
Abstract
The increasing production of nano-TiO2 has attracted extensive concerns about the ecological consequence and health risk of these compounds in natural ecosystem. However, little is known about its toxicity on zooplankton, especially its possibility to access to the food chain via dietary exposure. To address this concern, the toxic and cumulative effects of nano-TiO2 on an aquatic food chain were explored through two trophic levels independently or jointly including producer and consumer. The results revealed that exposure to suspensions of nanomaterials had negative effects on both producers and consumers. Specifically, nanoparticles reduced the density of algal cells in a concentration-dependent way, and hatching life expectancy, average lifespan, net reproductive rate, and population intrinsic growth rate of rotifers decreased significantly with the concentration of nanomaterials increased (P < 0.05). Notably, nanoparticles accumulated in algal cells and were transferred to consumers through dietary exposure. Biomagnification of nano-TiO2 was observed in this simplified food chain, as many of the biomagnification factor (BMF) values in this study were >1. Exposure concentration, exposure time and their interactions play a strong part in the accumulation of nanoparticles in algae and rotifers. Overall, the present findings confirmed that nano-TiO2 was deleterious to plankton, posing a significant environmental threat to aquatic ecosystems.

Graphical abstract
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As one of the most widely used nanomaterial, Titanium dioxide nanoparticles (Nano-TiO2) is applied in cosmetics, anti-ultraviolet materials, plastics, paints and other industries due to its special structure and properties, such as unique optical features, extremely high chemical stability, thermal stability, super hydrophilicity and non-migration, etc (Li et al. 2008; Weir et al. 2012). However, engineered nanoparticles are unavoidably released into natural ecosystems during the process of production and use, resulting in ecotoxicity to aquatic food chains (Gottschalk and Nowack 2011).
Algae, the primary producers in the aquatic food chain, are commonly employed as a water quality and ecosystem health monitors (Ji et al. 2011). Theoretically, algae possess a semi-permeable cell walls that serve as the main site for interaction with the surrounding environment and the potential barrier against pollutants (Yue et al. 2017; Fleischer et al. 1999). While researches showed that the cell wall with pores of 5–20 nm cannot effectively prevent nanoparticles from entering cells, thereby causing cell damage (Navarro et al. 2008). Up to now, the detrimental impacts of nanoparticles were determined in algal growth (Fekete-Kertész et al. 2016; Xia et al. 2015; Aruoja et al. 2009), metabolism (Cardinale et al. 2012), ROS (George et al. 2010), enzyme activity (Roy et al. 2018; Liu et al. 2018), membranes and other cell structures (Navarro et al. 2008). Aquatic animals, particularly zooplankton, are frequently used as test organisms to evaluate the ecological safety and health risks of pollutants. Previously, nanoparticles have been suggested to cause rapid toxicity mortality in Daphnia magna and D. galeata (Cui et al. 2017), reduction of Rotifera biomass (Jovanović et al. 2016), significant inhibitory effect on the heartbeat rate (Fekete-Kertész et al. 2016) and moulting inhibition in D. magna (Nasser et al. 2016). Moreover, metal nanoparticles may affect the rotifers across the generations (Martins et al. 2020). In general, the toxicological effects often change with the test organisms (Fekete-Kertész et al. 2016; Rotini et al. 2018), test media (Nogueira et al. 2015) and the factitial properties of nanoparticles (Clément et al. 2013). Besides the direct nanotoxicity on aquatic organisms, nanomaterial is also known to have indirect effects on organisms through food chain, transferring to higher trophic levels and subsequent accumulation (Maharramov et al. 2019). Using electron microscopy, transfer and accumulation of nanoparticles have been confirmed in the digestive system and tissue of higher trophic levels, potentially including humans, most of them focusing on the Cladocera (Iswarya et al. 2018; Chae and An 2016; Zhu et al. 2009; Lee and An 2013; Kwon et al. 2015; Pinheiro et al. 2013; Fouqueray et al. 2012). For example, the Nano-TiO2 can transfer from D. magna to zebrafish Danio rerio by dietary exposure, and no biomagnifications of nTiO2 was observed in this simplified food chain. Compared to the dietary intake, D. rerio could accumulate nTiO2 by aqueous exposure with high bioaccumulation factors (BCFs) (Zhu et al. 2010b). However, a limited number of studies have been conducted to determine the transfer efficiency of nano-TiO2 on rotifer, an important member of the zooplankton community, except for a report of limited bioconcentration but no biomagnification of a model engineered nanomaterial (carboxylated and biotinylated quantum dots) through dietary uptake in a simplified freshwater food web (bacteria Escherichia coli—ciliate Tetrahymena pyriformis—rotifer B. calyciflorus) (Holbrook et al. 2008).
B. calyciflorus is one of the most common freshwater rotifers, occurs in almost all types of waterbodies with the features of rapid growth, high reproductive rate, short generation time, and high sensitivity to toxic substances, making them excellent bioindicators of water quality (Sha et al. 2018). The algae Scenedesmus obliquus is the underlying good food for the rotifer in lab or field (Lürling and Beekman 2006), and both of them are important members in the freshwater ecosystem and have been designated as indicators of heavy pollution. With this study, applying the life table and spectral detection technologies, the response strategies of algae and rotifers to nanoparticles and its transfer performance were explored under the stress of nano-TiO2, with the aim of obtaining further insights into: (1) the ecotoxicity of nano-TiO2 on producer Scenedesmus obliquus and primary consumer B. calyciflorus, (2) the bioaccumulation and biomagnification effects nano-TiO2 along the food chain.
