
Abstract
In order to elucidate the cytological mechanism of 2n pollen formation in Chinese jujube, a cultivar named ‘Linglingzao’ (2n = 2x = 24) which produces relative more 2n pollens naturally was employed for microsporogenesis analysis. Chromosomes paired in 12 bivalents at diakinesis and the first meiotic division was normal, whereas, the second division was characterized by frequent abnormal spindle orientation (parallel and tripolar spindles). Perpendicular, tripolar, and parallel spindles at metaphase II accounted for 72.93, 17.22, and 9.85% respectively. Perpendicular and tripolar spindles led to tetrads and triads formation respectively. Two types of parallel spindles were observed and a significant association correlation (r = 0.84, P < 0.05) between parallel spindle II (fused spindles) and dyads was found which meant only fused spindles can form dyads, while parallel spindle I with a long distance between two sets of chromosomes seemed to lead tetrad formation. At tetrad stage, the observed frequencies of tetrads, triads and dyads were 80.99, 16.40, and 2.61% respectively. By the rule that each tetrad can form four n pollens, each triad can form one 2n pollen and two n pollens, each dyad can form two 2n pollens, the frequency of 2n pollen estimated was 5.71%. Based on the pollen diameter difference between n and 2n pollens, frequency of 2n pollen observed was 6.15%. No significant difference was observed between frequency of 2n pollen estimated and that of observed. The results indicated that, tripolar spindles and fused spindles at metaphase II followed by more triads and few dyads formation at tetrad stage were responsible for the production of 2n pollen in Chinese jujube. 2n gametes observed correspond to first division restitution. Research on the cytological mechanism of unreduced pollen will provide a platform for unreduced pollen induction and polyploidy breeding in Chinese jujube.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Polyploidization is widespread in plants and has played a major role in the evolution and diversification of the plant kingdom. Unreduced gametes are an important tool for polyploidization and the creation of genetic variation in plant breeding (Dewitte et al. 2009, 2010a, 2010b). Polyploidy is of great value to breeders (Dewitte et al. 2010a; Gallo et al. 2007). But among the rich germplasm resources of Chinese jujube (Z. jujuba Mill.), ‘Zanhuangdazao’ is the unique natural triploid identified up to now, which has the valued attributes of big fruit, high yield and good adaptation. Most of the cultivars of ber (Z. mauritiana Lam.), a close-related species of Chinese jujube, are tetraploid showing similar advantages. Considering the above-mentioned, polyploid induction of Chinese jujube should have attractive prospect.
Polyploidization may be asexual through somatic chromosome doubling and sexual through the formation of 2n gametes following both intra- and inter-ploidy matings (Ton et al. 1977). Asexual polyploidization (traditional chromosome duplication) is performed through the use of harmful mitosis inhibitors like colchicine, oryzalin and trifluralin (Hancock 1997; Wan et al. 1991). The major drawback of this technique is that it largely disables the recombination between chromosomes of different parents in allopolyploids. As a consequence, this form of polyploidization offers limited possibilities to create genetic variability. Sexual polyploidization is far more efficient than somatic chromosome doubling. Sexual polyploidization effected by functioning of 2n gametes in the parental species or in their hybrids has been got in ornamental plants, grain crops, potato, fruit-bearing trees and some other plants (Akutsu et al. 2007; Barba-Gonzalez et al. 2004, 2005; Carputo and Barone 2005; Carputo et al. 2003; Hayashi et al. 2009; Jauhar 2003; Lyrene et al. 2003; Ramanna et al. 2003; Sugiura et al. 2000; Van Laere et al. 2009; Van Tuyl and Lim 2003; Veilleux 1985; Yanagi et al. 2010; Zhou et al. 2008).
Advantages of 2n pollen in polyploidization have been proposed. Firstly, usage of 2n pollen can conquer homogeneity increasing, severe recession and chimaeras occurrence resulted from asexual polyploidization. Secondly, 2n pollen can simply transfer advantage genes from wild species and part-cultivars into cultivars that can be used in breeding and production directly. Thirdly, 2n gametes can transmit the parental heterozygosity to the offspring. The first division restitution (FDR) and the second division restitution (SDR) gametes can transfer at least 75–80% and 33–35% heterozygosity respectively (Camadro and Peloquin 1980; Peloquin et al. 2008). FDR gametes are particularly valuable in plant sexual polyploidization breeding programs (Bretagnolle and Thompson 1995; Ramanna 1983). Fourthly, using 2n gametes can improve self-fruitfulness much as well as the pollen fertility of triploids. Fifthly, endosperm balance number (EBN) can be conquered by 2n gametes, most breeding experiences showed that usage of 2n gametes can conquer this barrier directly and shorten the period of breeding (Johnston et al. 1980). In tuber-bearing solanums, it was reported that EBN in combination with 2n gametes maintains the ploidy integrity of diploid ancestral species, while proving the flexibility for either unilateral or bilateral sexual polyploidization (Carputo et al. 2003). Sixthly, seedless germplasm can be obtained by using 2n gametes. In citrus breeding, 2n gametes have been successfully used for the production of seedless variety, triploid from 2n female gametes (Geraci et al. 1975).
