
Abstract
Wide-spread fragmentation and isolation of habitats with high nature conservation value lends increasing importance to a better understanding of the factors which determine species richness in isolated habitat patches. Using data of one of the most abundant invertebrate groups in grasslands, Orthoptera, we analysed how species richness and distribution in 60 isolated semi-natural grassland remnants in Austria were affected by five environmental variables (altitude, habitat and land use diversity within each patch, habitat diversity of areas adjacent to each patch, patch size), and related to diversity of their main food source, i.e. vascular plants. We found a significant positive correlation between Orthoptera and vascular plant species richness, with threatened Orthoptera species having the lowest correlation coefficients. Life form diversity of plants was only moderately positively correlated with Orthoptera species richness. Habitat diversity within and adjacent to the grassland patch had by far the highest loadings on the first two axes of the principal component analysis, which jointly explained 99 % of the variance, and proved to be significant for total, threatened and not threatened Orthoptera, as well as for the two Orthoptera orders occurring in Central Europe (Caelifera, Ensifera). Additionally, the distribution of the majority of those 14 Orthoptera species analysed individually was mainly correlated with habitat diversity within and adjacent to the grassland patch. However, the distribution of a significant proportion of species was associated with other factors: five species were closely related to on-site land use diversity and patch size, and the distribution of three Ensifera species was not significantly correlated to any of the explanatory variables. We conclude that a surrogate taxa approach, i.e. the use of well-known taxonomic groups (e.g. vascular plants), may indeed deliver good results for capturing total, but less so for threatened, Orthoptera species richness in semi-natural grassland remnants. Small scale habitat diversity may be crucial to allow for the co-existence of a large number of Orthoptera species and has to be taken equally into account as patch size in nature conservation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Changes in land use practices associated with agricultural intensification have globally led to wide-spread fragmentation and isolation of habitats with high nature conservation value. This is particularly true for species-rich semi-natural grasslands in Central Europe, which harbour many rare and threatened species (Maurer et al. 2003; Zechmeister and Moser 2003; Zechmeister et al. 2007; Tscharntke et al. 2005; Marini et al. 2008). After significant—and still on-going—losses during the last decades (e.g. Settele et al. 1996; Braschler et al. 2009; Marini et al. 2010), only isolated remnants of semi-natural grasslands remain (e.g. Holzner 1986; Maurer et al. 2003).
Understanding the interplay of the determinants of local species richness and of individual species distribution in such remnant habitats is of prime importance for understanding and conserving remaining extant species diversity (e.g. Vessby et al. 2002; Sauberer et al. 2004; Tscharntke et al. 2005; Batary et al. 2007; Marini et al. 2008; Báldi 2008). Species richness in island-like habitat patches may be driven by two broad contrasting mechanisms (Kadmon and Allouche 2007; Kallimanis et al. 2008). Island biogeography theory suggests that area per se determines the number of species on an island as a result of an area-mediated equilibrium between extinction and colonisation rates (MacArthur and Wilson 1967; Rosenzweig 1995). In contrast, habitat diversity hypothesis and metapopulation theory predicts that habitat diversity and heterogeneity might be more important for affecting species richness, as more species may co-exist in a more diverse habitat patch of a given size as in a less diverse one (Rosenzweig 1995; Báldi 2008; Shi et al. 2010). Further, metapopulation theory suggests that the wider landscape context may influence local species diversity.
However, taxonomic groups may be differently correlated with indicators of site heterogeneity and landscape context which may result in divergent diversity patterns between taxonomic groups. So far, most studies have found significant cross-taxonomic correlations in species richness on the patch scale (e.g. Sauberer et al. 2004; Tscharntke et al. 2005; Batary et al. 2007; Marini et al. 2008; but see e.g. Vessby et al. 2002), albeit little research has focused on temperate grassland remnants. However, it remains insufficiently understood if and to which extent the distribution of individual species of a given taxonomic group follows the overall distribution pattern, and if and to which extent richness of declining and threatened species is correlated to overall species richness. For species conservation, a better understanding of these patterns and the underlying factors is crucial (Walter et al. 2004).