Materials and methods
Nanomaterial pretreatment and characteristic analysis
Nano-TiO2 was purchased from McLean Biochemistry Technology Co., Ltd. (Shanghai, China). According to the manufacturer, the property of nano-TiO2 was hydrophilic anatase, the diameter was 5–10 nm and the purity was 99.8%. In this study, a stock solution was prepared by sonication in distilled water. Briefly, 100 mg nano-TiO2 was added in 100 mL distilled water and bath-sonicated at room temperature for 30 min, with 9 s ultrasound time and 9 s rest at 400 W (SM-1000D, Shunma, China). Partial suspension was used to analyse the physico-chemical properties of nanoparticles by transmission electron microscopy (TEM, JEM-1200EX, JEOL, Japan) and X-Ray Diffractometer (ultima IV, Rigaku Corporation, Japan) in Science Spectrum R & D Center (Qingdao, China), and the rest was preserved at 4 °C for following experiment (Liu et al. 2018; Chen et al. 2015).
Growth inhibition experiment on algae exposure to nanoparticles independently
To investigate the ecological response of producer to nanomaterials, the growth inhibition test was performed with green alga (S. obliquus) followed OECD guidelines 201 (OECD 2011). S. obliquus was acquired from the Freshwater Algae Culture Collection at the Institute of Hydrobiology (FACHB-collection), then semi-continuously cultured with the OECD TG 201 medium in an incubator shaker with illumination (Temperature: 22 ± 1 °C, Illumination: 54 μmol·photons m−2 s−1, Light: Dark = 12 h: 12 h, Speed: 140 rpm). Based on preliminary dose-range finding tests, seven final nominal concentrations (0, 5, 10, 20, 40, 80 and 160 mg/L) of nano-TiO2 suspensions were selected to determine the effective concentration (ECx) which treated by nanoparticles at 72 h. The suspensions of nano-TiO2 were prepared by serially diluting stock solution to reach test solution concentration using OECD TG 201 medium for the experiments. In the experiment, S. obliquus in exponential growth phase were inoculated into the aforementioned seven test solutions to a final density of 5 × 104 cells/mL. The conditions used to culture algae were the same as in the semi-continuous culture. To confirm reproducibility, each treatment was conducted in three replicates. The morphological characteristics of algae S. obliquus exposed for 72 h were measured under a microscope using hemocytometer and the software Image J. Since the algae cell is spindle-shaped, its volume was calculated by the formula V = πab2/6, where: a is the long axis and b is the short axis of the cells (Sun and Liu, 2003).
Algal cell number in different experiment groups were monitored every 24 h using a hemocytometer (Leica, DM2000). According to daily change in cell number of algal culture, the specific growth rate (μ) was calculated with Eq. (1) (OECD 2011).
where μi-j is the specific growth rate from time i to j; Xj is the density at time j; and Xi is the density at the time i.
The growth inhibition rate was then calculated with Eqs. (2) and (3) (Wang et al. 2016).
where Yt is the area below growth curve; N0 is the algal density at t0 (5.0 × 104 cells/mL); N1 is the algal density at t1 (cells/mL); Nn is the algal density at tn (cells/mL); t1 is the first sampling time after the beginning of the experiment (h); tn is the n sampling time after the beginning of the experiment (h).
where I is the growth inhibition rate (%) of the treatments; Yc is the area below growth curve in control; Yt is the area below growth curve in different treatment.
In toxicology, to quickly assess the possible impact of pollutants on the tested organisms and provide reference for chronic toxicity studies, the concentration that can cause the 50% biological growth inhibition is called as the half-maximal effective concentration (EC50). According to OECD (2011) guidelines for testing of chemicals: For growth inhibition test in freshwater alga and cyanobacteria, a quantitative concentration - response relationship was obtained by regression analysis between the logarithm (10 based) of concentration and the probit of growth inhibition rate. Accordingly, ECx at 72 h was calculated with the regression equation at x% of growth inhibition rate (I).
Rotifer life history experiment
In this experiment, the rotifer B. calyciflorus was sampled from Lake Jinghu (118.3750E, 31.3298 N) in Wuhu City, Anhui Province, China. After collection, rotifers individuals were clonally cultured at 23 ± 1 °C, with the green algae S. obliquus as food at the density of 2.0 × 106 cells/mL. The medium used to support rotifers was freshly prepared EPA (pH 7.4–7.8). It can be obtained by dissolving 96 mg NaHCO3, 60 mg CaSO4, 60 mg MgSO4 and 4 mg KCl in 1 L distilled water (Peltier and Weber 1985). According to the acute toxicity test (Li et al. 2020), the concentrations of nano-TiO2 suspension were set at 0, 0.5, 1.0, 1.5, 2.0, 2.5 and 3.0 mg/L. For each treatment, 10 neonates, born within 4 h, were introduced into 8 ml glass beakers containing 5 ml culture medium with the algal density of 2.0 × 106 cells/mL. Each treatment consisted of three replicates. After that, the containers were put in a bioclimatic chamber at 23 ± 1 °C. The rotifers were checked every 8 h, meanwhile we suspended the culture for preventing the deposition of nano-materials in the experiment. The alive original individuals and neonates were recorded and then the neonates were discarded. Meanwhile, the original individuals which are still alive were transferred into freshly prepared test solutions every 24 h. The experiments were terminated when every individual in each cohort died.
Based on the collected data, the average lifespan (LS) was calculated. The life expectancy at hatching (e0), the net reproductive rate (R0) and generation time (T) were calculated with Eqs. (4) (5) and (6), respectively (Kerbs 1985). Intrinsic rate of population increases (rm), was first approximated using: r-rough = lnR0/T. For the final calculation, we solved with the Eq. (7).
where nx is the number of alive rotifers at time x; lx is the survival rates of rotifers at time x; mx is the fecundity of rotifers at time x.