Unreduced gametes are of importance for basic studies in cytogenetics as well as applied plant breeding and facilitate the production of new polyploidy species. According to the previous observation on 223 Chinese jujube cultivars, only four cultivars (‘Linglingzao’, ‘Yingluozao’, ‘Zanhuangchangzao’, ‘Zhongyangmuzao’) produce relative high percentage of 2n pollens and it was proposed that the diameter of 2n pollen was 1.5 times of n pollen (Peng 2008). The cytological basis for the formation of 2n pollen in Chinese jujube has not been clearly understood. The aim of this research was to elucidate the cytological mechanisms that lead to 2n pollen formation and provide new insights on utilization of 2n pollen in polyploidy breeding of Chinese jujube.
Materials and methods
Plant material
‘Linglingzao’ (2n = 2x = 24) which can produce relative high frequency of 2n pollen naturally was used for cytological analysis of meiosis. All the sampled trees were grown under regular management in the field.
Cytological analysis of meiosis
During May 15 to 22, 2009, flower buds were collected and fixed in ethanol:acetic acid (3:1) fixative for 24 h, then transferred into 70% ethanol and stored in refrigerator at 4°C. Before observation, anthers were dissected from the buds with a dissecting needle, then microsporocytes were prepared by squashing anthers with a nipper, and stained with propanoic acid-iron-hematoxylin-chloral hydrate (PIHCH). Chromosome number was determined at diakinesis and all phases of meiosis were observed with light microscope OLYMPUS. Photomicrographs were taken under binocular light microscope (×1000 magnification) with image-forming system Stk ‘MDC200(LY-HPCCD)’.
Observation of 2n pollen
Anthers from opening flowers were collected and squashed in a drop of 0.5% triphenyltetrazolium chloride (TTC) solution on a microscope slide. After a minimum of 1 h, the preparation was observed under a binocular light microscope (×100 magnification). Viable pollen grains were stained into full red, whereas aborted pollen grains were unstained. More than 5000 pollen grains were measured. Pollen which is 1.5 times of n pollen in diameter was regarded as 2n pollen.
Estimation of the frequency of 2n pollen
The number of dyads, triads, and tetrads were counted respectively. The number of observed dyads, triads, and tetrads during meiosis enabled an evaluation of the theoretical frequency of 2n pollen. Based on the rule that each dyad led to two unreduced pollen grains, each triad produced one unreduced pollen grain and two reduced pollen grains, and each tetrad gave rise to four reduced pollen grains, the frequency of 2n pollen (F) was estimated as below:
D = number of dyads, Tr = number of triads, Te = number of tetrads.
Statistical analysis
Correlation analysis was carried out with Statistical Product and Service Solutions version 16 (SPSS Inc, USA).
Results
Microsporogenesis
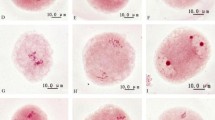
During microsporogenesis, chromosomes paired in 12 bivalents at diakinesis (Fig. 1a–d) and first meiosis was normal. However, abnormal spindle orientation (parallel and tripolar spindles) were observed at metaphase II (Fig. 1i, m, q). The frequencies of perpendicular, tripolar and parallel spindle were 72.93, 17.22, and 9.85% respectively (Table 1). While the frequencies of tetrad (Fig. 1g, h), triad (Fig. 1k, l) and dyad (Fig. 1s, t) were 80.99, 16.40, and 2.61% respectively at tetrad stage (Table 1). The correlation analysis showed that there was a significant association (r = 0.98, P < 0.01) between the frequency of tripolar spindle and that of triad (Table 2), which indicated that tripolar spindles resulted in triads. The correlation between parallel spindle and dyad and that between perpendicular spindle and tetrad were not significant, which indicated that not all the parallel spindles resulted in dyads formation.

