
Abstract
Almost all climate change studies of plants and animals adopt an ‘equilibrium assumption’ that analyses of natural distributions provide reliable estimates of species climatic requirements. Yet commercial forestry trials around the world have shown that many tree species can grow successfully under climatic conditions somewhat different from those of their natural distributions. Under climate change it is reasonable to assume that a long-lived tree species, already well-established at particular sites, may be able to display some of the climatic adaptability shown in trials outside its natural distribution. The purpose of this paper is to outline how some species distribution modelling (SDM) and ecological niche modelling (ENM) studies have estimated species climatic requirements beyond those shown by conventional analyses of just their natural distributions, and to show how recent developments are facilitating these analyses. Some of the earliest SDM studies of trees demonstrated the desirability of assessing species climatic requirements using data from outside, as well as within, their natural distributions. In recent years, with the advent of large biodiversity databases and some revised SDM analysis methods, there has been a revival of interest in measuring species climatic requirements using data from beyond their realized niches. It is recommended that at least for tree species, natural distribution data, and where possible results from plantings beyond natural distributions, should be analysed in climate change studies. When this is not possible, some alternative methods of estimating species climatic requirements are identified and some of their advantages and disadvantages are considered.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The vast majority of analyses of the likely impacts of climate change on both plant and animal species have been “dominated by studies using Ecological Niche Models (ENMs) that project future ranges based on correlative models of current relationships between environmental factors and species distribution” (see subsection 4.3.2.5.2 in IPCC 2014).
This approach was pioneered by Busby (1988) who used the BIOCLIM species distribution modelling (SDM) package (Nix 1986; Booth et al. 2014) to analyse likely climate change impacts in Australia on a tree species (Nothofagus cunninghamii, now known as Lophozonia cunninghamii ( Hook.f. ) Heenan & Smissen, Myrtle or Southern Beech) and two animal species (Potorous longipes Seebeck & Johnson, 1980, Long-footed Potoroo and Macropus antilopinus Gould, 1842, Antilopine Wallaroo). The method involved analysing the species current natural distributions to determine the range of their climatic conditions. A climatic change scenario was then applied to identify areas where climatically suitable conditions were likely to be found in the future. A key assumption of this approach is that species current distributions provide a reliable estimate of their climatic requirements. However, commercial forestry trials around the world have shown that many tree species can often grow successfully under climatic conditions somewhat different from those of their natural distributions (see, for example, Jacobs 1981; CAB International 2005).
The purpose of this paper is to show the importance of assessing tree species climatic requirements not only on the basis of natural distributions, but also if possible using data from plantings outside their natural distributions. This need was recognised in early SDM studies of tree species, and then widely (but not always) overlooked. This paper also outlines how recent biodiversity database and SDM developments are facilitating the estimation of species climatic requirements beyond those indicated by simple analyses of their natural distributions.
The study of the climatic requirements of invasive species is a very active area of research. However, as there is a vast literature on the subject and invasion examples are reasonably well covered in the two SDM/ENM books (see Franklin 2009 and chapter 13 in Peterson et al. 2011) the present paper touches on invasive species only briefly.
While correlative methods have come to dominate SDM/ENM studies, mechanistic methods are also important. They endeavour to describe basic physiological responses rather than estimating requirements only from analyses of occurrences (Kearney and Porter 2009). However, mechanistic models are complex to calibrate and are not well suited to analyses of hundreds of species, so the present paper only considers correlative approaches.
To provide some context for this paper, some of the ecological theory that underpins SDM/ENM studies is first briefly described in the section following this introduction. The emphasis is on trees, as unlike almost any other plant or animal group, commercial trials outside their natural distributions provide a rich source of information about their climatic requirements. Unimproved seed samples taken from natural stands of hundreds of tree species have been widely evaluated in thousands of trials around the world (see, for example, Jacobs 1981; Poynton 1979; CAB International 2005). Foresters do everything practical to reduce competition with other species in these trials. Unlike other plant introduction situations, such as ornamental or botanic gardens, human interventions such as irrigation and/or careful selection of microclimates, are not usually appropriate for commercial forestry trials assessing future broad-scale plantation potential.
2 Underpinning ecological theory prior to the release of the first SDM package in 1984
Important contributions to ecological niche theory were made by Grinnell (1917) and Elton (1927)(see Vetaas 2002 for a brief review of niche studies, as well as Franklin 2009 and Peterson et al. 2011 for more details and post-1984 advances). However, Hutchinson (1957) was the first to define the niche as an n-dimensional hypervolume. He also recognized both bionomic (i.e. resource-related) and scenopoetic (i.e. environmental) variables (Hutchinson 1978). The ‘realized niche’ of Hutchinson (1957) represents the conditions within a species’ natural distribution (i.e. when it is in the presence of other interacting species including positively as well as negatively interacting species e.g. pollinators as well as competitors). The ‘fundamental niche’ of Hutchinson (1957) represents a species ability to survive and reproduce within a broader range of conditions, when not in competition with other species. The fundamental niche is sometimes known as the physiological niche (Franklin 2009). See Austin (2007), and Soberón and Nakamura (2009) for more about niches and distributional areas.