Dietary exposure experiment along food chain
Accumulation and transfer of nano-TiO2 via the food chain (S. obliquus- B. calyciflorus) were investigated by dietary exposure method. To detect the accumulation of nano-TiO2 in the algae and prepare the algal food for rotifers in the subsequent food chain experiment, the algae was cultured under the different effect concentration (EC10, EC20, EC30, EC40 and EC50) obtained from the above algal growth inhibition experiments for ensuring the stable growth and reproduction of algae. The initial density was set as 1.0 × 106 cells/mL, and the other conditions were same as the growth inhibition experiment. Three replicates and a control group were set for each treatment. At the 3rd day, 5th day and 7th day, the algal cells were collected by centrifugation at 4000 rpm for 10 min. To separate nanoparticles from the algal cells surface, each sample was purified by sucrose density gradient centrifugation (Oukarroum et al. 2017; Xiong et al. 2011). Briefly, the sucrose solutions with mass fraction of 40, 60, 80, 100, 120 and 140% were prepared in sterile water, then 10 mL of each sucrose solution was gently placed in a 100 mL centrifuge tube by layering progressively less dense sucrose solutions upon one another. Subsequently, collected algae were slowly added on the top of the sucrose gradient and centrifuged at 2000 rpm for 10 min. Afterwards, the content of element Ti in algal were detected by ICP-OES (Thermo Fisher, iCAP7400) in Sremol Environmental Technology Co. LTD (Nanjing, China).
To ensure the quantity and quality of algae, we selected the algae exposed to nano-TiO2 for 5-days as the food for the rotifer. The exposure concentration of algae and experimental methods were followed by aforementioned methods to prepare algal food for subsequent consumers. In dietary exposure experiments, 50 rotifers were transferred into 8 mL beakers containing 5 mL EPA medium with the algal density of 2.0 × 106 cells/mL. Moreover, rotifers medium and algal food was renewed every 24 h. For preventing the deposition of algae with nano-materials in the experiment, we suspended the culture every 8 h. After 3 days, 5 days and 7 days of dietary exposure, 50 rotifers were sampled and rinsed three times with deionized water to remove any S. obliquus attached to their surface. We then determined the element Ti concentration in B. calyciflorus by ICP-OES (Thermo Fisher, iCAP7400) in Sremol Environmental Technology Co. LTD (Nanjing, China).
Based on the detected data obtained for both algae and rotifers, the biomagnification factor (BMF) was calculated to evaluate the trophic transfer efficiency of nano-TiO2 along the food chain (Arnot and Gobas 2006). The BMF of nano-TiO2 has been computed as the ratio of Ti concentration in the rotifers than that in the algal diet as represented in Eq. (8) (Iswarya et al. 2018).
Statistical analysis
The data were analysed by SPSS 25.0 and expressed as the mean ± standard deviation (SD). One-sample Kolmogorov–Smirnov procedure and Levene’s test were used to test all data for normality and homogeneity of variances, respectively. Kaplan–Meier analyses were conducted to test for the differences in the survivorship of the rotifer cohorts among seven exposure concentrations. One-way analysis of variance (ANOVA) was conducted to identify the significant influence of exposure concentrations and time on life table parameters, as well as accumulation of nanoparticles in algae and rotifers, respectively. Two-way ANOVA of variance (ANOVAs) was executed to resolve the influence of exposure concentration, exposure time and their interaction on accumulation of nanoparticles in producers and consumers. To determine which groups were significantly different among the different treatments, multiple comparisons were carried out using Student-Neuman-Keuls (SNK). The significant level of statistics (P) was set as <0.05.
Results
Characteristics of nano-TiO2
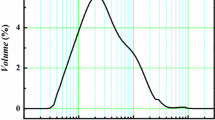
The physico-chemical properties of nano-TiO2 suspensions were detected (Fig. 1). Analysis by TEM showed that nano-TiO2 presents an oval shape and agglomerated into irregular structures in the distilled water (Fig. 1-I, II, III), with diameters ranging from 13 to 36 nm and average of 21.58 nm (Fig. 1-IV), which were higher than the product description by manufacturer. The analysis showed that the zeta potential of these nanoparticles was 28.2 mV (Fig. 1-V). The value of the Zeta potential is related to the stability of the colloidal dispersion. The higher the Zeta value, the more stable the system is. Thus, the nanoparticle solution used in this study was homogeneous and unstable.

Characteristic of nano-TiO2 in the distilled water (1.0 g/L): (I), (II) and (III) are TEM image; (IV) is size distribution; (V) is the zeta potential
Growth inhabitation of nano-TiO2 on algae
The growth of S. obliquus was inhibited at 72 h gradually with the increasing nano-TiO2 dose. In addition, the results suggested a positive linear correlation (R2 = 0.9526) between logarithm (base 10) of concentrations and the probit of inhibition rate (Fig. 2). With the increasing of nano-particle concentration, the maximum population density and specific growth rate decreased significantly, additionally the algae cells got smaller gradually (Table 1). According to the algal growth inhibition experiment, different effect concentrations at 72 h were EC10 = 6.46 mg/L, EC20 = 12.58 mg/L, EC30 = 19.90 mg/L, EC40 = 30.20 mg/L and EC50 = 43.65 mg/L, respectively, and the EC50 = 57.54 mg/L at 48 h.

