
Abstract
This study tested whether multiple domains of social adversity, including neighborhood opportunity/deprivation and life stress, moderate genetic (A), common environmental (C), and unique environmental (E) influences on externalizing behaviors in 760 same-sex twin pairs (332 monozygotic; 428 dizygotic) ages 10–11 from the ABCD Study. Proportion of C influences on externalizing behavior increased at higher neighborhood adversity (lower overall opportunity). A decreased and C and E increased at lower levels of educational opportunity. A increased at lower health-environment and social-economic opportunity levels. For life stress, A decreased and E increased with number of experienced events. Results for educational opportunity and stressful life experiences suggest a bioecological gene-environment interaction pattern such that environmental influences predominate at higher levels of adversity, whereas limited access to healthcare, housing, and employment stability may potentiate genetic liability for externalizing behavior via a diathesis-stress mechanism. More detailed operationalization of social adversity in gene-environment interaction studies is needed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Externalizing behavior in childhood, including proactive aggression, reactive aggression, and irritability/anger dysregulation, has been linked to negative health sequelae, such as increased risk for a myriad of mental health problems (Loth et al. 2014; Bornstein et al. 2013; Molina et al. 2018), and disruption in life opportunities, such as educational and occupational achievement during adulthood (Fairchild et al. 2019; Dardani et al. 2021; Ramos-Olazagasti et al. 2018; Jangmo et al. 2021; Lewis et al. 2017). Twin studies suggest that externalizing behavior is moderately to highly heritable (~ 35-65%), with some additive influence attributable to both common and unique environmental factors; gene-by-environment interaction (GxE) effects related to exogenous environmental exposures are also etiologically important (Hatoum et al. 2018; Barr and Dick 2019; Pearson et al. 2022; Burt 2022). GxE as presently operationalized refers to a statistical (not biological) dependence of genetic expression on environmental factors and/or the statistical dependence of environmental effects on genetic characteristics (Dick 2011).
Experiences of social adversity may escalate long-term risk for negative outcomes among youth at risk for externalizing behavior. Social adversity includes factors such as neighborhood disadvantage (e.g., poverty; lack of access to health and educational resources) and life stressors (e.g., parental incarceration, injury, and/or death; witnessing or experiencing violence) (Gartland et al. 2019). Several neighborhood-level facets of social adversity have been associated with externalizing behavior, including structural neighborhood factors (e.g., agricultural land use; alcohol outlet density), environmental toxicants (Pearson et al. 2022), limited exposure to greenspace (Sakhvidi et al. 2022), economic stress, neighborhood poverty (Burt 2022; Wickham et al. 2017; Devenish et al. 2017; Maxwell et al. 2022), and school classroom quality (Hoglund et al. 2015; Abry et al. 2017). Youth living in marginalized communities and/or low-resource neighborhoods are disproportionately exposed to negative and stressful life events (Reiss 2013; Reiss et al. 2019; Choi et al. 2021), which are also associated with externalizing behavior (March-Llanes et al. 2017; Womack et al. 2022; McNeilly et al. 2021; Turney 2022).
While the phenotypic associations between adversity and externalizing behavior are well-characterized, there are competing conceptualizations of how GxE effects may operate and, in turn, inconsistent findings regarding how facets of social adversity relate to the etiology of externalizing behavior (Burt 2015, 2022). Part of this inconsistency may stem from differences in how proximal versus distal aspects of social adversity (e.g., stressful life events versus neighborhood disadvantage) moderate genetic influences on externalizing behaviors (Holz et al. 2018). Specifically, it has been posited that more proximal factors (e.g., experienced stressors) moderate genetic influences on externalizing behavior via a diathesis-stress process, while more distal factors (e.g., neighborhood deprivation) moderate genetic influences on externalizing behavior via a bioecological “social push” process (Holz et al. 2018).
The diathesis-stress model of psychopathology (Monroe and Simons 1991) posits that genetic liability related to externalizing behavior can be potentiated by experience of environmental stressors among youth. Consistent with this, several studies have shown that genetic influences on externalizing behavior are enhanced at higher levels of adversity, particularly higher levels of life stress (Fairchild et al. 2019; Samek and Hicks 2014; Hicks et al. 2009). For example, one biometric analysis of twin data found that genetic influences on externalizing behavior were stronger among 17-year-old twins who experienced more stressful life events, while shared environmental influences were stronger among those who experienced fewer stressful life events (Hicks et al. 2009). Similar patterns have also been observed in candidate gene-by-environment interaction studies, wherein higher levels of life stress were associated with externalizing behavior among youth carrying the 7-repeat allele of the dopamine D4 receptor (DRD4) gene (Zandstra et al. 2018), and childhood maltreatment was associated with genetic risk for antisocial behavior among youth carrying the low-activity allele of the MAOA gene (Assary et al. 2018; Caspi et al. 2002). However, it should be noted that candidate gene studies such as these often fail to replicate (Border et al. 2019; Farrell et al. 2015). While there is some evidence for polygenic score by proximal environment interactions on externalizing behavior outcomes that support a diathesis-stress model, this approach typically does not explain substantial variance beyond main effects of genes and environment (Plomin et al. 2022).
Conversely, the bioecological model posits that environmental influences become more potent in the context of greater social adversity (Raine 2002) or the converse, that when adversity is low and social environments are “good-enough,” then individual differences are more likely to be related to genetic variation. There has been support for this theory in the context of GxE effects of neighborhood disadvantage and related contextual factors (Burt 2022; Burt et al. 2019). A twin study among 4- to 5-year-olds found that genetic influences on externalizing behaviors were less robust at lower levels of socioeconomic status (SES) (Cheung et al. 2014). Similarly, a study of 16- to 17-year-old twins similarly found that heritability of antisocial behavior was higher among youth residing in more advantaged neighborhoods, while shared environmental influences were stronger among youth residing in more disadvantaged neighborhoods (Tuvblad et al. 2006). Longitudinal findings from the Add Health study showed that heritability of externalizing behavior was higher among youth who experienced fewer adverse childhood events, as indexed by an aggregate measure of individual-level stressors and neighborhood disadvantage (Wright and Schwartz 2021). Such patterns have also been replicated in several studies of 6- to 10-year-old and 3- to 17-year-old twin samples (Burt 2022; Burt et al. 2016, 2020).