Dyads and triads formation of Chinese jujube (Ziziphus jujuba Mill. ‘Linglingzao’). Bar 5 μm. a–d Diakinesis with 12 bivalents. e Normal perpendicular spindles. f Anaphase II with two sets of chromosomes being pulled to four poles. g–h Tetrads as a result of perpendicular spindles formation. i Tripolar spindles at Metaphase II. j–l Triads as a result of tripolar spindles formation. m Parallel spindle I with a distance between two sets of chromosomes. n Anaphase II with sets of chromosomes being separated. o Tetrads resulted from parallel spindle I. p Tetraspore. q Parallel spindle II (Fused spindle). r Tripolar, parallel and fused spindle. s – t Dyads following the formation of fused spindles
Actually, two types of parallel spindles were observed. For parallel spindle I (Fig. 1m), a long distance between the two sets of chromosomes led to tetrads formation (Fig. 1o). Differed with the regular perpendicular tetrads, the tetrads resulted from the parallel spindle I were arranged at the same division plane. For parallel spindle II (fused spindle), the two sets of chromosomes were very close, even integrated with each other (Fig. 1q). Fused spindles were similar to spindles at metaphase I. However, fused spindles could be distinguished based on the synchronization of meiosis (Fig. 1r). The correlation analysis showed a significant association (r = 0.84, P < 0.05) between the frequency of parallel spindle II and dyads (data not show). This result further indicated that fused parallel spindles (Fig. 1q) led to dyads formation (Fig. 1s, t).
2n pollen formation
Each triad gave rise to one 2n microspore and two n microspores, while each dyad produced two 2n microspores. 2n microspore formed 2n pollen grain. The frequencies of 2n pollen from different types of sporads were estimated (Table 3). The average frequency of 2n pollen from both dyad and triad was estimated as 5.71%, which was not significantly different with the observed one, 6.15% (Table 3). It indicated that the 2n pollens in ‘Linglingzao’ are resulted from the formation of triads and dyads at tetrad stage which originated from the abnormal spindle orientation at metaphase II.
Discussion
The formation of 2n gametes is a common phenomenon in the plant kingdom. 2n gametes may result from a variety of different cytological abnormalities (Bretagnolle and Thompson 1995; Harlan and De Wet 1975; Veilleux 1985).
Five main cytological mechanisms of 2n gamets formation were summarized: (i) premeiotic chromosome doubling during the process from mitosis to meiosis, disturbances appeared and resulted in chromosomes of archesporial cell being doubled followed by 2n gametes formation; (ii) meiosis disturbances, most 2n gametes were formed by this reason. Meiosis abnormalities led to 2n gametes of different types, second division with crossing-over (SDR-CO), first division with crossing-over (FDR-CO), first division restitution without crossing-over (FDR-NCO), transmitting ±40, ±80, 100% of the parental heterozygosity to the offspring, respectively (Crespel et al. 2002; El Mokadem et al. 2002); (iii) abnormal cytokinesis, in Paspalum, 52 accessions presented failure of cytokinesis during first or second meiosis and resulted in dyads and triads formation respectively, followed by 2n pollen formation (Pagliarini et al. 1999). In Brachiaria, it was reported that the failure of cytokinesis at metaphase I and total absence of cytokinesis resulted in unreduced pollen formation (Gallo et al. 2007); (iv) desynaptic mutants, desynaptic mutants of diploid potato (2n = 2x = 24) were found to produce a fairly high frequency of fertile 2n pollen grains (Ramanna 1983); (v) abnormal spindles of meiosis, this often happened in simultaneous dicotyledonous plants. Parallel or tripolar spindles during metaphase II of microsporogenesis were responsible for the production of 2n pollen. Triads formation resulted in 2n pollen formation in Brazilian gingseng (Taschetto and Pagliarini 2003) and rose (Crespel et al. 2006). In Populus tomentosa Carr, parallel, fused and tripolar spindles were considered to be leading to 2n pollen formation (Zhang and Kang 2010; Zhang et al. 2009). Parallel and tripolar spindles have been reported leading to 2n pollen formation in the wild potato Solanum okadae (Camadro et al. 2008).
