
Abstract
This work aimed to analyze the chemical composition and evaluate the anti-inflammatory and antinociceptive properties of the essential oil obtained from the leaves of Lantana montevidensis (OEFLM) in mice. The chemical composition of the OEFLM was analyzed by GC–MS, and its effects on the central nervous system were determined by the open field and rotarod tests. The antinociceptive and anti-inflammatory screenings were conducted using the formalin test, as well as the acetic acid-induced abdominal contortion assay. The antinociceptive activity was assessed by the tail-flick and hot plate tests. The systemic anti-inflammatory activity was determined using models of paw edema induced by carrageenan, dextran, histamine, or arachidonic acid. The effects of the essential oil on vascular permeability and granuloma formation were also investigated. The chemical composition of the OEFLM revealed the presence of valencene (17.93%), bicyclogermacrene (16.35%), trans-caryophyllene (12.84%) and germacrene D (12.18%). The administration of OEFLM at the dose of 2000 mg/Kg caused no evident toxicity. While OEFLM (25, 50, 100, and 200 mg/kg) did not present significant anxiolytic-loke or sedative effects, it showed promising antinociceptive and anti-inflammatory activities. The results of this study indicate that the OEFLM has analgesic and anti-inflammatory activities in animal models by inhibiting acute and chronic inflammation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Inflammation is a protective response of the body against tissue injury, which aims to remove the aggressor agent, initiating the tissue repair process (Netea et al. 2018; Pahwa et al. 2020). Although the inflammatory response is an essential physiological mechanism, the intense and continuous release of inflammatory mediators can significantly damage tissues, causing pain and frequently evolving into a chronic process (Adedayo et al. 2019; Drexler et al. 2008). In this context, inflammatory pain is caused by the production and secretion of mediators such as prostaglandins, pro-inflammatory cytokines, and chemokines that activate nociceptors in the inflammatory site (Matsuda et al. 2019).
Nonsteroidal anti-inflammatory drugs (NSAIDs) are among the most commonly used medications to treat inflammation and pain worldwide. However, especially in the long term, these drugs can cause significant side effects, inducing damage to organs such as the stomach, kidneys, bronchi, as well as to the cardiovascular system (Wongrakpanich et al. 2018; Zapata‐Morales et al. 2016). Therefore, the search for new drugs with greater efficacy and safety has been increasing, placing medicinal plants as important sources of bioactive molecules (Olorukooba and Odoma, 2019).
Essential oils are plant-derived substances rich in secondary metabolites whose biological activities have been consistently demonstrated (Bakkali et al. 2008; Ribeiro-Santos et al. 2017). The Verbenaceae is a family composed of 35 genera and about 1.200 species (Xu and Chang, 2017), in which the production of biologically active essential oils is associated with the presence of secret trichomes (Favorito 2009). The species Lantana montevidensis (Spreng) Briq., popularly known as "chumbinho", is used in Brazilian folk medicine for the treatment of rheumatism, bronchitis, and gastric disorders (Nagao et al. 2002). Accordingly, previous research has demonstrated that this species has biological activities such as antibacterial (Barreto et al. 2010; Costa et al. 2011; De Sousa et al. 2013; Sousa e Costa, 2012), antiproliferative (against adenocarcinoma, carcinoma, and melanoma) (Nagao et al. 2002), anti-inflammatory, antinociceptive, antipyretic (Makboul et al. 2013), antioxidant (Barros et al. 2017; Makboul et al. 2013; De Oliveira et al. 2019), insecticide (Weverton et al. 2019) and acaricide (De Sousa et al. 2020). Nevertheless, the anti-inflammatory and antinociceptive effects of this species remain to be better investigated.
Therefore, based on the traditional use of Lantana montevidensis, as well as the biological activities already proven in the literature, the present study characterized for the first time, the antinociceptive and anti-inflammatory effects of the essential oil obtained from the leaves of Lantana montevidensis. Also, study highlights the importance of biological conservation, for the use of natural products with pharmacological activity.
2 Materials and methods
2.1 Collection and extraction of the essential oil from leaves of Lantana montevidensis (OEFLM)
The leaves of Lantana montevidensis were collected in the botanical garden of the Regional University of Cariri-URCA (Brazil) with the coordinates 7°22′ S; 39°28′ W at 492 m above the level of the sea. A voucher specimen was identified by prof. Drª. Maria Arlene Pessoa da Silva and registered in the Herbarium Caririense Dárdano de Andrade Lima (HCDAL/(Brazil), under number 12.790. The extraction of the essential oil was carried out using a Clevenger apparatus, obtaining a yield of 0.0582%.
2.2 Analysis of the chemical profile of the essential oil
The chemical analysis of the OEFLM was performed using a Shimadzu QP-2010 gas chromatograph connected to a mass spectrometer (GC–MS). The analyses were carried out in an Agilent DB-5MS column (30 m × 0.25 mm × 0.25 μm) using helium (99.99%) as a carrier gas at a constant flow of 1.1 ml.min−1; injection volume: 1.0 μL; 1:10 injector split ratio; injector temperature of 250 °C; electron impact mode at 70 eV; ion source temperature of 280 °C and transfer line temperature 260 °C. The quantitative analysis was performed using the internal standard addition method (alkanes C8H18–C40H82) injected under the same experimental conditions as the samples. The identification of the constituents was performed by comparing their retention indices (RI) values and their mass spectral fragmentation pattern with those reported in the literature (Adams 2007) and equipment database (Wiley 7 lib and NIST 08 lib) using the Kovats index. The data were processed using the Shimadzu GC–MS Solution software.
2.3 In vivo tests
2.3.1 Chemical reagents and drugs
Arachidonic acid, histamine, carrageenan, dextran, Evans blue, indomethacin, morphine, dexamethasone, acetic acid, and formalin were purchased from Sigma-Aldrich (St. Louis, MO, USA); xylazine and ketamine were purchased from Ceva Santé Animale (Paulínia, SP, Brazil). All solutions were prepared immediately before use and administered orally (p.o.) or intraperitoneally (i.p.) according to the animal weight (0.01 mL/g body weight) and specific experimental protocol. The negative control group received injectable water (0.01 mL/g, p.o.), while the OEFLM was diluted in 5% Tween-80 aqueous solution.
2.3.2 Animals and ethical aspects of the study
For in vivo analysis, male and female Swiss mice (Mus musculus) weighing between 20 and 30 g were kept in polypropylene boxes at room temperature (23 ± 2 °C) with food (Labina, Purina ®) and water ad libitum, under a light /dark cycle of 12 h. The Commission of Ethics in Research in Animals (CEUA/Brazil) of the Regional University of Cariri approved all protocols described in this study (reference number 00029/2017.1).
2.3.3 Determination of non-clinical acute toxicity of the OEFLM.
The assessment of the acute toxicity was performed according to the guidelines of the Organization for Economic Cooperation and Development (OECD) (Guideline 2001), with adaptations, as well as according to the table of Malone e Robichaud (Malone and Robichaud 1962). The animals were divided into two groups (n = 3). The control group received injectable water (Control; 0.01 mL/g, p.o.), and the OEFLM-treated group received a single dose of 2000 mg/kg (p.o.). Following the treatments, the animals were observed at 30, 60, 120, 180 and 360 min and every 24 h for 14 days.
2.3.4 Evaluation of the effects of the OEFLM on the central nervous system (CNS)
2.3.4.1 The open field test
The animals (n = 3/group) were treated with injectable water (Control: 0.01 mL/g, p.o.) or OEFLM (2000 mg/Kg) or diazepam (DZP, 5 mg/Kg, i.p.). The motor capacity of the animals was analyzed in a 30 cm side open field, with 9 squares of 10 cm side, for 5 min, and the number of rearing (vertical holding), grooming (self-cleaning), and crosses was determined (Lapa et al. 2003).
