
Abstract
Context
Species distribution models (SDM) establish statistical relationships between the current distribution of species and key attributes whereas process-based models simulate ecosystem and tree species dynamics based on representations of physical and biological processes. TreeAtlas, which uses DISTRIB SDM, and Linkages and LANDIS PRO, process-based ecosystem and landscape models, respectively, were used concurrently on four regional climate change assessments in the eastern Unites States.
Objectives
We compared predictions for 30 species from TreeAtlas, Linkages, and LANDIS PRO, using two climate change scenarios on four regions, to derive a more robust assessment of species change in response to climate change.
Methods
We calculated the ratio of future importance or biomass to current for each species, then compared agreement among models by species, region, and climate scenario using change classes, an ordinal agreement score, spearman rank correlations, and model averaged change ratios.
Results
Comparisons indicated high agreement for many species, especially northern species modeled to lose habitat. TreeAtlas and Linkages agreed the most but each also agreed with many species outputs from LANDIS PRO, particularly when succession within LANDIS PRO was simulated to 2300. A geographic analysis showed that a simple difference (in latitude degrees) of the weighted mean center of a species distribution versus the geographic center of the region of interest provides an initial estimate for the species’ potential to gain, lose, or remain stable under climate change.
Conclusions
This analysis of multiple models provides a useful approach to compare among disparate models and a more consistent interpretation of the future for use in vulnerability assessments and adaptation planning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Average global land temperatures have risen approximately 1 °C since the mid-1950s (Rohde et al. 2012) and increasingly rapid warming is projected to occur during the twenty first century (IPCC 2014). Predictions of future precipitation also indicate seasonal changes, but the timing and magnitude of precipitation events are not well understood. A more vigorous hydrologic cycle is expected to result in heavy precipitation events that will increasingly provide a larger proportion of the total annual precipitation. Indeed, this pattern has been observed across the eastern United States with an increase of 27, 37, and 71 % of precipitation falling in very heavy events (the heaviest 1 %) from 1958 to 2012 for the Southeast, Midwest, and Northeast, respectively (Melillo et al. 2014). The increased frequency of these events is expected to result in more runoff and floods (Lenderink and van Meijgaard 2008) as well as more and longer periods without rain and droughts (IPCC 2014). Indeed, the elevated number, severity, and impact of disastrous heat and precipitation events have been linked to the human influence on climate (Coumou and Rahmstorf 2012) and amplified heating of the Arctic (Seminov 2012).
Climate is the primary driving force at a coarse scale for the location, composition, and productivity of forests (Woodward and Williams 1987), and past changes in climate and tree distributions, responding individually as species (not communities) are evident in the paleoecological record (e.g., Webb III and Bartlein 1992; DeHayes et al. 2000). There is already evidence that tree species have moved to higher altitudes (Beckage et al. 2008; Holzinger et al. 2008; Lenoir et al. 2008) and latitudes (Woodall et al. 2009; Boisvert-Marsh et al. 2014; Reich et al. 2015) in recent decades, and future climate changes are expected to result in further range shifts.
A number of different modeling approaches have been developed to project future distributions of trees and forests under a changing climate. Species distribution models (SDMs) establish a statistical relationship between the current distribution of species and key attributes of their habitats and are used to predict how species distributions might shift as climate change affects those attributes. SDMs are computationally inexpensive and typically provide projections for many species over large areas. SDMs carry caveats, discussed elsewhere, that limit their scope to the realized niche (the habitat currently occupied) rather than fundamental niche (the habitat that could potentially be occupied in the absence of competitors, diseases, or herbivores) and fail to consider migration or successional patterns of forest growth (e.g., Wiens et al. 2009; Iverson and McKenzie 2013). Rather, SDMs assess whether species will be exposed to bioclimatic conditions outside their realized ranges into the future, suggesting a potential for changes to suitable habitats.
Process models simulate ecosystem and tree species dynamics based on mathematical representations of physical and biological processes. Process models can simulate future change in tree species dispersal, succession, biomass and nutrient dynamics over space and time, often with competition considered among several species; they most often operate at a finer pixel size and require more computational power than a SDM. Process models also have assumptions and uncertainties that should be taken into consideration when applying results to management decisions (Lawler et al. 2006; Kennedy and Ford 2011). For example, they rely on species parameterization and empirical and theoretical relationships that are specified by the modeler using imperfect knowledge, and any uncertainties in these relationships can be compounded over time and space, leading to potential biases.
SDM and process models have forecasted tree habitats to move variously, but generally poleward, with climate change (Iverson et al. 2008b; Keith et al. 2008; Dobrowski et al. 2011; McKenney et al. 2011). Changes in potential tree habitat are often modeled to show much larger changes that can be expected to be colonized within 100 years through unassisted migration (e.g., Iverson et al. 2004; Serra-Diaz et al. 2014). Uncertainty in data inputs and model outputs, as well as other challenges will continue to confront species modeling (Pearson et al. 2006; Thuiller et al. 2008; Xu et al. 2009). Meanwhile, multiple approaches are being developed in attempts to improve projections (Morin and Thuiller 2009; Franklin 2010; Iverson et al. 2011; Matthews et al. 2011). Assuredly, within the landscape of interest, managers and decision makers will need to use model outputs in concert with an evaluation of legacies in land use, past ecological trends, and local conditions prior to determining a management prescription. For example, identification of potential refugia for species losing habitat or possible sites for encouraging species expansion or new establishment may be important.
As part of a Climate Change Response Framework that is being applied across the Northeastern United States (Swanston and Janowiak 2012), the predictions from TreeAtlas (SDM) and process models Linkages and LANDIS PRO were applied using consistent climate and forest inventory data sets and future climate scenarios to assess changes in tree species importance at the end of this century. Projects within this area have been the focus of extensive ecosystem vulnerability assessment and synthesis efforts coordinated by the Northern Institute of Applied Climate Science (Brandt et al. 2014; Butler et al. 2015). These efforts effectively created a platform for scientists and managers to provide early feedback on model outputs in each region, thereby improving model results, and were also the basis for adaptation efforts towards informed forest management in the face of climate change (Janowiak et al. 2014).
Our goal was to derive a robust assessment of potential changes in trees species importance or abundance in response to climate change through multi-model inference. We compared predictions from a species distribution model (TreeAtlas, or DISTRIB), a hybrid empirical-process model (Linkages), both of which projected habitat suitability in 2100, and a process-based landscape change model (LANDIS PRO), which projected forest composition and structure in 2100 and 2300, for four regions of the eastern United States using two climate change scenarios. We modeled predictions for two ‘bookends’ of climate scenarios for each forest change model, and assessed species change that were not dependent on any one model framework. This allowed us to address uncertainties in future climate by considering climate scenarios with lesser and greater amounts of change in precipitation and temperature. We hypothesized that TreeAtlas and Linkages, both projecting suitable habitat, should be more in agreement with each other than with the LANDIS PRO succession model outputs, but that as succession is allowed to proceed within LANDIS PRO, agreement with the suitable habitat outputs would improve. Furthermore, we expected LANDIS PRO and Linkages predictions to be in closer agreement over longer time periods as stand dynamics equilibrated because species specific establishment probabilities and maximum growing space were parameterized in LANDIS based on Linkages predictions for each climate scenario.