Unreduced gametes formation of Chinese jujube was due to the abnormal configuration of spindles according to this research. Detailed studies have been made on pollen mother cells (PMCs) at all stages of meiosis to determine whether or not meiotic disturbances were responsible for 2n gametes formation. During the observation of microsporogenesis of Chinese jujube, chromosomes paired in 12 bivalents at diakinesis and the first meiotic division was normal, whereas the second division was characterized by frequent abnormal spindle orientation (parallel and tripolar spindles), as have been reported in some plants (Bretagnolle and Thompson 1995; Camadro et al. 2008; Veilleux 1985; Xu et al. 2008). However, the occurrence of parallel spindles at the second division does not necessarily mean that 2n pollen has been formed (Ramanna 1979; Tavoletti et al. 1991). The ‘fused spindle’ at metaphase II has been associated with the formation of dyads (Ramanna 1983; Veilleux et al. 1982; Xu et al. 2008). In this study, a significant association correlation (r = 0.84, P < 0.05) between fused spindles and dyads was found. This meant only parallel spindle II (fused spindle) can form dyads thereby result in 2n gametes, while parallel spindles I (with a long distance between two sets of chromosomes) seemed to be leading tetrad formation followed by n gametes in Chinese jujube.
Most researches indicated that formation of 2n gamete was controlled by genes (Calderini and Mariani 1997; Katsiotis and Forsberg 1995). d’Erfurth et al. (2008) isolated and characterized a gene (AtPS1) involved in the abnormal orientation of spindles at meiosis II that controlled 2n gamete formation apparently in Arabidopsis thaliana. It was reported that the mechanism of 2n pollen formation of Solanum is under control of Mendelian recessive ps (Camadro et al. 1980). Jumbo pollen formation in alfalfa is dependent on the homozygous recessive condition of the gene jp that causes complete failure of post-meiotic cytokinesis which results in 100% of 4x pollen (Tavoletti et al. 2000). Desynapsis was found to be controlled by a single recessive gene ds (Ramanna 1983). Although the production of 2n gametes is under genetic control, it can be also affected by environmental conditions. The variation of 2n pollen production could be related to environmental fluctuations (Crespel et al. 2006).
Depending on the cytological mechanisms involved in the production of 2n gametes in Chinese jujube, such gametes are generally categorized as being equivalent to FDR. FDR gametes could transmit at least 75% of parental genotype to their progenies. Further research is needed to reveal the actual level of heterozygosity of the unreduced gametes and their contributions to the progeny.
References
Akutsu M, Kitamura S, Toda R, Miyajima I, Okazaki K (2007) Production of 2n pollen of Asiatic hybrid lilies by nitrous oxide treatment. Euphytica 155:143–152. doi:10.1007/s10681-006-9317-y
Barba-Gonzalez R, Lokker AC, Lim KB, Ramanna MS, Van Tuyl JM (2004) Use of 2n gametes for the production of sexual polyploids from sterile Oriental × Asiatic hybrids of lilies (Lilium). Theor Appl Genet 109:1125–1132. doi:10.1007/s00122-004-1739-0
Barba-Gonzalez R, Lim KB, Ramanna MS, Visser RGF, Van Tuyl JM (2005) Occurrence of 2n gametes in the F1 hybrids of Oriental × Asiatic lilies (Lilium): relevance to intergenomic recombination and backcrossing. Euphytica 143:67–73. doi:10.1007/s10681-005-2657-1
Bretagnolle F, Thompson JD (1995) Gametes with the somatic chromosome number: mechanisms of their formation and role in the evolution of autopolyploid plants. New Phytol 129:1–22. doi:10.1111/j.1469-8137.1995.tb03005.x
Calderini O, Mariani A (1997) Increasing 2n gamete production of diploid alfafa by cycles of phenotypic recurrent selection. Euphytica 93:113–118. doi:10.1023/A:1002978405500
Camadro EL, Peloquin SJ (1980) The occurrence and frequency of 2n pollen in three diploid solanums from northwest Argentina. Theor Appl Genet 56:11–15. doi:10.1007/BF00264421