2.3.4.2 The rotarod test
Mice were selected through sessions with 3 min of duration, before treatment. Thus, those mice who remained during that period in the rotating bar were chosen. Animals (n = 3/group) were treated with injectable water (control: 0.01 mL/g; p.o.) or OEFLM (2000 mg/kg; p.o) or diazepam (DZP; 5 mg/kg; i.p.) and then evaluated for both number of falls and time of permanence in the rotating bar (Dunham and Miya 1957).
2.3.5 Evaluation of antinociceptive screening of OEFLM
2.3.5.1 Acetic acid-induced abdominal contortions
Mice (n = 6/group) were treated orally with water (control: 0.01 mL/g) or OEFLM (25, 50, 100 or 200 mg/Kg) or indomethacin (10 mg/Kg) 1 h before receiving an intraperitoneal injection of 0.6% acetic acid. After administration of acetic acid, the number of abdominal contortions was recorded for each animal over 30 min (Lapa et al. 2003).
2.3.5.2 The formalin test
Mice (n = 6/group) were treated orally with injectable water (Control; 0.01 mL/g) or OEFLM (25,50, 100, 200 mg/Kg) or indomethacin (10 mg/Kg). Following 1 h of the treatments, a 2.5% formalin solution (20 μl) was administered into the intraplantar region via the right paw's ventral surface. The nociceptive response (licking and/or paw bite) was observed from 0 to 5 min (first Phase/Neurogenic Phase) and 15–30 min (second Phase/Inflammatory Phase) after formalin injection (Tjølsen et al. 1992).
2.3.5.3 The tail-flick test
Mice (n = 6/group) were orally treated with water (0.01 mL/g, control), morphine (5 mg/Kg) or OEFLM (25, 50, 100 or 200 mg/Kg). Each animal was placed in a suitable container (acrylic tube, with ventilation), with the tail's dorsal surface to the thermal stimulus. The heating was applied to the distal third of the tail, which consisted of focusing a light beam at 50 °C on these animals' tails. The time (in seconds) to remove the tail is considered the latency time, measured automatically by a recorder connected to the apparatus. The latency time was measured before the treatments (baseline) and at the times of 30, 60, 90, and 120 min after treatment (Lapa et al. 2003).
2.3.5.4 The hot plate test
Mice (n = 6/group) were pretreated with water (0.01 mL/g, p.o.), morphine (5 mg/Kg, s.c.) or OEFLM (25, 50, 100 and 200 mg/Kg; p.o.). The nociceptive response was evaluated by the latency time (s) until the animals licked the paw or jumped from the hot plate. The readings were performed at the times of 30, 60, 90, 120, 180 min after the corresponding treatment (Lapa et al. 2003).
2.3.6 Evaluation of acute anti-inflammatory activity of the OEFLM
The anti-inflammatory activity was analyzed using murine models of acute and chronic inflammation. The essential oil activity in acute responses was evaluated through paw edema models induced by different stimuli, including carrageenan, dextran, histamine, and arachidonic acid. The vascular permeability of the animals was assessed by Evans blue extravasation. The animals (n = 6/group) were treated orally with water (0.01 mL/g), indomethacin (10 mg/Kg), promethazine (6 mg/Kg), dexamethasone (5 mg/Kg) or OEFLM (25, 50, 100 or 200 mg/Kg). In the paw edema protocol, the animals had the initial volume (Vi) of both paws measured by plethysmometry. Following the treatments and challenges, the volumes were measured according to the time established in each protocol. The results were expressed as a percentage of edema at each time using the following formula: (Vpd-Vpe / Vpe) × 100, in which: Vpd = volume of the right paw; Vpe = volume of the left paw.
2.3.6.1 Carrageenan-induced paw edema
Mice (n = 6/group) had the initial volume of the right and left hind paws evaluated by plethysmometry. Subsequently, the animals were pretreated orally with water (0.01 mL/g), indomethacin (10 mg/Kg) or OEFLM (25, 50,100 or 200 mg/Kg). After 1 h, the animals received an intraplantar injection of carrageenan (20 μl, 1% (w/v)) in the right paw and saline (20 μl) in the left paw. The volume of the hind paws of each animal was recorded at times 1, 2, 3, 4 h after the application of carrageenan (Lapa et al. 2003).
2.3.6.2 Dextran-induced paw edema
Mice (n = 6/group) had the initial volume of the right and left hind paws evaluated by plethysmometry. Subsequently, the animals were pretreated orally with water (0.01 mL/g), promethazine (6 mg/kg) or OEFLM (25, 50, 100 or 200 mg/kg). After 1 h, the animals received an intraplantar injection of dextran 1% (w/v) and saline (20 μl) in the left paw. The volume of each animal's hind paws was recorded at times 1, 2, 3, 4 h, after the administration of dextran (Lapa et al. 2003).
2.3.6.3 Histamine-induced paw edema
Mice (n = 6/group) had the initial volume of the right and left hind paws evaluated by plethysmometry. Subsequently, the animals were pretreated orally with water (0.01 mL/g), promethazine (6 mg/Kg) or OEFLM (50 mg/Kg). After 1 h, the animals received an intraplantar injection of 1% (w/v) histamine (20 μl/paw) in the right hind paw and saline (20 μl) in the left paw. The hind paw volume of each animal was recorded 30, 60, 90, and 120 min after the histamine challenge (Maling et al. 1974).
2.3.6.4 Arachidonic acid-induced paw edema
Mice (n = 6/group) had the initial volume of the right and left hind paws evaluated by plethysmometry. Subsequently, the animals were pretreated orally with water (0.01 mL/g), indomethacin (10 mg/Kg) or OEFLM (50 mg/Kg). After 1 h, the animals received an intraplantar injection of 1% (w/v) arachidonic acid (20 μl / paw) in the right hind paw and saline (20 μl) in the left paw. Each animal's hind paw volume was recorded 15, 30, 45, and 60 min after the administration of arachidonic acid (DiMartino et al. 1987).
2.3.6.5 Evaluation of vascular permeability by the Evans blue extravasation method
Mice (n = 6/group) were orally treated with water (0.01 mL/g), indomethacin (10 mg/Kg) or OEFLM (25, 50, 100 or 200 mg/Kg). A group of animals was neither treated nor challenged (negative control). Immediately after treatments, the animals received an injection of Evans blue (0.2 mL/ animal) in the retro-orbital plexus. After 1 h of the treatments, the animals were intraperitoneally administered with 1% carrageenan. After 4 h, the animals were euthanized, and the peritoneal cavities were washed with 3 mL of PBS buffer. The peritoneal fluid was collected, the samples were centrifuged for 2 min at 6000 rpm, and the supernatants were read using a spectrophotometer at 520 nm (Lapa et al. 2003).
2.3.6.6 Evaluation of the myeloperoxidase activity
The myeloperoxidase activity (MPO) was assessed using samples of the peritoneal fluid according to the technique described by Bradley and Colleagues (Bradley et al. 1982), using a 0.0005% hydrogen peroxide solution as the substrate for MPO. A MPO unit was defined as the amount capable of converting 1 μmol of hydrogen peroxide to water in 1 min at 22° C. The variation of the optical density of the sample mixture with the o-dianisidine solution as a function of the reaction time was read in a spectrophotometer at 600 nm. The results were expressed as UMPO/μl.
2.3.7 Evaluation of the OEFLM activity in a chronic inflammatory model
To investigate the effectiveness of the OEFLM in chronic inflammation, we used a murine model of cotton pellet-induced granuloma. To this end, mice were anesthetized with xylazine (20 mg/Kg) and ketamine (80 mg/Kg). Four cotton pellets weighing 10 mg (0.01 g) were then introduced in the back of each animal. The animals were treated by an oral administration of water (0.01 mL/g), dexamethasone (5 mg/Kg) or OEFLM (50 mg/Kg), daily until the 10th day. On the 11th day, the animals were euthanized, and the pellets were removed from the dorsal region and placed in a drying oven for 24 h at 37° C and weighed. Then, a homogenate was prepared, and total proteins were quantified by the biuret method. In this method, the biuret reagent reacts with the proteins present in the sample generating a purple coloration proportional to the protein concentration. After 10 min, the samples were read using a spectrophotometer at 520 nm (Swingle and Shideman 1972).