In temperate regions, Orthoptera species richness is highest in semi-natural grasslands. In Central Europe, 82 % of the Orthoptera species are associated with, and 60 % are completely restricted to, grasslands (Ingrisch and Köhler 1998). They constitute one of the most abundant invertebrate groups and are leading consumers of plant biomass, and thus play a key role in ecosystem processes (Ingrisch and Köhler 1998). Most Orthoptera have very specific habitat requirements regarding vegetation structure (Köhler 1999; Gardiner et al. 2002; Reinhardt et al. 2005) and microclimate (Ingrisch and Köhler 1998, Schirmel et al. 2010a). This specialization contributes to their high level of endangerment (Reinhardt et al. 2005), e.g. 57 % of all Austrian Orthoptera species are included in the national Red List (Berg et al. 2005). The ecology and distribution of Orthoptera in Central Europe is exceptionally well-studied, hence they are a key-group for monitoring and bioindication of grasslands (e.g. Ingrisch and Köhler 1998; Köhler 1999; Maas et al. 2002; Marini et al. 2008, 2010; Poniatowski and Fartmann 2010; Schirmel et al. 2010a).
The purpose of this paper is to elucidate how patch characteristics (area, altitude, habitat and land use diversity) as well as landscape context (adjacent habitat diversity) affect Orthoptera species richness in semi-natural dry grasslands in Central Europe and how their diversity is correlated to that of their main food source, i.e. vascular plants. We use data from 60 isolated grassland remnants to answer the following questions: (1) How is species richness of Orthoptera affected by species and functional diversity (life form strategies) of vascular plants? (2) How do patch size, altitude, within and adjacent habitat diversity and land use interact on Orthoptera species richness? (3) Is the richness of not threatened and threatened species controlled by the same set of factors? (4) Which factors control the distribution of individual Orthoptera species?
Materials and methods
Study region and study sites
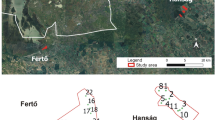
We have chosen a transect extending from the Northern Limestone Alps to the southern rim of the Bohemian massif (14°10′–50′ longitude, 47°50′–48°20′ latitude) in Northern Austria (Fig. 1). This transect extends 50 km in both the N–S and W–E directions. The climate is temperate, with cool winters and warm summers, with maximum precipitation in summer. Depending on the altitude, the annual medium temperature is between 7 and 9 °C (1961–1990). Annual precipitation increases from 700 to 900 mm in the Bohemian massif to 1,200–1,400 mm in the Limestone Alps (Auer et al. 1998). Bedrock at the study sites mainly consists of limestone or basic sediments (calcareous gravel, loess), and a few sites comprise neutral bedrock (e.g. flysch). The agricultural landscape of this area is dominated by fields and forests with interspersed villages and small towns. Semi-natural grassland remnants are mostly restricted to steep slopes and other marginal lands, where land use intensification is unfeasible.

Study area and location of the 60 dry grassland patches in northern Austria
Data
In the years 1999–2003 a total of 60 isolated dry grassland remnants have been investigated. Random selection of the sites was based on analyses of the Austrian inventory of dry grasslands (Holzner 1986), on results of the biotope mapping scheme of the federal province of Upper Austria and on our own data on distribution of dry grasslands across the study area (Essl unpublished). The altitude of the patches ranges between 260 and 690 m a.s.l, the size of the grassland remnants between 0.032 and 7 ha (Electronic supplementary material S1), and minimum distance between patches is 0.7 km.
Vegetation types were documented by 95 relevés for the entire study area to produce a conclusive list of vegetation types per grassland site. Prevailing vegetation types were mapped on all sites (Essl unpublished; vegetation classification follows Mucina et al. 1993). All study sites are dominated by semi-dry temperate grasslands, mostly belonging to the phytosociological association Onobrychido viciifoliae-Brometum. On moderately fertilized sites this community is replaced by the Ranunculi bulbosi-Arrhenatheretum. More rarely found were plant communities of thermophilic forest fringes (Trifolion medii), of nutrient-poor and acidic grasslands (Polygalo-Nardetum), open dry meadows (Saxifrago-Poetum), and moderately fertilized pastures (Festuco-Cynosuretum). We also mapped current land use on all sites. Most sites are mown once a year in early summer, a few sites are grazed by cattle. Due to changes in the agricultural system, land use has ceased altogether on a significant proportion of the sites in the last decades. For a more detailed description of the study sites see Essl (2000, 2002, 2004) and Essl and Weissmair (2002).
Each grassland site was visited four times during one vegetation period in order to sample a complete inventory of vascular plant species per site. Each plant species was assigned to a life form category: macro-phanerophytes (trees), nano-phanerophytes (shrubs), hemicryptophytes (long-lived herbs), geophytes (herbs with bulbs, tubers or rhizomes), and therophytes (annuals and biennals).