For the purposes of this paper, ‘species climatic requirements’ are taken to mean not only the climatic conditions under which a species grows within its natural distribution, but also the conditions under which it can grow outside its natural distribution when no biotic or movement limitations apply. An important assumption in SDM analyses is that a species is in equilibrium or pseudo-equilibrium with its environment (Guisan and Thuiller 2005). Typically this involves the assumption that the range of conditions within the natural distribution provides a reliable estimate of the species requirements. This equilibrium assumption appears to work reasonably well when climatic conditions are largely static, and there are very many examples of SDM analyses helping to clarify species distributions under current conditions (Franklin 2009). The situation is different with climate change studies, particularly for long-lived species, such as trees. Under climate change a species already well-established at a particular site in its natural distribution, does not have to undergo the competitive processes that led to its establishment at that site. It is reasonable to assume that it may be able to display some of the climatic adaptability that is shown in trials outside its natural distribution, where it is not in competition with other species.
3 Early SDM niche-relevant studies 1984-1998
Modern SDM studies began with the release of the BIOCLIM package in January 1984 (Nix 1986; Booth et al. 2014). A search on Google ScholarTM for ‘BIOCLIM’ since 1984 identifies more than 8000 publications, the great majority of which relate to the BIOCLIM SDM package or its associated variables. The concepts behind the BIOCLIM package are very closely related to the niche concepts of Hutchinson (1957), as it describes climatically suitable environments in n-dimensional space. There were 12 variables used in the first version, including factors such as mean annual temperature, mean annual precipitation, mean maximum temperature of the warmest month and mean precipitation of the driest quarter. Later versions of the program increased the n-dimensional space to include 16, 19, 24 or 35 variables. The 19 BIOCLIM variables available from WORLDCLIM (Hijmans et al. 2005) are still used in many SDM analyses (Booth et al. 2014). SDM analyses of a tree species’ natural distribution can provide an estimate of its realized niche, while evaluation of successful trials or plantation sites outside its natural distribution can provide an estimate, in part at least, of its fundamental niche (Booth et al. 1988). For example, the range of mean annual temperature within the natural distribution of E. cladocalyx F. Muell. in Australia was 14-17oC, but it was grown successfully at some sites in Africa where the mean annual temperature was as high as 21oC.
Some of the early forestry SDM studies have been outlined by Booth et al. (2015, see especially subsection 3.1) and Booth (2016, see the introduction). The importance of these early results for climate change studies was summarised in a paper proposing “a global climatological audit to assist conservation and sustainable development” (Booth 1991). A key sentence stated that “in many cases species can grow successfully in conditions somewhat different from those within their natural distributions” and it was therefore suggested that data from trials outside natural distributions should be collected where possible. The need for collation of data from both natural and planted sites was highlighted in a study by Richardson and McMahon (1992), which provided an extreme example of a species climatic adaptability. They showed that Eucalyptus nitens (Deane and Maiden) Maiden grows in successful plantations in southern Africa “under much warmer and drier conditions than any that exist within the natural range of E. nitens” in Australia.
Hughes et al. (1996) carried out one of the first large scale SDM analyses, in terms of analysing more than 800 eucalypt taxa. They found 41% of eucalypt species have a mean annual temperature range within their natural distribution of less than 2oC and 25% have a range of less than 1oC. However, they acknowledged that “the actual climatic tolerances of many species are wider than the climatic envelope they currently occupy”. This dataset was later used as part of the Thomas et al. (2004) SDM extinction risk study, which has been cited more than 5000 times. Unfortunately, they did not discuss the possibility that the climatic envelopes they measured may not fully represent conditions that are limiting species distributions. In particular, some of the eucalypt species may have climatic requirements beyond those suggested by their natural distributions (see, for example, Booth et al. 1988).
4 SDM climate change studies 1999-2011
In November 2009 ScienceWatchTM published a review of ‘Climate Change and Species Distributions’, which identified 38 key highly cited papers from 1999-2009. Only seven of these papers discuss SDM in relation to the fundamental, as well as the realized, niche. Generally, these accept that “SDMs are de facto quantifying Hutchinson’s realized niche of species” (Guisan and Thuiller 2005). Only two of the 38 papers make any comment about using observations from outside natural distributions to estimate conditions beyond the realized niche. Guisan and Thuiller (2005) briefly mention the use of data from botanical gardens and arboreta, as well as natural distributions, by Vetaas (2002). Pearson et al. (2004) mention a small number of occurrences corresponding to “alien introductions” for one of the species they analysed.