There is substantial nuance in the presentation of GxE effects on externalizing behavior. A study in twin pairs aged 6 to 15 years from the Australian Twin-Study of the National Assessment Program- Literacy and Numeracy did not find any evidence for GxE effects on externalizing behavior (Gould et al. 2018). Other studies have found differential support for the diathesis-stress versus bioecological patterns of effect across development, with the former appearing in adolescence and the latter appearing in childhood (Burt and Klump 2014), and yet others have found different GxE patterns across disparate facets of externalizing behavior (e.g., ADHD versus callous-unemotional traits; aggression versus rule-breaking) (Burt 2022; Burt et al. 2016; Carroll et al. 2022). Taken together, results are inconsistent in terms of the presence of GxE effects and the circumstances under which genes versus environment may exert the most influence.
Also complicating this picture is that the construct of social adversity in GxE studies is often narrowly- or under-defined. While we know that both proximal and distal aspects of social adversity operate as social determinants of health across the lifespan (Viner et al. 2012), these constructs are multidimensional and there are increasing calls for attention to be paid to the nuance of educational, economic, and health-related aspects of social adversity. Such lack of specificity in how adversity, including neighborhood disadvantage and life stress, is defined may contribute to inconsistencies in the GxE literature if different facets of adversity display different patterns of GxE effects. For example, recent conceptualizations of neighborhood-level influences on youth development have highlighted the importance of several specific domains, including those of educational, health and environmental, and social and economic opportunity (Pearson et al. 2022; Acevedo-Garcia et al. 2014). Because neighborhood opportunity and life stress are multidimensional constructs reflecting overall resources available within a given community and subjectively valanced experiences, it is important to identify how the critical elements within a larger, more comprehensive construct of adversity may differentially moderate genetic and environmental influences on youth externalizing behaviors.
Clarifying GxE effects using clearly defined and differentiated aspects of social adversity is salient given that environments can be modified to some extent and can thus be leveraged in family- and school-based interventions such as Multisystemic Therapy (Henggeler et al. 1986) and the Family Check-Up intervention (Dishion and Kavanagh 2003; Hentges et al. 2020). In that context, stratification by genetic risk can help identify risk groups who may benefit from prevention policy. Further, genotype may moderate externalizing intervention efficacy within interventions that leverage both family- and school-based components (e.g., Albert et al. 2015), which can aid the shift toward personalized prevention/intervention efforts. Better understanding such dynamics under different contexts can help point to optimal opportunities for intervention or treatment-matching. This is also relevant for public and school policy, juvenile/criminal justice decision making, and issues of genetic discrimination. For example, if aggression and other externalizing behaviors are erroneously viewed by policymakers and criminal justice authorities as unmalleable due to heritable influences, GxE studies can aid in informing and clarifying the critical malleability of risk and outcome. This can prevent poorly-conceived policies that inadvertently exacerbate health disparities.
Present Study
The present study aimed to test whether multidimensionally measured neighborhood opportunity and stressful life events moderate genetic and environmental influences on externalizing behaviors within a twin sample derived from the Adolescent Brain Cognitive Development (ABCD) Study, a 10-year longitudinal study of child behavioral, brain development, and health outcomes conducted across 21 research sites involving nearly 12,000 youth in the United States (Jernigan et al., 2018). Four “Twin Hub” sites in Minnesota, Colorado, Virginia, and Missouri conducted targeted recruitment of complete, same-sex twin pairs with a target of 200 pairs per site (Iacono et al. 2017). The advantage of this data source is the wide representativeness of the sample (although this was a volunteer study so some volunteer biases can occur, as noted in the discussion). Analyses focused on how GxE effects may differ across different domains related to neighborhood opportunity, including education, health-environment, and social-economic, as well as experiences of life stress as cumulative experiences or experiences subjectively experienced as “bad.” For all domains, we tested competing predictions from diathesis-stress and bioecological models of psychopathology that posit either increasing (diathesis-stress) or decreasing (bioecological) genetic effects in the context of higher levels of social adversity.
Methods
Participants & Procedure
Participants were drawn from the ABCD Study twin sample. The analytic sample was comprised of 760 complete same-sex twin pairs from the ABCD Study (n = 332 monozygotic [MZ] pairs, n = 428 dizygotic [DZ] pairs). Zygosity was determined via saliva and blood samples. The sample included 49% cisgender girls, was 82% white; 10% Hispanic/Latinx; and had a mean age of 10.12 years old at baseline (SD = 0.53; range = 9–11; see Table 1). The data used in the present study were collected prior to the onset of the COVID-19 pandemic.
Informed Consent was obtained from participant parent/guardians and youth provided informed assent. Comprehensive interviews were conducted separately with youth and their parents at baseline and at 1-year follow-up. Data collection was approved by the IRB at UCSD (i.e., the ABCD coordinating center). Detailed information about ABCD Study protocol is available at https://abcdstudy.org. Additional IRB review was not required for these secondary analyses of deidentified data, which did not meet criteria for human subject research.
Measures
Neighborhood Opportunity
We aimed to capture the multidimensional nature of neighborhood opportunity via the Child Opportunity Index (COI). The COI represents an index of neighborhood environment and is intended to capture neighborhood resources and conditions that facilitate healthy child development (Acevedo-Garcia et al. 2014). COI index scores were derived from residential geocodes by the ABCD team based on primary address information provided at baseline. Details on computational procedure and ethical considerations can be found in Fan et al. (2021).
The present analyses used family-level COI scores for the domains of: (1) education (e.g., school poverty rate, student math and reading proficiency, high school graduation rate), (2) health and environment (e.g., proximity to healthcare facilities, retail health food environment index, proximity to parks and open spaces), (3) social and economic opportunity (e.g., foreclosure rate, unemployment rate, public assistance rate), and (4) an overall index based on the weighted average of those three domains. Nationally-normed overall and domain opportunity scores ranged from 1 to 100 (see Table 1). Raw scores were z-transformed for analysis.