Camadro EL, Saffarano SK, Espinillo JC, Castro M, Simon PW (2008) Cytological mechanisms of 2n pollen formation in the wild potato Solanum okadae and pollen–pistil relations with the cultivated potato, Solanum tuberosum. Genet Resour Crop Evol 55:471–477. doi:10.1007/s10722-007-9254-1
Carputo D, Barone A (2005) Ploidy level manipulations in potato through sexual hybridisation. Ann Appl Biol 146:71–79. doi:10.1111/j.1744-7348.2005.04070.x
Carputo D, Frusciante L, Peloquin SJ (2003) The role of 2n gametes and endosperm balance number in the origin and evolution of polyploids in the tuber-bearing solanums. Genetics 163:287–294
Crespel L, Gudin S, Meynet J, Zhang D (2002) AFLP-based estimation of 2n gametophytic heterozygosity in two parthenogenetically derived dihaploids of Rosa hybrida L. Theor Appl Genet 104:451–456. doi:10.1007/s001220100695
Crespel L, Ricci SC, Gudin S (2006) The production of 2n pollen in rose. Euphytica 151:155–164. doi:10.1007/s10681-006-9136-1
d’Erfurth I, Jolivet S, Froger N, Catrice O, Novatchkova M, Simon M et al (2008) Mutations in AtPS1 (Arabidopsis thaliana Parallel Spindle 1) lead to the production of diploid pollen grains. PLoS Genet 4(11):1000274. doi:10.1371/journal.pgen.1000274
Dewitte A, Eeckhaut T, Van Huylenbroeck J, Van Bockstaele E (2009) Occurrence of viable unreduced pollen in a Begonia collection. Euphytica 168:81–94. doi:10.1007/s10681-009-9891-x
Dewitte A, Eeckhaut T, Van Huylenbroeck J, Van Bockstaele E (2010a) Induction of 2n pollen formation in Begonia by trifluralin and N2O treatments. Euphytica 171:283–293. doi:10.1007/s10681-009-0060-z
Dewitte A, Eeckhaut T, Van Huylenbroeck J, Van Bockstaele E (2010b) Meiotic aberrations during 2n pollen formation in Begonia. Heredity 104:215–223. doi:10.1038/hdy.2009.111
El Mokadem H, Crespel L, Meynet J, Gudin S (2002) The occurrence of 2n-pollen and the origin of sexual polyploids in dihaploid roses (Rosa hybrida L.). Euphytica 125:169–177. doi:10.1023/A:1015830803459
Gallo PH, Micheletti PL, Boldrini KR, Risso-Pascotto C, Pagliarini MS, do Valle CB (2007) 2n Gamete formation in the genus Brachiaria (Poaceae:Paniceae). Euphytica 154:255–260. doi:10.1007/s10681-006-9294-1
Geraci G, Esen A, Soost RK (1975) Triploid progenies from 2x × 2x cross of Citrus cultivars. J Hered 66:177–178
Hancock JF (1997) The colchicine story. HortScience 32:1011–1012
Harlan JR, De Wet JMJ (1975) On Ö Winge and a prayer: the origins of polyploidy. Bot Rev 41:361–390. doi:10.1007/BF02860830
Hayashi M, Kato J, Ohashi H, Mii M (2009) Unreduced 3x gamete formation of allotriploid hybrid derived from the cross of Primula denticulata (4x) × P. rosea (2x) as a causal factor for producing pentaploid hybrids in the backcross with pollen of tetraploid P. denticulata. Euphytica 169:123–131. doi:10.1007/s10681-009-9955-y
Jauhar PP (2003) Formation of 2n gametes in durum wheat haploids: sexual polyploidization. Euphytica 133:81–94. doi:10.1023/A:1025692422665
Johnston SA, den Nijs TPM, Hanneman RE Jr, Peloquin SJ (1980) The significance of genic balance to endosperm development in interspecific crosses. Theor Appl Genet 57:5–9. doi:10.1007/BF00276002
Katsiotis A, Forsberg RA (1995) Discovery of 2n gametes in tetraploid oat Avena vaviloviana. Euphytica 81:1–6. doi:10.1007/BF00022452
Lyrene PM, Vorsa N, Ballington JR (2003) Polyploidy and sexual polyploidization in the genus Vaccinium. Euphytica 133:27–36. doi:10.1023/A:1025608408727
Pagliarini MS, Takayama SY, de Freitas PM, Carraro LR, Admowski EV, Silva N, Batista LAR (1999) Failure of cytokinesis and 2n gamete formation in Brazilian accessions of Paspalum. Euphytica 108:129–135. doi:10.1023/A:1003660327223
Peloquin SJ, Boiteux LS, Simon PW, Jansky SH (2008) A chromosome-specific estimate of transmission of heterozygosity by 2n gametes in potato. J Hered 99:177–181. doi:10.1093/jhered/esm110
Peng B (2008) Study on 2n pollen in Chinese jujube. Dissertation, Agricultural University of Hebei