The vast majority of SDM climate change studies, both in the ScienceWatch review and elsewhere, have simply analysed the climatic conditions of species’ natural distributions and then applied climate change scenarios. For example, Thuiller et al. (2005) wrote “ ’the bioclimatic envelope’ describes the conditions under which populations of a species persist in the presence of other biota as well as climatic constraints. Future distributions are projected on the assumption that current envelopes reflect species environmental preferences, which will be retained under climate change”. They followed the equilibrium assumption that species are incapable of coping with climates other than those within their natural distributions. Many researchers have made this assumption simply because, in contrast to many tree species, for most plant and animal species results from trials outside natural distributions are not available.
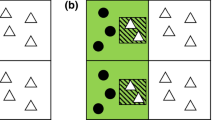
Though the majority of SDM climate change studies have focussed on analysing natural distributions, an intriguing exception amongst the 38 highly cited papers was the work of Svenning and Skov (2004), which involved 55 European tree species. They also analysed natural distributions, but used a minimal rectilinear envelope modelling approach and just three variables related to winter cold, growing season heat and drought to estimate their potential range. That is, the range that would be achieved should all dispersal constraints be overcome (Gaston 2003), but including biotic interactions with other species i.e. the union between the occupied distributional area and the invadable distributional area (see Figure 1 here and also Peterson et al. 2011 for a more detailed discussion and definitions of niche types). Svenning and Skov (2004) found that 36 of the 55 species occupied less than 50% of their potential range. This approach is particularly interesting, as it may provide a generic method to estimate at least part of the fundamental niche beyond the realized niche, if results from plantings outside the natural distribution are not available for a particular species.

A BAM diagram (from Barve et al. 2011) illustrating the three major interacting factors that determine species geographic (G) distributions: Biotic, Abiotic and Movement. The small closed circles indicate presences, while the small open circles indicate absences in terms of the natural distribution. GA is the abiotically suitable area (c.f. Hutchinson’s (1957) fundamental niche), GO is the occupied distributional area (c.f. Hutchinson’s (1957) realized niche) and GI is the invadable distributional area. ‘Potential range’ is the occupied plus invadable areas (i.e. GO + GI). See Peterson et al. (2011) for more information
Two books have provided very useful overviews of methods relevant to SDM/ENM, and briefly considered climate change studies. The book on “Mapping Species Distributions” (Franklin 2009) concentrated on SDM as its name suggests. The early forestry-related SDM research, showing how trials could be used to determine species climatic requirements, was not mentioned in either book, but was very relevant. For example, in a short section (1.5.5) considering the effects of climate change, Franklin (2009) wrote that in using SDMs “it must be assumed that species distributions tend to be in equilibrium with climate..”. This assumption is not necessary under climate change, if results from trials outside a species’ natural distribution are available.
The book by Peterson et al. (2011) on ‘Ecological Niches and Geographic Distributions’ concentrated on ecological niche modelling (ENM). The book concentrates on defining and analysing different niche types, rather than on distribution mapping in particular. Again, the early forestry-related SDM papers were not described, but are very relevant. For example, Peterson et al. (2011) wrote “we will frequently face questions like ‘if the species of interest ranges 10-18oC and the maximum temperature on the calibration landscape is 18oC, then what will the species do under conditions of 20oC?’. In theory, and probably in practice, as well, correlative models are incapable of providing robust projections for non-analog climate conditions..”. This is exactly the type of question that the Booth et al. (1988) SDM niche paper had shown how to address.
5 SDM climate change studies since 2012
Most climate change studies since 2012 have continued to determine species climatic requirements solely on the basis of analyses of their natural distributions. For example, more than 1820 papers since 2012 have mentioned “species distribution model” and “climate change”, but only 271 of these have also mentioned the “fundamental niche”. Many of the studies that mention the fundamental niche continue to be based on analyses of natural distributions, without taking into account information on species climatic requirements derived from plantings beyond their natural distributions. For example, Iverson et al. (2016) wrote that SDMs “.limit their scope to the realized niche (the habitat currently occupied) rather than the fundamental niche (the habitat that could potentially be occupied in the absence of competitors, diseases, or herbivores)…”.