Recording of Orthoptera species was done during four visits between June and September in one of the observation years between 1999 and 2003. Sampling was hence extensive enough to fulfil standards for a near-complete inventory of Orthoptera species in small grassland remnants (e.g. Ingrisch and Köhler 1998). As we use presence/absence data of Orthoptera occurrence in the analyses, potential effects of population fluctuations are negligible. Recording of vascular plants and Orthoptera was done in the same year by the only observer (the first author of this paper). Total recording time was related to the size of the site, ranging from 1.5 h for sites <0.1 ha to 4 h for sites >1 ha for each of the two taxonomic groups. Orthoptera species were identified in the field based on morphological and acoustic characteristics. Orthoptera species were assigned to a threat category according to the Austrian Red List (Berg et al. 2005). Nomenclature of vascular plants follows Fischer et al. (2008), of Orthoptera follows Berg et al. (2005).
Five environmental variables were collected which were assumed to determine the variation in Orthoptera species richness. The grassland remnants have different sizes so that the log of size was used as a covariate in the analysis in order to control for the size effect on species richness. All habitat variables were standardized to the same size unit (ha). Habitat diversity of the sites was expressed as the number of mapped vegetation types. To analyse the effects of habitat diversity in the surrounding, the presence and absence of habitats adjacent to the patch (<100 m distance) was recorded for all grassland sites. Classification of adjacent habitats is based on a simplified version of the Austrian habitat type catalogue (Umweltbundesamt 2010). A land use diversity index was calculated as the inverse of the variance of the areas exposed to mowing, grazing, and fallow. The altitudes of the sites were expressed as the mean value of minimum and maximum altitude; due to the small size of most grassland patches, the within-patch elevational range was small.
Statistical analyses
For analyses, we calculated total Orthoptera species richness, as well as richness of threatened (i.e., species listed as CR, VU, EN, NT in the national Red List, Berg et al. 2005) and not threatened (i.e. species not included in the national Red list) Orthoptera species, as well as with species numbers of the two Orthoptera orders occurring in Central Europe, i.e. Caelifera (=grasshoppers, with short antennae), which mostly occur in low vegetation (e.g. grasslands, bare soil) and Ensifera (=crickets, bush crickets and allied species, with long antennae), which mostly prefer higher, vertically structured vegetation (e.g. bushes, fallow grassland, forest edges) (Ingrisch and Köhler 1998).
First we test how vascular plant species and life form diversity correlate with Orthoptera species richness. We calculated vascular plant life form diversity as the Shannon diversity index of the number of the respective species in each grassland patch. The Shannon index accounts for the degree of heterogeneity of different life forms (Magurran 1988). We used Pearson Product moment correlation with a two-sided test. In order to relate Orthoptera species richness to the above series of explanatory variables, we used generalised linear models (GLM) from the Poisson family with a log-link function. For total, not threatened and threatened Orthoptera species as well as for Caelifera and Ensifera separate models were fitted with the same predictor variables. Since these variables are correlated we used the first three axes (also known as components) of a principal component analysis (PCA) as explanatory variables, a preferred method when dealing with co-linearity in regression analysis (Quinn and Keough 2002). No scaling of variables has been performed. Response curves were visually inspected for curvilinear relationships. However, since no such responses were found only linear relationships were used. The importance of the three PCA axes in explaining the variance of the number of species was calculated by the change in deviance when dropping a variable from the GLM while all other variables remained in the model. The deviance is higher for variables responsible for a higher proportion of the variance of species richness. Significance was determined using the likelihood ratio statistic assuming a χ² test distribution.
We further analysed if the distribution of individual Orthoptera species is affected by the same set of explanatory variables. Again we used GLMs and followed the same procedure as described above albeit assuming a binomial error distribution for the presence of the respective species in a grassland patch. We selected those Orthoptera species which occur in medium abundance in the grassland patches (Electronic supplementary material S2), in total 14 species. We excluded abundant (>40 grassland patches colonized) and rare species (<10 grassland patches colonized) as model fitting for these species is not giving reliable enough results.
Statistical analyses were carried out in R, Version 2.13.1 (R Development Core Team 2011). We used the “prcomp” function for PCA and the “glm” function for GLM, which are both part of the “stats” package.
Results
Orthoptera species richness
We recorded 32 Orthoptera species (25 % of 127 Austrian species, Berg et al. 2005) (Electronic supplementary material S2) in the 60 dry grassland remnants, of which 16 were Ensifera respectively Caelifera species. Average species number per site was 11.2, with a minimum of 5 and a maximum of 17. Those Orthoptera species which are listed on the national Red List (11 taxa) have been recorded on average in 12.3 sites and hence are significantly more rare than not threatened species, which have been recorded in 26.1 sites on average.