Growth inhibition of algae under different exposure concentration. x is the logarithm (base 10) of concentration; y is the probit of inhibition rate
Rotifer life history experiment
We compared the age-specific survivoship and fecundity of rotifers exposed in different concentrations of nanoparticles using standard life table methods. The results showed that the age-specific survivoship (lx) of treatment groups tended to decrease earlier compared with control (Fig. S1). However, there was no significant difference among exposure concentrations (Kaplan–Merier analysis, Fig. 3, P > 0.05). As for the age-specific fecundity (mx), the maximum values of rotifers neonates in exposure concentration 0, 0.5, 1.0, 1.5, 2.0, 2.5 and 3.0 mg/L were 51.67 ± 4.62, 42.33 ± 2.08, 42.00 ± 1.00, 42.67 ± 8.62, 35.67 ± 1.53, 33.00 ± 2.00 and 32.33 ± 1.15 ind., respectively. Besides, some specific death and reproduction time in life history, such as time for death, time for all death, time for reproduction, peak time of reproduction and reproduction end time were further analysed. However, no sinificant difference was found except for thr time for death (P > 0.05, Fig. 4).

Survival rates of rotifers exposure in different nanoparticles concentrations

Specific death and reproduction time for rotifers (mean ± SD). For each parameter, values in each column with * indicate significant different compared with control according to SNK at P < 0.05
The one-way analysis of variance indicated the life table parameters were significantly affected among exposure concentration of nanoparticle (Table 2). Under the stress of nano-materials, the net reproductive rate (R0) and intrinsic rate of population increase (rm) of rotifers decreased significantly with the raising of exposure concentration (P < 0.05). The results revealed the reduced life expectancy at hatching (e0) and average lifespan (LS), although non-significant. However, no statistical evidence was found to confirmed significant difference in generation time (P > 0.05).
Dietary exposure experiment
The accumulation of nanoparticles was observed in algal cells after exposure to nano-TiO2 suspension for different times (Fig. 5I). The results indicated that the content of Ti element in algal cells increased significantly with the prolongation of exposure time and increasing of exposure concentration (P < 0.05).

Accumulation of nano-TiO2 in S. obliquus and rotifers at different exposure concentration and time (Mean ± SD, N = 3). I is the accumulation of nanoparticles in algae; II is the accumulation of nanoparticles in rotifers. For each nanoparticle concentration, values in each column with different capital letters indicate significant different among treatments according to SNK at P < 0.05; For each exposure time, values in each column with different lowercase letters indicate significant different among treatments according to SNK at P < 0.05
The impact of a dietary exposure experiment was investigated using a basic algae-rotifer model to see if they may transfer across trophic levels in the aquatic food chain. As an unselective filter-feeder, B. calyciflorus feeding on algae which were contaminated by nano-TiO2 resulted in accumulation of nanoparticles in rotifers (Fig. 5II). The content level of nano-TiO2 in algal food had a significant influence on its accumulation in rotifers by dietary exposure (P < 0.05). From the analyses, the accumulative content of nanoparticles in the rotifers increased with the prolong of feeding time. The concentration of nano-TiO2 elevated significantly at the 7th day for the EC10 group. However, this significance started at the 5th day when the concentration was equal or higher than EC20. As the rotifer was fed on the algae with nanoparticles for short time (3 days), the accumulation of nano-TiO2 increased firstly and decreased then at the highest exposure (EC50). After 5-days and 7-days feeding, the nano-TiO2 content kept at high level constantly.
We also explored the biomagnification factors (BMF) of nano-TiO2 in the food chain, and different BMFs occurred alone the food chain due to the different levels of nano-TiO2 in food and feeding time. The result showed that when rotifers ingested contaminated algae for 3 days, its BMF values were in the range of 0.80–2.36, 1.13–6.57 BMF for 5 days, and 1.77–7.40 BMF for 7 days (Table 3). Compared with 3 days of feeding, the BMF values of 5 days and 7 days were all >1, indicating that there is a phenomenon of biomagnification in the nano-TiO2 accumulation by rotifers. The two-way ANOVA analyses suggested that the dietary exposure time and concentration significantly increased the values of BMF (P < 0.05). The BMF at the concentration of EC40 for 3 days was greater than the other ones significantly, and the BMF increased firstly and then decreased with the raising of effective concentrations when feeding rotifer for 5 days, however after stressed for 7 days the BMF value was the significant highest at the EC10 and EC30, then decreased at EC20, EC40 and EC50 gradually. At the EC10 and EC50, the value of BMF reached a maximum at the dietary exposure time of 7 days, while they were significant maximal at 5 days for the EC20, EC30 and EC40. Two-way ANOVA showed that dietary exposure time, exposure concentration and their interaction significantly affected the accumulation of nano-TiO2 in algae and rotifers as well as the value of BMF (P < 0.05, Table 4).
Discussion
Algae are an important primary producer in aquatic ecosystems, and it has been the main test organism in the studies of ecotoxicity effect of nano-TiO2 on aquatic producers. The most intuitionistic toxic response of algal cells to the nano TiO2 is growth inhibition (Wong et al. 2020). This is reflected by EC50, which varied greatly with the range of 2.53–179.05 mg/L depending on the types and diameters of nanoparticles, algae species, exposure time (Table 5). For this study, the EC50 was almost two times (43.65 vs. 21.2 mg/L) comparing to the results by Sadiq et al. (2011) based on the same genus Scenedesmus and similar features of nanoparticles. In addition, the results suggested the significant decreases of the maximum population density, specific growth rate and the cells size with the increasing of nano-TiO2 concentration. Different levels of Ti in algal indicated that nanoparticles can penetrated the cell wall and accumulated in algal cells. Exposure concentration and time significantly affected the accumulation of nanoparticles in algal cells. This result is coincident with the previous findings about nanotoxicity on algae. Many explanations on the algal growth inhibition have been addressed in many aspects, including alteration of enzymatic activity, cell membrane impairment and production of ROS (Li et al. 2015; Fu et al. 2015). Further, we assumed that many nanoparticles were absorbed by algae during the experiment, resulting in the increased cellular weight and algae sinking, which will reduce the amount of light accessible for photosynthesis and thus lessen the algal biomass. These underlying effects will impede utilising of CO2, which in turn prevents the achieving of carbon neutrality.