Methods
Study areas
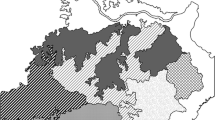
The study area consisted of much of the forested land in the central and northern portions of the eastern United States (Fig. 1). The area was modeled in four regions: the Central Hardwoods (CH), Central Appalachians (CA), Mid-Atlantic (MA), and New England (NE). The MA and NE are much larger in area and had more species (24) modeled than the other two regions (20 for CH and 15 for CA, Table 1) in this study. The climates in the regions represented north–south and east–west gradients. Mean annual temperatures ranged from 13.0 °C in CH to 6.4 °C in NE at present (1981–2010) and are projected to increase to 14.2–17.5 °C in CH and 7.6–10.3 °C in NE, depending on scenario, by the end of century (Table 1). Estimates of annual precipitation did not vary widely across regions or scenarios, although CH is modeled to have higher precipitation under the mild scenario of climate change and lower under the harsh scenario, while the other regions had estimates of level or slightly higher annual precipitation under either scenario. However, with the warmer conditions, and larger precipitation events followed by longer drought periods, greater physiological “hot droughts” can be expected to increase forest mortality (Allen et al. 2015). Elevations are substantially higher and more variable in the CA (mean maximum elevation of 570 m) and lower in the CH (288 m mean maximum elevation). Edaphic variables also provide important features which describe ecological conditions and, in the models, create variation across the regions (Table 1).

Locations of the four study regions in the eastern United States for which we compare projected changes in tree species importance
Data sources
The same climate scenarios and downscaled climate data were used for the TreeAtlas, Linkages, and LANDIS PRO models. We selected two climate scenarios that generally spanned the range of potential future conditions by pairing general circulation models (GCM) with emission scenarios. The Parallel Climate Model (PCM, the mild scenario) has relatively low sensitivity to changes in greenhouse gas concentrations (PCM, Washington et al. 2000) and was combined with the B1 emission scenario, which represented a rapid conversion by humans to low carbon energy sources (Nakicenovic et al. 2000). The Geophysical Fluid Dynamics Laboratory (GFDL, the harsh scenario) is moderately sensitive to changes in greenhouse gas concentrations (Delworth et al. 2006) and was combined with the A1FI emission scenario, which predicted the highest greenhouse gas emissions (Nakicenovic et al. 2000). Therefore our PCM-B1 and GFDL-A1FI scenarios represent bookends to potential future climates out to 2100. A statistically downscaled dataset at ~12 km resolution was used to calculate daily precipitation and mean, maximum, and minimum temperature through 2100 for the two scenarios (Stoner et al. 2013), which provided the climate inputs to the forest models described here. All three model approaches used U.S. Forest Service Forest Inventory and Analysis data (FIA) to model current habitat or initialize current conditions for simulations (Woodall et al. 2010).
TreeAtlas
TreeAtlas is a collection of DISTRIB models that use a statistical approach (Random Forest) to predict current and potential future suitable habitat (Prasad et al. 2006). Model inputs include US Forest Service forest inventory data and other environmental variables (Iverson et al. 2008b; Prasad et al. 2016). Each species model was given a reliability rating, using four statistical and spatial metrics, to provide an indication of the quality of the species model; species with fewer FIA observations tended to have lower reliability (Iverson et al. 2011). Each species also was rated, via literature, on 9 biological characteristics that may assist in adaptation to climate change as well as its capacity to deal with 12 disturbance types (Matthews et al. 2011). The outputs are presented in a web-based Climate Change Tree Atlas, which incorporates a diverse set of information about potential shifts in the distribution and abundance of tree species’ habitat in the eastern United States over the next century (Landscape Change Research Group 2014). Importantly, TreeAtlas projects where suitable habitat may change for a particular species, but does not project how long it may take for a species’ range to change. The actual rate of migration into newly suitable habitat will be influenced by large time lags, dispersal and establishment limitations, and availability of refugia. These have been evaluated in other work for a few species through our model SHIFT and the multi-stage approach, but were not used here because of insufficient numbers of modeled species (Iverson et al. 2004; Prasad et al. 2013, 2016).
The models use inputs of tree abundance, climate, and the environment to simulate species’ habitats into 20 × 20 km cells; details are presented elsewhere (Iverson et al. 2008b; Landscape Change Research Group 2014; Butler et al. 2015). Because the forest inventory plots are non-biased and extensive across the eastern US, the 30 common species reported here have high model reliability within TreeAtlas.
Linkages
Linkages is a hybrid empirical-physiological model that simulates forest ecosystem functions such as soil–water balance, litter decomposition, nitrogen cycling, soil hydrology, and evapotranspiration to predict tree establishment, growth, and death. Model inputs include daily maximum and minimum temperature, precipitation, wind speed, solar radiation, soil moisture capacity for multiple soil layers, wilting point, percent rock, percent clay, percent sand, initial organic matter, and nitrogen contents (Wullschleger et al. 2003; Dijak et al. in press). We used Linkages to model the relative performance of individual tree species for the ecological subsections and climate scenarios we considered in this study (for details see Brandt et al. 2014; Butler et al. 2015; Wang et al. (in press); Wang et al. 2015). We predicted the maximum biomass reached for individual species based on 20 replicate simulations for 30 years starting from bare ground on virtual plots that represented 8–10 landforms in each subsection under current climate (1980–2009) and each future climate scenario (2070–2099). We aggregated the biomass estimates from these studies from landforms to subsections and subsections to sections using area-weighted means and calculated change ratios as the quotient of future biomass divided by current biomass. Like TreeAtlas, this application of Linkages provided a prediction of the change in future habitat suitability for individual tree species and is not a simulation of actual forest change.
LANDIS PRO
LANDIS PRO (hereafter Landis) is a forest landscape model that projects changes in forest composition and structure due to processes occurring at species-, stand-, and landscape-levels (Wang et al. 2013, 2014a). We used Landis to simulate changes in forest composition and structure due to succession, dispersal, windthrow, harvest, and climate change for each region and climate scenario (for details see Brandt et al. 2014; Butler et al. 2015; Wang et al. 2015). Landis simulates tree growth, longevity-caused mortality, competition-caused mortality (self-thinning), disturbance-caused mortality, resprouting, fecundity, seed dispersal, and establishment (He et al. 2005). It tracks the number of trees and diameter at breast height (DBH) by species and age cohort in each 270-m raster cell and after establishing initial forest conditions at year 2000 from 1995 to 2005 FIA data including trees >2.54 cm (Wang et al. 2014b). Landis does not directly consider climate, however; effects of climate change were incorporated by estimating climate change impacts on species establishment probabilities (SEP), which affect species colonization, and resources availability (measured as maximum growing space, MGSO), which, in turn, affects tree mortality as a function of climate, soil, and terrain. MGSO and SEP were modeled using Linkages under current climate and each future climate scenario, then linearly interpolated from current climate to the end of the century. We calculated species importance values for each cell at the start of simulations and at year 2100 (hereafter Landis100) and 2300 (hereafter Landis300) under each scenario and the ratio of future to current importance values. However, because the climate scenarios only forecasted climate change through 2100, simulations for years 2100–2300 did not consider climate change beyond 2100 and held SEP and MGSO at year 2100 levels. We recognize the high degree of uncertainty around 300 year projections; indeed a recent publication reports immense climatological changes should humans continue to burn fossil fuels to 2300 (Tokarska et al. 2016). However, we wished to demonstrate the effects of an additional 200 years of stand dynamics in response to the climate change that occurred through 2100 because species turnover can take a long time (Wang et al. 2015). Unlike TreeAtlas and Linkages, Landis simulated stand dynamics, colonization, and extinction; representing projections of expected change in forest composition and structure at 2100 and 2300. We simulated the current level of tree harvest and approximated the silvicultural methods used in the region to make our simulations as realistic as possible (Wang et al. 2015). We created regional management units and parameterized the level of harvest based on FIA data and we varied the amount of basal area harvested in a stand to simulate clearcutting or partial harvest at the pixel level. In addition, we simulated windthrow as a stochastic gap-scale disturbance that affected the oldest trees on an average of 1.4 % of pixels per decade. (Wang et al. 2015, ).