The most wide-spread species were mesophilic grassland-species (Chorthippus parallelus, Ch. biguttulus, Ch. dorsatus, Metrioptera roeseli) and forest edge and shrub-species (Pholidoptera griseoaptera), all recorded in >50 sites. The most common species restricted to nutrient-poor grasslands were Gryllus campestris (50 sites), Stenobothrus lineatus (36 sites) and Leptophyes albovittata (34 sites).
Correlation of vascular plant and Orthoptera species richness
Overall 544 vascular plant species (18 % of 2,950 Austrian plant species, Fischer et al. 2008) have been found in the grassland sites (Essl and Dirnböck 2008). We found a significant correlation between Orthoptera and vascular plant species richness (Table 1). The more plant species occur in a site the more Orthoptera species are to be expected. This is true for total as well as for not threatened and threatened Orthoptera species and for Caelifera and Ensifera. Correlations are close for total and not threatened Orthoptera species, whereas threatened species and Caelifera have the lowest correlation coefficients with vascular plant species richness. Grouping vascular plants into plant functional types (i. e. life forms) resulted in much lower correlation coefficients, but again the correlation is exclusively positive. Only not threatened Orthoptera and Ensifera species have a significant correlation (P < 0.05) with plant life form diversity (Table 1).
Results of the PCA
The first axis of the PCA with the habitat and land use variables explained 88 % of the total variance, the second axis another 11 %, whereas the third axis explained only 0.3 %. The correlation (“loadings”) of the habitat and land use variables with these axes is shown in Table 2. Habitat diversity within and adjacent to the grassland patch have by far the highest loadings on the first two PCA axes. Since habitat diversity within and adjacent to the grassland patch are highly correlated the PCA is not capable at finding independent components. This means that the explanatory power of the two variables in the following GLM can only be interpreted jointly. The third PCA axis was most strongly related to land use diversity within the habitat patch and by patch size. Altitude had very low loadings on all PCA axes.
Factors determining Orthoptera species richness
All explored responses, i.e. total, not threatened and threatened Orthoptera species richness as well as richness of Caelifera and Ensifera, were significantly related with the first PCA axis (Table 2). This axis is determined by within and adjacent patch habitat diversity. PCA axis 2 is not significant but as mentioned this is an artefact of the high correlation of within and adjacent patch habitat diversity. Land use diversity and patch size, the determinants of PCA axis 3, only explain an insignificant fraction of the variance of Orthoptera species richness. The GLMs explain between 16 % (threatened species) and 61 % (all species) of the total variance of Orthoptera richness (Table 2).
Factors affecting individual Orthoptera species distributions
The distribution of the majority of the 14 Orthoptera species which occur in medium abundance is mainly associated with habitat diversity within and adjacent to the grassland patch, which are the two variables with the highest loadings on the PCA axes 1 and 2. In total, the distribution of eight of the 14 species is closely linked to axis 1 (Table 3). The distribution of five species, of which three are not related to the first two axes (Mecostethus parapleurus, Metrioptera bicolor, Tetrix tenuicornis) is closely related to axis 3, which is shaped by land use diversity within the site and patch size. The distribution of three Ensifera species (Leptophyes albovittata, Phaneroptera falcata, Tettigonia viridissima) is not significantly correlated to any of the three axes. In general, Ensifera species had lower correlations with the used explanatory variables than Caelifera species.
Discussion
Correlation of vascular plant and Orthoptera species richness
Our results show that species numbers of vascular plants are closely correlated with those of Orthoptera species in dry grassland remnants. This relation proved to be stable for total, not threatened, threatened Orthoptera species as well as for Caelifera and Ensifera species, being least close with threatened Orthoptera. Although still significant in some cases, life form diversity of vascular plants is much less correlated with Orthoptera species richness. These results suggest that Orthoptera richness in semi-natural grasslands is largely associated with the same set of environmental factors as the diversity of their main food source, vascular plants. These results corroborate most previous studies in grassland ecosystems which identified positive correlations between vascular plants and invertebrates in general (e.g. Panzer and Schwartz 1998; Collinge et al. 2003; Sauberer et al. 2004), although few of these studies have dealt with Orthoptera in particular (Sauberer et al. 2004; Marini et al. 2008), on the patch scale. In a study including eight taxonomic groups in an agricultural landscape in Eastern Austria, correlation of Orthoptera species richness with vascular plant species number was the highest compared with other taxonomic groups (pearson correlation coefficient 0.77, Sauberer et al. 2004). The correlation in our study is even stronger suggesting that Orthoptera and vascular plant species richness is particularly closely related in dry grasslands.