A recent review considered 131 extinction risk analyses from climate change studies published from 1992-2014 for plant and animal species (Urban 2015). These were mainly multispecies SDM-based predictions. Their chief limitations were outlined in terms of often omitting important biological mechanisms “including species interactions, evolution, landscape dispersal barriers, habitat degradation and intraspecific trait variation”. However, no mention was made of the importance of assessing climatic adaptability, beyond that shown by analysis of natural distributions, for long-lived plant species.
While most SDM climate change studies continue to ignore species inherent adaptive capacity, there has been some increasing interest in the problem in recent years. For example, Sax et al. (2013) recognised that “the degree to which species can tolerate or even thrive under conditions beyond their natural distributions” will alter extinction risks under climate change. Similarly, Catullo et al. (2015) acknowledged that “species often have physiological limits beyond the range of environmental conditions experienced in their present realized niche..”. Many tree species exhibit considerable phenotypic plasticity and Prober et al. (2016) have reviewed recent and ongoing studies with eucalypts.
The development of major biodiversity databases has made analyses of not only natural distributions, but also of invasive occurrences and/or plantings outside natural distributions easier. The Global Biodiversity Information Facility (GBIF, www.gbif.org, Gaiji et al. 2013) has led the way in collation of worldwide data. GBIF was established in 2001, and currently has over 600 million records of species occurrence. At the national scale, the Atlas of Living Australia (ALA, www.ala.org.au, Belbin and Williams 2015), which was established in 2007, has been an outstanding achievement. It has been used by GBIF as a basis to assist development of other national biodiversity databases, such as those of France and Spain. Until 2016, GBIF included an SDM analysis facility (a variation of BIOCLIM) in its interface, but with the increasing volume of records it is now concentrating on providing occurrence data. In contrast, the ALA not only provides occurrence data, currently more than 64 million occurrence records, but also nearly 500 layers of gridded surfaces for various environmental, social and political variables. As well as plotting facilities, the ALA provides sophisticated SDM analysis facilities, including a version of the Maxent SDM (Phillips et al. 2006). Maxent has replaced BIOCLIM as one of the most widely used SDM packages. Maxent provides information about the responses of particular variables, rather than the simple ranges of suitable conditions provided by BIOCLIM.
Both the GBIF and ALA lack information from forestry species elimination trials, though occurrences related to plantations and invasive tree species are sometimes available. So their data and facilities can be used to develop improved descriptions of tree species climatic requirements at least for some of the most widely grown species (Booth 2014). The ALA is also complemented by the Biodiversity and Climate Change Virtual Laboratory (BVCCL). Together these facilitate climate change analyses using distributional data from the ALA, and climate change scenario data as well as analysis methods via the BCCVL (Hallgren et al. 2016).
Early and Sax (2014) have used information from multiple databases, including GBIF, to compare the native (European) and naturalized (USA) distributions of 51 plant species’ climatic conditions. They found that for 22 species most of their naturalized ranges fell outside those of their natural distributions.
Zhang et al. (2016) have also used an ensemble of SDM analyses with data from GBIF, as well as data from forest inventory plots, to assess the likely effects of climate and land-use change on 7465 seed plant taxa in North America. By including data for the whole USA, as well as Canada and Mexico, they encompassed observations for plantings outside natural distributions for species such as Pinus radiata D. Don.
Bocsi et al. (2016) used SDM analyses and data from GBIF, and several other herbarium compendia, to compare natural distributions in the United States with non-native ranges for 144 non-invasive plant species. The title of their paper “Plants’ native distributions do not reflect climatic tolerance“ is indicative of their conclusions. These showed that 86% of the species had non-native occurrences in climates outside those of their natural distributions.
A recent study, comparing distributional data from three tree species over the last 10,000 years, has come to somewhat similar conclusions. In their paper, which includes two of the most highly cited authors in the SDM field (Guisan and Araújo) as co-authors, (Cheddadi et al. 2016) wrote “predicting future species distributions should not solely be based on modern realized niches”.
6 Measuring tree species climatic requirements
One essential step and five options for assessing a tree species climatic requirements are outlined below. The first step is to analyse conditions within its natural distribution. This provides a basic analysis of the species climatic envelope or realized niche (i.e. when in presence of other interacting species). Five optional methods for estimating the possibly greater range of species climatic requirements are then outlined (i.e. including parts of the fundamental niche).
6.1 Natural distribution
Modern biodiversity databases, such as the ALA and GBIF make access to natural distribution data relatively easy. However, some caution is needed and Anderson et al. (2016) have provided a detailed discussion of the potential pitfalls of using data from GBIF in SDM/ENM analyses. Heap (2016) has described a program designed to assist refining data from sources such as GBIF and demonstrated its use with a dataset for Abies fraseri (Pursh) Poir (Fraser fir).
