, Singh K et al (2024) Genetic diversity dissection and population structure analysis for augmentation of bread wheat (Triticum aestivum L.) germplasm using morpho-molecular markers. Genet Resour Crop Evol. https://doi.org/10.1007/s10722-023-01851-x
Vikas VK, Kumar S, Archak S, Tyagi RK, Kumar J et al (2020) Screening of 19,460 genotypes of wheat species for resistance to powdery mildew and identification of potential candidates using focused identification of germplasm strategy (FIGS). Crop Sci 60:2857–2866
Wang M, Chen X (2017) Stripe rust resistance. Str Rust 2017:353–558
Witham FH, Blaydes DF, Devlin RM (1986) Exercises in Plant Physiology 2nd Edn. PWS Publishers, Boston, USA
Xu HX, Yi YJ, Ma PT, Qie YM, Fu XY, Xu YF et al (2015) Molecular tagging of a new broad-spectrum powdery mildew resistance allele Pm2c in Chinese wheat landrace Niaomai. Theor Appl Genet 128:2077–2084
Yan W, Kang MS, Ma B, Cornelius WS, PL, (2007) GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci 47:643–653
Yan W, Tinker NA (2006) Biplot analysis of multi-environment trial data: principles and applications. Canadian J Pl Sci 86:623–645
Yang L, Zhang X, Zhang X, Wang J, Luo M et al (2017) Identification and evaluation of resistance to powdery mildew and yellow rust in a wheat mapping population. PLoS ONE 12(5):e0177905
Zhao NJ, Xue F, Wang CY, Han JR, JI WQ, Zhen L (2010) SSR analysis of powdery mildew resistance gene in Chinese wheat landrace Baihulu. J Triticeae Crop 30:411–414
Acknowledgements
The authors are grateful to the Department of Genetics and Plant Breeding, CSK Himachal Pradesh Agricultural University, Palampur for providing the wheat germplasm, facilities and financial support during the research.
Funding
The authors have not disclosed any funding.
Author information
Authors and Affiliations
Contributions
HKC conceptualized and planned the experiments; SV executed the experiments, drafted the manuscript and analyzed the data & HKC, SV, AB, KS, KSD, AP and MS provided suggestions and polished the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interests regarding the publication of this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Verma, S., Chaudhary, H.K., Badiyal, A. et al. Assessing dual resistance to stripe rust and powdery mildew in wheat germplasm through molecular and field studies across the north-western Himalayas. Euphytica 220, 128 (2024). https://doi.org/10.1007/s10681-024-03385-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-024-03385-4