Interestingly, the correlation with vascular plants is weakest for the richness of threatened Orthoptera species, an indication that the occurrence of specialized and declining Orthoptera species is only partly captured by the diversity of vascular plants (see e.g. Reinhardt et al. 2005). Several previous studies have identified factors which might account for differential responses to environmental factors between vascular plants and Orthoptera species, and these differences seem to be of particular relevance for threatened Orthoptera species. These factors include the wider landscape composition, which influences the distribution of Orthoptera species in alpine grasslands, but not that of vascular plant species (Marini et al. 2008). In contrast to vascular plants, site conditions such as the pH value of the bedrock or soil depth are of little direct importance for Orthoptera. Life cycle differences—most vascular plant grassland species are long-lived (many years to decades) whereas Orthoptera have an annual life cycle—may lead to different relaxation times to habitat fragmentation or degradation (Krauss et al. 2010), as long-lived species may persist longer under deteriorating conditions (Öckinger et al. 2010; Sang et al. 2010).
We conclude that a surrogate taxa approach, i.e. the use of well-known taxonomic groups (e.g. vascular plants) to capture overall species richness (Lewandowski et al. 2010), and which has proven to be applicable in many cases on the habitat scale (e.g. Panzer and Schwartz 1998; Sauberer et al. 2004; Kruess and Tscharntke 2002; but see Vessby et al. 2002 for contrasting results), may indeed deliver good results for capturing total, but less so for threatened, Orthoptera species richness in semi-natural grassland remnants.
Importance of patch size, land use and environmental factors on Orthoptera richness
Habitat diversity within and adjacent to the grassland patches were the principal factors influencing total, not threatened and threatened Orthoptera as well as Caelifera and Ensifera species richness. The presence of a wide range of habitats including open dry pioneer grasslands, semi-dry grasslands, shrubs, and unmown forest fringe vegetation allows Orthoptera species with contrasting habitat requirements to co-exist.
Land use diversity within the grassland patches did only marginally influence Orthoptera species richness, although the prevailing type (grazing, mowing, fallow) and intensity (e.g. one or two annual cuts) affects sward structure and height (Gardiner et al. 2002; Kruess and Tscharntke 2002; Batary et al. 2007; Marini et al. 2009), and may cause high mortality of larvae and adults (Gardiner and Hill 2006). A higher diversity of land uses types on a grassland patch may also offer temporal refuges for mobile Orthoptera, as most Orthoptera species—with the notable exception of geophilic species (e.g. Oedipoda coerulescens, Tetrix spp.)—avoid very short freshly mown or grazed swards, to reduce predation risks due to missing shelter. We assume that due to the predominance of habitat diversity in our study, land use, which is moreover related to habitat diversity, had too little additional, independent explanatory power to offer.
The altitudinal position of the grassland sites had very little influence on Orthoptera species richness. We argue that this result can be explained by the rather small altitudinal range (260–690 m a.s.l) covered by the study sites. In Central Europe, Orthoptera species richness in grasslands decrease strongly in cool climates prevalent in medium to high altitudes (Ingrisch and Köhler 1998; Wettstein and Schmid 1999), however we have not sampled the upper elevation belts.
Biogeography theory suggests that patch size might be an important factor in controlling species richness (MacArthur and Wilson 1967; Rosenzweig 1995). However, we found that patch sizes provided only a small contribution to explain Orthoptera species richness. This is somewhat surprising as our case study included many very small grassland remnants (0.032–7.0 ha), which were just above or well below minimum area sizes for viable populations of most Central European Orthoptera confined to species-rich grasslands (Ingrisch and Köhler 1998; Köhler 1999). Edge effects, which are known to have strong negative effect on Orthoptera species richness, may further reduce the carrying capacity. Orthoptera species richness in dry grassland in eastern Austria is strongly reduced by adjacent forests, which reduce irradiance and soil temperature (Bieringer and Zulka 2003). As our patches are isolated by intensively used agricultural landscapes, rapid re-colonisation is unlikely in most cases (Poniatowski and Fartmann 2010), although Orthoptera with higher dispersal capacity may be capable to colonize isolated small dry grassland remnants (Marini et al. 2010). Generalist Orthoptera species may be able to co-occur in intensively used grasslands in the wider landscape, so isolation of patches may well be less pronounced for these species. Possibly, the size effect may also be masked by delayed local extinctions (Krauss et al. 2010). A significant proportion of habitat loss of dry grasslands in Austria in general (BMLFUW 2011) and in the study area (Essl unpublished) happened quite recently (>1985), so we argue that even without further habitat loss future losses of Orthoptera species due to stochasticity and population dynamics on the small grassland patches might be expected (Ingrisch and Köhler 1998), a process which may however take several decades to happen (Krauss et al. 2010; Sang et al. 2010).