Model assumptions
All models have limitations and assumptions, and are approximations of reality. Each of these models carry the assumptions for the climate change scenarios for GFDL and PCM, and that the FIA data captures the distribution and abundance for the species under study. For TreeAtlas, we assume that the selected variables do in fact reflect the niche requirements of a species, that species are in equilibrium with their suitable habitat, that predictions can be made into novel climates and land covers, that the effects of adaptation and evolution are minimal, and that the effects of biotic interactions (including human interactions) are minimal. For Linkages, we assume that the variables selected and the parameterization of those variables accurately drive the establishment, growth, and death of individual tree species over the first 30 years of growth from bare ground. For Landis, we assume that we have correctly parameterized life histories and physiologies for each species, captured the complexity of interacting biotic and disturbance factors, and controlled cumulative errors associated with the sheer number and diverse spatio-temporal scales of parameters. Despite these limitations, this study represents a pathway to further understand the range of possibilities for tree species under a changing climate.
Analysis
We calculated a future:current change ratio as the ratio of a species’ future importance values (as calculated equally from tree density and basal area in TreeAtlas and Landis, and as biomass in Linkages) to current importance values or biomass. For example, the future:current ratio for species X according to TreeAtlas and GFDL scenario = modeled future importance value for species X at 2100 according to GFDL scenario/current importance for species X. Future values were calculated for 2100 (100 years) for TreeAtlas and Linkages and 2100 and 2300 (100 and 300 years) for Landis. If the change ratio is close to one (i.e., 0.8–1.2), essentially no change in suitable habitat is projected by the models. If the ratio exceeds 1.2, an increase is projected, while if the ratio is less than 0.8, a decrease is projected. We created the following change classes based on change ratios to facilitate interpretation of some results: Large Increase (>2), Small Increase (1.2–2.0), No Change (0.8–1.2), Small Decrease (0.5–0.8), Large Decrease (<0.5). Change ratios were derived using each model in each region for the two climate scenarios to assess overall species vulnerability.
We tallied agreement between the following pairs of models based on change ratios: (1) TreeAtlas & Linkages, (2) TreeAtlas & Landis100, (3) TreeAtlas & Landis300, (4) Linkages & Landis100, (5) Linkages & Landis300, and (6) Landis100 & Landis300. To present a simple, single metric with higher values indicating higher agreement, we created an ordinal scale of 0–4 and assigned 4 points if both change classes were identical = full agreement; 3 points if one class apart (e.g., No Change and Small Increase or Small Decrease); 2 points if two classes apart but still trending in same direction (e.g., No Change and Large Increase or Large Decrease); 1 point if Small Decrease & Small Increase (opposite trend); and 0 points if opposite trend and one or both are Large Decrease or Large Increase. In this way, we assign increasing penalties as the classes are more in disagreement.
We also calculated Spearman’s rank correlations of future:current ratios among combinations of models and scenarios as another measurement of agreement. We report correlations and p-values for the hypothesis that change ratios from a pair of models were not positively correlated and used Holm’s method to adjust p-values to account for familywise error rate with multiple comparisons (Holm 1979), a fairly restrictive adjustment.
To assess association of potential change to their geographic location for the 30 species, we evaluated the mean centers of the species distribution, now and into the future, against the geographic centers of each of the four regions. We calculated the weighted mean centers of species’ current and future habitats based on importance values reported within TreeAtlas (www.nrs.fs.fed.us/atlas/tree/meancenter_latlong2f.html), then calculated the difference between a species center of distribution and the geographic center of our study regions (species mean center—region mean center = mean offset from range center) to assess if a species center was North (>2°N), Central (2°N to 2°S) or South (>2°S) with respect to centers of the four regions. For example, balsam fir is a species with a species mean center to the far North (latitude 46.2°N), while the geographic center for the study region MA is at 41.2°N, a mean offset of 5°, so that the species is classed as North for this region. For species intersecting Canada, only the US portion (for which importance values are available) was used for calculating mean centers, which compressed some latitude differences, but results were robust relative to the regional locations. We included the mean offset from range center in our correlation analysis, and summarized future:current ratios by North, Central, or South latitude classes to assess relationships between model outputs and species geographic positioning.
Lastly, we calculated mean future:current ratios across species and for each model system for both PCM and GFDL scenarios. These ratios represent single estimates of change derived from all three models and the range between PCM and GFDL can be interpreted as a measure of uncertainty across scenarios.
Results
By tabulating change classes for species by region, model, and climate scenarios, some general patterns in species changes were apparent: (1) substantially less change was projected by the models under PCM than GFDL for all models; (2) Landis100 has small changes under both scenarios but Landis300 shows larger changes; (3) more species show projected declines than increases, with a possible exception in New England (Fig. 2).


Comparison of change classes for 30 species for TreeAtlas, Linkages, and Landis100 and Landis300 for PCM and GFDL climate change scenarios, for the New England (NE, N = 24), MidAtlantic (MA, N = 24), Central Appalachians (CA, N = 15), and Central Hardwoods (CH, N = 20) regions. Numbers indicate the future:current ratios, while colors represent the change class, where red = Large Decrease (future:current ratio <0.5), pink = Small Decrease (future:current ratio >0.5 and <0.8), yellow = No Change (future:current ratio >0.8 and <1.2), light green = Small Increase (future:current ratio >1.2 and <2.0), and darker green = Large Increase (future:current ratio >2.0)
TreeAtlas-Linkages had the highest agreement scores on our 0–4 ordinal scale when averaged across species, with 3.4 for PCM and 3.16 for GFDL (Table S1). Next in agreement was Landis100-Landis300 (3.07–3.11), followed by Linkages-Landis300, Linkages-Landis100 ≅ TreeAtlas-Landis300, and TreeAtlas-Landis100. TreeAtlas-Landis300 tended to agree the most for the GFDL scenario (2.78 vs. 2.48 on PCM), while Linkages-Landis300 agreed relatively more for the PCM scenario (2.92 vs 2.60 on GFDL). These relationships generally held across regions. Among the 30 species, there was a variation in agreement, with some of the northernmost species (e.g., black spruce, balsam fir, northern white cedar, and red spruce) showing maximum agreement among models (Table 2, Note: scientific names for all species are presented in Table 2). On the low end of agreement are several pines (shortleaf, pitch, loblolly, and Virginia—the latter two with low prominence in these regions), quaking aspen, post oak, tulip poplar, yellow birch, and American beech. Quaking aspen had model outputs in NE with large increases under Landis100/300 and large decreases in TreeAtlas and Linkages, while the reverse was true for shortleaf pine, and to a lesser extent, post oak, in CH. Many of the oaks, hickories, and maples tended to have moderate agreement; for example, most models show a decline in sugar maple especially in the more southerly regions of CH and CA.