Life Stress
At the 1-year follow-up, youth were administered the Life Events Scale (LES) (Hoffman et al. 2019; Grant et al. 2004; Tiet et al. 1998), a 26-item self-report measure of stressful events experienced in their lifetime (e.g., death of a family member, witnessing a crime, negative change parents’ financial situation). Youth indicated whether they had experienced each stressor and, if so, whether it was a “bad” experience. Composite child-level sum scores were created for both total number of experienced events and number of experienced events that the youth indicated were “bad” events.
Externalizing Behaviors
At both baseline and 1-year follow-up, parents were administered the Child Behavior Checklist (CBCL) (Karcher and Barch 2021; Achenbach et al. 2011; Achenbach and Rescorla 2014; Achenbach and Edelbrock 1991) about the child’s behavior over the past 6 months. The CBCL is 112-item dimensional assessment of youth mental health, with externalizing and internalizing scales; for this study, we utilized the externalizing scale (α = 0.89). Items were rated on a 0 (“not true”) to 2 (“very true”) scale and summed to create a raw score total, which was z-transformed for analysis.
Summary of Data Availability
Geocoded neighborhood data (the COI) were available only at baseline, as the data for 1-year follow-up period were still in preparation at the time of the present analyses. The Life Events Scale was only administered at 1-year follow up, thus at a different time point than the COI measure, although changes in address during that 1 year should be restricted to a small minority. Externalizing behaviors were assessed at both baseline and 1-year follow-up. Based on data availability, models focused on COI were examined only at baseline while models focused on stressful life events were examined only at 1-year follow-up. Children were age 10 and 11 at each time point, respectively. Given the narrow age range, major age-related changes in the magnitude of genetic, environmental, and/or GxE effects were not expected during this period.
Analytic Plan
Analyses were conducted in R using the OpenMx (Neale et al. 2016) and umx (Bates et al. 2019) packages in R (Team 2013). First, univariate biometric models were run to estimate the proportion of variance in externalizing behaviors attributable to additive genes (A), common environment (C), and unique environment (E). Models were run for both baseline and follow-up measures of externalizing behaviors. Next, a series of six two-group gene-by-environment interaction models were run (Purcell 2002). These models tested whether the magnitude of genetic or environmental influence varied parametrically across levels of COI and life events, which were each tested as moderators in separate models.
Models using baseline data included (1) overall COI, (2) education COI, (3) health and environment COI, and (4) social and economic COI as separate moderators of externalizing behaviors as reported at baseline. Linear effects of the moderator, as well as linear and quadratic means effects were tested. Twenty-two MZ pairs and 28 DZ pairs were missing data on the moderator; because cases with missing data on the definition variable cannot be included in the model, these pairs were excluded from analyses. Models using follow-up data included (1) cumulative life events and (2) life events experienced as “bad” as separate moderators of externalizing behaviors as reported at the 1-year follow-up. Fourteen MZ pairs and 16 DZ pairs were missing data on the moderator and were excluded from analysis. In all six models, age and sex were included as covariates by creating residualized externalizing variables for both the baseline and the 1-year follow-up measures. Best-fit models were identified by first testing whether moderation parameters could be dropped without decrement in model fit. For moderation parameters that could be dropped, we then tested whether the associated component parameter could also be dropped; we did not attempt to drop the unique environmental component parameter, as it is unlikely that constructs were measured without error. Model comparison was conducted with likelihood ratio tests (Bates et al. 2019).
Results
Descriptive Statistics
Sample characteristics and descriptive statistics of study variables are presented in Table 1. Raw scores for the externalizing composite were 3.45 (observed range = 0–42) and 3.21 (observed range = 0–36) at baseline and 1-year follow up, respectively; t-scores for the externalizing composite were 43.61 (observed range = 33–80) and 43.22 (observed range = 33–75), respectively. The sample had an average overall nationally-normed COI index in the 70th percentile, where higher percentiles reflect higher opportunity scores. The education domain had the lowest average percentile in the sample (approximately 63rd ) and the social and economic domain had the highest (approximately 71st ). COI subdomains were moderately correlated, with the weakest association between education and health-environment domains (r = .51, p < .001) and stronger associations between social-economic and education (r = .71, p < .001) and health-environment domains (r = .76, p < .001). At the 1-year follow-up, youth endorsed experiencing approximately 5 events from the life events checklist, on average; youth endorsed fewer “bad” events (approximately 2, on average). The most frequently endorsed events were death of a family member (50.26%), serious injury of a family member (25.66%), and witnessing a crime or accident (15.72%).
Univariate Models
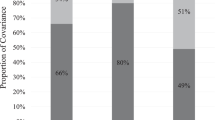
Estimates of genetic, common environmental, and unique environmental influences on externalizing behavior derived from the best-fit models at baseline and 1-year follow-up are depicted in Fig. 1 (complete model comparison is detailed Supplemental Table S1). In the freely estimated baseline model, externalizing behaviors were 74% attributable to A and 26% attributable to E. The C parameter was estimated at 0% and could be constrained to 0. Results were similar for externalizing behavior at 1-year follow-up. In the freely estimated model, externalizing behaviors were 74% attributable to A, 3% attributable to C (ns), and 23% attributable to E. The common environmental parameter could again be constrained to 0, such that 77% of the variance in externalizing was attributable to A and the remaining 23% was attributable to E under the best-fit model.

