
Abstract
Organic farming has gained in importance in Germany during recent years. Therefore an increasing demand exists for varieties with specific adaptation to this farming system. In the present study we therefore conducted comparative field experiments with modern maize breeding materials under organic versus conventional farming conditions (ORG and CON, respectively) to estimate quantitative genetic parameters needed for developing optimal breeding strategies and to investigate the perspectives of selection for specific adaptation to ORG. Starting from two broad samples of elite germplasm, consisting of 178 flint and dent lines, respectively, fractions of 11 flint and 11 dent lines were selected based on their testcross performance under ORG. A corresponding set of lines was selected under CON. Testcross performance was evaluated in three regions of Germany and selection of superior lines was practiced across two stages in 2004 and 2005, respectively. The specifically selected lines were crossed in a factorial manner for production of experimental inter-pool single-cross hybrids which were field-tested under ORG and CON in two regions in 2006. Average grain yields were about 16% lower under ORG than under CON. Variance components and entry-mean heritability coefficients under ORG largely resembled those obtained under CON. Phenotypic correlations between ORG and CON were moderate for grain yield and strong for grain dry matter content. No consistent estimates were obtained for the corresponding genotypic correlation for grain yield. At the first stage of testcross selection no evidence of specific adaptation to ORG or CON was observed whereas the factorial crosses tested in 2006 displayed distinct, yet non-significant, advantages when evaluated under the respective target farming system. A small top fraction of hybrids showed outstanding performance under both ORG and CON. The chances of detecting such broadly adapted genotypes are increased if ORG test sites are included in the regular testing system.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Organic agriculture has gained in importance in Germany during the past 20 years. As a consequence, there is a growing demand for plant varieties meeting the requirements of this farming system. Developing such varieties may require specific testing conditions and alternative breeding strategies. Extensive breeding work has already been carried out in the field of bio-dynamic agriculture (Kunz 2000). Generally, these activities have been conducted under organic farming conditions, only. First results of comparative yield trials under high input, low input, and organic farming conditions in wheat were published by Baresel (2006). Also in wheat, Löschenberger et al. (2008) suggested specific features of a breeding strategy targeting organic farming.
In maize (Zea mays L.), to our knowledge, no comparative studies on breeding varieties with specific adaptation to organic versus conventional farming (ORG and CON, respectively) have yet been reported. First results on the estimation of quantitative genetic parameters in both systems were recently published by Lorenzana and Bernardo (2007). They conducted their research in Minnesota, USA, using testcrosses of a sample of recombinant inbred lines derived from the hybrid B73 × Mo17. Based on the parameter estimates, the authors concluded that a separate breeding program for ORG may not be needed. No such experiments have been reported from European maize research groups.
In the present study we conducted field experiments with modern Central European maize breeding materials under ORG as well as CON to estimate quantitative genetic parameters needed for developing optimal breeding strategies and to investigate the perspectives of selection for specific adaptation to ORG as well as for broad adaptation to both ORG and CON. More specifically, the objectives of the present study were.
-
(i)
to assess the amount of genetic variation observed in the two farming systems,
-
(ii)
to compare the precision of field trials under these systems,
-
(iii)
to estimate correlation coefficients as criteria for the agreement between the performing ability of modern maize under ORG and CON,
-
(iv)
to investigate whether specific adaptation to ORG can be achieved by continued selection under ORG,
-
(v)
to find out the best strategy of breeding for broad adaptation to both ORG and CON.
Materials and methods
Genetic materials
The starting material consisted of two genetically broad based samples of doubled haploid (DH) dent and flint lines taken from the most advanced KWS breeding program for Germany in that year. Each of the two samples comprised 178 lines. The dent lines were derived from 110 single crosses among 30 parent lines and the flint lines from 43 three-way crosses among 20 parent lines. Generally 1 or 2 dent lines and 3–5 flint lines were taken from each cross. The dent lines were crossed with a flint tester and the flint lines with a dent tester. Testers were single crosses between unrelated elite lines (Table 1).