Distribution of individual Orthoptera species
As for total Orthoptera species richness, habitat diversity within and adjacent to the grassland patches were best correlated with the distribution of the majority of the 14 Orthoptera species of medium abundance, whose distribution we analysed individually. However, this pattern is not uniform across these species. The distribution of three Ensifera species is not related to any of the three axes of the PCA, and the occurrence of three species is only related to the third axis, which is dominated by land use diversity within the patch and patch size.
Differential factors controlling the distribution of individual Orthoptera species makes setting conservation priorities delicate (Tscharntke et al. 2005; Marini et al. 2010), as conservation measures targeted to individual species may be best achieved by setting other, or even opposite, management priorities as to optimizing total species richness (Walter et al. 2004).
Implications for nature conservation
Given limited taxonomic expertise and financial resources, the assessment of taxonomic diversity for appraising the conservation value of habitat patches or of nature reserves is often restricted to one or few taxonomic groups, in particular vascular plants (Sauberer et al. 2004; Lewandowski et al. 2010). Our study shows, that in Central European grassland remnants vascular plant species richness is indeed a reasonable surrogate for Orthoptera species richness, although less so for threatened Orthoptera species.
Habitat diversity within and adjacent to the grassland patches proved to be the most important predictors of Orthoptera species richness, whereas the contribution of patch size was minor. Our results thus suggest that habitat diversity on small scales may be crucial to allow for the co-existence of a large number of Orthoptera species. Hence, habitat heterogeneity and diversity has to be taken equally into account as size in nature conservation (Báldi 2008; Kallimanis et al. 2008: Schirmel et al. 2010b). In order to safeguard the whole species assemblage conserving and re-creating heterogenous habitats (e.g. Knop et al. 2006) as may be achieved by well-designed agri-environmental schemes (Kleijn et al. 2006), may be the optimal conservation strategy. Unfortunately, recent trends in land use and habitat configuration in Europe indicate decreasing habitat diversity on the patch scale driven mostly by increasing mechanization of land use, increasing field parcel sizes and abandonment of traditional land use systems (Zechmeister et al. 2003; Moser et al. 2004; BMLFUW 2011).
Abbreviations
- GLM:
-
General linear model
- PCA:
-
Principal component analysis
References
Auer I, Böhm R, Dobesch H, Hammer N, Koch E et al (1998) Klimatographie und Klimaatlas von Oberösterreich. Beiträge zur Landeskunde Oberösterreich, II. naturwiss. Reihe 2 + 3
Báldi A (2008) Habitat heterogeneity overrides the species–area relationship. J Biogeogr 35:675–681
Batary P, Orci KM, Baldi A, Kleijn D, Kisbenedek T, Erdös S (2007) Effects of local and landscape scale and cattle grazing intensity on Orthoptera assemblages of Hungarian Great Plain. Basic Appl Ecol 8:280–290
Berg HM, Bieringer G, Zechner L (2005) Rote Liste der Heuschrecken (Orthoptera) Österreichs. In. Zulka K (Red.). Rote Listen gefährdeter Tiere Österreichs. Grüne Reihe 14/1: 167–211
Bieringer G, Zulka KP (2003) Shading out species richness. Edge effect of a pine plantation on the Orthoptera (Tettigoniidae and Acrididae) assemblage of an adjacent dry grassland. Biodiv Conserv 12:1481–1495
BMLFUW (2011) Grüner Bericht 2011. http://www.gruenbericht.at. Accessed 06 Oct 2011
Braschler B, Marini L, Thommen GH, Baur B (2009) Effects of small-scale grassland fragmentation and frequent mowing on population density and species diversity of Orthopterans—a long-term study. Ecol Entomol 34:321–329
Collinge SK, Prudic KL, Oliver JC (2003) Effects of local habitat quality characteristics and landscape context on grassland butterfly diversity. Conserv Biol 17:178–187