There were 83 species-region combinations when we pooled species for the correlation analysis. The highest correlations between model-scenario pairings were between Landis100 PCM and Landis100 GFDL (Fig. 3; Table S2), which was not surprising because change is predominately driven by succession, not climate, in the first 100 years in Landis (Wang et al. 2015). Landis100 versus Landis300 were also highly correlated regardless of scenario, but especially so within the same scenario. High correlations also occurred between PCM and GFDL scenarios within both TreeAtlas and Linkages across most combinations of species and regions, except for CA with its low species count of 15 (Table S2). There were also highly significant correlations between TreeAtlas and Linkages outputs for both scenarios except for in the CA (Table S2).

Spearman rank correlations future:current ratios of predicted tree species importance or biomass by three models (TreeAtlas, Linkages, Landis) and two climate scenarios (PCM B1, GFDL A1FI) for the four regions (CA, CH, MA, NE) pooled together. Landis was modeled to 2100 (e.g., Landis.gfdl100) and 2300 (Landis.gfdl300). Latitude difference is also included
Importantly, however, correlations between TreeAtlas and Landis increased as Landis simulations increased from 100 to 300 years, especially under the GFDL scenario where the correlation increased from 0.23 to 0.25 (P > 0.05) for 100-year to 0.48-0.51 (P < 0.0001) for 300-year Landis simulations (Fig. 3, Table S2). An increase in correlation would be expected over time because of the role of the Linkages outputs as inputs to Landis, and because Linkages and TreeAtlas outputs were quite highly correlated (r = 0.60–0.77). The same pattern of increasing correlation over time is true for both Linkages and TreeAtlas throughout regions although not always significantly so (Fig. 3; Table S2). Notably however, rank correlations between Landis even at 300 years and the habitat suitability models did not attain very high values, probably because certain species that did not model well fell in highly variable ranks among the models.
There was also a highly significant negative correlation between the latitude difference and change ratios from TreeAtlas and Linkages across species-region combinations (~−0.8, Fig. 3; Table S2). This pattern indicates that as the distance between mean centers of the species and the region center increased, the future:current ratios (~habitat suitability) decreased. The relationship was also highly significant for Landis300 GFDL but not the other outputs from Landis, again indicating a progression towards the habitat models given harsh conditions and 300-year Landis simulations.
When species were evaluated according to their latitudinal differences between the region and mean centers, the 16 species/region combinations categorized as North had very low future:current ratios under both PCM (average = 0.68) and especially GFDL (average = 0.23) scenarios (Table 3). Those combinations categorized as South had future:current ratios >1, indicating increasing tendencies for habitat within their region, while the Central class was between these extremes but tended towards decreasing habitat, especially under GFDL. Graphically, future:current ratios versus latitude difference by species (for TreeAtlas and GFDL scenario) generally shows higher future:current ratios (gains or less loss in habitat) for the most northerly region (NE) and less gain or more loss for the southerly regions CH and CA (Fig. 4). With data from these graphs and the latitude of the mean centers (imbedded on the graph), users can, for the majority of the 24 species presented, be given a crude idea of the habitat tendencies based on their latitude of interest. For example, sugar maple has a mean center latitude of 42.1°N (Fig. 4). For someone interested in a location at 40°N (i.e., 42.1−40 = ~+ 2 difference in latitude), a straight line interpolation from the sugar maple graph on Fig. 4 would yield a future:current ratio of ~0.35, or roughly a loss of 65 % of habitat, and presumably the ecosystem services it provides, by 2100 under the GFDL scenario. In contrast, the habitat for black oak would roughly double at that same latitude (Fig. 4). Exceptions to the north to south trend were with loblolly pine, a large gainer in the MA relative to other regions, and red maple with much higher gains in CH, an artifact because this species is already prevalent in the other three regions) (Fig. 4). Landis300 versus GFDL showed similar trends except that northern red oak follows the trend of loblolly pine and tulip poplar follows the trend of red maple shown in Fig. 4 (Fig. S1).

The relationship of difference in latitude to future:current ratios, by region (red = NE; green = MA, pink = CH, blue = CA) for each of 24 species as forecast by TreeAtlas for 2100 under the GFDL climate scenario. Numbers in each species block indicate current mean latitude. Difference in latitude is calculated as (region mean center)—(species mean center), such that species mean centers to the north of region center have negative numbers
We compared the mean and ranked future:current change ratios for each species across all four models, all four regions, and under PCM, GFDL, or both scenarios (Table 4), and based on all output data (Table S3). Seven northern conifers were predicted to decline the most under climate change: black spruce, balsam fir, northern white cedar, red spruce, eastern hemlock, eastern white pine, and pitch pine. All had future:current ratios <0.8 so were categorized as small to large decreasers. Some species had a wide variation among models, including trends of opposite sign (future:current change ratio both <1 and >1) among models. Most striking is pitch pine, with Landis300 projecting a complete collapse in the NE and MA regions, while TreeAtlas and Linkages project small increases in NE. Sugar maple would only decrease slightly under PCM but lose substantial habitat, especially in the more southern CA and CH regions, under GFDL. Many of the hardwoods and southern pines show overall average increases in future:current ratios across regions and models (Tables 4, S3). However, some averages are skewed by very high ratios caused by very low current importance under one or more models (e.g., Virginia and loblolly pine), and there is a wide variation including substantial losses in some regions under some models.
Discussion
This comparative study, a first of its kind as far as we know, standardizes outputs from three unique forest landscape models across spatial (four regions) and temporal (2000, 2100, 2300) scales, a range of climate scenarios (PCM B1 and GFDL A1fi), species (30 total, most common species; region totals varied from 15 to 24), and evaluation metric (future:current ratios). Standardizing in this manner enables an increased understanding of underlying drivers of differences across models that use diverse approaches and assumptions. Divergence among models for certain species could indicate insufficient knowledge of the species’ life history traits or ecological complexities, a mismatch between current tree distributions and ecological drivers selected for modeling, or individual model biases. Divergence indicators provide fodder for further investigations into model behavior versus drivers. Agreement common to models could indicate a better understanding of species and their drivers, or could be indicative of a systemic bias among all models. The three models represent different approaches to assess potential species changes. TreeAtlas provides a robust statistical approach to model projected habitat, assuming the trees are where they are currently due to ecological and legacy conditions that will hold true into the future. Linkages also projects habitat as a hybrid empirical-physiological model working at the species level, and also feeding into Landis, which is a forest succession model estimating expected species importance through time.