Proportion of variance in externalizing behavior attributable to additive genetic, common environmental, and unique environmental influences at baseline and the 1-year follow-up
Gene-by-Environment Interaction Models
Overall COI Score (Baseline)
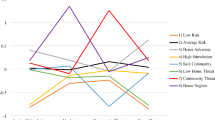
Model comparison and associated fit statistics are available in Supplemental Table S2. Despite no C influence emerging in the univariate model, the component and moderation parameters could not be dropped from the GxE model without decrement in model fit; this is not unprecedented (Hicks et al. 2009). The A and E moderation and the means moderation parameters could be dropped, as could linear and quadratic effects of the mean. Parameter estimates are presented in Table 2. A and E components were statistically significant but were not moderated, meaning that the absolute (unstandardized) variance did not significantly vary across levels of overall COI. Proportional (standardized) results showed that the relative influence of C on externalizing behaviors was substantially higher (approximately 43%) at extremely low COI scores (see Fig. 2). Influence of C dropped to nearly 0% at the mean, explaining the absence of a main effect. The pattern of proportional (standardized) effects showed a relative decrease in A at lower levels of overall COI and a more modest decrease in E at lower levels of overall COI.

Moderation effects for gene-by-environment interaction models. (Note: Figures reflect standardized variance estimates across the spectrum of neighborhood opportunity levels and number of life events experienced; for neighborhood opportunity, higher levels of adversity are reflected in lower values; for life events, higher levels of adversity are reflected in higher values; Va = additive genetic variance; Vc = common environmental variance; Ve = unique environmental variance; *significant moderation parameter)
Education COI Domain (Baseline)
Model comparison and associated fit statistics are available in Supplemental Table S3. Again, the component and moderation parameters of C could not be dropped from the GxE model without decrement in model fit, despite no C influence emerging in the univariate model. The means moderation parameters could be dropped from the model, as could linear and quadratic effects of the mean. Parameter estimates are presented in Table 2. The component effects of A and E were statistically significant, as were the A, C, and E moderation parameters. Proportional (standardized) results showed that the influence of A was approximately 60% at the mean, but 5% and approximately 78% at the extreme low and high education COI scores, respectively (Fig. 2). Conversely, the proportional influence of C was substantially higher at extreme low scores (> 60%), dropping to nearly 0% at and above the mean. This may again explain the absence of a significant main effect of C. The moderation effect was less pronounced for E, as reflected in its effect size, but was still statistically significant. The pattern of effect suggested that the proportional influence of E peaks at approximately 40% for individuals at 1 SD below the mean, and then begins to decrease at higher education COI scores.
Health and Environment COI Domain (Baseline)
Model comparison and associated fit statistics are available in Supplemental Table S4. Both the component and moderation effects of C could be dropped from the model, as could the E and means moderation parameters and the linear and quadratic effects of the mean. Parameter estimates are presented in Table 2. The A and E components were statistically significant, as was the A moderation parameter. Results showed that the proportional (standardized) influence of A increased at lower health and environmental COI scores, peaking at over 80% at extreme low scores and declining to approximately 63% at 1 SD above the mean (Fig. 2). While the absolute (unstandardized) variance contribution for E did not significantly vary across levels of health and environment COI score, the pattern of proportional (standardized) effects showed a slight increase at higher health and environment COI scores, as would be necessitated given the pattern of A influence across levels of this domain.
Social and Economic COI Domain (Baseline)
Model comparison and associated fit statistics are available in Supplemental Table S5. Results were similar to those for the health and environment domains. Both the component and moderation effects of C could be dropped from the model, as could the E and means moderation parameters and the linear and quadratic effects of the mean. Parameter estimates are presented in Table 2. The component effects of A and E were statistically significant, as was the A moderation parameter. Results showed that the proportional (standardized) influence of A increased at lower social and economic opportunity COI scores, peaking at approximately 80% at extreme low scores and declining to approximately 67% at 1 SD above the mean (Fig. 2). Again here, the pattern of standardized effects showed a slight increase in proportion of E influence at higher health and environment COI scores.
Life Stress and Externalizing Behaviors (Follow-Up)
Youth-Reported Cumulative Events
Model comparison and associated fit statistics are available in Supplemental Table S6. Both the component and moderation effects of C could be dropped from the model. Parameter estimates are presented in Table 2. The component effects of A and E were statistically significant, as were the A and E moderation parameters. Proportional (standardized) results showed that A influence declined as number of experienced life events increased, peaking at approximately 87% at 0 events and declining to approximately 63% at over 20 events. Conversely, the influence of E increased as number of experienced life events increased, starting at approximately 13% at 0 events and increasing to approximately 37% at over 20 events (Fig. 2).
Youth-Reported “Bad” Events
Model comparison and associated fit statistics are available in Supplemental Table S7. Results were similar to those from the cumulative events model. Both the main and moderation effects of C could be dropped from the model. Parameter estimates are presented in Table 2. The component effects of A and E were statistically significant, as were the A and E moderation parameters. Proportional (standardized) results showed that A variance declined as number of experience life events increased, peaking at approximately 78% at 0 “bad” events and declining to approximately 67% at over 15 events. Conversely, the influence of E increased as number of experienced life events increased, starting at approximately 22% at 0 events and increasing to approximately 33% at over 15 events (Fig. 2).
Discussion
The present study aimed to evaluate how multiple different domains related to social adversity, including multidimensionally defined neighborhood opportunity and life stress, moderate genetic influences on externalizing behavior among twin pairs of youth participating in the ABCD Study. When considering neighborhood opportunity as a unitary construct, results identified proportionally increasing environmental influence at higher levels of adversity, consistent with the bioecological model, despite mean level of externalizing behavior remaining consistent across COI scores. However, these proportional GxE effects were not uniform across COI and stress-related domains, with some domains showing an opposite pattern and supporting a diathesis-stress model. Overall, results highlight the importance of decomposing the construct of social adversity, and particularly domains of neighborhood opportunity, to better understand optimal environmental targets for intervention.