Essl F (2000) Botanische (Flora, Vegetation) und zoologische Aspekte (Heuschrecken, Reptilien) der Halbtrockenrasen am Westabfall des Kürnberger Waldes (Oberösterreich). Naturkundliches Jahrbuch der Stadt Linz 45:135–167
Essl F (2002) Flora, Vegetation und zoologische Untersuchungen (Heuschrecken und Reptilien) der Halbtrockenrasen im Ostteil der Traun-Enns-Platte (Oberösterreich). Naturkundliches Jahrbuch der Stadt Linz 48:193–244
Essl F (2004) Flora, Vegetation und zoologische Untersuchungen (Heuschrecken und Reptilien) ausgewählter Halbtrockenrasen der Ennstaler Flysch- und Kalkvoralpen (Oberösterreich). Naturkundliches Jahrbuch der Stadt Linz 50:11–58
Essl F, Dirnböck T (2008) Diversity of native and alien vascular plants of dry grasslands in Central Europe. Appl Veget Science 11:441–451
Essl F, Weissmair W (2002) Flora, Vegetation und zoologische Untersuchungen (Heuschrecken und Reptilien) der Halbtrockenrasen am Südrand der Böhmischen Masse östlich von Linz (Oberösterreich). Beitr Naturk Oberösterreichs 11:267–320
Fischer MA, Adler W, Oswald K (2008) Exkursionsflora für Österreich, Liechtenstein und Südtirol. Biologiezentrum Oberösterreich, Linz
Gardiner T, Hill J (2006) Mortality of Orthoptera caused by mechanical mowing of grassland. British J Entomol Nat Hist 19:38–40
Gardiner T, Pye M, Field R, Hill J (2002) The influence of sward height and vegetation composition in determining the habitat preferences of three Chorthippus species (Orthoptera: Acrididae) in Chemlsford, Essex, UK. J Orthoptera Res 11:207–213
Holzner W (ed) (1986) Österreichischer Trockenrasenkatalog. Grüne Reihe des Bundesministeriums für Gesundheit und Umweltschutz, Wien
Ingrisch S, Köhler G (1998) Die Heuschrecken Mitteleuropas. Westarp Wissenschaften, Magdeburg
Kadmon R, Allouche O (2007) Integrating the effects of area, isolation, and habitat heterogeneity on species diversity: a unification of island biogeography and niche theory. Amer Naturalist 170:443–454
Kallimanis AS, Mazaris AD, Tzanopoulos J, Halley JM, Pantis JD, Sgardelis SP (2008) How does habitat diversity affect the species—area relationship? Glob Ecol Biogeogr 17:532–538
Kleijn B, Baquero RA, Clough Y, Diaz M, De Esteban J et al (2006) Mixed biodiversity benefits of agri-environmental schemes in five European countries. Ecol Lett 9:243–254
Knop E, Kleijn D, Herzog F, Schmid B (2006) Effectiveness of the Swiss agri-environmental scheme to promote biodiversity. J Appl Ecol 43:120–127
Köhler G (1999) Ökologische Grundlagen von Aussterbeprozessen. Fallstudien an Heuschrecken. Laurenti, Jena
Krauss J, Bommarco R, Guardiola M, Heikkinen RK, Helm A et al (2010) Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels. Ecol Lett 13:597–605
Kruess A, Tscharntke T (2002) Grazing intensity and the diversity of grasshoppers, butterflies and trap-nesting bees and wasps. Conserv Biol 16:1570–1580
Lewandowski AS, Noss RF, Parsons DR (2010) The effectiveness of surrogate taxa for the representation of biodiversity. Conserv Biol 24:1367–1377
Maas S, Detzel P, Staudt A (2002) Gefährdungsanalyse der Heuschrecken Deutschlands. Bundesamt für Naturschutz, Bonn
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Magurran AE (1988) Ecological diversity and its measurement. Cambridge University Press,Cambridge
Marini L, Fontana P, Scotton M, Klimek S (2008) Vascular plant and Orthoptera diversity in relation to grassland management and landscape composition in the European Alps. J Appl Ecol 45:361–370
Marini L, Fontana P, Battisti A, Gaston KJ (2009) Response of orthopteran diversity to abandonment of semi-natural meadows. Agric Ecosyst Environ 132:232–236
Marini L, Bommarco R, Fontana P, Battisti A (2010) Disentangling effects of habitat diversity and area on orthopteran species with contrasting mobility. Biol Conserv 143:2164–2171
Maurer K, Durka W, Stöcklin J (2003) Frequency of plant species in remnants of calcareous grassland and their dispersal and persistence characteristics. Basic Appl Ecol 4:307–316