6.2 Commercial forestry trials outside the natural distribution
Data from trials outside natural distributions may be used to refine knowledge of tree species climatic requirements. The approach used by Booth et al. (1988) was to collate information from published papers and reports of commercial forestry trials. In contrast to information from invasive studies an advantage of using data from commercial trials is that competition with other species is reduced as far as possible. Forest scientists generally use sites typical of broad areas and, aside from initial watering in, irrigation is rarely used. Disadvantages include the fact that collating individual observations from the literature can be slow, and the results of many trials are only available in grey literature sources, such as unpublished internal reports.
6.3 Botanic gardens or arboreta
Data from botanical gardens or arboreta (see Vetaas 2002) may be used. However, this approach should be used with great caution as high levels of management intervention may limit the value of the observations. For example, careful site selection and/or permanent irrigation may produce conditions that are distinctly different to interpolated climatic estimates. An advantage is that the PlantSearch database can be interrogated to find if a particular plant is being grown at any of more than a thousand contributing institutions (https://www.bgci.org/plant_search.php). However, to protect rare plants, approval is required for access to locational data, and further approval is required from individual providers to use data in publications (Booth 2014).
6.4 Biodiversity databases
Data on occurrences outside natural distributions may be obtained from biodiversity databases such as the ALA and GBIF (see, for example, Zhang et al. 2016). In some cases this may include observations for invasive species. The main advantage of using data from biodiversity databases is that these data are quickly and easily available. The main disadvantage is that there are many species in GBIF for which information for plantings outside their natural distributions are not readily available. Not surprisingly biodiversity databases tend to focus on natural occurrences. Information from forestry trials and/or commercial plantations are not generally captured, unless a species becomes invasive.
6.5 Growth analyses
A recent climate change study of forests in southern Germany has used data from ecological niche analyses and tree ring analyses to complement conventional SDM analyses (Walentowski et al. 2017). Using these it was possible to infer that “the oak and beech forests that currently dominate in the area are not yet at their climatic limit”, though confirming these conclusions with transplant experiments would be desirable.
6.6 Unconventional SDM/ENM analyses
Species inherent adaptive capacity may be estimated using unconventional SDM/ENM analyses, for example, the potential range estimation procedure described by Svenning and Skov (2004). A similar approach was used by Nogués-Bravo et al. (2014) and another variation on the method was tried for three eucalypt species (Booth 2016). Results have been encouraging. For example, the method described by Booth (2016) identified climatically suitable areas in Western Australia for E. globulus subsp. globulus Labill., where it is a major plantation species, based only on analysis of its natural distribution in colder and wetter locations in eastern Australia. However, it is likely that the fundamental niche is wider than the realized niche in some directions but not others, so further testing is needed to demonstrate that these methods are reliable.
Phillips and Dudik (2008) described some extensions to the Maxent package, including ‘hinge features’, which some researchers have used to estimate species fundamental niches. In comparison to other options, hinge features provide smoother model fits (Elith et al. 2011). These do not overfit the data, so may provide better estimates of the fundamental niche. González-Orozco et al. (2016) were aware of the climatic adaptability problem and used Maxent’s hinge features to “more closely approximate the species fundamental niche”, rather than their realized niche for more than 600 eucalypt species. However, they tested the fit of their model against natural distribution data. It would have been desirable, for at least a few species, to test how well the SDM method was predicting species climatic requirements, as indicated by results from both the natural distributions and plantings outside the natural distributions (see Booth 2017 for more details).
6.7 Combined approaches
Some of the preceding options can be combined. For example, McKenney et al. (2007) gathered more than a million observations for 286 North American species. They used natural distribution data derived from Canadian provincial and US Forest Service sources, as well as information for planted locations obtained from botanical gardens, herbaria, master gardeners and horticultural society members.
7 Discussion and conclusions
The importance of assessing the climatic requirements of tree species, beyond those determined from analyses of their natural distributions, was well understood in early SDM papers. However, almost all SDM climate change studies of plant and animal species between 1988 and 2014 only estimated species climatic requirements on the basis of analyses of their natural distributions (see, for example, studies reviewed by Guisan and Thuiller 2005; Franklin 2009; Peterson et al. 2011; Urban 2015). Notable exceptions, which made efforts to estimate conditions beyond the realized niche, included the forest-related studies of Vetaas (2002), McKenney et al. (2007) and Svenning and Skov (2004). Since 2014, there has been increasing interest in using data from biodiversity databases, such as the GBIF and the ALA, to help estimate climatic requirements beyond those shown by simple analyses of natural distributions (see, for example, Booth 2014; Early and Sax 2014; Bocsi et al. 2016; Zhang et al. 2016; Boiffin et al. 2017).
