
Abstract
We herein investigated Lactobacillus rhamnosus as a probiotic, paraprobiotic and their synbiotic combination in the diet of dourado Salminus brasiliensis. Two hundred and forty juvenile S. brasiliensis with average weight and length of 6.78 ± 1.65 g and 8.97 ± 0.42 cm were randomly distributed in 16 experimental units (300 L) under four treatments: diet without additive (control), 0.02% probiotic (L. rhamnosus 108 CFU), 2.0% paraprobiotic (inactive L. rhamnosus 1010 CFU) and synbiotic (probiotic + paraprobiotic), all in quadruplicate, for 45 days. Then, zootechnical performance, hemato-biochemicals (pre-and post-challenge), intestinal and hepatic histomorphometric analyses were performed, in addition to a bacterial challenge with Aeromonas hydrophila. The feed intake was significantly lower (p < 0.05) in the probiotic group. The allomeric condition factors were significantly higher (p < 0.05) in fish from the paraprobiotic and synbiotic groups. No significant differences (p > 0.05) were observed in the blood count between the supplemented groups and the control group. However, significant differences (p < 0.05) were observed in the blood count between the pre- and post-challenge periods. Fish in the synbiotic groups had higher total villi height and villi height when compared to fish in the probiotic and control group. The serosa were significantly (p < 0.05) thicker in the intestines of fish from the probiotic and synbiotic groups. Goblet cells were significantly numerous (p < 0.05) in fish from the synbiotic group. In conclusion, the use of 0.02% probiotic, 2.0% paraprobiotic, and synbiotic proved to be a promising practice due to their action on productive performance, enabling weight gain similar to that of control group with lower feed intake. In addition to improving intestinal immune-histomosphometric parameters, possibly promoting an improved condition in facing intestinal pathogens.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Even with the predominance of Nile tilapia (Oreochromis niloticus) in Brazilian fish farming, the native Brazilian species also increased by 1.8% in statistics, reaching the mark of 267,060 tons in 2022 (PeixeBR 2023). It is not surprising that indigenous fish species contribute to the advancement of Brazilian aquaculture, given that in recent years, the emphasis of production and national research has given priority to native species (Saint-Paul 2017; Valenti et al. 2021) and that more than 30% of total production is currently derived from native species (PeixeBR 2023). This supports the notion that enhancing species diversification can enhance aquaculture (Oboh 2022). Within this scenario emerges the dourado (Salminus brasiliensis).
Dourado S. brasiliensis belongs to the Characidae family and has a wide geographic distribution, with great potential for fish farming characterized by high weight gain and accelerated growth in the nursery phase, spending less time in this phase (30% shorter) than most species (Della Flora et al. 2010). However, the growth performance of S. brasiliensis can be negatively affected by the stress of frequent handling in cage systems (Braun et al. 2010), while nutritional strategies with adequate inclusion of vitamin A in the diet (Koch et al. 2018) or vegetable-based diets supplemented with swine liver hydrolysate (Lorenz et al. 2022) increase the immunomodulation activity (Koch et al. 2018) and the growth performance of the species (Lorenz et al. 2022).
Research results such as those mentioned above boost the use of potentially immunostimulating bioproducts in fish feeding as a strategy to enhance defense mechanisms in the prevention of stress and resistance against infectious diseases (Vallejos-Vidal et al. 2016; Hoseini et al. 2020). In this context, probiotics stand out as dietary supplements to improve the immune response and zootechnical performance (Navarrete and Tovar-Ramírez 2014; Akhter et al. 2015; Hoseinifar et al. 2018).
Research has already reported that non-viable counterparts of probiotic organisms, known as paraprobiotics (Choudhury and Kamilya 2019; Cuevas-González et al. 2020), inactivated probiotics or ghost probiotics (Taverniti and Guglielmetti 2011; Almada et al. 2016; Piqué et al. 2019), when administered in adequate amounts and frequency, also provide benefits to the animal. On the other hand, synbiotics are a combined mixture of food additives that can confer a synergistic and potentiated effect of benefits to the host and have been highlighted as immunostimulants in research in several areas of animal production, such as broiler chickens (Soomro et al. 2019; Abd El-Hack et al. 2020), ruminants (Arowolo and He 2018; Raabis et al. 2019; Mahesh et al. 2021) and fish (Elumalai et al. 2020; El-Saadony et al. 2021; Oliveira et al. 2022).