2.4 Statistical analysis
All data were analyzed using the GraphPad Prism program (v. 6.0). The differences were compared by ANOVA (one-way or two-way), followed by Tukey’s post hoc test. Differences with a value of p < 0.05 were considered as significant (Fig. 1).
3 Results
3.1 Analysis of the chemical profile of the essential oil
The analysis of the chemical composition of the OEFLM revealed the presence of valencene (17.93%), bicyclogermacrene (16.35%), trans-caryophyllene (12.84%), and germacrene D (12.18%) as major constituents, in addition to several minor compounds, as shown in Table 1. Figure 2 represents the chromatogram (CG-MS) of the essential oil obtained from the leaves of Lantana montevidensis.

Photograph of Lantana montevidensi leaves

Chromatogram of the essential oil obtained from the leaves of Lantana montevidensis (CG-MS)

Chemical structures of the major constituents of the OEFLM
3.2 Non-clinical acute toxicity of the OEFLM
According to the parameters established by Malone; Robichaud, 1962, the animals treated with OEFLM at the dose of 2000 mg/Kg showed no signs of toxicity. In addition, this dose caused no mortality, indicating that the oil presented no evident toxicity at these conditions. Then, we consider the down limit of 10% of the LD50, as recommended by the OECD (2008), to select the doses used in this study (25, 50, 100, and 200 mg/Kg) (Fig. 3).
3.3 Characterization of the effects of the OEFLM on the CNS
An oral administration of the OEFLM (2000 mg/Kg) did not alter (p > 0.05 vs. Control) the behavioral reactions (rearing, grooming and crossing) of the mice (Fig. 4a–c), indicating that in this test, the oil showed no evident anxiolytic or sedative activity. In addition, the oral treatment of the animals with OEFLM (2000 mg/Kg) did not affect the motor coordination compared to the control (Fig. 4d).

Central nervous system effects of OEFLM on mice behavior in the open field test and rotarod tests. a Number of rearing (NR), b number of Grooming (NG), c number of crossings (NC), and d effect of the OEFLM on the motor coordination on the mice in the rotarod performance test. The data were expressed as mean ± S. E. M (standard error of the mean). Analysis by one-way ANOVA followed by the Tukey's test (a1p < 0.05; a3p < 0.001 vs control)
3.4 Screening of analgesic activity
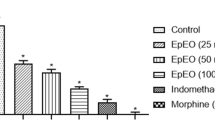
In the first phase of the formalin test (neurogenic phase), the administration of OEFLM at 25, 50 or 100 mg/Kg (p.o) significantly reduced the formalin-induced nociceptive behavior by 71.5% (p < 0.001), 84.58% (p < 0.0001), and 62.15% (p < 0.01), respectively. On the other hand, the administration OEFLM at 200 mg/kg (p.o.), as well as indomethacin (10 mg/Kg. p.o.), did not inhibit (p > 0.05) the formalin-induced nociceptive behavior compared to the negative control. Nevertheless, the second (inflammatory) phase of the test, all doses of the oil, as well as indomethacin significantly (p > 0.05) reduced the nociceptive response, suggesting that the oil presented an analgesic effect that is associated to anti-inflammatory mechanisms (Fig. 5a).

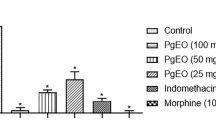
Characterization of the analgesic activity of the OEFLM in mice. a) formalin test to phase I and II and b) abdominal writhes response induced by acetic acid. The data were expressed as mean ± S. E. M (standard error of the mean). Analysis by one-way ANOVA followed by the Tukey's test (a1p < 0,05;a2p < 0,01; a3p < 0,001; a4p < 0,0001 vs negative control)
A single oral administration of the OEFLM at 25, 50, 100, and 200 mg/Kg significantly reduced the number of abdominal writhes induced by acetic acid by 79.53% (p < 0.0001), 63% (p < 0.001), 79.53% (p < 0.0001), and 88.19% (p < 0.0001), respectively (Fig. 5b). These results were significantly (p < 0.05) comparable to the effect of indomethacin (10 mg/Kg), which significantly reduced (p < 0.001 vs. control) by 66.54% the number of abdominal contortions (Fig. 5b).
3.5 Evaluation CNS antinociceptive activity of the OEFLM by tail-flick and hot plate tests
To evaluate central nervous system participation in the analgesic effect of the essential oil, a group of animals was orally treated with the doses of 25, 50, 100, or 200 mg/Kg. None of these doses increased the latency time compared with the negative control group both in the hot plate (Fig. 6a) and tail-flick (Fig. 6b) tests. On the other hand, the animals treated with morphine showed a greater latency time in comparison with the untreated animals.

Antinociceptive effect of the OEFLM in the nociceptive response triggered by a) tail-flick and b) hot plate test. The data were expressed as mean ± S. E. M (standard error of the mean). Analysis by one-way ANOVA followed by the Tukey's test (a4p < 0.0001 vs negative control)
3.6 Effects of the OEFLM on paw edema models induced by carrageenan and dextran
At the dose of 25 mg/kg (p.o.), the OEFLM decreased (p < 0.01 vs. negative control) by 60.51% the paw edema from 1 h after the carrageenan challenge (Fig. 7a). The other doses of the oil also presented different degrees of inhibition between the 2nd and 4th hours after the carrageenan challenge. Indomethacin (10 mg/kg; p.o.), used as a control drug, inhibited the edema at all time points of analysis. Together, these results indicate that the essential oil could inhibit edema formation in response to the inflammatory reaction triggered by carrageenan.

Effect of the OEFLM on pow edema formation a) carrageenan-induced paw edema and b) dextran-induced paw edema. The data were expressed as mean ± S. E. M (standard error of the mean). Analysis by two-way ANOVA followed by the Tukey's test. (a1p < 0,05; a2p < 0,01; a3p < 0,001; a4p < 0,0001 vs. negative control)
The analysis of dextran-induced paw edema (Fig. 7b) shows that all doses of OEFLM exerted an antidematogenic activity. Compared with the group of challenged and untreated animals, the doses of 25, 50, 100, and 200 mg/Kg (p.o.) reduced the edema by 30.29% (p < 0.01), 55.19% (p < 0.0001), 50.56% (p < 0.0001), and 32, 87% (p < 0.01), respectively, in the first hour after the challenge. At the second hour after the challenge, these doses caused inhibitions of 30.44% (p < 0.01), 57.32% (p < 0.0001), 45.91% (p < 0.0001), and 43.62% (p < 0.0001), respectively. At the third hour, these treatments caused inhibitions of 45.35% (p < 0.001), 79.72% (p < 0.0001), 52.97% (p < 0.0001), and 45.35% (p < 0.001), respectively. However, at the 4th hour, only the doses of 25 or 50 or 200 mg/Kg caused significant inhibition of the edema induced by dextran. Promethazine, a histamine H1 receptor antagonist, significantly inhibited the edema at all time intervals evaluated in this study.
3.7 Mechanism involved in the anti-inflammatory action of the OEFLM
To investigate the effect of the OEFLM on specific inflammatory pathways, the animals were challenged with histamine and treated with the oil (50 mg/Kg, p.o) or promethazine (6 mg/Kg; p.o.). An intraplantar administration of histamine to untreated animals induced significant edema formation within 2 h after the challenge, with a peak at 30 min. Both treatments significantly inhibited the edema at all time points analyzed. While the OEFLM showed maximal inhibition of 92.08% (p < 0.0001) in the last interval (120 min), promethazine showed maximum inhibition in the first range of 80.13% (p < 0.0001) compared to the histamine-stimulated and vehicle-treated group. (Fig. 8a).