To learn more about tree species climatic adaptability it would be desirable to establish new transplant experiments for many tree species. However, this would be expensive and would take several years to yield useful results. Consequently, one of the greatest needs is to capture some of the wealth of existing data from commercial introduction trials of species and provenances. Given that these often well-designed and replicated trials were specifically created in order to understand the climatic requirements of tree species and provenances it is surprising how little use has been made of these data in climate change studies. Commercial forestry has largely moved on from the species elimination and provenance trial phases, using unimproved material, to selective breeding of a relatively small number of species (Eldridge et al. 1993; Harwood 2011). Results concerning the climatic tolerances of poorer performing species and provenances are generally of no commercial interest, but may be extremely useful in helping to predict how natural stands may respond under climate change.
Fortunately, in some cases the information from all species assessed in trials has been well recorded and published in detail. The book by Poynton (1979) on eucalypt trials in southern Africa is an outstanding example. Detailed data are provided for 88 species and 271 trial locations. At the time it was written reliable climatic interpolation surfaces were not available, so only partial climatic data are included. However, as latitude, longitude and elevation data are provided, reliable estimates of climatic conditions can now be made. These data could be made available for importing into biodiversity databases. A greater problem is the vast amount of data from other trials around the world that have not been published, but are left in filing cabinets, or worst still, have been discarded. Nevertheless, collating available information from forestry trials could assist climate change studies.
A coordinated international effort should be undertaken to collate information from tree trials and to make these available for climate change analyses. As a proof of concept, data for about a dozen of the most widely planted eucalypts species could be assessed first. The project could involve the Australian Tree Seed Centre at CSIRO, which was responsible for distributing most of the eucalypt seed used in international trials. The ALA and BVCCL might contribute to the analysis of the data. If results were encouraging, data for a greater range of eucalypt species could be collated. Eventually GBIF might support collation of data for other genera, such as the pines.
Commercial tree species trials provide valuable information about conditions under which species will survive and grow, but a limitation is that they rarely provide information about successful reproduction. It is often the case that trees may grow successfully under conditions different from those of their natural distributions, but warmer winters may reduce flowering and they may set seed either poorly or not at all (see, for example, Germishuizen and Gardner 2015). In the context of climate change it would be useful to collect more data from trials about successful flowering and seeding. In the meantime, many eucalypt species can grow for hundreds of years (see Table 1.1 in Williams and Brooker 1997), so in terms of climate change, survival of existing individual trees over the coming decades is the critical issue.
Any project assessing likely climate change impacts on any plant or animal species should carefully consider if climatic requirements beyond those shown by just analyses of natural distributions need to be examined. There is some evidence that just SDM analyses of natural distributions may be appropriate for some highly mobile species, such as birds (Russell et al. 2015). However, the influence of competitors is also likely to be important, so more research is needed to determine which groups of species, other than trees, require estimates of climatic requirements in addition to conventional assessments of their climatic requirements derived solely from natural distributions.
For tree species it is recommended that efforts should be made in climate change studies to estimate their climatic requirements beyond just conventional SDM analyses of their natural distributions. If adequate data are available for occurrences beyond natural distributions from published trials and/or sources such as GBIF, those are to be preferred. If not, then some effort should be made to estimate species climatic requirements using methods similar to those of Svenning and Skov (2004) or Phillips and Dudik (2008). However, it is important that the reliability of such analysis methods should be demonstrated where possible, using some examples of well-known species for which adequate occurrences beyond their natural distributions are available to validate the method used.