Zooplankton, a filter feeder in water, can take in the nanoparticles released into the environment unavoidably. The present study discovered the impact of nanotoxicity on rotifer life history strategy, including life expectancy at hatching, intrinsic rate of population growth, net reproduction rate, and average longevity. Jacobasch et al. (2014) also found that nano-TiO2 chronic exposure resulted in population breakdown of Daphnia magna at concentration of 1.78 mg/L. Li et al. (2020) suggested that treatments with 1 mg/L nano-TiO2 significantly decreased the population density of rotifers. Dong et al. (2020) also discovered that nano-TiO2 treatment affected the life-table characteristics of B. calyciflorus. The results herein described are directly in line with the previous findings, and the species of B. calyciflorus play an excellent test model in toxicity assessment of nano-TiO2.
Because some pollutants have the ability to accumulate in primary producers and eventually reach the end of the food chain, understanding bioaccumulation and dietary transfer of contaminants in aquatic environments is critical. By comparing the results of Ti content from producers and consumers, we determined that nanoparticles were transferred via dietary exposure in the aquatic food chain. From these results, it is clear that dietary exposure concentration, exposure time and their interaction significantly affected the accumulation of nano-TiO2 in rotifers (P < 0.01). However, the cumulations of nano-TiO2 in rotifers gradually increased and might reach saturation (Because of the limitations of lifespan in rotifers we can’t investigate the exposure time for longer) with the delay of dietary exposure time. Chen et al. (2015) found that nanoparticles were accumulated in D. magna rapidly from day 1 to 7, and reached saturation from day 8 to 21. Suggest: Zhu et al. (2010b) also found the content of nano-TiO2 in D. rerio accumulated rapidly at the dietary exposure time of the first day but tended to be saturated by the 5th day. This is consistent with the findings of previous studies.
Usually, a specific contaminant through environment exposure or dietary uptake were quantified by bioconcentration and biomagnification factors (Holbrook et al. 2008). Previous studies investigated bioaccumulation and biotransformation of nano-TiO2 in freshwater food chain, the results showed that nano-TiO2 was accumulated in Daphnia and elimination was difficult (Zhu et al. 2010a). At the same time, they found that nano-TiO2 can transfer from D. magna to Danio rerio by dietary exposure. However, no biomagnifications were observed in the simplified food chain because the values of the biomagnification factors (BMF) in this study (0.024 and 0.009) were all less than one (<1) (Zhu et al. 2010b). Similarly, biomagnification of QDs nanoparticles was not found in a food web which including bacteria (Escherichia coli), ciliate (Tetrahymena pyriformis) and rotifer species (B. calyciflorus) (Holbrook et al. 2008). On contrary, many BMF values of nanoparticles in the present study were >1, indicating that biomagnification of nano-TiO2 from S. obliquus to B. calyciflorus occurred. The result is line with that of Chen et al. (2015) who also found biomagnification (nano-TiO2 BMF = 7.83, nano-TiO2-SDBS = 2.66) of nano-TiO2 in a simplified invertebrate food chain including S. obliquus and D. magna. Moreover, Chen et al. (2019) also demonstrated that BMFs of daphnia fed with 1 and 10 mg/L nTiO2-exposed algae were consistently >1.0 (5.7–122). Previous study indicated that nanoparticles trophic transfer occurs through the food chain and it is more efficient than through direct contact an organism with nanoparticles (Unrine et al. 2012). This may be the reason for the variable findings in previous studies. In addition, a necessary condition for biomagnification in food chains or webs is that the pollutant must be soluble in fats or lipophilic (Mader 1996). However, lipophobic and insoluble nanoparticles have a high BMF through food chain transfer, demonstrating important impacts of nanoparticles size, exposure methods and surfactant on bioaccumulation and biomagnification. In case of biomagnification of these nanoparticles in food chains, organisms at higher trophic levels or human consumers in the food chains are at greater ecological and health risks as a result of constantly accumulation of trace elements.
Conclusions
Our studies showed that algal growth, reproduction and cells size were inhibited by nano-TiO2 treatment. Although algae possess cell walls serving as the barrier for pollutant entry, nanoparticles can still pass through the cell walls and accumulate in the cells. The concentration and duration of exposure had an effect on the content of nanoparticles in the algal cells. Moreover, the life history strategies of rotifers were changed negatively under the stress of nano-TiO2, including the longevity, the net reproductive rate, the life expectancy at hatching and the intrinsic rate of population growth. In addition, nanoparticles in algae were transferred to the rotifers via dietary exposure. The content of nanoparticles in consumers was closely related to that in primary producers due to dietary exposure. Similarly, dietary exposure concentration, time and their interaction have significant influence on bioaccumulation and biomagnification of nanoparticles. To determine the relative relevance of aqueous and dietary exposure to different nanoparticles more research is needed.
Data availability
Supplementary material submitted with paper. Any further data required can be requested from corresponding author.