Mechanism involved in effect of the OEFLM a) histamine-triggered paw edema; b) arachidonic acid-triggered paw edema c) carrageenan-induced vascular permeability; and d) mieloperoxidase activity. The data were expressed as mean ± S. E. M (standard error of the mean). Analysis by one-way ANOVA followed by the Tukey's test. (a1p < 0,05; a2p < 0,01; a3p < 0,001; a4p < 0,0001 vs. negative control)
To evaluate the participation of specific inflammatory pathways, the animals were challenged with arachidonic acid and treated orally with OEFLM (50 mg/ kg, p.o) or indomethacin (10 mg/ kg; p.o.). An intraplantar administration of arachidonic acid to untreated animals induced significant edema within 2 h after challenge, with a peak at 30 min. Both OEFLM and indomethacin significantly inhibited the edema with maximal inhibition of 76,32% (p < 0.0001) and 80,51% (p < 0.0001), respectively, in the last interval (120 min), compared to the arachidonic acid-stimulated and vehicle-treated group (Fig. 8b).
The effects of the essential oil treatment on vascular changes were assesses through the Evans blue method. An intraperitoneal administration of 1% carrageenan in the animals caused a significant increase in Evans blue extravasation compared with non-stimulated animals, indicating increased vascular permeability. A single oral treatment with OEFLM at doses of 50, 100, 200 mg/kg or indomethacin (10 mg/kg) significantly reduced vascular permeability compared to vehicle-treated animals. However, the dose of 25 mg/kg of OEFLM did not significantly affect the permeability compared to the control group, indicating that, under these conditions, the highest doses were more effective in reducing carrageenan-induced vascular permeability (Fig. 8c).
To investigate a potential inhibition of leukocyte activation by the essential oil, we analyzed the myeloperoxidase activity. Although no significant difference (p > 0.05) between the untreated animals and those treated with the essential oil, the treated animals presented lower levels of myeloperoxidase activity compared with the group of animals challenged with carrageenan and treated with saline (Fig. 8d). In contrast, the treatment with indomethacin was found to significantly inhibit myeloperoxidase activity compared to the negative control group. These findings suggest that antiedematogenic effects of the essential oil occur due to interference with vascular mechanisms, although its effects on leukocyte recruitment and activation remain to be further investigated.
3.7.1 Effects of the OEFLM on granuloma formation
The oral treatment with the OEFLM (50 mg/kg) or dexamethasone (5 mg/kg) significantly reduced the granuloma induced by the implantation of cotton pellets. Compared with the group of untreated animals, the weight of the dried pellets obtained from treated animals with OEFLM or dexamethasone were significantly reduced 65.42% (p < 0.001) and 88.55% (p < 0.0001), respectively (Fig. 9a). In addition, the homogenates of these pellets presented reduction in the protein content of 76.36% (p < 0.0001) and 82.56% (p < 0.0001), respectively, in comparison with the negative control group, indicating inhibition of granuloma formation (Fig. 9b). However, the effects of the treatment with the OEFLM on other aspects of the granuloma, including the cellular content, and tissue changes remain to be characterized.

Effect of the OEFLM on cotton pellet-induced granuloma. a) Dry mass of the pellets and b) protein content by the biuret method. The data were expressed as mean ± S. E. M (standard error of the mean). Analysis by one-way ANOVA followed by the Tukey's test (a3p < 0,001; a4p < 0,0001 vs. negative control)
4 Discussion
The analysis of the chemical composition of OEFLM revealed the presence of major compounds such as valencene (17.93%), bicyclogermacrene (16.35%), trans-caryophyllene (12.84%), and germacrene D (12.18%). These compounds are chemical markers of the genus Lantana and are found in other species such as L. caatingensis (de Aguiar et al. 2015), L. camara (Chowdhury et al. 2007; Costa et al. 2009). L. montevidensis, as well as other species of the genus Lantana, present variations in the chemical composition of their essential oils. Such differences, can be attributed to biotic and abiotic intrinsic factors, such as genetic specificity or extrinsic, such as environmental conditions, which can cause chemical variation depending on the time and site of collection (Facanali et al. 2009).
The biological properties of the components of this oil have been previously described in the literature. Previous research demonstrated that valencene presented antioxidant (Liu et al. 2012) and anti-inflammatory activities in models of sepsis induced by cecal ligation and puncture (CLP) (Tsoyi et al. 2011) and reduced the levels of pro-inflammatory cytokines and chemokines in atopic dermatitis model (Yang et al. 2016). In addition, this compound inhibited TRPV1 and ORAI 1 ion channels, as well as a UV-induced melanogenesis in melanoma cells (Nam et al. 2016). A study by Marques et al. (2019), showed that valencene was able to suppress: pro-inflammatory enzymes, such as iNOS and NADH and NADPH oxidase; mediators (TNF-α and IL-1α), as well as NF-κB signaling.
Trans-caryophyllene, also known as β-caryophyllene or (E)-caryophyllene or caryophyllene, has an anti-inflammatory activity reported in the literature with results similar to those used in the present study (de Morais Oliveira-Tintino et al. 2018). In addition, this compound was able to inhibit the concentration of cytokines: TNF-α, IL-β, IL-6, NO, and PGE2, expressions of mRNA and proteins of iNOS and COX-2, as well as inhibition of the activation of the tlr4/NF-кB signaling in neuroinflammation in bv-2 microglia cells (Hu et al. 2017). A study conducted by Zhang et al. (2017) demonstrated that trans-caryophyllene inhibited the expression of the VCAM-1 protein and macrophage infiltration levels on the aortic surface in vivo and in vitro assay.
With regard to the chemical composition, the findings of the present research are in accordance with previous studies, which found a similar chemical composition for the essential oil of L. montevidensis, including concerning the presence of (E)-caryophyllene, germacrene D, and bicyclogermacrene (De Sousa et al. 2013; Montanari et al. 2011).
The administration of OEFLM to mice did not promote significant behavioral changes in the open field and rotarod tests. The open field model is used to identify both the anxiolytic and sedative effects of a given substance. While anxiolytic effects are identified by an increase in the parameters of the test, substances with sedative effects cause a decrease in these parameters (Prut and Belzung, 2003). On the other hand, the rotarod test is used to assess the effect of substances on neuromotor functions as well as to identify a CNS depression action (de Moraes Pultrini et al. 2006). Thus, since the OEFLM treatment did not cause behavioral or motor changes in these models, there is no evidence that the essential oil exhibits significant toxicity to the central nervous system.
An intraperitoneal injection of acetic acid inducing abdominal contortions has been widely employed to investigate the action of compounds with anti-inflammatory and antinociceptive activity. This method is sensitive to substances both with central or peripheral actions, and therefore, it is a nonspecific test (Brito da Matta et al. 2011). The pathophysiological mechanisms involved in the acetic acid action include the release of inflammatory mediators, such as histamine, serotonin, bradykinin, cytokines (TNF-α, IL-1β, and IL-8), which orchestrate inflammatory and pain responses (Coura et al. 2012; Mohamad et al. 2010; Xie et al. 2014). The results of the present study demonstrated that OEFLM significantly inhibited the development of abdominal contortions suggesting that the components of this essential oil may reduce the production or interfere with the biological actions of inflammatory mediators released in response to acetic acid.
Other species of the genus Lantana, including Lantana camara (Silva et al. 2015) and Lantana trifolia (Silva et al. 2005), have already been shown to be effective in inhibiting acetic acid-induced responses in this model. Accordingly, β-caryophyllene, a major constituent of L. trifolia, was found to inhibit abdominal contortions in this model, indicating that it is, at least in part, one of the constituents responsible for the pharmacological activity of this species (de Morais Oliveira-Tintino et al. 2018). In addition, previous studies demonstrated that valencene and trans-caryophyllene van inhibit the release of inflammatory mediators (Yang et al. 2016; Hu et al. 2017; Marques et al. 2019), indicating that they can contribute to the analgesic/anti-inflammatory properties of the essential oil of L. montevidensis.