Going into this study, we expected, based on what we know about the models, that (1) species with very low IV or biomass in the region will likely have less reliable models, and therefore less consistency among models; (2) Landis outputs will be dominated by current growth and succession of existing vegetation, and those legacies will persist; (3) migration does not occur in TreeAtlas or Linkages (only movement of suitable habitat), while migration within Landis will be extremely slow, even out 300 years; (4) the scenario with the least climate change, PCM, should also have the least predicted changes in IV or biomass; and (5) each of the models have a set of assumptions and limitations that need to be taken into account upon application (Iverson and McKenzie 2013).
Overall estimated impacts on species
With exceptions, there was overall agreement among models on species expected to do poorly under climate change in the future, especially under the hotter, drier GFDL scenario (see Table 1 for temperature and precipitation forecasts by region and scenario). The northern conifers of black spruce, balsam fir, northern white cedar, eastern hemlock, and red spruce are modeled to have large reductions in the future (Fig. 2). In the near term, however, red spruce has been shown to be expanding due to release from earlier human pressures (Foster and D’Amato 2015). This expanding trend may not continue for long because it favors cool, no-fire, and low spruce budworm conditions (Blum 1990), all conditions forecast to change to opposing trends in future. Historically, the southern margin of spruce and to some degree, fir, was likely limited by summer heat and drought, and they have only existed prominently in New England over the past 1000–1500 years, a time when July temperatures were perhaps 1 °C cooler than present (DeHayes et al. 2000). Thus, the warning of large future reductions for spruce and fir is warranted.
Sugar maple is a species modeled to stay roughly constant under the mild PCM scenario with all models, but likely to diminish substantially at the southern portions of its range under GFDL. In NE, it may stay relatively constant even under GFDL as it has some buffering because of its positive modification factors (Matthews et al. 2011). The red maple models show a large increase projected for the CH and more of a tendency to decline in the other three regions (Fig. 2), but it is the most adaptive species of all 134 species evaluated in TreeAtlas and should generally do fine under climate change (Matthews et al. 2011).
The oaks and hickories as a group generally show agreement across the modeling approaches, lending more support that, as a group, they may fare better under climate change, with the overall future:current averages for these species consistently above 1.0 (increase in importance or biomass) (Fig. 2, Table S3). They are often projected to have increased habitat or expanded ranges in the northern regions (NE and MA) but fare less well in the southern regions (CA and CH) where their southern limits are reduced within the region. With exceptions (e.g., northern red oak), this group of species is physiologically favored to withstand drought and heat (Johnson et al. 2009) and should be competitive under hotter and drier conditions of GFDL. However, succession of oak-dominated stands in the eastern United States is currently leading to ‘mesophication’ and dominance by maple (Nowacki and Abrams 2008), potentially due to lack of fire (Brose et al. 2014) and a more moist climate regime in the past century (Pederson et al. 2014). In much of the oak-hickory forests of the eastern United States, current management often relies upon increased fire and harvest to restore oak-hickory communities in locations otherwise trending towards maple (Hutchinson et al. 2005; Iverson et al. 2008a; Brose et al. 2013). A hotter climate may assist in enabling oak regeneration, but management to favor oak will likely still be needed because the highly competitive capabilities of maple (Abrams 1998; Matthews et al. 2011).
The pines generally have the most variable species projections among the models. The southern loblolly and Virginia pines have very low densities currently in these regions which cause less reliable future:current ratios for Landis and Linkages (the TreeAtlas models are built from entire eastern US distribution). Expansion by natural dispersal from low density populations is very slow within Landis. Pitch pine requires fire which is not directly included in the models, though may be indirectly included in the TreeAtlas model as a historic component of its current distribution. Eastern white pine generally shows a decreasing status in future.
Agreement among models
Several metrics were used to assess agreement among models. We hypothesized that TreeAtlas and Linkages, both projecting suitable habitat, should be more in agreement than with the Landis succession model outputs. All of our metrics appear to align with this hypothesis. A side-by-side visual comparison (Fig. 2), the agreement metric by species (Table 2) and region (Table S2), along with the Spearman rank coefficients (Fig. 3; Table S2), all point to the greatest overall agreement between the TreeAtlas and Linkages models. Therefore, for many species-region combinations with high agreement, uncertainty is lessened and confidence is built indicating probable tendencies of species behavior in coming decades. As in a similar study comparing Linkages and TreeAtlas for southern Missouri (Schneiderman et al. 2015), we found highest correlations under the scenario with the most change, GFDL (Spearman r = 0.77), compared to PCM (r = 0.60) for 83 species-region combinations. However, there was a large variation among regions: correlations were highest for 24 species simulated in NE (r = 0.91–0.96, P < 0.0001), followed by 24 species in the MA (r = 0.75–0.88, P < 0.01–0.0001), 20 species in the CH (r = 0.35–0.77, P = NS - < 0.05), and 15 species in the CA (r = 0.02–0.40, P = NS). We attribute much of the reduction in correlation to lower sample sizes in CH and CA. A further analysis of model sensitivity to species parameters would allow a more complete assessment of those species-region combinations showing disagreement between the two habitat models, but currently these disagreements underscore uncertainty for those combinations (Fig. 2; Table S3). Importantly, combinations of uncertainty between TreeAtlas and Linkages also indicate more uncertainty for Landis outputs because Linkages provide species establishment probabilities and maximum growing space inputs into Landis.
Some key differences between the two habitat models (TreeAtlas and Linkages) and Landis are attributed to the added components of succession and dispersal accounted for in Landis. For example, TreeAtlas and Linkages project large increases for loblolly pine habitat in CH and MA while Landis projects small to large decreases. This species currently has limited distributions in those regions and are constrained by natural dispersal. Of course, the future picture would change towards the habitat model projections should assisted migration of loblolly pine be implemented (Iverson and McKenzie 2013). In NE, TreeAtlas and Linkages predict large decreases in quaking aspen but Landis predicts increases because aspen, as an early successional species, fills growing space vacated by other northern species that are removed through harvest or are more severely affected by climate change. These differences illustrate the strength of a multi-model approach especially when the outcomes necessitate clear interpretability such as when being applied to regional vulnerability assessments. The importance of successional dynamics is a key ecological process that will regulate future forest transitions, yet the drivers of such change are dynamic and precise predictions are untenable. Therefore, by being able to evaluate side-by-side results originating from different approaches, one can begin to formulate quantitative and biologically informed assessments of likely outcomes, while clearly acknowledging potential sources of uncertainty.
We also hypothesized that Landis outcomes should move towards the habitat suitability outputs of TreeAtlas and Linkages as time moves from 100 to 300 years out. The metrics we used bore this out, but primarily with the harsh GFDL model (Figs. 2, 3; Tables S1, S2). Both the Spearman rank statistic, showing a large increase in correlations going from TreeAtlas and Linkages versus Landis100 to Landis300 (Fig. 3), and the class agreement scores, higher for TreeAtlas/Landis300 than for TreeAtlas/Landis100 (Table S1), showed these trends for GFDL but inconsistently so for PCM. Because Landis is a succession model, and a high proportion of established tree cohorts are likely to stay in place through 2100, most species will not see drastic changes by that time. However, most tree cohorts will be replaced by 2300 in Landis, by which time climate conditions (through Linkages) will dictate a greater similarity to the habitat suitability outcomes from Linkages and TreeAtlas. Landis simulations for the CH and CA indicated that the amount of variation in tree importance explained by succession and harvest decreased from 80 to 67 %, and by climate increased from 13 to 26 %, from 2100 to 2300 (Wang et al. 2015). Therefore, since TreeAtlas and Linkages did not consider succession and harvest, we expected agreement with Landis to increase as climate became more important over time and in the harsher GFDL scenario.