References
Anderson RP, Araújo M, Guisan A, Lobo JM, Martinez-Meyer E, Peterson AT, Soberón J (2016) Final report of the task group on GBIF data fitness for use in distribution modelling. Global Biodiversity Information Facility, Geneva http://www.gbif.org/resource/82612
Austin M (2007) Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol Model 200:1–19
Barve N, Barve V, Jimenez-Valverde A, Lira-Noriega A, Maher SP, Peterson AT, Soberón J, Villalobos F (2011) The crucial role of the accessible area in ecological niche modelling and species distribution modelling. Ecol Model 222:1810–1819
Belbin L, Williams KJ (2015) Towards a national bio-environmental data facility: experiences from the Atlas of Living Australia. Int J Geogr Inf Syst 30:108–125
Bocsi T, Allen JA, Bellemare J, Kartesz J, Nishino M, Bradley B (2016) Plants’ native distributions do not reflect climatic tolerance. Divers Distrib 22:615–624
Boiffin J, Badeau V, Bréda N (2017) Species distribution models may misdirect assisted migration: insights from the introduction of Douglas-fir to Europe. Ecol Appl 27:446–457
Booth TH (1991) A global climatological audit to assist conservation and sustainable development. Proceedings of 10th World Forestry Congress, Paris, 2:65-70
Booth TH (2014) Using biodiversity databases to verify and improve descriptions of tree species climatic requirements. Forest Ecol Manag 315:95–102
Booth TH (2016) Estimating potential range and hence climatic adaptability in selected tree species. Forest Ecol Manag 366:175–183
Booth TH (2017) Impacts of climate change on eucalypt distributions in Australia: an examination of a recent study. Aust Forestry. https://doi.org/10.1080/00049158.2017.1365402
Booth TH, Nix HA, Hutchinson MF, Jovanovic T (1988) Niche analysis and tree species introduction. Forest Ecol Manag 23:47–59
Booth TH, Nix HA, Busby JR, Hutchinson MF (2014) BIOCLIM: the first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers Distrib 20:1–9
Booth TH, Broadhurst LM, Pinkard E, Prober SM, Dillon SK, Bush D, Pinyopusarerk K, Doran JC, Ivkovisch M, Young AG (2015) Native forests and climate change: Lessons from eucalypts. Forest Ecol Manage 247:18–29
Busby JR (1988) Potential implications of climate change on Australia’s flora and fauna. In: Pearman GI (ed) Greenhouse; Planning for Climate Change. CSIRO, Melbourne, pp 387–398
CAB International (2005) Forestry Compendium. CABI, Wallingford CD-ROM
Catullo RA, Ferrier S, Hoffmann AA (2015) Extending spatial modelling of climate change responses beyond the realized niche: estimating and accommodating, physiological limits and adaptive evolution. Global Ecol Biogeogr 24:1192–1202
Cheddadi R, Araújo MB, Maiorano L, Edwards M, Guisan A, Carré M, Chevalier M, Pearman PB (2016) Temperature range shifts for three European tree species over the last 10,000 years. Front Plant Sci 7:doi:https://doi.org/10.3389/fpls.2016.01581
Early R, Sax DF (2014) Climatic niche shifts between species’ native and naturalized ranges raise concern for ecological forecasts during invasions and climate change. Global Ecol Biogeogr 23:1356–1365
Eldridge K, Davidson J, Harwood C, van Wyk G (1993) Eucalypt Domestication and Breeding. Clarendon Press, Oxford
Elith J, Phillips SJ, Hastie T, Dudik M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57
Elton CS (1927) Animal ecology. Sidgwick and Jackson, London
Franklin J (2009) Mapping species distributions: spatial inference and prediction. Cambridge University Press, Cambridge
Gaiji S, Chavan V, Arino AH, Otegui J, Hober D, Sood R, Robles E (2013) Content assessment of the primary biodiversity data published through GBIF network: status, challenges and potentials. Biodiv Info 8:94–172
Gaston KJ (2003) The Structure and Dynamics of Geographic Ranges. Oxford University Press, Oxford
Germishuizen I, Gardner RAW (2015) A tool for identifying potential Eucalyptus nitens seed orchard sites based on climate and topography. South Forests 77:123–130
González-Orozco C, Pollock LJ, Thornhill AH, Mischler BD, Knerr N, Laffan SW, Miller JT, Roscuer DF, Faith DP, Nipperess DA, Kujala H, Linke S, Butt N, Kűlheim C, Crisp MD, Gruber B (2016) Phylogenetic approaches reveal biodiversity threats under climate change. Nat Clim Change 6:1110–1114
Grinnell J (1917) The niche-relationships of the California Thrasher. Auk 34:427–433
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8:993–1009
Hallgren W, Beaumont L, Bowness A, Chambers L, Graham E, Holewa H, Laffan S, Mackey B, Nix H, Price J, Vanderwal J, Warren R, Weis G (2016) The Biodiversity and Climate Change Virtual Laboratory: Where ecology meets big data. Environ Modell Softw 76:182–186
Harwood C (2011) Introductions: doing it right. In: Walker J (ed) Developing a Eucalypt Resource. Learning from Australia and Elsewhere. Wood Technology Research Centre, University of Canterbury, New Zealand, pp 43–54
Heap MJ (2016) Introducing BioSARN-an ecological niche model refinement tool. Ecol Evol 6:5808–5816