Ramanna MS (1979) A re-examination of the mechanisms of 2n gamete formation in potato and its implications for breeding. Euphytica 28:537–561. doi:10.1007/BF00038921
Ramanna MS (1983) First division restitution gametes through fertile desynaptic mutants of potato. Euphytica 32:337–350. doi:10.1007/BF00021442
Ramanna MS, Kuipers AGJ, Jacobsen E (2003) Occurrence of numerically unreduced (2n) gametes in Alstroemeria interspecific hybrids and their significance for sexual polyploidisation. Euphytica 133:95–106. doi:10.1023/A:1025652808553
Sugiura A, Ohkuma T, Choi YA, Tao R, Tamura M (2000) Production of nonaploid (2n = 9x) Japanese persimmons (Diospyros kaki) by pollination with unreduced (2n = 6x) pollen and embryo rescue culture. J Am Soc Hortic Sci 125(5):609–614
Taschetto OM, Pagliarini MS (2003) Occurrence of 2n and jumbo pollen in the Brazilian ginseng (Pfaffia glomerata and P. tuberosa). Euphytica 133:139–145. doi:10.1023/A:1025683511162
Tavoletti S, Mariani A, Veronesi F (1991) Cytological analysis of macro- and micro-sporogenesis of a diploid alfalfa clone producing male and female 2n gamete. Crop Sci 31:1258–1263
Tavoletti S, Pesaresi P, Barcaccia G, Albertini E, Veronesi F (2000) Mapping the jp (jumbo pollen) gene and QTLs involved in multinucleate microspore formation in diploid alfalfa. Theor Appl Genet 101:372–378. doi:10.1007/s001220051493
Ton PM, Nijs D, Peloquin SJ (1977) Polyploid evolution via 2n gametes. Am Potato J 54:377–386. doi:10.1007/BF02852758
Van Laere K, Leus L, Van Huylenbroeck J, Van Bockstaele E (2009) Interspecific hybridisation and genome size analysis in Buddleja. Euphytica 166:445–456. doi:10.1007/s10681-008-9844-9
Van Tuyl JM, Lim KB (2003) Interspecific hybridisation and polyploidisation as tools in ornamental plant breeding. Acta Hortic 612:13–22
Veilleux R (1985) Diploid and polyploid gametes in crop plants: mechanisms of formation and utilization in plant breeding. Plant Breed Rev 3:253–288
Veilleux RE, McHale NA, Lauer FI (1982) 2n gametes in diploid Solanum: frequency and types of spindle abnormalities. Genome 24:301–314. doi:10.1139/g82-032
Wan Y, Duncan DR, Rayburn AL, Petolino JF, Widholm JM (1991) The use of antimicrotubule herbicides for the production of doubled haploid plants from anther-derived maize callus. Theor Appl Genet 81:205–211. doi:10.1007/BF00215724
Xu LQ, Zhang QL, Luo ZR (2008) Occurrence and cytological mechanism of 2n pollen formation in Chinese Diospyros spp. (Ebenaceae) staminate germplasm. J Hortic Sci Biotech 83(5):668–672
Yanagi T, Hummer KE, Iwata T, Sone K, Nathewet P, Takamura T (2010) Aneuploid strawberry (2n = 8x + 2 = 58) was developed from homozygous unreduced gamete (8x) produced by second division restitution in pollen. Sci Hortic 125:123–128
Zhang ZH, Kang XY (2010) Cytological characteristics of numerically unreduced pollen production in Populus tomentosa Carr. Euphytica 173:151–159. doi:10.1007/s10681-009-0051-0
Zhang JF, Wei ZZ, Li D, Li BL (2009) Using SSR markers to study the mechanism of 2n pollen formation in Populus × euramericana (Dode) Guinier and P. × popularis. Ann For Sci 66:506. doi:10.1051/forest/2009032
Zhou SJ, Ramanna MS, Visser RGF, van Tuyl JM (2008) Analysis of the meiosis in the F1 hybrids of Longiflorum × Asiatic (LA) of lilies (Lilium) using genomic in situ hybridization. J Genet Genomics 35:687–695. doi:10.1016/S1673-8527(08)60091-0
Acknowledgments
The authors want to thank the National Key Technology R&D Program (2007BAD36B07, 2008BAD92B03), the Provincial Natural Science Fund of Hebei (C2007000452) and Agricultural Scientific Research Achievement Transformation Program (2010GB2A200027) for financial assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Xue, Z., Liu, P. & Liu, M. Cytological mechanism of 2n pollen formation in Chinese jujube (Ziziphus jujuba Mill. ‘Linglingzao’). Euphytica 182, 231–238 (2011). https://doi.org/10.1007/s10681-011-0461-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-011-0461-7