As for the higher correlations with the harsher, GFDL scenario, we suggest that the greater extremes in temperature and moisture, as analyzed through a daily database through 2100 (Matthews et al. in press), creates conditions likely to hasten succession and disturbance such that Landis300 will approximate changes in suitable habitat much faster than in the moderated climatic conditions projected through the PCM scenario. Notably, the large contrast between GFDL and PCM in this regard again points to the value of global society holding down emissions in coming decades (IPCC 2014).
The only disturbances we simulated in Landis were tree harvest and windthrow. In reality, additional disturbances from pests, pathogens, more frequent and hotter droughts, fires, floods, warm springs followed by late spring frosts, and wind damage are all expected to increase under climate change, and more so under the harsh scenario than the mild scenario (Dale et al. 2001; IPCC 2014; Melillo et al. 2014). Therefore, these disturbances will likely hasten transitions so that Landis300 along with TreeAtlas and Linkages projections of habitat may be closer to expectations by 2100. We emphasize the need to review species outputs, be they potential suitable habitat changes or succession models, in concert with other species and locality information, such as the TreeAtlas modification factors, so that disturbance and biological features of the species can be included into interpretations for management going forward (Iverson et al. 2011).
Relationships depending on geographic location
With these analyses, we confirm that the geographic position of the species range in comparison to the area of interest influences the potential for the species to thrive or not under a changing climate. Species that are already centered north of the region of interest are more likely to undergo a loss in habitat, as shown by the 16 species-region combinations classed as ‘North’ (species center is >2°N of region center) in Table 3; this pattern is consistent across all models and is much worse for the harsh GFDL scenario than the PCM scenario. Similarly, those species with mean centers >2°S of the region centers (in this case 30 species-region combinations averaging 4.2°S of region centers) had future:current ratios above 1 for all models and would be expected to increase in importance under climate change. For the 36 species-region combinations falling between 2°N and 2°S, the tendency is for a slight loss in importance according to all models, especially under GFDL, so the fate of these species may depend more on local conditions and specific attributes of the species.
Further confirmation of these geographic trends, by species, shows that, for TreeAtlas (Fig. 4) and Landis300 (Fig. S1), most species follow an expected north to south trend, with high future:current ratio/northerly latitude differences trending to low ratio/southerly latitude differences going from NE to MA to CA to CH. The exceptions to the pattern, such as red maple and loblolly pine for both TreeAtlas and Landis300, can be explained by a huge spatial niche becoming suitable in one of the regions (CH for red maple and MA for loblolly pine), creating a proportionately greater boost in those particular regions. The same can be said for some of the oaks and hickories in the Landis300 graphs. The geographic analysis presented here may therefore be a simple way for managers to get a perspective on tendencies for how species may fare in coming decades in their area of interest. By comparing the latitude of interest to the mean center latitude for the species, and then extrapolating to the points on species graphs of future:current ratio versus difference in latitude on Fig, 4, one can obtain an estimate of potential climatic pressures on the species by 2100 at that location. It is important to calculate mean centers based on both abundance and occupancy, not just presence so that a realistic center of species is generated. Low abundance and occupancy in expanding edges of the range may limit a species capacity to migrate (Iverson et al. 2004; Prasad et al. 2013), while low abundance at the trailing, or southern latitude edges, may result in range erosion and regional extirpation of suitable habitats. Examples of the former may be black oak in NE (i.e., rare propagules available to migrate into NE) while of the latter may be sugar maple in the CH (i.e., large loss of habitat in the species’ southern range) in this examination. Murphy et al. (2010), using TreeAtlas data, examined abundance–occupancy distributions across the geographic ranges of 102 eastern U.S. trees and found that 62 % of the species tended towards higher capacity to respond to climate change in the northern latitudes of their range but that most species were found to have range contraction in the south and limited range expansion in the north. Evaluating mean center positions of species importance (available on the TreeAtlas web site, cited in Methods) simultaneously with area of interest can thus provide a first look at potentials for species changes.
Informing management decisions
Our approach can help forest managers who are interested in adapting their management to current and future climate change (Janowiak et al. 2015). We have successfully incorporated the comparison of these models into climate change vulnerability assessments designed to support forest management decision-making (Brandt et al. 2014; Butler et al. 2015). The use of this information varies by individual management objectives, local site conditions, and differences in model projections by species and location. When there is greater agreement among models for a species of interest in a particular location, managers may choose to favor a species projected to do well or disfavor a species that is projected to do poorly in a manner than supports their land management objectives. When there is less agreement among models in the timing or direction of change, managers may choose alternative strategies such as favoring a greater diversity of species or choosing a mix of strategies in different areas (Swanston and Janowiak 2012). These areas of greater uncertainty can also provide an indication of when additional modeling, scientific evidence, ancillary information such as the modification factors, or just plain expertise gained from local, on-the-ground experience is needed to help inform decisions.
Conclusions
We evaluated three models and four model outcomes, for 30 common species and across four regions in the eastern United States, for agreement and enriched interpretation for forest management. Comparisons indicated high agreement for many species, especially northern species modeled to lose habitat in coming decades. TreeAtlas and Linkages outputs had the greatest agreement, but each had reasonable agreement with many species outputs from Landis, particularly when Landis was simulated to 2300. Most differences in projected change could be rationalized based on whether models considered realized versus fundamental niches, inclusion or not of succession, disturbance, and dispersal components, and the errors associated with incomplete knowledge and model constructs of ecological drivers, species parameters, and species interactions. Clearly there is still uncertainty imbedded in each of the models, such that any application stemming from them requires a thorough vetting through the lens of experts. A geographic analysis provided evidence that a simple calculation of mean center of species versus location of interest provides a basic idea on potential for the species under climate change. Of course the mantra “all models are wrong, but some are useful” (Box and Draper 1987) holds here but we suggest a unified analysis of multiple models provides a more consistent interpretation of the future for use in vulnerability assessments and adaptation planning.