Following the analysis by the acetic acid test, we investigated the analgesic properties of the essential oil in the formalin test, which more specifically evaluates the participation of central and anti-inflammatory mechanisms in the antinociceptive activity of drugs (Bonjardim et al. 2012). The formalin test comprises two distinct phases. The first, also known as the nociceptive phase, occurs within the first 5 min after the formalin injection. This phase is characterized by the stimulation of nociceptors present in the C type and part of the Aδ type fibers in response to glutamate and substance P. The second, also known as the inflammatory phase, occurs between 15 and 30 min after the formalin injection. This phase is characterized by a nociceptive response triggered by the action of pro-inflammatory mediators such as adenosine, bradykinin, histamine, prostaglandins, and serotonin (Denny et al. 2013; dos Santos et al. 2010). A single oral administration of OEFLM significantly inhibited the formalin-induced nociception in both phases, suggesting that the constituents of the essential oil may interfere with neurogenic and inflammatory mechanisms associated with the nociceptive responses. However, de Morais Oliveira-Tintino et al. 2018 demonstrated that β-caryophyllene was shown to be more effective in inhibiting the second phase of the formalin test.
The tail-flick and hot plate tests are well-established models to evaluate the antinociceptive activity of substances that interfere with neurogenic mechanisms at a central level. In this test, substances with analgesic activity cause an increase in the latency time. Nevertheless, these tests evaluate distinct mechanisms, since the stimulus in the tail-flick test induces a spinal response, whereas in the hot plate test the response is supraspinatus (Bannon and Malmberg, 2007; Rezaee-Asl et al. 2014). Because OEFLM did not show an antinociceptive effect in these models, it is suggested that this substance does not have analgesic effects at a central nervous system level.
An intraplantar injection of carrageenan is usually known to cause an inflammatory response associated with the production of ROS and inflammatory mediators that orchestrate the tissue changes observed in acute inflammation (Li et al. 2018). In the paw edema test, the administration of carrageenan triggers a two-step response. In the first phase, the activation of resident cells results in the release of mediators that increase vascular permeability, such as histamine, bradykinin, serotonin, and prostaglandins. On the other hand, in the second phase, the migration and activation of leukocytes amplify the inflammatory response, causing additional edema peaks (de Brito et al. 2013; Vendramini-Costa et al. 2015). In the present study, the oral treatment with the essential oil obtained from the leaves of Lantana montevidensis significantly inhibited the paw edema induced by carrageenan. Earlier reports demonstrated that Lantana trifólia (Silva et al. 2005; Uzcátegui et al. 2004) and Lantana camara (Gidwani et al. 2009) inhibited the edema formation in this model, corroborating the data of the present research.
The dextran-induced paw edema is orchestrated by the sudden release of preformed mediators. In this context, histamine induces vascular changes through the activation of H1 receptors. In addition, in combination with serotonin, this mediator contributes to the activation of various cell types, leading to neutrophil recruitment and production of protein mediators (Ribeiro et al. 2014; Silva et al. 2014). As observed in carrageenan-induced paw edema, the administration of OEFLM caused a significant inhibition of dextran-induced edema. Because no difference was observed between the doses, we chose the dose of 50 mg/Kg to continue the experiments. Since histamine is one of the first mediators released in the inflammatory site (Benly, 2015), it is assumed that the constituents of the oil could act in the initial phase of the inflammatory reaction, possibly by directly impairing the vascular changes orchestrated by this mediator. On the other hand, histamine stimulates the production of platelet activating factor (PAF), interleukin 8 (IL-8), and leukotriene B4 (LTB4), which in turn cause neutrophil recruitment and contribute to increased vascular permeability (Mostafa et al. 2010; Tamaddonfard et al. 2012). Nevertheless, because of the variety of constituents present in the oil, multiple steps of the inflammatory cascade could potentially be affected by the treatment with the OEFLM.
The biosynthesis of eicosanoids (Yui et al. 2015) from the arachidonic acid (AA) (Dennis and Norris, 2015) is an important step in edema formation. The metabolism of this fatty acid is catalyzed by three main enzymatic pathways: cyclooxygenase (COX), lipoxygenase (LO), and cytochrome P450 (Dennis and Norris, 2015; Wang et al. 2005). The activation of the COX pathway, resulting in the production of prostaglandins and thromboxanes, and the activation of 5- LO, generating leukotrienes, contributes significantly to the development of acute inflammation (Huang et al. 2016; Kawahara et al. 2015). Given the importance of these pathways for developing inflammatory reactions, the model of arachidonic acid administration has been used in the search for new anti-inflammatory substances (Bulani et al. 2011; Kumar and Jain, 2014). In fact, one of the most important products of the AA metabolism is the eicosanoid PGE2. This mediator is involved in several cellular and molecular inflammatory events, including those associated with vascular changes and edema formation (Ray et al. 2015). As observed in the present study, the dose of 50 mg /Kg of OEFLM inhibited AA-induced edema, suggesting that the treatment inhibited the synthesis or the action of eicosanoids in the acute inflammatory phase. Since compound trans-caryophyllene has demonstrated the ability to inhibit the production of inflammatory mediators, including PGE2 (Hu et al. 2017), it is hypothesized that this compound contributes to the antiedematogenic effects of the OEFLM.
Increased vascular permeability resulting in protein extravasation is one of the first events of acute inflammation (Gehlen et al. 2004). This event is caused by the combined action of several mediators such as histamine, cytokines, and arachidonic acid products (Martins et al. 2017). A simple way to analyze this event in vivo is through the Evans blue method. As the dye binds to plasma proteins, its concentration can be quantified by spectrophotometry (Cruz et al. 2016). In the present study, the OEFLM reduced the transudate infiltration, as evidenced by a decrease in Evans blue concentrations, suggesting that this oil decreased vascular permeability in vivo. This result corroborates the data obtained in the paw edema models since a decrease in permeability justifies an inhibition of the edema induced by different stimuli. Additionally, this phenomenon may be related to the performance of compounds such as valencene, which inhibited the production of cytokines and chemokines in vitro (Yang et al. 2016; Marques et al. 2019), and trans-caryophyllene, which was found to reduce the production of mediators such as TNF-α, IL-β, IL-6, NO, and PGE2, in addition to inhibiting the expression of iNOS and COX-2 (Hu et al. 2017).
Myeloperoxidase (MPO) is an enzyme expressed by leukocytes such as neutrophils and other phagocytic cells. An increase in MPO activity is associated with leukocyte activation and contributes to a series of events triggered by inflammatory agents (Cruz et al. 2016; Khan et al. 2017). Although the treatment with the OEFLM did not significantly inhibit MPO activity in vivo, the group of animals treated with the essential oil presented reduced levels of MPO activity. These findings suggest that antiedematogenic effects of the essential oil occur due to interference with vascular mechanisms, although its effects on leukocyte recruitment and activation remain to be further investigated.
Animal models of granuloma formation have been effectively used to study the influence of substances on chronic inflammation (Ray et al. 2015). These models allow the investigation of key parameters of chronic inflammation, such as transudates, exudates, and cell proliferation (Ramirez et al. 2013). In cotton pellet-induced granuloma, the inflammatory events are divided into three phases: the first one begins 3 h after introduction of the cotton pellets, characterized by increased vascular permeability and fluid extravasation; the second (exudative) phase occurs between 3 and 72 h with protein release for the granuloma; the third and last (proliferative) phase occurs from 3 to 6 days, with the formation of granulomatous tissue (Patil and Patil, 2017). The OEFLM treatment reduced the mass of the dried pellets and the total proteins that were evaluated in the homogenate of the pellets, which possibly contributed to an inhibition of the formation of the granuloma, suggesting that the oil may have a modulating effect in the development of chronic inflammatory reactions.