Based on testcross performance in 2004, the best 44 dent lines each under ORG and CON were selected. Seventeen of them were superior under both conditions leading to 88 − 17 = 71 dent lines in total. These were crossed with two elite flint inbred lines as testers. The latter were unrelated with each other and with the single-cross tester used at the first selection stage. In the flint lines, selection was practised analogously. Twenty-one lines were overlapping resulting in 88 − 21 = 67 selected flint lines in total. These were tested with two elite dent inbred lines. The latter originate from the same single cross and both of them are related (coancestry: f = 0.25) with the single-cross tester used the year before. Due to lack of sufficient seed, a few testcrosses in each group could not be included in the yield trials (Table 1).
Based on testcross performance in 2005, the top 11 dent and flint lines each under ORG and CON were selected for production of two sets of inter-pool (flint × dent) hybrids being specifically adapted to ORG and CON, respectively. It was attempted to produce 77 crosses per set according to a partially balanced factorial mating design (Melchinger 1984). However only 46 “ORG hybrids” and 44 “CON hybrids” had enough seed for the field experiments.
At all selection stages, selection was based on an index (I) composed of grain yield (GY) and dry matter content (DMC) according to the formula I = GY + 2.5 DMC, where GY is measured in 100 kg ha−1 and DMC in 10 g kg−1.
Field experiments
Genetic materials were evaluated in nine field experiments under ORG and CON in three or two regions of Germany between 2004 and 2006 (Tables 1 and 2). All organically managed trials were carried out on fields which are certified according to EC regulation 2092/91. As a consequence, the conventionally managed trials could not be placed side by side to the organically managed ones. Distances between experimental fields within a region varied between 2 and 15 km. Each experiment was laid out as a lattice design with two replicates. “Free” entries were filled up with appropriate checks. Two-row plots with a row distance of 75 cm were used throughout. In the Einbeck and Erding regions the trials were sown to a final stand of 9 and 11 plants m−2, respectively. In the Hohenheim region the trials were overplanted and thinned to 9 plants m−2 in the 6- to 8-leaf stage. Field emergence was fast and even in all environments. General characteristics of the test sites are given in Table 2.
Statistical analyses
Analyses of variance were first performed for each lattice experiment separately (Cochran and Cox 1957). Adjusted entry means were used in combined analyses across regions within farming systems and years. Variance components were then calculated according to Snedecor and Cochran (1980) and entry-mean heritability coefficients were estimated from the variance components of the combined analyses. Confidence intervals of the estimated heritability coefficients were calculated according to Knapp and Bridges (1987). Coefficients of genotypic correlation and their standard errors were calculated according to Mode and Robinson (1959). All statistical computations were performed with the PLABSTAT software package (UTZ 2004).
Results
Average grain yield under organic farming was 16% lower than under conventional farming. Inferiority ranged from 8.4% in Exp. 3 to 21.4% in Exp. 6 (Table 3). In 2004 differences were smaller than in 2005 and 2006. In all nine experiments, grain dry matter content was slightly lower under ORG than under CON.
Coefficients of genotypic (r g) and phenotypic (r p) correlation between ORG and CON for grain yield varied greatly between experiments. Estimates ranged from 0.08 to 0.63 for r p and from 0.07 to 1.66 for r g. In only four experiments, r g exceeded twice its standard error. In contrast, estimates of r p for grain dry matter content were high to very high in all experiments, and those of r g were close to one.
Most estimates of the genotypic variance and all of the genotype × location interaction variance for grain yield were significant at P = 0.05 or 0.01 (Table 4). On average, estimates of the genetic component of variance were of similar size under ORG and CON, but great differences existed between experiments, and limited agreement was found between ORG and CON at a single-experiment basis. In most experiments, the genotype × location interaction variance was 1.5 to 3 times larger than the genotypic variance, and there was good agreement between ORG and CON for the interaction variance. Error variances were generally about 50% higher than the interaction variances and were also of similar magnitude under ORG and CON.