Findings related to educational COI indicated drastically diminished proportional genetic influence on externalizing behaviors at extreme low levels (-2 SD) of educational opportunity that increased rapidly as educational opportunity scores increased. Proportionally decreasing genetic effects at higher levels of adversity support a bioecological GxE process (Raine 2002), which posits that genetic influences are relatively less important in the manifestation of externalizing behavior at higher levels of environmental adversity. One mechanism posited for this effect is social spread (Burt et al. 2019), wherein youth observe externalizing behavior within their neighborhood environment, which in turn enhances the likelihood of youth engaging in externalizing behaviors themselves via a shared environmental (as opposed to genetic) mechanism. Youth in higher-opportunity neighborhoods may observe fewer such behaviors among peers and family members, such that genetic influences proportionally predominate in these contexts. Given that the developmental literature has phenotypically linked factors such as negative school climate with youth expression of externalizing behavior (Reaves et al. 2018; Hendron and Kearney 2016), findings here indicate that community-based prevention programs targeting environmental factors such as school climate, quality, resources, and educational opportunities could help attenuate and mitigate development of externalizing behavior among youth in lower-opportunity neighborhoods, whose liability for externalizing behavior is predominantly environmental.
Results also found that genetic influences proportionally decrease with more stressful life events experienced, regardless of whether events were experienced by youth as “bad.” While this does not align with results reported in several studies conducted among older adolescents (age 17) (Fairchild et al. 2019; Hicks et al. 2009; Samek and Hicks 2014), a pattern of dominant environmental influences in the context of increased stressful life events such as the one identified in the present results is not wholly unprecedented (Button et al. 2005, 2008). For example, a study of 12–19 year old twins found that the proportional influence of genes decreased at higher levels of adversity while the proportional influence of unique environment increased. However, it should be noted that this proportional shift was the result of moderation of unique environmental variance (with absolute values being higher at higher levels of adversity) rather than differences in absolute genetic variance across levels of adversity (which did not vary across level of adversity) (Button et al. 2008). Such inconsistencies within the existing literature may be attributable to distinct patterns of effect across developmental periods (Burt and Klump 2014; Samek et al. 2017). As evidenced both in prior studies and the present findings, GxE studies of life stress that include a younger age group (< 17 years) tend to find support for a bioecological process while studies that focus on older adolescents tend to find support for a diathesis-stress process.
In contrast to the findings for educational opportunity and life stress, effects for health-environment and social-economic COI domains indicated that genetic liability for externalizing behavior proportionally increased at higher levels of social adversity (or, alternately, lower levels of neighborhood-level opportunity). These findings align with diathesis-stress models of psychopathology (Monroe and Simons 1991). The specificity of results supporting a diathesis-stress process to health and social-economic domains suggests that a lack of access to healthcare, housing, and employment may potentiate genetic liability for youth externalizing behavior. As such, public and healthcare policy addressing healthcare access, housing security, and employment stability may be an optimal strategy for mitigating genetic liability for externalizing behavior in the context of limited resources.
Overall, findings offer strong support for the multidimensionality of adversity, with different facets of opportunity and stress differentially moderating genetic, common environmental, and unique environmental influences on externalizing behaviors. Whereas limited educational opportunity and stressful life events may enhance the likelihood of a bioecological “social push” towards externalizing behavior, lack of access to healthcare, housing, and employment opportunities may function via a diathesis-stress mechanism wherein such facets of adversity potentiate latent genetic risk for externalizing behavior. When only the overall COI metric is considered, findings will be driven by whichever effects are strongest in the particular sample being studied. These conflicting patterns also indicate the inherently complicated and multidimensional nature of these constructs and indicate that composite metrics of SES may obscure important nuance of relevant subdomains.
Limitations
While the current study takes a novel approach to disaggregating the impact of complicated interactions within the multidimensional construct of adversity, findings should be interpreted in light of the following limitations. First, although the ABCD sample is a rich and unique dataset derived from a contemporary and geographically diverse sample, it is a volunteer sample and not a true epidemiological sample; as such, it does not in practice reflect the demographics of the United States in terms of race, ethnicity, or SES. Second, COI data were only available at baseline and life stress data were only available at the 1-year follow up, limiting our ability to test simultaneous effects of the moderators or how effects change over time.
Additionally, the degree of externalizing behavior in the sample was quite low overall, which may reflect a degree of volunteer bias or may reflect lower overall rates in national versus local samples. While the ABCD sample is an exceptional resource, additional studies in case-control samples enriched for specific types of psychopathology and studies that include more nuanced and in-depth measures of psychopathology are both needed. For example, the broad operationalization of externalizing behaviors implemented here combines items related to proactive aggression, reactive aggression, and irritability/anger dysregulation, potentially obscuring unique effects in each domain. There is evidence that, even among related behaviors and conditions, GxE effects can emerge in opposing patterns (i.e., one may be more heritable in a favorable environment than in a risk environment and vice versa) (Pennington et al. 2009). A valuable next step will be to examine these GxE relationships with more specific constructs.
Conclusion
Neighborhood-level opportunity in the educational, health and environmental, and social and economic domains, as well as life stress, are critical environmental facets of social adversity that have a well-established link to externalizing behavior in youth. Though commonly researched, social adversity is often operationalized simply as a unidimensional metric of neighborhood disadvantage (e.g., poverty). The present study suggests that a more fine-grained assessment of social adversity is warranted, and that magnitude and source of its effects vary across facets of neighborhood-level opportunity. Understanding the varied influence of the environment and genetics on externalizing behavior is an important step in identifying targets for intervention and the optimal level at which to direct resources to maximize the prevention and intervention efforts.
Data Availability
De-identified data is available for this study; please see https://nda.nih.gov/abcd. The ABCD data repository grows and changes over time. The ABCD data used in this report came from https://doi.org/10.15154/1527738.
Code Availability
Code is available upon request to the first author.