5 Conclusion
The findings of the present study indicate the oral treatment with the essential oil of the leaves of Lantana montevidensis presented analgesic effects at the peripheral level and demonstrated antiedematogenic and anti-inflammatory activity in acute inflammation and chronic inflammation models. Importantly, the oral treatment with this essential oil induced no evident toxicity and did not present central antinociceptive activity. However, further research is required to characterize the effects of individual constituents, as well as to determine the molecular mechanisms underlying the anti-inflammatory, analgesic, and antidematogenic action of the OEFLM.
Abbreviations
- AA:
-
Arachidonic acid
- ANOVA:
-
Analysis of variance
- CEUA:
-
Commission of Ethics in Research in Animals
- COX:
-
Cyclooxygenase
- CNS:
-
Central nervous system
- DZP:
-
Diazepam
- GC–MS:
-
Gas chromatography-mass spectrometry
- HCDAL:
-
Herbarium Caririense Dárdano de Andrade Lima
- IL-1β:
-
Interleukin-1 β
- IL-8:
-
Interleukin-8
- LOX:
-
Lipoxygenase LTB4, leukotriene B4
- MPO:
-
Myeloperoxidase
- OECD:
-
Organization for Economic Cooperation and Development
- OEFLM:
-
Essential oil of leaves of Lantana montevidensis
- PAF:
-
Platelet activating factor
- PBS:
-
Phosphate buffered saline
- PGE2 :
-
Prostaglandin E2
- p.i.:
-
Per os (intraperitoneal administration)
- p.o.:
-
Per os (oral administration)
- TNF-α:
-
Tumor necrosis factors alpha
- SEM:
-
Standard error of the mean
- URCA:
-
Regional University of Cariri
References
Adams, R. P. (2007). Identification of essential oil components by gas chromatography/mass spectrometry. Allured publishing corporation.
Adedayo, L. D., Ojo, A. O., Awobajo, F. O., Adeboye, B. A., Adebisi, J. A., & Bankole, T. J., et al. (2019). Methanol extract of Cola nitida ameliorates inflammation and nociception in experimental animals. Neurobiology of Pain , 5, 100027. https://doi.org/10.1016/j.ynpai.2019.100027.
Bannon, A., & Malmberg, A. (2007). Models of Nociception: Hot-Plate, Tail-Flick, and Formalin Tests in Rodents, Current protocols in neuroscience/editorial board, Jacqueline N. Crawley ... [et al.]. https://doi.org/10.1002/0471142301.ns0809s41
Benly, P. (2015). Role of histamine in acute inflammation. Journal of Pharmaceutical Sciences and Research, 7, 373.
Bonjardim, L. R., Cunha, E. S., Guimarães, A. G., Santana, M. F., Oliveira, M. G. B., & Serafini, M. R., et al. (2012). Evaluation of the anti-inflammatory and antinociceptive properties of p-cymene in mice. Zeitschrift für Naturforsch. C, 67, 15–21.
Bradley, P. P., Christensen, R. D., & Rothstein, G. (1982). Cellular and extracellular myeloperoxidase in pyogenic inflammation. Blood, 60, 618–622.
Brito da Matta, C. B., De Souza, É. T., De Queiroz, A. C., De Lira, D. P., De Araújo, M. V., Cavalcante-Silva, L. H. A., et al. (2011). Antinociceptive and anti-inflammatory activity from algae of the genus Caulerpa. Marine Drugs, 9, 307–318.
Bulani, V., Biyani, K., Kale, R., Joshi, U., Charhate, K., Kumar, D., & Pagore, R. (2011). Inhibitory effect of Calotropis gigantea extract on ovalbumin-induced airway inflammation and Arachidonic acid induced inflammation in a murine model of asthma. IInternational Journal of Biomedical Science, 1, 19–25.
Chowdhury, J. U., Nandi, N. C., & Bhuiyan, M. N. I. (2007). Chemical composition of leaf essential oil of Lantana camara L. from Bangladesh. Bangladesh Journal of Botany, 36, 193–194.
Costa, J. G. M., de Sousa, E. O., Rodrigues, F. F. G., de Lima, S. G., & Braz-Filho, R. (2009). Composição química e avaliação das atividades antibacteriana e de toxicidade dos óleos essenciais de Lantana camara L. e Lantana sp. Brazilian Journal of Pharmacognosy, 19, 710–714.
Coura, C. O., de Araújo, I. W. F., Vanderlei, E. S. O., Rodrigues, J. A. G., Quinderé, A. L. G., & Fontes, B. P., et al. (2012). Antinociceptive and anti-inflammatory activities of sulphated polysaccharides from the red seaweed Gracilaria cornea. Basic & Clinical Pharmacology & Toxicology, 110, 335–341.
Cruz, M.P., Andrade, C.M.F., Silva, K.O., de Souza, E.P., Yatsuda, R., Marques, L.M., David, J.P., David, J.M., Napimoga, M.H., & Clemente-Napimoga, J.T. (2016). Antinoceptive and anti-inflammatory activities of the ethanolic extract, fractions and flavones isolated from Mimosa tenuiflora (Willd.) Poir (Leguminosae). PLoS One 11, e0150839.
de Aguiar, U.N., de Lima, S.G., Rocha, M. dos S., Citó, A.M. das G.L., Sousa, A.J.P., Silva, R.M., Silva, I.S.A., da Costa, J.G.M., 2015. Chemical composition and modulation of antibiotic activity of essential oil of Lantana caatingensis M. (Verbenaceae). Industrial Crops and Products 74, 165–170. https://doi.org/10.1016/j.indcrop.2015.04.011
de Brito, T. V., da Prudêncio, R., & S., Sales, A.B., Vieira Júnior, F. das C., Candeira, S.J.N., Franco, Á.X., Aragão, K.S., Ribeiro, R. de A., Ponte de Souza, M.H.L., & Chaves, L. de S. (2013). Anti-inflammatory effect of a sulphated polysaccharide fraction extracted from the red algae H ypnea musciformis via the suppression of neutrophil migration by the nitric oxide signalling pathway. Journal of Pharmacy and Pharmacology, 65, 724–733.
de Moraes Pultrini, A., Galindo, L. A., & Costa, M. (2006). Effects of the essential oil from Citrus aurantium L. in experimental anxiety models in mice. Life Sciences, 78, 1720–1725.
de Morais Oliveira-Tintino, C. D., Pessoa, R. T., Fernandes, M. N. M., Alcantara, I. S., da Silva, B. A. F., & de Oliveira, M. R. C., et al. (2018). Anti-inflammatory and anti-edematogenic action of the Croton campestris A. St.-Hil (Euphorbiaceae) essential oil and the compound β-caryophyllene in in vivo models. Phytomedicine, 41, 82–95.
De Oliveira, M. R. C., Barros, L. M., Duarte, A. E., De Lima Silva, M. G., Da Silva, B. A. F., & Pereira Bezerra, A. O. B., et al. (2019). Gc-ms chemical characterization and in vitro evaluation of antioxidant and toxic effects using drosophila melanogaster model of the essential oil of lantana montevidensis (Spreng) briq. Medicine,. https://doi.org/10.3390/medicina55050194.
De Sousa, E. O., Lima, A. D. S., Lopes, S. G., Costa-Junior, L. M., & Da Costa, J. G. M. (2020). Chemical composition and acaricidal activity of Lantana camara L. and Lantana montevidensis Briq. essential oils on the tick Rhipicephalus microplus. Journal of Essential Oil Research, 32, 316–322. https://doi.org/10.1080/10412905.2020.1752320.