Heritability coefficients for grain yield were moderate to low in the eight testcross experiments as well as in Exp. 9 containing the specifically selected hybrids (Table 5). On average, the same level of heritability was obtained under ORG and CON. In five experiments the estimates were larger under ORG than under CON and in the remainder four the reverse was true. However, the differences were generally small and in no case significant.
The first stage of testcross selection for performance under ORG and CON, respectively, did not result in specific adaptation as judged from the performance of the selected fractions in the next year (2005) and with other testers (Table 6). However, the experimental hybrids built up from the best dent and flint lines after the second selection stage indicated a distinct, yet statistically non-significant, degree of differentiation: The hybrids developed under ORG performed better than those developed under CON when tested under ORG. Likewise the “CON hybrids” performed better than the “ORG hybrids” under CON. The respective differences in relative grain yield amounted to 2.2% and 1.6%, respectively.
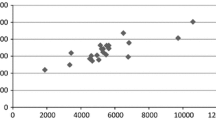
The beginning of a divergent selection process resulting in specifically adapted groups of hybrids can also be recognized in Fig. 1. It demonstrates that a hybrid excelling under ORG may be only mediocre or even fail under CON and vice versa. On the other hand, it also appears that there is a small top fraction of hybrids showing outstanding performance under both ORG and CON.

Relationship between grain yields (g m−2) under organic and conventional farming of two groups of experimental hybrids selected under organic (ORG-hybrids) and conventional farming (CON-hybrids) conditions; means across two regions 2006 (r p = coefficient of phenotypic correlation; *,** significantly different from zero at the 0.05 and 0.01 probability levels; a excluding the outlying dot in the upper left part of the graph)
Discussion
Under organic farming, plants have to comply with specific stress conditions requiring a number of characteristics which are less important under CON. For instance, lack of seed treatment and pesticides application demands high germination ability and early vigour as well as disease and pest resistance and superior competitiveness against weeds. Moreover, omitting mineral nitrogen (N) fertilizers frequently leads to N deficiency in spring when low temperatures retard N mineralization from organic fertilizers (Mengel and Kirkby 2001). High N-use efficiency therefore is an important component of a maize plant’s adaptedness to ORG. Taken together, more yield limiting factors may be expected under ORG compared to CON. Indeed the present study revealed lower grain yields under ORG in all nine experiments (Table 3) and in all three regions (data not shown). This is in agreement with results of variety trials in Germany (Meyercordt and Mücke 2006) and with Lorenzana and Bernardo’s (2007) study in Minnesota.
Experiments under ORG are often assumed to be less precise than if they were grown in high- input CON environments. This is not confirmed by our results. Estimates of both error variances and heritability coefficients were in the same order of magnitude under ORG and CON. Accordingly, similar genetic gains can be expected from selection under the two farming systems. This was confirmed by the response realized at the first stage of testcross selection (data not shown). Assuming that genotype × year and line × tester interaction effects largely cancel when averaged across many genotypes, we calculated the realized response to selection by contrasting the relative testcross performance of the candidate lines evaluated in 2004 with that of the selected fractions tested in 2005. As a reference basis for computing the relative values we used the average testcross performance of four elite dent lines and three elite flint lines which had been topcrossed with the same flint resp. dent testers as the candidate lines. Averaged across experiments, the realized response for grain yield amounted to 4.8% under ORG and 5.6% under CON on the flint side (Exps. 1 and 2 vs. Exps. 5 and 6). On the dent side, a respective comparison did not make sense because of bias due to green-snapping damage in Exps. 7 and 8.
Presterl et al. (2003) reviewed a comprehensive series of experiments under low versus high soil N supply in Germany and North France. As in the present study, the authors found similar precision measures and heritability coefficients under various levels of N-deficiency stress. However, on highly heterogeneous soils the precision of experiments may be more impaired under stress than under conventional high input conditions (Brun and Dudley 1989; Bänziger et al. 1997; Bertin and Gallais 2000). On the other hand, the genetic variance may be greater under stress than non-stress conditions which may counterbalance an increased error variance and even lead to higher heritability under low-input conditions (Ceccarelli 1994; Lafitte and Edmeades 1994; Agrama et al. 1999).