References
Arnot JA, Gobas FA (2006) A review of bioconcentration factor (BCF) and bioaccumulation factor (BAF) assessments for organic chemicals in aquatic organisms. Environ Rev 14:257–297. https://doi.org/10.1139/A06-005
Aruoja V, Dubourguier HC, Kasemets K, Kahru A (2009) Toxicity of nanoparticles of CuO, ZnO and TiO2 to microalgae Pseudokirchneriella subcapitata. Sci Total Environ 407:1461–1468. https://doi.org/10.1016/j.scitotenv.2008.10.053
Cardinale BJ, Bier R, Kwan C (2012) Effects of TiO2 nanoparticles on the growth and metabolism of three species of freshwater algae. J Nanoparticle Res 14:913–921. https://doi.org/10.1007/s11051-012-0913-6
Chae Y, An YJ (2016) Toxicity and transfer of polyvinylpyrrolidone-coated silver nanowires in an aquatic food chain consisting of algae, water fleas, and zebrafish. Aquat Toxicol 173:94–104. https://doi.org/10.1016/j.aquatox.2016.01.011
Chen JY, Li HR, Han XQ, Wei XZ (2015) Transmission and accumulation of nano-TiO2 in a 2-step food chain (Scenedesmus obliquus to Daphnia magna). Bull Environ Contam Toxicol 95:145–149. https://doi.org/10.1007/s00128-015-1580-y
Chen XJ, Zhu Y, Yang K, Zhu LZ, Lin DH (2019) Nanoparticle TiO2 size and rutile content impact bioconcentration and biomagnification from algae to daphnia. Environ Pollut 247:421–430. https://doi.org/10.1016/j.envpol.2019.01.022
Clément L, Hurel C, Marmier N (2013) Toxicity of TiO2 nanoparticles to cladocerans, algae, rotifers and plants—Effects of size and crystalline structure. Chemosphere 90:1083–1090. https://doi.org/10.1016/j.chemosphere.2012.09.013
Cui RX, Chae Y, An YJ (2017) Dimension-dependent toxicity of silver nanomaterials on the cladocerans Daphnia magna and Daphnia galeata. Chemosphere 185:205–212. https://doi.org/10.1016/j.chemosphere.2017.07.011
Dong LL, Wang HX, Ding T, Li W, Zhang G (2020) Effects of TiO2 nanoparticles on the life-table parameters, antioxidant indices, and swimming speed of the freshwater rotifer Brachionus calyciflorus. J Exp Zool A Ecol Genet Physiol 333:230–239. https://doi.org/10.1002/jez.2343
Fekete-Kertész I, Maros G, Gruiz K, Molnár M (2016) The effect of TiO2 nanoparticles on the aquatic ecosystem: a comparative ecotoxicity study with test organisms of different trophic levels. Period Polytech-chem 60:231–243. https://doi.org/10.3311/ppch.8869
Fleischer A, O’Neill MA, Ehwald R (1999) The pore size of non‑graminaceous plant cell walls is rapidly decreased by borate ester cross‑linking of the pectic polysaccharide rhamnogalacturonan II1. Plant Physiol 121:829–838. https://doi.org/10.1104/pp.121.3.829
Fouqueray M, Dufils B, Vollat B, Chaurand P, Botta C, Abacci K, Labille J, Rose J, Garric J (2012) Effects of aged TiO2 nanomaterial from sunscreen on Daphnia magna exposed by dietary route. Environ Pollut 163:55–61. https://doi.org/10.1016/j.envpol.2011.11.035
Fu L, Hamzeh M, Dodard S, Zhao YH, Sunahara GI (2015) Effects of TiO2 nanoparticles on ROS production and growth inhibition using freshwater green algae pre-exposed to UV irradiation. Environ Toxicol Pharmacol 39:1074–1080. https://doi.org/10.1016/j.etap.2015.03.015
George S, Pokhrel S, Xia T, Gilbert B, Ji Z, Schowalter M, Rosenauer A, Damoiseaux R, Bradley KA, M„dler L, Nel AE (2010) Use of a rapid cytotoxicity screening approach to engineer a safer zinc oxide nanoparticle through iron doping. ACS Nano 4:15–29. https://doi.org/10.1021/nn901503q
Gottschalk F, Nowack B (2011) The release of engineered nanomaterials to the environment. J Environ Monitor 13:1145–1155. https://doi.org/10.1039/c0em00547a
Hartmann NB, Kammer FV, Hofmann T, Ottofuelling S, Baun A (2010) Algal testing of titanium dioxide nanoparticles-testing considerations, inhibitory effects and modification of cadmium bioavailability. Toxicology 269:190–197. https://doi.org/10.1016/j.tox.2009.08.008
Holbrook RD, Murphy KE, Morrow JB, Cole KD (2008) Trophic transfer of nanoparticles in a simplified invertebrate food web. Nat Nanotech 3:352–355. https://doi.org/10.1038/nnano.2008.110
Hund-Rinke K, Simon M (2006) Ecotoxic effect of photocatalytic active nanoparticles (TiO2) on algae and daphnids. Environ Sci Pollut Res 13:225–232. https://doi.org/10.1065/espr2006.06.311
Iswarya V, Bhuvaneshwari M, Chandrasekaran N, Mukherjee A (2018) Trophic transfer potential of two different crystalline phases of TiO2 NPs from Chlorella sp. to Ceriodaphnia dubia. Aquat Toxicol 197:89–97. https://doi.org/10.1016/j.aquatox.2018.02.003
Jacobasch C, Voelker C, Giebner S, Völker J, Alsenz H, Potouridis T, Heidenreich H, Kayser G, Oehlmann J, Oetken M (2014) Long-term effects of nanoscaled titanium dioxide on the cladoceran Daphnia magna over six generations. Environ Pollut 186:180–186. https://doi.org/10.1016/j.envpol.2013.12.008
Ji J, Long ZF, Lin DH (2011) Toxicity of oxide nanoparticles to the green algae Chlorella sp. Chem Eng J 170:525–530. https://doi.org/10.1016/j.cej.2010.11.026
Jovanović B, Bezirci G, Çağan AS, Coppens J, Levi EE, Oluz Z, Tuncel E, Duran H, Beklioğlu M (2016) Food web effects of titanium dioxide nanoparticles in an outdoor freshwater mesocosm experiment. Nanotoxicology 10:902–912. https://doi.org/10.3109/17435390.2016.1140242.