De Sousa, E. O., Rodrigues, F. F. G., Campos, A. R., Lima, S. G., & Da Costa, J. G. M. (2013). Chemical composition and synergistic interaction between aminoglycosides antibiotics and essential oil of Lantana montevidensis Briq. Natural Product Research, 27, 942–945. https://doi.org/10.1080/14786419.2012.678351.
Dennis, E. A., & Norris, P. C. (2015). Eicosanoid storm in infection and inflammation. Nature Reviews Immunology, 15, 511.
Denny, C., Melo, P. S., Franchin, M., Massarioli, A. P., Bergamaschi, K. B., de Alencar, S. M., & Rosalen, P. L. (2013). Guava pomace: a new source of anti-inflammatory and analgesic bioactives. BMC Complementary Medicine, 13, 235.
DiMartino, M. J., Campbell, G. K., Wolff, C. E., & Hanna, N. (1987). The pharmacology of arachidonic acid-induced rat paw edema. Agents and Actions, 21, 303–305.
dos Santos, D. A., de Fukui, M., & J., Nanayakkara, N.P.D., Khan, S.I., Sousa, J.P.B., Bastos, J.K., Andrade, S.F., da Silva Filho, A.A., & Quintão, N.L.M., . (2010). Anti-inflammatory and antinociceptive effects of Baccharis dracunculifolia DC (Asteraceae) in different experimental models. Journal of Ethnopharmacology, 127, 543–550.
Drexler, S. K., Kong, P. L., Wales, J., & Foxwell, B. M. (2008). Cell signalling in macrophages, the principal innate immune effector cells of rheumatoid arthritis. Arthritis Research and Therapy, 10, 216.
Dunham, N. W., & Miya, T. S. (1957). A note on a simple apparatus for detecting neurological deficit in rats and mice. Journal of the American Pharmaceutical Association, 46, 208–209.
Facanali, R., Campos, M. M. S., Pocius, O., Ming, L. C., Soares-Scott, M. D., & Marques, M. O. M. (2009). Reproductive biology of Ocimum selloi Benth. populations. Biologia reprodutiva de populações de Ocimum selloi Benth, 11, 141–146.
Gehlen, M. L., Moreira, H., Moreira, L., Sabag, F. P., & Repka, J. C. D. (2004). Avaliação espectrofotométrica do azul de Evans na reação inflamatória da córnea: estudo experimental em coelhos. Arquivos brasileiros de oftalmologia, 67, 219–225.
Gidwani, B. K., Bhargava, S., Rao, S. P., Majoomdar, A., Pawar, D. P., & Alaspure, R. N. (2009). Analgesic, anti-inflammatory and antihemorrhoidal activity of aqueous extract of Lantana camara Linn. Research Journal of Pharmacy and Technology, 2, 378–381.
Guideline, O. O. (2001). 425: acute oral toxicity—up-and-down procedure. OECD GuidelinesTesting Chemical, 2, 12–16.
Hu, Y., Zeng, Z., Wang, B., & Guo, S. (2017). Trans-caryophyllene inhibits amyloid β (Aβ) oligomer-induced neuroinflammation in BV-2 microglial cells. International Immunopharmacology, 51, 91–98. https://doi.org/10.1016/j.intimp.2017.07.009.
Huang, H., Al-Shabrawey, M., & Wang, M.-H. (2016). Cyclooxygenase-and cytochrome P450-derived eicosanoids in stroke. Prostaglandins & Other Lipid Mediators, 122, 45–53.
Kawahara, K., Hohjoh, H., Inazumi, T., Tsuchiya, S., & Sugimoto, Y. (2015). Prostaglandin E 2-induced inflammation: Relevance of prostaglandin E receptors. Biochim. Biophys. Acta (BBA)-Molecular Cell Biol. Lipids, 1851, 414–421.
Khan, M. J., Saraf, S., & Saraf, S. (2017). Anti-inflammatory and associated analgesic activities of HPLC standardized alcoholic extract of known ayurvedic plant Schleichera oleosa. Journal of Ethnopharmacology, 197, 257–265.
Kumar, T., & Jain, V. (2014). Antinociceptive and anti-inflammatory activities of bridelia retusa methanolic fruit extract in experimental animals. The Scientific World Journal.
Lapa, A. J., Souccar, C., Lima-Landman, M. T. R., de Castro, M. S., & A., Lima, T.C.M. (2003). Métodos de avaliação da atividade farmacológica de plantas medicinais (p. 2017). Plantas Med: Soc. Bras.
Li, K. K., Shen, S. S., Deng, X., Shiu, H. T., Siu, W. S., & Leung, P. C., et al. (2018). dihydrofisetin exerts its anti-inflammatory effects associated with suppressing Erk/p38 Mapk and Heme Oxygenase-1 activation in lipopolysaccharide-stimulated Raw 264.7 macrophages and carrageenan-induced mice paw edema. International Immunopharmacology, 54, 366–374.
Liu, K., Chen, Q., Liu, Y., Zhou, X., & Wang, X. (2012). Isolation and biological activities of decanal, linalool, valencene, and octanal from sweet orange oil. Journal of Food Science, 77, C1156–C1161.
Maling, H. M., Webster, M. E., Williams, M. A., Saul, W., & Anderson, W. (1974). Inflammation induced by histamine, serotonin, bradykinin and compound 48/80 in the rat: antagonists and mechanisms of action. Journal of Pharmacology and Experimental Therapeutics, 191, 300–310.
Malone, M. H., & Robichaud, R. C. (1962). A Hippocratic screen for pure or crude drug materials, in: Lloydia. pp. 320-+.
Marques, F. M., Figueira, M. M., Schmitt, E. F. P., Kondratyuk, T. P., Endringer, D. C., Scherer, R., & Fronza, M. (2019). In vitro anti-inflammatory activity of terpenes via suppression of superoxide and nitric oxide generation and the NF-κB signalling pathway. Inflammopharmacology, 27, 281–289. https://doi.org/10.1007/s10787-018-0483-z.
Martins, A. O. B. P. B., Rodrigues, L. B., Cesário, F. R. A. S., de Oliveira, M. R. C., Tintino, C. D. M., & e Castro, F.F., Alcântara, I.S., Fernandes, M.N.M., de Albuquerque, T.R., & da Silva, M.S.A. (2017). Anti-edematogenic and anti-inflammatory activity of the essential oil from Croton rhamnifolioides leaves and its major constituent 1, 8-cineole (eucalyptol). Biomedicine & Pharmacotherapy, 96, 384–395.
Matsuda, M., Huh, Y., & Ji, R. R. (2019). Roles of inflammation, neurogenic inflammation, and neuroinflammation in pain. J. Anesth., 33, 131–139. https://doi.org/10.1007/s00540-018-2579-4.
Mohamad, A. S., Akhtar, M. N., Zakaria, Z. A., Perimal, E. K., Khalid, S., & Mohd, P. A., et al. (2010). Antinociceptive activity of a synthetic chalcone, flavokawin B on chemical and thermal models of nociception in mice. European Journal of Pharmacology, 647, 103–109.
Montanari, R. M., Barbosa, L. C. A., Demuner, A. J., Silva, C. J., Carvalho, L. S., & Andrade, N. J. (2011). Chemical composition and antibacterial activity of essential oils from Verbenaceae species: Alternative sources of (E)-caryophyllene and germacrene-D. Quimica Nova, 34, 1550–1555.
Mostafa, M., Appidi, J. R., Yakubu, M. T., & Afolayan, A. J. (2010). Anti-inflammatory, antinociceptive and antipyretic properties of the aqueous extract of Clematis brachiata leaf in male rats. Pharmaceutical Biology, 48, 682–689.
Nam, J. H., Nam, D.-Y., & Lee, D.-U. (2016). Valencene from the rhizomes of cyperus rotundus inhibits skin photoaging-related ion channels and UV-induced melanogenesis in B16F10 melanoma cells. Journal of Natural Products, 79, 1091–1096.