References
Abrams MD (1998) The red maple paradox. BioScience 48:355–364
Allen CD, Breshears DD, McDowell NG (2015) On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6(8):1–55
Beckage B, Osborne B, Gavin DG, Pucko C, Siccama T, Perkins T (2008) A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc Natl Acad Sci USA 105(11):4197–4202
Blum B (1990) Picea rubens Sarg. Red Spruce. In: Burns RM, Honkala BH (eds) Silvics of North America, Conifers, vol 1. USDA Forest Service, Agriculture Handbook 654, Washington, DC, pp 250–259
Boisvert-Marsh L, Périé C, de Blois S (2014) Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 5(7):1–33
Box G, Draper NR (1987) Empirical model-building and response surfaces. Wiley, New York
Brandt L, He H, Iverson L, Thompson FR, Butler P, Handler S, Blume-Weaver R et al (2014) Central Hardwoods ecosystem vulnerability assessment and synthesis: a report from the Central Hardwoods climate change response framework project. U.S. Department of Agriculture, Forest Service, Northern Research Station, General technical report NRS-124, Newtown Square, PA
Brose PH, Dey DC, Phillips RJ, Waldrop TA (2013) A meta-analysis of the fire-oak hypothesis: does prescribed burning promote oak reproduction in Eastern North America? For Sci 59(3):322–334
Brose PH, Dey DC, Waldrop TA (2014) The fire-oak literature of Eastern North America: synthesis and guidelines. General technical report NRS-135. U.S. Department of Agriculture, Forest Service, Northern Research Station, Newtown Square, PA
Butler PR, Iverson L, Thompson FR, Brandt L, Handler S, Janowiak M, Connolly S et al (2015) Central Appalachians forest ecosystem vulnerability assessment and synthesis: a report from the Central Appalachians climate change response framework project. U.S. Department of Agriculture, Forest Service, Northern Research Station, General technical report NRS-146, Newtown Square, PA
Coumou D, Rahmstorf S (2012) A decade of weather extremes. Nat Clim Change 2:491–496
Dale VH, Joyce LA, McNulty S, Neilson RP, Ayres MP, Flannigan MD, Simberloff D (2001) Climate change and forest disturbances. BioScience 51(9):723–734
DeHayes DH, Jacobson GL, Schaberg PG, Bongarten B, Iverson L, Dieffenbacher-Krall AC (2000) Forest responses to changing climate: lessons from the past and uncertainty for the future. In: Mickler RA, Birdsey RA, Hom JL (eds) Responses of northern forests to environmental change. Ecological studies series. Springer, New York, pp 495–540
Delworth TL, Broccoli AJ, Rosati A, Stouffer RJ, Balaji V, Beesley JA, Durachta JW (2006) GFDL’s CM2 global coupled climate models. Part I: formulation and simulation characteristics. J Clim 19(5):643–674
Dijak W, Hanberry B, Fraser JS, He HS, Thompson III FR, Wang WJ (in press) Revision and application of the LINKAGES model to simulate forest growth in Central Hardwood landscapes in response to climate change. Landscape Ecol
Dobrowski SZ, Thorne JH, Greenberg JA, Safford HD, Mynsberge AR, Crimmins SM, Swanson AK (2011) Modeling plant ranges over 75 years of climate change in California, USA: temporal transferability and species traits. Ecol Monogr 81(2):241–257
Foster JR, D’Amato AW (2015) Montane forest ecotones moved downslope in Northeastern US in spite of warming between 1984 and 2011. Glob Change Biol 21:4497–4507
Franklin J (2010) Moving beyond static species distribution models in support of conservation biogeography. Divers Distrib 16(3):321–330
He HS, Hao Z, Mladenoff DJ, Shao G, Hu Y, Chang Y (2005) Simulating forest ecosystem response to climate warming incorporating spatial effects in northeastern China. J Biogeogr 32:2043–2056
Holm S (1979) A simple sequentially rejective multiple test procedure. Scand J Stat 6(2):65–70
Holzinger B, Hulber K, Camenisch M, Grabherr G (2008) Changes in plant species richness over the last century in the eastern Swiss Alps: elevational gradient, bedrock effects and migration rates. Plant Ecol 195(2):179–196
Hutchinson TF, Sutherland EK, Yaussy DA (2005) Effects of repeated fires on the structure, composition, and regeneration of mixed-oak forests in Ohio. For Ecol Manag 218:210–228
IPCC (2014) Climate change 2014: impacts, adaptation, and vulnerability. Working group II contribution to the IPCC 5th assessment report. Intergovernmental Panel on Climate Change, Stanford
Iverson L, McKenzie D (2013) Tree-species range shifts in a changing climate—detecting, modeling, assisting. Landscape Ecol 28:879–889
Iverson LR, Hutchinson TF, Prasad AM, Peters MP (2008a) Thinning, fire, and oak regeneration across a heterogeneous landscape in the eastern U.S.: 7-year results. For Ecol Manag 255(7):3035–3050
Iverson LR, Prasad AM, Matthews SN, Peters M (2008b) Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For Ecol Manag 254:390–406
Iverson LR , Prasad AM, Matthews SN , Peters M (2011) Lessons learned while integrating habitat, dispersal, disturbance, and life-history traits into species habitat models under climate change. Ecosystems 14:1005–1020
Iverson LR, Schwartz MW, Prasad A (2004) How fast and far might tree species migrate under climate change in the eastern United States? Glob Ecol Biogeogr 13:209–219
Janowiak MK, Swanston CW, Nagel LM, Brandt LA, Butler PR, Handler SD, Shannon PD, Iverson LR, Matthews SN, Prasad A, Peters MP (2014) A practical approach for translating climate change adaptation principles into forest management actions. J For 112:423–433
Johnson P, Shifley S, Rogers R (2009) The ecology and silviculture of oaks. CABI, New York
Keith DA, Akçakaya HR, Thuiller W, Midgley GF, Pearson RG, Phillips SJ, Rebelo TG (2008) Predicting extinction risks under climate change: coupling stochastic population models with dynamic bioclimatic habitat models. Biol Lett 4(5):560–563
Kennedy M, Ford E (2011) Using multi-criteria analysis of simulation models to understand complex biological systems. BioScience 61:994–1004
Landscape Change Research Group (2014) Climate change atlas. Northern Research Station, US Forest Service, Delaware. www.nrs.fs.fed.us/atlas
Lawler JJ, White D, Neilson RP, Blaustein AR (2006) Predicting climate-induced range shifts: model differences and model reliability. Glob Change Biol 12:1568–1584
Lenderink G, van Meijgaard E (2008) Increase in hourly precipitation extremes beyond expectations from temperature changes. Nat Geosci 1(8):511–514
Lenoir J, Gégout JC, Marquet PA, de Ruffray P, Brisse H (2008) A significant upward shift in plant species optimum elevation during the 20th century. Science 320(5884):1768–1771
Matthews SN, Iverson L, Peters M, Prasad AM (in press) Assessing potential climate change pressures throughout this century across the Conterminous United States: mapping plant hardiness zones, heat zones, and growing degree days. Northern Research Station Research map.