Kerbs CJ (1985) Ecology: the experimental analysis of distribution and abundance. Q Rev Biol 48:133–148. https://doi.org/10.1086/407551
Kwon D, Nho HW, Yoon TH (2015) Transmission electron microscopy and scanning transmission x-ray microscopy studies on the bioaccumulation and tissue level absorption of TiO2 nanoparticles in Daphnia magna. J Nanosci Nanotechnol 15:4229–4238. https://doi.org/10.1166/jnn.2015.9695
Lee SW, Obregón S, Rodríguez-González V (2013) The role of silver nanoparticles functionalized on TiO2 for photocatalytic disinfection of harmful algae. RSC Adv 5:44470–44475. https://doi.org/10.1039/C5RA08313C
Lee WM, An YJ (2013) Effects of zinc oxide and titanium dioxide nanoparticles on green algae under visible, UVA, and UVB irradiations: No evidence of enhanced algal toxicity under UV pre-irradiation. Chemosphere 91:536–544. https://doi.org/10.1016/j.chemosphere.2012.12.033
Li F, Liang Z, Zheng X, Zhao W, Wu M, Wang ZY (2015) Toxicity of nano-TiO2 on algae and the site of reactive oxygen species production. Aquat Toxicol 158:1–13. https://doi.org/10.1016/j.aquatox.2014.10.014
Li FM, Zhao W, Li YY, Tian Z, Wang ZY (2012) Toxic Effects of Nano-TiO2 on Gymnodinium breve. Environ Sci 33:233–238
Li M, Huang R, Xi YL, Xiang XL (2020) Effects of nano-TiO2 exposure on population dynamics of Brachionus calyciflorus. Asian J Ecotoxicol 15:234–242. https://doi.org/10.7524/AJE.1673-5897.20200103002
Li QL, Mahendra S, Lyon DY, Brunet L, Liga MV, Li D, Alvarez PJJ (2008) Antimicrobial nanomaterials for water disinfection and microbial control: potential applications and implications. Water Res 42:4591–4602. https://doi.org/10.1016/j.watres.2008.08.015
Liu YH, Wang S, Wang Z, Ye N, Fang H, Wang DG (2018) TiO2, SiO2 and ZrO2 nanoparticles synergistically provoke cellular oxidative damage in freshwater microalgae. Nanomaterials 8:95–107. https://doi.org/10.3390/nano8020095
Lürling M, Beekman W (2006) Influence of food-type on the population growth rate of the rotifier Brachionus calyciflorus in short-chronic assays. Acta Zoologica Sinica 52(1):70–78
Mader SS (1996) Biology, Fifth Edition. Wm. C. Brown Publishers, Dubuque, IA, USA
Martins N, Pradhan A, Pascoal C, Cássio F (2020) Effects of metal nanoparticles on freshwater rotifers may persist across generations. Aquat Toxicol. 229:105652. https://doi.org/10.1016/j.aquatox.2020.105652
Maharramov AM, Hasanova UA, Suleymanova IA, Osmanova GE, Hajiyeva NE (2019) The engineered nanoparticles in food chain: potential toxicity and effects. SN Appl Sci 1:1362. https://doi.org/10.1007/s42452-019-1412-5
Nasser F, Davis A, Valsami-Jones E, Lynch I (2016) Shape and charge of gold nanomaterials influence survivorship, oxidative stress and moulting of Daphnia magna. Nanomaterials 6:222–235. https://doi.org/10.3390/nano6120222
Navarro E, Baun A, Behra R, Hartmann NB, Filser J, Miao AJ, Quigg A, Santschi PH, Sigg L (2008) Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 17:372–386. https://doi.org/10.1007/s10646-008-0214-0
Nogueira A, Lopes I, Rocha-Santos TAP, Rasteiro MG, Abrantes N, Gocalves F, Soares AMVM, Duarte AC, Pereira R (2015) Assessing the ecotoxicity of metal nano-oxides with potential for wastewater treatment. Environ Sci Pollut Res 22:13212–13224. https://doi.org/10.1007/s11356-015-4581-9
OECD (2011) Test No. 201: Freshwater alga and cyanobacteria, growth inhibition test, OECD Guidelines for the Testing of Chemicals, Section 2. OECD Publishing, Paris, https://doi.org/10.1787/9789264069923-en
Oukarroum A, Zaidi W, Samadani M, Dewez D (2017) Toxicity of nickel oxide nanoparticles on a freshwater green algal strain of Chlorella vulgaris. Biomed Res Int 2017:1–8. https://doi.org/10.1155/2017/9528180
Peltier WH, Weber CI (1985) Methods for measuring the acute toxicity of effluents to freshwater and marine organisms. United States Environmental Protect Agency, Cincinnati, Ohio
Pinheiro T, Moita L, Silva L, Mendonca E, Picado A (2013) Nuclear microscopy as a tool in TiO2 nanoparticles bioaccumulation studies in aquatic species. Nucl Instrum Methods Phys Res B 306:117–120. https://doi.org/10.1016/j.nimb.2012.12.049
Rotini A, Gallo A, Parlapiano I, Berducci MT, Boni R, Tosti E, Prato E, Maggi C, Cicero AM, Migliore L, Manfra L (2018) Insights into the CuO nanoparticle ecotoxicity with suitable marine model species. Ecotox Environ Safe 147:852–860. https://doi.org/10.1016/j.ecoenv.2017.09.053