Netea, M. G., Balkwill, F., Chonchol, M., Cominelli, F., Marc, Y., Giamarellos-bourboulis, E. J., et al. (2018). Europe PMC Funders Group Europe PMC Funders Author Manuscripts A guiding map for inflammation, 18, 826–831. https://doi.org/10.1038/ni.3790.A.
Olorukooba, A. B., & Odoma, S. (2019). Elucidation of the possible mechanism of analgesic action of methanol stem bark extract of Uapaca togoensis pax in mice. Journal of Ethnopharmacology, 245, 112156. https://doi.org/10.1016/j.jep.2019.112156.
Pahwa, R., Goyal, A., Bansal, P., & Jialal, I. (2020). Chronic Inflammation. Treasure Island (FL): StatPearls Publishing.
Patil, K. R., & Patil, C. R. (2017). Anti-inflammatory activity of bartogenic acid containing fraction of fruits of Barringtonia racemosa Roxb. in acute and chronic animal models of inflammation. Journal of Traditional and Complementary Medicine, 7, 86–93.
Prut, L., & Belzung, C. (2003). The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. European Journal of Pharmacology, 463, 3–33.
Ramirez, A. M., Cotera, L. B. F., & Gutierrez, R. M. Z. (2013). Anti-inflammatory activity of the hexane extract of byrsonimacrassi folia seeds in experimental animal models. Alternative Therapy, 19, 26–36.
Ray, S.D., Ray, S., Zia-Ul-Haq, M., De Feo, V., & Dewanjee, S. (2015). Pharmacological basis of the use of the root bark of Zizyphus nummularia Aubrev.(Rhamnaceae) as anti-inflammatory agent. BMC Complementary Alternative Medicine, 15, 416.
Rezaee-Asl, M., Sabour, M., Nikoui, V., Ostadhadi, S., & Bakhtiarian, A. (2014). The Study of Analgesic Effects of Leonurus cardiaca L. in Mice by Formalin, Tail Flick and Hot Plate Tests. Int. Sch. Res. Not. 2014, 687697. https://doi.org/10.1155/2014/687697
Ribeiro, N. A., Abreu, T. M., Chaves, H. V., Bezerra, M. M., Monteiro, H. S. A., Jorge, R. J. B., & Benevides, N. M. B. (2014). Sulfated polysaccharides isolated from the green seaweed Caulerpa racemosa plays antinociceptive and anti-inflammatory activities in a way dependent on HO-1 pathway activation. Inflammation Research, 63, 569–580.
Silva, G. N., Martins, F. R., Matheus, M. E., Leitão, S. G., & Fernandes, P. D. (2005). Investigation of anti-inflammatory and antinociceptive activities of Lantana trifolia. Journal of Ethnopharmacology, 100, 254–259.
Silva, T. S. C., Suffredini, I. B., Ricci, E. L., Fernandes, S. R. C., Gonçalves, V., Jr., & Romoff, P., et al. (2015). Antinociceptive and anti-inflammatory effects of Lantana camara L. extract in mice. The Revista Brasileira de Plantas Medicinais, 17, 224–229.
Silva, F. V., Oliveira, I. S., Figueiredo, K. A., Melo Júnior, F. B., Costa, D. A., & Chaves, M. H., et al. (2014). Anti-inflammatory and antinociceptive effects of Sterculia striata A. St.-Hil. & Naudin (Malvaceae) in rodents. Journal of Medicinal Food, 17, 694–700.
Swingle, K. F., & Shideman, F. E. (1972). Phases of the inflammatory response to subcutaneous implantation of a cotton pellet and their modification by certain antiinflammatory agents. Journal of Pharmacology and Experimental Therapeutics, 183, 226–234.
Tamaddonfard, E., Farshid, A. A., & Hosseini, L. (2012). Crocin alleviates the local paw edema induced by histamine in rats. Avicenna J. phytomedicine, 2, 97.
Tjølsen, A., Berge, O.-G., Hunskaar, S., Rosland, J. H., & Hole, K. (1992). The formalin test: an evaluation of the method. Pain, 51, 5–17.
Tsoyi, K., Jang, H. J., Lee, Y. S., Kim, Y. M., Kim, H. J., & Seo, H. G., et al. (2011). (+)-Nootkatone and (+)-valencene from rhizomes of Cyperus rotundus increase survival rates in septic mice due to heme oxygenase-1 induction. Journal of Ethnopharmacology, 137, 1311–1317.
Uzcátegui, B., Ávila, D., Suárez-Roca, H., Quintero, L., Ortega, J., & González, B. (2004). Anti-inflammatory, antinociceptive, and antipyretic effects of Lantana trifolia Linnaeus in experimental animals. Investigacion Clinica, 45, 317–322.
Vendramini-Costa, D. B., Spindola, H. M., de Mello, G. C., Antunes, E., Pilli, R. A., & de Carvalho, J. E. (2015). Anti-inflammatory and antinociceptive effects of racemic goniothalamin, a styryl lactone. Life Sciences, 139, 83–90.
Wang, D., Mann, J. R., & Dubois, R. N. (2005). The role of prostaglandins and other eicosanoids in the gastrointestinal tract. Gastroenterology, 128, 1445–1461.
Weverton, J., Bezerra, A., Rocha, M. I., & Sousa, J. D. (2019). Chemical composition and insecticidal action of essential oil from Lantana montevidensis ( Spreng .) Briq . (Chumbinho) against Nauphoeta cinerea Composición química y evaluación inseticida del aceite esencial de Lantana montevidensis ( Spreng .) Briq . 24, 1–11.
Wongrakpanich, S., Wongrakpanich, A., Melhado, K., & Rangaswami, J. (2018). A comprehensive review of non-steroidal anti-inflammatory drug use in the elderly. Aging Dis. 9, 143–150. https://doi.org/10.14336/AD.2017.0306
Xie, Y., Wang, K., Huang, Q., & Lei, C. (2014). Evaluation toxicity of monoterpenes to subterranean termite. Reticulitermes chinensis Snyder. Industrial Crops and Products, 53, 163–166. https://doi.org/10.1016/j.indcrop.2013.12.021.
Xu, Z., & Chang, L. (2017). Verbenaceae, in: Identification and Control of Common Weeds: Volume 3. Springer, pp. 163–179.
Yang, I.J., Lee, D.-U., & Shin, H.M. (2016). Inhibitory effect of valencene on the development of atopic dermatitis-Like skin lesions in NC/Nga mice. Evidence-Based Complement. Alternative Medicine.
Yui, K., Imataka, G., Nakamura, H., Ohara, N., & Naito, Y. (2015). Eicosanoids derived from arachidonic acid and their family prostaglandins and cyclooxygenase in psychiatric disorders. Current Neuropharmacology, 13, 776–785.
Zapata-Morales, J. R., Alonso-Castro, A. J., Domínguez, F., Carranza-Álvarez, C., Castellanos, L. M. O., Martínez-Medina, R. M., & Pérez-Urizar, J. (2016). Antinociceptive activity of an ethanol extract of justicia spicigera. Drug Development Research, 77, 180–186.
Zhang, Z., Yang, C., Dai, X., Ao, Y., & Li, Y. (2017). Inhibitory effect of trans-caryophyllene (TC) on leukocyte-endothelial attachment. Toxicology and Applied Pharmacology, 329, 326–333. https://doi.org/10.1016/j.taap.2017.06.016.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001, Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP)—Finance Code BPI and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Financiadora de Estudos e Projetos—Brasil (FINEP).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interests
The author(s) deny the existence of any potential conflict of interest with respect to the research, authorship, and/or publication of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
de Oliveira, M.R.C., de Lima Silva, M.G., Alcântara, I.S. et al. Biological activities of the essential oil from the leaves of Lantana montevidensis (Spreng) Briq. in mice. Environ Dev Sustain 23, 14958–14981 (2021). https://doi.org/10.1007/s10668-021-01281-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10668-021-01281-8