Moser D, Zechmeister HG, Plutzar C, Sauberer N, Wrbka T, Grabherr G (2004) Landscape patch shape complexity as an effective measure for plant species richness in rural landscapes. Landscape Ecol 17:657–669
Mucina L, Grabherr G, Ellmauer T, Wallnöfer S (1993) Die Pflanzengesellschaften Österreichs. G. Fischer, Stuttgart
Öckinger E, Schweiger O, Crist TO, Debinski DM, Krauss J et al (2010) Life history traits predict species responses to habitat area and isolation—a cross-continental synthesis. Ecol Lett 13:969–979
Panzer R, Schwartz MW (1998) Effectiveness of a vegetation-based approach to insect conservation. Conserv Biol 12:693–702
Poniatowski D, Fartmann T (2010) What determines the distribution of a flightless bush-cricket (Metrioptera brachyptera) in a fragmented landscape? J Insect Conserv 14:637–645
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge University Press, Cambridge
Reinhardt K, Köhler G, Maas S, Detzel P (2005) Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: the Orthoptera of Germany. Ecography 28:593–602
Rosenzweig ML (1995) Species diversity in space and Time. Cambridge University Press, Cambridge
Sang A, Teder T, Helm A, Pärtel M (2010) Indirect evidence for an extinction debt of grassland butterflies half century after habitat loss. Biol Conserv 143:1405–1413
Sauberer N, Zulka KP, Abensperg-Traun M, Berg HM, Bieringer G et al (2004) Surrogate taxa for biodiversity in agricultural landscapes of eastern Austria. Biol Conserv 117:181–190
Schirmel J, Mantilla-Contreras J, Blindow I, Fartmann T (2010a) Impact of succession and grass encroachment on heathland Orthoptera. J Insect Conserv 15:633–642
Schirmel J, Blindow I, Fartmann T (2010b) The importance of habitat mosaics for Orthoptera (Caelifera and Ensifera) in dry heathlands. Eur. J. Entomol. 107:129–132
Settele J, Margules C, Poschlod P, Henle K (1996) Species survival in fragmented landscapes. Kluwer, Dordrecht
Shi JM, Ma KM, Wang JF, Zhao JZ, He K (2010) Vascular plant species richness on wetland remnants is determined by area and habitat heterogeneity. Biol Conserv 19:1279–1295
R Development Core Team (2011) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, http://www.R-project.org
Tscharntke T, Kleijn AM, Kruess A, Steffan-Dewenter I, Thiess C (2005) Landscape perspectives on agricultural intensification and biodiversity: ecosystem service management. Ecol Lett 8:857–874
Umweltbundesamt (2010) Naturschutzfachliches Informationssystem Austria (NISA). http://gis.umweltbundesamt.at/austria/natur/nisa/Default.faces. Accessed 14 Aug 2010
Vessby K, Söderström B, Glimskär A, Svensson B (2002) Species-richness of six different taxa in Swedish seminatural grasslands. Conserv Biol 16:430–439
Walter T, Hunziker M, Peter B, Ward F (2004) Threatened grasshopper species profit from ecological compensation areas. Grassland Sciences in Europe 9:234–236
Wettstein W, Schmid B (1999) Conservation of arthropod diversity in montane wetlands: effect of altitude, habitat quality and habitat fragmentation on butterflies and grasshoppers. J Appl Ecol 36:363–373
Zechmeister HG, Moser D (2003) The influence of agricultural land use intensity on bryophyte species richness. Biodiv Conserv 10:1609–1625
Zechmeister HG, Schmitzberger I, Steurer B, Peterseil J, Wrbka T (2003) The influence of land use practices and economics on plant species richness in meadows. Biol Conserv 114:165–177
Zechmeister HG, Moser D, Milasowszky N (2007) Spatial distribution patterns of Rhynchostegium megapolitanum at the landscape scale—an expanding species? J Appl Veget Science 10:111–120
Acknowledgments
We are obliged to one anonymous reviewer and to W. Rabitsch for comments on a previous version of the paper.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Essl, F., Dirnböck, T. What determines Orthoptera species distribution and richness in temperate semi-natural dry grassland remnants?. Biodivers Conserv 21, 2525–2537 (2012). https://doi.org/10.1007/s10531-012-0315-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-012-0315-1