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978
Hughes L, Cawsey EM, Westoby M (1996) Climatic range sizes of Eucalyptus species in relation to future climate change. Global Ecol Biogeogr 5:23–29
Hutchinson GE (1957) Concluding remarks. Cold Spring Harb Sym 22:415–427
Hutchinson GE (1978) An Introduction to Population Ecology. Yale University Press, New Haven
IPCC (2014) Climate Change 2014: impacts, adaptation, and vulnerability. part a: global and sectoral aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, 1132 pp
Iverson LR, Thompson FR, Matthew S, Peters M, Prasad A, Dijak WD, Fraser J, Wang WJ, Hanberry B, He H, Janowiak M, Butler P, Brandt L, Swanston C (2016) Multi-model comparison on the effects of climate change on tree species in the eastern U.S.: results from an enhanced niche model and process-based ecosystem and landscape models. Landscape Ecol 32:1327–1346
Jacobs MR (1981) Eucalypts for Planting, 2nd edn. FAO, Rome
Kearney M, Porter W (2009) Mechanistic niche modelling: combining physiological and spatial data to predict species’ ranges. Ecol Lett 12:334–350
McKenney DW, Pedlar JH, Lawrence K, Campbell K, Hutchinson MF (2007) Potential impacts of climate change on the distribution of North American trees. BioScience 57:939–948
Nix HA (1986) In: Longmore R (ed) A biogeographic analysis of Australian elapid snakes. Atlas of Elapid Snakes of Australia. Australian Flora and Fauna Series 7. Bureau of Flora and Fauna, Canberra, pp 4–15
Nogués-Bravo D, Pulido F, Araujo MB et al (2014) Phenotypic correlates of potential range size and range filling in European trees. Perspect Plant Ecol 16:219–227
Pearson RG, Dawson TP, Liu C (2004) Modelling species distributions in Britain: a hierarchical integration of climate and land-cover data. Ecography 27:285–298
Peterson AT, Soberon J, Pearson RG, Anderson RP, Martinez-Meyer E, Nakamura M, Araujo MB (2011) Ecological niches and geographic distributions. Princeton University Press, Princeton
Phillips SJ, Dudik M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modelling of species geographic distributions. Ecol Model 190:231–259
Poynton RJ (1979) Tree Planting in Southern Africa. Vol. 2 The Eucalypts. Department of Forestry, Southern African Regional Commission for the Conservation and Utilisation of the Soil, Pretoria
Prober SM, Potts BM, Bailey T et al (2016) Climate adaptation and ecological restoration in eucalypts. Roy Soc Vic 128:40–53
Richardson DM, McMahon JP (1992) A bioclimatic analysis of E. nitens to identify potential planting regions in southern Africa. S Afr J Sci 88:380–387
Russell DJF, Wanless S, Collingham YC, Anderson BJ, Beale C, Reid JB, Huntley B, Hamer KC (2015) Beyond climate envelopes: bio-climate modelling accords with observed 25-year changes in seabird populations of the British Isles. Divers Distrib 21:211–222
Sax DF, Early R, Bellemare J (2013) Niche syndromes, species extinction risks, and management under climate change. Trends Ecol Evol 28:517–523
Soberón J, Nakamura M (2009) Niches and distributional areas: concepts, methods and assumptions. P Natl Acad Sci USA 106:19644–19650
Svenning J-C, Skov F (2004) Limited filling of the potential range in European tree species. Ecol Lett 7:565–573
Thomas CD, Cameron A, Green RE et al (2004) Extinction risk from climate change. Nature 427:145–147
Thuiller W, Lavorel S, Araujo MB, Sykes MT, Prentice IC (2005) Climate change threats to plant diversity in Europe. P Natl Acad Sci USA 102:8245–8250
Urban MC (2015) Accelerating extinction risk from climate change. Science 348:571–573
Vetaas OR (2002) Realized and potential climatic niches: a comparison of four Rhododendron tree species. J Biogeogr 29:545–554
Walentowski H, Falk W, Mette T, Kunz J, Bräuning A, Meinardus C, Zang C, Sutcliffe L, Leuschner C (2017) Assessing the future suitability of tree species under climate change by multiple methods: a case study in southern Germany. Ann For Res 60:101–126
Williams JE, Brooker MIH (1997) Eucalypts: an introduction. In: Williams JE, Woinarski JCZ (eds) Eucalypt Ecology. Cambridge University Press, Cambridge, pp 1–15
Zhang J, Nielsen SE, Chen Y, Georges D, Qin Y, Wang S-S, Svenning J-C, Thuiller W (2016) Extinction risk of North American seed plants elevated by climate and land-use change. J Appl Ecol 54:303–312
Acknowledgements
This paper was funded by CSIRO. I am grateful to Libby Pinkard, Sadanandan Nambiar and David Bush, as well as the anonymous referees, for their comments on earlier versions of this paper. Thanks to Elsevier for permission to use Fig. 1 under the terms of their STM permissions guidelines.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Booth, T.H. Assessing species climatic requirements beyond the realized niche: some lessons mainly from tree species distribution modelling. Climatic Change 145, 259–271 (2017). https://doi.org/10.1007/s10584-017-2107-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-017-2107-9