An important question for breeders is whether selection under CON will sufficiently improve performance under ORG or, put in other words, whether indirect (correlated) selection for adaptation to ORG is as effective as direct selection. A quantitative genetic criterion for the efficiency of indirect selection is the ratio of the expected indirect (CR) to direct (R) gain from selection (Falconer and Mackay 1996):
In this formula, i is the standardized selection differential (under CON and ORG, respectively), h is the square root of the heritability coefficient, σg is the genotypic standard deviation and r g is the genotypic correlation coefficient. If the same selection intensity is practiced under ORG and CON, and if the heritability coefficients are equal as well, the efficiency of indirect selection only depends on the genotypic correlation between the two farming systems. This actually applies to the present study (Table 4). However, the estimated coefficients of the genotypic correlation between ORG and CON (Table 3) have to be interpreted with caution since they refer to populations with complicated substructures and since the ORG and CON experiments could not be performed side by side in the same experimental field. This may explain the great variation in the estimates obtained from the different experiments and may be a reason for the much lower estimates of the phenotypic correlation coefficients (Table 3). In view of the above considerations we assume that the true genotypic correlation coefficients are distinctly lower than r g = 1 which means that direct selection for performance under ORG is expected to be more effective than indirect selection. Following the same reasoning, the analogous conclusions apply to direct vs. indirect selection for performance under CON. The superiority of the ORG-SEL hybrids under ORG and the CON-SEL hybrids under CON in Exp. 9 (Table 6) agrees with this interpretation. In contrast, Lorenzana and Bernardo (2007) found a higher heritability under CON and a higher genetic correlation (r g ≥ 0.8) between ORG and CON for grain yield. Thus they concluded that direct and correlated selection for adaptation to ORG may be similarly effective. Possible reasons for the lack of agreement between the two studies include differences in the genetic materials (narrow in the US and broad based in the present study), differences in the soil nutrient supply (yield reductions under ORG were much smaller in the US study), and differences between the macro-environments in which the experiments were conducted.
Surprisingly, no specific adaptation to ORG or CON was observed in the selected flint lines tested in 2005 (Table 6) although selection in 2004 was effective under both farming systems (see above). Possibly the testers used in 2005 masked genetic variation for traits causing specific adaptation.
An important result of this study relates to green snapping, i.e. the breakage of the stem below the ear-bearing node at the end of the vegetative growth phase under heavy storm. Green snapping had a much greater impact under CON than under ORG. This was specifically true for one of the hybrids which performed excellently under ORG but failed completely under CON (upper left dot in Fig. 1).
Conclusions
Results of the present study demonstrate that modern European dent and flint maize genotypes may reach remarkable grain yields under ORG. Estimated variance components and heritability coefficients indicate that considerable genetic gains can be achieved under each of the two farming systems. Moderate phenotypic and highly erratic genotypic correlations between ORG and CON point at strong genotype × farming system interactions. Apparently, specific yield associated characteristics are necessary to reach maximum performance under ORG and CON, respectively. Such traits include germination vigour, competitiveness to weeds, and high N-use efficiency under ORG on the one hand, and resistance to green snapping and early root lodging under CON on the other. Thus, developing varieties with specific adaptation to ORG may be more promising if breeding is carried out under ORG than under CON. Analogously, if hybrids with adaptation to both farming systems are aimed at, an adequate number of ORG sites should be included in the regular testing system.