Roy B, Chandrasekaran H, Krishnan SP, Chandrasekaran N, Mukherjee A (2018) UVA pre-irradiation to P25 titanium dioxide nanoparticles enhanced its toxicity towards freshwater algae Scenedesmus obliquus. Environ Sci Pollut Res 25:1–14. https://doi.org/10.1007/s11356-018-1860-2
Sadiq MI, Dalai S, Chandrasekaran N, Mukherjee A (2011) Ecotoxicity study of titania (TiO2) NPs on two microalgae species: Scenedesmus sp. and Chlorella sp. Ecotoxicol Environ Saf 74:1180–1187. https://doi.org/10.1016/j.ecoenv.2011.03.006
Sha JJ, Dai YY, Pan YL, Li J, Xu ZD, Zhang JM (2018) Research progress in using rotifers in ecotoxicological studies. Asian J Ecotoxic 13:56–70. https://doi.org/10.7524/AJE.1673-5897.20171022001
Sun J, Liu D (2003) Geometric models for calculating cell biovolume and surface area for phytoplankton. J Plankton Res 25:1331–1346. https://doi.org/10.1093/plankt/fbg096
Unrine JM, Aaron Shoults-Wilson W, Zhurbich O, Bertsch PM, Tsyusko OV (2012) Trophic transfer of Au nanoparticles from soil along a simulated terrestrial food chain. Environ Sci Technol 46(17):9753–9760. https://doi.org/10.1021/es3025325
Wang YX, Zhu XS, Lao YM, Lv XH, Tao Y, Huang BM, Wang JX, Zhou J, Cai ZH (2016) TiO2 nanoparticles in the marine environment: physical effects responsible for the toxicity on algae Phaeodactylum tricornutum. Sci Total Environ 565:818–826. https://doi.org/10.1016/j.scitotenv.2016.03.164
Weir A, Westerhoff P, Fabricius L, Goetz N (2012) Titanium dioxide nanoparticles in food and personal care products. Environ Sci Technol 46:2242–2250. https://doi.org/10.1021/es204168d
Wong SWY, Zhou GJ, Kwok KWH, Djurišić AB, Han J, Lee J, Leung KMY (2020) In vivo toxicities of nine engineered nano metal oxides to the marine diatom Skeletonema costatum and rotifer Brachionus koreanus. Mar Pollut Bull 14:913–921. https://doi.org/10.1007/s11051-012-0913-6
Xia B, Chen BJ, Sun XM, Qu KM, Ma FF, Du MR (2015) Interaction of TiO2 nanoparticles with the marine microalga Nitzschia closterium: growth inhibition, oxidative stress and internalization. Sci Total Environ 508:525–533. https://doi.org/10.1016/j.scitotenv.2014.11.066
Xiong B, Cheng J, Qiao YX, Zhou R, He Y, Yeung ES (2011) Separation of nanorods by density gradient centrifugation. J Chromatogr A 1218:3823–3829. https://doi.org/10.1016/j.chroma.2011.04.038
Yue Y, Li XM, Sigg L, Suter MJ, Pillai S, Behra R, Schirmer K (2017) Interaction of silver nanoparticles with algae and fish cells: a side by side comparison. J Nanobiotechnol 15:16–27. https://doi.org/10.1186/s12951-017-0254-9
Zhu XS, Chang Y, Chen YS (2010a) Toxicity and bioaccumulation of TiO2 nanoparticle aggregates in Daphnia magna. Chemosphere 78(3):209–215. https://doi.org/10.1016/j.chemosphere.2009.11.013
Zhu XS, Wang JX, Zhang XZ, Chang Y, Chen YS (2010b) Trophic transfer of TiO2 nanoparticles from daphnia to zebrafish in a simplified freshwater food chain. Chemosphere 79:928–933. https://doi.org/10.1016/j.chemosphere.2010.03.022
Zhu XS, Zhu L, Chen Y, Tian S (2009) Acute toxicities of six manufactured nanomaterial suspensions to Daphnia magna. J Nanoparticle Res 11:67–75. https://doi.org/10.1007/s11051-008-9426-8
Acknowledgements
We are grateful to the anonymous referees for their valuable comments, constructive suggestions and editorial assistance, which have greatly improved the paper.
Funding
This research was funded by National Natural Science Foundation of China (31872208), the State Key Laboratory of Pollution Control and Resource Reuse under Grant (PCRRF20040) and the University Synergy Innovation Program of Anhui Province under Grant (GXXT-2020-075).
Author information
Authors and Affiliations
Contributions
ML wrote the paper and performed the experiments and analyses. YZ and SF participated in the daily experiments. XZ, YX and XX putted forward suggestions on the design of the experiment and the writing of the paper. All authors contributed to the paper and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Li, M., Zhang, Y., Feng, S. et al. Bioaccumulation and biomagnification effects of nano-TiO2 in the aquatic food chain. Ecotoxicology 31, 1023–1034 (2022). https://doi.org/10.1007/s10646-022-02572-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-022-02572-0