References
Abry T, Bryce CI, Swanson J, Bradley RH, Fabes RA, Corwyn RF (2017) Classroom-level adversity: Associations with children’s internalizing and externalizing behaviors across elementary school. Dev Psychol 53(3):497
Acevedo-Garcia D, McArdle N, Hardy EF, Crisan UI, Romano B, Norris D, Baek M, Reece J (2014) The child opportunity index: improving collaboration between community development and public health. Health Aff 33(11):1948–1957
Achenbach TM, Edelbrock C (1991) Child behavior checklist. Burlington (Vt, 7371 – 392
Achenbach TM, McConaughy S, Ivanova M, Rescorla L (2011) Manual for the ASEBA brief problem monitor (BPM). ASEBA 33, Burlington, VT
Achenbach TM, Rescorla LA (2014) The Achenbach system of empirically based assessment (ASEBA) for ages 1.5 to 18 years. InThe use of psychological testing for treatment planning and outcomes assessment. Routledge, pp 179–214
Albert D, Belsky DW, Crowley DM, Latendresse SJ, Aliev F, Riley B, Sun C, Conduct Problems Prevention Research Group, Dick DM, Dodge KA (2015) Can genetics predict response to complex behavioral interventions? Evidence from a genetic analysis of the fast Track Randomized Control Trial. J Policy Anal Management: 34(3):497–518
Assary E, Vincent JP, Keers R, Pluess M (2018) Gene-environment interaction and psychiatric disorders: review and future directions. InSeminars in cell & developmental biology. Elsevier, pp 133–143
Barr PB, Dick DM (2019) The genetics of externalizing problems. Recent advances in research on impulsivity and impulsive behaviors 93–112
Bates TC, Maes H, Neale MC (2019) umx: twin and path-based structural equation modeling in R. Twin Res Hum Genet 22(1):27–41
Border R, Johnson EC, Evans LM, Smolen A, Berley N, Sullivan PF, Keller MC (2019) No support for historical candidate gene or candidate gene-by-interaction hypotheses for major depression across multiple large samples. Am J Psychiatry 176(5):376–387
Bornstein MH, Hahn C-S, Suwalsky JT (2013) Developmental pathways among adaptive functioning and externalizing and internalizing behavioral problems: cascades from childhood into adolescence. Appl Dev Sci 17(2):76–87
Burt SA (2015) Evidence that the gene–environment interactions underlying youth conduct problems vary across development. Child Dev Perspect 9(4):217–221
Burt SA (2022) The genetic, environmental, and cultural forces influencing youth antisocial behavior are tightly intertwined.Annual review of clinical psychology18155–178
Burt S, Klump K (2014) Parent–child conflict as an etiological moderator of childhood conduct problems: an example of a ‘bioecological’gene–environment interaction. Psychol Med 44(5):1065–1076
Burt SA, Klump KL, Gorman-Smith D, Neiderhiser JM (2016) Neighborhood disadvantage alters the origins of children’s nonaggressive conduct problems. Clin Psychol Sci 4(3):511–526
Burt SA, Pearson AL, Rzotkiewicz A, Klump KL, Neiderhiser JM (2019) It really does take a village: the role of neighbors in the etiology of nonaggressive rule-breaking behavior. Dev Psychopathol 31(2):713–725
Burt SA, Pearson AL, Carroll S, Klump KL, Neiderhiser JM (2020) Child antisocial behavior is more environmental in origin in disadvantaged neighborhoods: evidence across residents’ perceptions and geographic scales in two samples. J Abnorm Child Psychol 48(2):265–276
Button TM, Lau JY, Maughan B, Eley T (2008) Parental punitive discipline, negative life events and gene–environment interplay in the development of externalizing behavior. Psychol Med 38(1):29–39
Button TMM, Scourfield J, Martin N, Purcell S, McGuffin P (2005) Family dysfunction interacts with genes in the causation of antisocial symptoms. Behav Genet 35(2):115–120
Carroll SL, Klump KL, Burt SA (2022) Understanding the effects of neighborhood disadvantage on youth psychopathology.Psychological Medicine1–11
Caspi A, McClay J, Moffitt TE, Mill J, Martin J, Craig IW, Taylor A, Poulton R (2002) Role of genotype in the cycle of violence in maltreated children. Science 297(5582):851–854
Cheung AK, Harden KP, Tucker-Drob EM (2014) Gene×Environment interactions in early externalizing behaviors: parental emotional support and socioeconomic context as moderators of genetic influences? Behav Genet 44(5):468–486
Choi J-K, Teshome T, Smith J (2021) Neighborhood disadvantage, childhood adversity, bullying victimization, and adolescent depression: a multiple mediational analysis.Journal of affective disorders279554–562
Dardani C, Riglin L, Leppert B, Sanderson E, Rai D, Howe LD, Davey Smith G, Tilling K, Thapar A, Davies NM (2021) Is genetic liability to ADHD and ASD causally linked to educational attainment? Int J Epidemiol 50(6):2011–2023
Devenish B, Hooley M, Mellor D (2017) The pathways between socioeconomic status and adolescent outcomes: a systematic review. Am J Community Psychol 59(1–2):219–238
Dick DM (2011) Gene-environment interaction in psychological traits and disorders. Annual review of clinical psychology 7383 – 409
Dishion TJ, Kavanagh K (2003) Adolescent problem behavior: an intervention and assessment sourcebook for working with families in schools. Guilford, New York
Fairchild G, Hawes DJ, Frick PJ, Copeland WE, Odgers CL, Franke B, Freitag CM, De Brito SA (2019) Conduct disorder. Nat Reviews Disease Primers 5(1):1–25
Fan CC, Marshall A, Smolker H, Gonzalez MR, Tapert SF, Barch DM, Sowell E, Dowling GJ, Cardenas-Iniguez C, Ross J (2021) Adolescent brain Cognitive Development (ABCD) study linked External Data (LED): protocol and practices for geocoding and assignment of environmental data.Developmental cognitive neuroscience52101030
Farrell MS, Werge T, Sklar P, Owen MJ, Ophoff RA, O’Donovan MC, Corvin A, Cichon S, Sullivan PF (2015) Evaluating historical candidate genes for schizophrenia. Mol Psychiatry 20(5):555–562