Matthews SN, Iverson LR, Prasad AM, Peters MP, Rodewald PG (2011) Modifying climate change habitat models using tree species-specific assessments of model uncertainty and life history factors. For Ecol Manag 262:1460–1472
McKenney DW, Pedlar JH, Rood RB, Price D (2011) Revisiting projected shifts in the climate envelopes of North American trees using updated general circulation models. Glob Change Biol 17(8):2720–2730
Melillo JM, Melillo TC, Richmond T, Yohe GW (2014) Climate change impacts in the United States: the third national climate assessment. U.S. Global Change Research Program, Washington, DC
Morin X, Thuiller W (2009) Comparing niche- and process-based models to reduce prediction uncertainty in species range shifts under climate change. Ecology 90:1301–1313
Murphy HT, VanDerWal J, Lovett-Doust J (2010) Signatures of range expansion and erosion in eastern North American trees. Ecol Lett 13(10):1233–1244
Nakicenovic N, Alcamo J, Davis G, Cambridgede Vries B, Fenhann J, Gaffin S (2000) IPCC special report on emissions scenarios. Cambridge University Press, Cambridge
Nowacki GJ, Abrams MD (2008) The demise of fire and “mesophication” of forests in the eastern United States. BioScience 58(2):123–138
Pearson RG, Thuiller W, Araújo MB, Martinez-Meyer E, Brotons L, McClean C, Lees DC (2006) Model-based uncertainty in species range prediction. J Biogeogr 33(10):1704–1711
Pederson N, D’Amato AW, Dyer JM, Foster DR, Goldblum D, Hart JL et al (2014) Climate remains an important driver of post-European vegetation change in the eastern United States. Glob Change Biol 2:2105–2110
Prasad AM, Gardiner J, Iverson L, Matthews S, Peters M (2013) Exploring tree species colonization potentials using a spatially explicit simulation model: implications for four oaks under climate change. Glob Change Biol 19(7):2196–2208
Prasad AM, Iverson LR, Liaw A (2006) Newer classification and regression tree techniques: bagging and random forests for ecological prediction. Ecosystems 9:181–199
Prasad AM, Iverson LR, Matthews SN, Peters MP (2016) A multistage decision support framework to guide tree species management under climate change via habitat suitability and colonization models, and a knowledge-based scoring system. Landscape Ecol. doi:10.1007/s10980-016-0369-7
Reich PB, Sendall KM, Rice K, Rich RL, Stefanski A, Hobbie SE, Montgomery RA (2015) Geographic range predicts photosynthetic and growth response to warming in co-occurring tree species. Nat Clim Change 5(2):148–152
Rohde R, Muller RA, Jacobsen R, Muller E, Perlmutter S, Rosenfeld A, Wurtele J, Groom D, Wickham C (2012) A new estimate of the average earth surface land temperature spanning 1753–2011. Geoinfor Geostat 1:1
Seminov V (2012) Arctic warming favours extremes. Nat Clim Change 2:315–316
Serra-Diaz JM, Franklin J, Ninyerola M, Davis FW, Syphard AD, Regan HM, Ikegami M (2014) Bioclimatic velocity: the pace of species exposure to climate change. Divers Distrib 20(2):169–180
Stoner AMK, Hayhoe K, Yang X, Wuebbles DJ (2013) An asynchronous regional regression model for statistical downscaling of daily climate variables. Int J Climatol 33(11):2473–2494
Swanston CW, Janowiak MK (2012) Forest adaptation resources: climate change tools and approaches for land managers. General technical report NRS-87. U.S. Department of Agriculture, Forest Service, Northern Research Station, Newtown Square, PA
Schneiderman J, He H, Thompson F III, Dijak W, Fraser J (2015) Comparison of a species distribution model and a process model from a hierarchical perspective to quantify effects of projected climate change on tree species. Landscape Ecol 30:1879–1892
Thuiller W, Albert C, Araújo MB, Berry PM, Cabeza M, Guisan A, Sykes MT (2008) Predicting global change impacts on plant species’ distributions: Future challenges. Perspect Plant Ecol 9(3–4):137–152
Tokarska KB, Gillett NP, Weaver AJ, Arora VK, Eby M (2016) The climate response to five trillion tonnes of carbon. Nat Clim Change. doi:10.1038/nclimate3036
Wang WJ, He HS, Fraser JS, Thompson FR III, Shifley SR, Spetich MA (2014a) LANDIS PRO: a landscape model that predicts forest composition and structure changes at regional scales. Ecography 37(3):225–229
Wang WJ, He HS, Spetich MA, Shifley SR, Thompson FR III (2014b) Evaluating forest landscape model predictions using empirical data and knowledge. Environ Model Softw 62:230–239
Wang WJ, He HS, Spetich MA, Shifley SR, Thompson FR, Larsen DR, Yang J (2013) A large-scale forest landscape model incorporating multi-scale processes and utilizing forest inventory data. Ecosphere 4(9):1–22
Wang W, He H, Thompson FR III, Fraser J, Hanberry B, Dijak W (2015) Importance of succession, harvest, and climate change in determining future composition in U.S. Central Hardwood Forests. Ecosphere 6(12):art277
Wang W, He HS, Thompson III FR, Fraser J, Dijak W (in press) Forest biomass and species distributions under climate change in the Northeastern U.S.: accounting for effects of succession and harvest. Landscape Ecol
Washington WM, Weatherly JW, Meehl GA, Semtner AJ Jr, Bettge TW, Craig AP, Zhang Y (2000) Parallel climate model (PCM) control and transient simulations. Clim Dyn 16:755–774
Webb T III, Bartlein PJ (1992) Global changes during the last 3 million years: climatic controls and biotic responses. Annu Rev Ecol Syst 23:141–173
Wiens JA, Stralberg D, Jongsomjit D, Howell CA, Snyder MA (2009) Niches, models, and climate change: assessing the assumptions and uncertainties. Proc Natl Acad Sci USA 106(Supplement 2):19729–19736
Woodall C, Conkling B, Amacher M, Coulston J, Jovan S, Perry C, Schulz B, Smith G, Wolf SW (2010) The forest inventory and analysis database. Version 4.0: database description and users manual for Phase 3. USDA Forest Service, Northern Research Station, Newtown Square
Woodall C, Oswalt CM, Westfall JA, Perry CH, Nelson MD, Finley AO (2009) An indicator of tree migration in forests of the Eastern United States. For Ecol Manag 257:1434–1444
Woodward FI, Williams BG (1987) Climate and plant distribution at global and local scales. Vegetatio 69:189–197
Wullschleger S, Gunderson C, Tharp ML, West D, Post W (2003) Simulated patterns of forest succession and productivity as a consequence of altered precipitation. In: Hanson P, Wullschleger S (eds) North American temperate deciduous forest responses to changing precipitation regimes. Ecological studies. Springer, New York, pp 433–446
Xu C, Gertner GZ, Scheller RM (2009) Uncertainties in the response of a forest landscape to global climatic change. Glob Change Biol 15(1):116–131
Acknowledgments
Many people are to be thanked, for we authors are dependent on data collected by others! Forest inventory data are paramount, as are historic and potential future climate data along with environmental data, all geographically referenced. We thank Eric Gustafson and journal reviewers for reviewing the manuscript. This project was funded by the USDA Forest Service Northern Research Station and Eastern Region, the United States Geological Survey Northeast Climate Science Center, and the University of Missouri-Columbia. Its contents are solely the responsibility of the authors and do not necessarily represent views of the funding agencies. This manuscript is submitted for publication with the understanding that the United States Government is authorized to reproduce and distribute reprints for governmental purposes.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Iverson, L.R., Thompson, F.R., Matthews, S. et al. Multi-model comparison on the effects of climate change on tree species in the eastern U.S.: results from an enhanced niche model and process-based ecosystem and landscape models. Landscape Ecol 32, 1327–1346 (2017). https://doi.org/10.1007/s10980-016-0404-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0404-8