References
Agrama HAS, Zakaria AG, Said FB, Tuinstra M (1999) Identification of quantitative trait loci for nitrogen use efficiency in maize. Mol Breed 5:187–195
Baresel JP (2006) Weizenzüchtung für den Ökologischen Landbau. Diss TU München, Germany
Bänziger M, Betran FJ, Lafitte HR (1997) Efficiency of high-nitrogen selection environments for improving maize for low-nitrogen target environments. Crop Sci 37:1103–1109
Bertin P, Gallais A (2000) Genetic variation for nitrogen use efficiency in a set of recombinant maize inbred lines I. Agrophysiological results. Maydica 45:53–66
Brun EL, Dudley JW (1989) Nitrogen response in the USA and Argentina of corn populations with different proportions of flint and dent germplasm. Crop Sci 29:565–569
Ceccarelli S (1994) Specific adaptation and breeding for marginal conditions. Euphytica 77:205–219
Cochran WG, Cox G (1957) Experimental designs. Wiley, New York
Falconer DS, Mackay TFC (1996) Introduction to quantitative genetics, 4th edn. Longman, Essex, UK
Knapp SJ, Bridges WL Jr (1987) Confidence interval estimates for heritability for several mating and experimental designs. Theor Appl Genet 73:759–763
Kunz P (2000) Backqualität und/oder Brotqualität? Lebendige Erde 5:38–41
Lafitte HR, Edmeades GO (1994) Improvement for tolerance to low soil nitrogen in tropical maize. II. Grain yield, biomass production, and N accumulation. Field Crops Res 39:15–25
Lorenzana RE, Bernardo R (2007) Genetic correlation between corn performance in organic and conventional production systems. North Central Regional Corn Breeding Research Meeting, St. Charles, IL, 21–22 March 2007. http://corn2.agron.iastate.edu/NCR167/Abstracts/Default.html. Cited 01 December 2007
Löschenberger F, Fleck A, Grausgruber H, Hetzendorfer H, Hof H, Lafferty J, Marn M, Neumayer A, Pfaffinger G, Birschitzky J (2008) Breeding for organic agriculture: the example of winter wheat in Austria. Euphytica (this issue)
Melchinger AE (1984) Analysis of incomplete factorial mating designs. Vorträge für Pflanzenzüchtung 7:131–150
Mengel K, Kirkby EA (2001) Principles of plant nutrition, 5th edn. Kluwer Academic Publishers, Dordrecht, Boston, London
Meyercordt A, Mücke M (2006) Ergebnisse der Sortenversuche Öko-Silomais und - Körnermais 2006 der Landwirtschaftskammer Niedersachsen. http://basis.zadi.de/VIPDOKVOLLPFLANZEN/DDD/4c788.pdf. Cited 01 December 2007
Mode CJ, Robinson HF (1959) Pleiotropism and the genetic variance and covariance. Biometrics 15:518–537
Presterl T, Seitz G, Landbeck M, Thiemt EM, Schmidt W, Geiger HH (2003) Improving nitrogen-use efficiency in European maize - estimation of quantitative genetic parameters. Crop Sci 43:1259–1265
Snedecor GW, Cochran WG (1980) Statistical methods, 7th edn. Iowa State University Press, Ames, IA, USA
Utz HF (2004) PLABSTAT, Version 3Awin. Universität Hohenheim, Institut für Pflanzenzüchtung, Saatgutforschung und Populationsgenetik, Stuttgart
Acknowledgements
The study would not have been possible without the efforts of A. Altenweger, H. Hilscher, O. Hübner, J. Jesse, S. Koch, E. Löhnhardt, H. Meyer, U. Mund, Mrs. and Mr. Lanzinger, Mr. Pfanzelt, G. Wegener, and the support staff of the experimental stations, who carefully managed the field experiments and assisted in data collection. The authors are grateful for financial support from “Bundesministerium für Verbraucherschutz, Ernährung und Landwirtschaft, Bundesprogramm Ökologischer Landbau” (Kenn-Nr.03OE651), Bonn, and from KWS SAAT AG, Einbeck.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Burger, H., Schloen, M., Schmidt, W. et al. Quantitative genetic studies on breeding maize for adaptation to organic farming. Euphytica 163, 501–510 (2008). https://doi.org/10.1007/s10681-008-9723-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-008-9723-4