Gartland D, Riggs E, Muyeen S, Giallo R, Afifi TO, MacMillan H, Herrman H, Bulford E, Brown SJ (2019) What factors are associated with resilient outcomes in children exposed to social adversity? A systematic review. BMJ open 9(4):e024870
Gould KL, Coventry WL, Olson RK, Byrne B (2018) Gene-environment interactions in ADHD: the roles of SES and chaos. J Abnorm Child Psychol 46(2):251–263
Grant KE, Compas BE, Thurm AE, McMahon SD, Gipson PY (2004) Stressors and child and adolescent psychopathology: measurement issues and prospective effects. J Clin Child Adolesc Psychol 33(2):412–425
Hatoum AS, Rhee SH, Corley RP, Hewitt JK, Friedman NP (2018) Etiology of stability and growth of internalizing and externalizing behavior problems across childhood and adolescence. Behav Genet 48(4):298–314
Hendron M, Kearney CA (2016) School climate and student absenteeism and internalizing and externalizing behavioral problems. Child Schools 38(2):109–116
Henggeler SW, Rodick JD, Borduin CM, Hanson CL, Watson SM, Urey JR (1986) Multisystemic treatment of juvenile offenders: Effects on adolescent behavior and family interaction. Dev Psychol 22(1):132–141
Hentges RF, Krug CM, Shaw DS, Wilson MN, Dishion TJ, Lemery-Chalfant K (2020) The long-term indirect effect of the early Family Check-Up intervention on adolescent internalizing and externalizing symptoms via inhibitory control Development and psychopathology 32(4):1544-54
Hicks BM, South SC, DiRago AC, Iacono WG, McGue M (2009) Environmental adversity and increasing genetic risk for externalizing disorders. Arch Gen Psychiatry 66(6):640–648
Hoffman EA, Clark DB, Orendain N, Hudziak J, Squeglia LM, Dowling GJ (2019) Stress exposures, neurodevelopment and health measures in the ABCD study.Neurobiology of stress10100157
Hoglund WL, Klingle KE, Hosan NE (2015) Classroom risks and resources: teacher burnout, classroom quality and children’s adjustment in high needs elementary schools. J Sch Psychol 53(5):337–357
Holz NE, Zohsel K, Laucht M, Banaschewski T, Hohmann S, Brandeis D (2018) Gene x environment interactions in conduct disorder: implications for future treatments.Neuroscience & Biobehavioral Reviews91239–258
Iacono WG, Heath AC, Hewitt JK, Neale MC, Banich MT, Luciana MM, Madden PA, Barch DM, Bjork JM (2018) The utility of twins in developmental cognitive neuroscience research: how twins strengthen the ABCD research design. Dev Cogn Neurosci 32:30–42
Jangmo A, Kuja-Halkola R, Pérez-Vigil A, Almqvist C, Bulik CM, D’Onofrio B, Lichtenstein P, Ahnemark E, Werner-Kiechle T, Larsson H (2021) Attention-deficit/hyperactivity disorder and occupational outcomes: the role of educational attainment, comorbid developmental disorders, and intellectual disability. PLoS ONE 16(3):e0247724
Jernigan TL, Brown SA, Dowling GJ (2018) The adolescent brain cognitive development study. J Res adolescence: official J Soc Res Adolescence 28(1):154–156
Karcher NR, Barch DM (2021) The ABCD study: understanding the development of risk for mental and physical health outcomes. Neuropsychopharmacology 46(1):131–142
Lewis GJ, Asbury K, Plomin R (2017) Externalizing problems in childhood and adolescence predict subsequent educational achievement but for different genetic and environmental reasons. J Child Psychol Psychiatry 58(3):292–304
Loth AK, Drabick DA, Leibenluft E, Hulvershorn LA (2014) Do childhood externalizing disorders predict adult depression? A meta-analysis. J Abnorm Child Psychol 42(7):1103–1113
March-Llanes J, Marqués-Feixa L, Mezquita L, Fañanás L, Moya-Higueras J (2017) Stressful life events during adolescence and risk for externalizing and internalizing psychopathology: a meta-analysis. Eur Child Adolesc Psychiatry 26(12):1409–1422
Maxwell MY, Taylor RL, Barch DM (2022) Relationship between Neighborhood Poverty and externalizing symptoms in children: mediation and moderation by environmental factors and brain structure. Child Psychiatry & Human Development, pp 1–13
McNeilly EA, Peverill M, Jung J, McLaughlin KA (2021) Executive function as a mechanism linking socioeconomic status to internalizing and externalizing psychopathology in children and adolescents.Journal of Adolescence89149–160
Molina BS, Howard AL, Swanson JM, Stehli A, Mitchell JT, Kennedy TM, Epstein JN, Arnold LE, Hechtman L, Vitiello B (2018) Substance use through adolescence into early adulthood after childhood-diagnosed ADHD: findings from the MTA longitudinal study. J Child Psychol Psychiatry 59(6):692–702
Monroe SM, Simons AD (1991) Diathesis-stress theories in the context of life stress research: implications for the depressive disorders. Psychol Bull 110(3):406
Neale MC, Hunter MD, Pritikin JN, Zahery M, Brick TR, Kirkpatrick RM, Estabrook R, Bates TC, Maes HH, Boker SM (2016) OpenMx 2.0: extended structural equation and statistical modeling. Psychometrika 81(2):535–549
Pearson AL, Shewark EA, Burt SA (2022) Associations between neighborhood built, social, or toxicant conditions and child externalizing behaviors in the Detroit metro area: a cross-sectional study of the neighborhood ‘exposome’. BMC Public Health 22(1):1–13
Pennington BF, McGrath LM, Rosenberg J, Barnard H, Smith SD, Willcutt EG, Friend A, DeFries JC, Olson RK (2009) Gene× environment interactions in reading disability and attention-deficit/hyperactivity disorder. Dev Psychol 45(1):77
Plomin R, Gidziela A, Malanchini M, von Stumm S (2022) Gene–environment interaction using polygenic scores: do polygenic scores for psychopathology moderate predictions from environmental risk to behavior problems?Development and Psychopathology1–11
Purcell S (2002) Variance components models for gene–environment interaction in twin analysis. Twin Res Hum Genet 5(6):554–571
Raine A (2002) Biosocial studies of antisocial and violent behavior in children and adults: a review. J Abnorm Child Psychol 30(4):311–326
Ramos-Olazagasti MA, Castellanos FX, Mannuzza S, Klein RG (2018) Predicting the adult functional outcomes of boys with ADHD 33 years later. J Am Acad Child Adolesc Psychiatry 57(8):571–582 e571
R Core Team (2013) R: A language and environment for statistical computing
Reaves S, McMahon SD, Duffy SN, Ruiz L (2018) The test of time: a meta-analytic review of the relation between school climate and problem behavior.Aggression and violent behavior39100–108
Reiss F (2013) Socioeconomic inequalities and mental health problems in children and adolescents: a systematic review.Social science & medicine9024–31
Reiss F, Meyrose A-K, Otto C, Lampert T, Klasen F, Ravens-Sieberer U (2019) Socioeconomic status, stressful life situations and mental health problems in children and adolescents: results of the german BELLA cohort-study. PLoS ONE 14(3):e0213700
Sakhvidi MJZ, Knobel P, Bauwelinck M, de Keijzer C, Boll LM, Spano G, Ubalde-Lopez M, Sanesi G, Mehrparvar AH, Jacquemin B (2022) Greenspace exposure and children behavior: a systematic review. Science of the Total Environment 153608
Samek DR, Hicks BM (2014) Externalizing disorders and environmental risk: Mechanisms of gene-environment interplay and strategies for intervention. Clinical practice (London, England) 11(5):537
Samek DR, Hicks BM, Keyes MA, Iacono WG, McGue M (2017) Antisocial peer affiliation and externalizing disorders: evidence for Gene× Environment× development interaction. Dev Psychopathol 29(1):155–172
Tiet QQ, Bird HR, Davies M, Hoven C, Cohen P, Jensen PS, Goodman S (1998) Adverse life events and resilience. J Am Acad Child Adolesc Psychiatry 37(11):1191–1200
Turney K (2022) Chains of adversity: the time-varying consequences of paternal incarceration for adolescent behavior. J Quant Criminol 38(1):159–196
Tuvblad C, Grann M, Lichtenstein P (2006) Heritability for adolescent antisocial behavior differs with socioeconomic status: gene–environment interaction. J Child Psychol Psychiatry 47(7):734–743
Viner RM, Ozer EM, Denny S, Marmot M, Resnick M, Fatusi A, Currie C (2012) Adolescence and the social determinants of health. The lancet 379(9826):1641–1652
Wickham S, Whitehead M, Taylor-Robinson D, Barr B (2017) The effect of a transition into poverty on child and maternal mental health: a longitudinal analysis of the UK Millennium Cohort Study. The Lancet Public Health 2(3):e141–e148
Womack SR, Wilson MN, Tong X, Lemery-Chalfant K, Shaw DS (2022) Trajectories of early childhood family instability and the development of externalizing behaviors from middle childhood to adolescence: a prospective study of at‐risk families. Child development
Wright EM, Schwartz JA (2021) The influence of adverse childhood experiences on internalizing and externalizing problems in early adulthood: evidence of a gene× environment× sex interaction. Child Abuse & Neglect 114104962
Zandstra ARE, Ormel J, Hoekstra PJ, Hartman CA (2018) Chronic stressors and adolescents’ externalizing problems: genetic moderation by dopamine receptor D4. The TRAILS study. J Abnorm Child Psychol 46(1):73–82
Funding
This research was supported by the National Institute on Drug Abuse grant numbers F31DA054701 (PI: Dash) and U01DA041148-01 (MPIs: Nagel, Feldstein Ewing, Fair), the National Institute on Alcohol Abuse and Alcoholism grant number K24AA026876-01 (PI: Feldstein Ewing), and the National Institute on Mental Health grant number R37MH059105 (PI: Nigg). Data used in the preparation of this article were obtained from the Adolescent Brain Cognitive DevelopmentSM(ABCD) Study (https://abcdstudy.org), held in the NIMH Data Archive (NDA). This is a multisite, longitudinal study designed to recruit more than 10,000 children age 9–10 and follow them over 10 years into early adulthood. The ABCD Study® is supported by the National Institutes of Health and additional federal partners under award numbers U01DA041048, U01DA050989, U01DA051016, U01DA041022, U01DA051018, U01DA051037, U01DA050987, U01DA041174, U01DA041106, U01DA041117, U01DA041028, U01DA041134, U01DA050988, U01DA051039, U01DA041156, U01DA041025, U01DA041120, U01DA051038, U01DA041148, U01DA041093, U01DA041089, U24DA041123, U24DA041147. A full list of supporters is available at https://abcdstudy.org/federal-partners.html. A listing of participating sites and a complete listing of the study investigators can be found at https://abcdstudy.org/consortium_members/. ABCD consortium investigators designed and implemented the study and/or provided data but did not necessarily participate in the analysis or writing of this report. This manuscript reflects the views of the authors and may not reflect the opinions or views of the NIH or ABCD consortium investigators.
Author information
Authors and Affiliations
Contributions
Secondary study design and data access (GFD, SLK, SWFE), data analysis (GFD), drafting of manuscript (all authors), critical revision of manuscript (all authors), final approval of manuscript (all authors).
Corresponding author
Ethics declarations
Conflicts of Interest/Competing Interests
The authors have no conflict of interest to disclose.
Ethics Approval
Original data collection was approved by the IRB at UCSD (i.e., the ABCD coordinating center). Detailed information about ABCD Study protocol is available at https://abcdstudy.org. IRB review was not required for these secondary analyses of deidentified data, which did not meet criteria for human subjects research.
Consent to Participate
Not applicable (secondary analysis).
Consent for Publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Sylia Wilson
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Dash, G.F., Karalunas, S.L., Kenyon, E.A. et al. Gene-by-Environment Interaction Effects of Social Adversity on Externalizing Behavior in ABCD Youth. Behav Genet 53, 219–231 (2023). https://doi.org/10.1007/s10519-023-10136-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10519-023-10136-z