In this diverse universe of beneficial microorganisms, there are those of the genus Lactobacillus, –belonging to the group of gram-positive lactic acid bacteria (BAL), nonsporeforming (Tamang 2014)– which can adapt and colonize the intestinal epithelium in different environmental conditions (Pfeiler and Klaenhammer 2007). In fish diets, the use of Lactobacillus rhamnosus was related to improvements in intestinal morphology and immunity, and disease resistance in Nile tilapia (Pirarat et al. 2011; Xia et al. 2018), while in Pargus major improves the immune system and physiological responses (Dawood et al. 2017). On the other hand, dietary supplementation of L. rhamnosus for Oncorhynchus mykiss was efficient in controlling infections by Vibrio anguillarum, Flavobacterium psychrophilum, and Aeromonas salmonicida (Nikoskelainen et al. 2003).
The genus Aeromonas are the main causing agents for diseases frequently found in fish farms, where the species A. hydrophila, which are freshwater, facultatively anaerobic bacterium, cause disease outbreaks with high mortality rates (Austin et al. 2016; Tavares-Dias and Martins 2017; Semwal et al. 2023) and growth impairments (Carraschi et al. 2011), becoming a limiting factor for the development of aquaculture.
Species with a high productive potential, such as the dourado S. brasiliensis, are in evidence in Brazilian aquaculture due to their great economic, social and environmental importance (Della Flora et al. 2010). In view of this purpose, the aim of the present study was to investigate the probiotic, paraprobiotic and combined effects of dietary supplementation with L. rhamnosos on growth performance, hematology, intestinal and hepatic histology and resistance of S. brasiliensis after bacterial challenge with A. hydrophila.
Material and methods
Experimental design
Two hundred and forty healthy juvenile dourado S. brasiliensis with average initial weight and length of 3.49 ± 0.20 g and 7.32 ± 0.12 cm obtained from the commercial fish farm Projeto Pacu, Terenos, Mato Grosso do Sul, Brazil, were randomly distributed in 16 circular experimental units (300 L) with constant aeration and continuous water-flow, housed in a greenhouse, totaling 15 fish per tank. The fish went through a pre-assay acclimatization period for three weeks and were fed a commercial diet (Laguna carnivorous fish / Socil® crude protein 450 g kg−1; ethereal extract 120 g kg−1; crude fiber 45 g kg−1; mineral matter 140 g kg−1; calcium (Min.) 15 g kg−1; calcium (Max.) 25 g kg−1; phosphorus 10 g kg−1; moisture 120 g kg−1; vitamin C 600 mg kg−1) three times daily at a rate of 5%. During this period, no clinical signs such as erratic swimming, opacity of the cornea, lesions on the body or any other signs indicating diseases were observed.
After the acclimatization period, the fish had an average weight and length of 6.78 ± 1.65 g and 8.97 ± 0.42 cm and were submitted to four established treatments: feed supplemented with (manufacturer recommended inclusions) 0.02% of probiotic (L. rhamnosus 108 colony forming unit—CFU); 2.0% of paraprobiotic (inactive strains of L. rhamnosus 1010 CFU); synbiotic supplementation (0.02% probiotic + 2.0% paraprobiotic), and feed without additive (control group); all in quadruplicate according to Ruiz et al (2020).
The additives were added in an oily medium (soybean oil) and sprinkled directly on the commercial feed described above (Laguna pescado carnivores / Socil®) and then placed to dry at room temperature according to Nakandakare et al. (2018). For the control diet, only soybean oil was added to the feed. The probiotic Rham GB® (lyophilized microbial culture of L. rhamnosus) and the paraprobiotic Neoimuno® HealthCare (inactive strains of L. rhamnosus) were donated by the company GABBIA Biotecnologia®. The experimental diets were analysed by the company for the microbiological presence of the probiotic L. rhamnosus in MRS agar medium. After 48 h in a bacteriological incubator at 37° C, the analyses of the plates were performed. The microbiological analyzes for the presence of L. rhamnosus showed control diet = absence; probiotic diet = L. rhamnosus 2.50 × 107 CFU g−1; paraprobiotic diet = absence; and synbiotic diet (viable + inactive) = L. rhamnosus 2.35 × 107 CFU g−1.
Fish were fed twice daily (8:00 am and 4:00 pm) until apparent satiation for 45 days, and the amounts of feed provided were calculated. Monitoring of water quality variables were performed daily and mean values in the morning remained at pH 6.90 ± 0.64, dissolved oxygen 6.19 ± 0.77 mg L−1 and temperature 25.03 ± 0.87 °C; while in the afternoon they remained with pH 7.52 ± 0.61, dissolved oxygen 5.14 ± 0.93 mg L−1 and temperature 28.37 ± 0.87 °C. Ammonia, nitrite and nitrate remained undetectable. Whenever necessary, the experimental units were siphoned to remove feces and leftover feed.
Zootechnical performance
After 45 days of supplementation, the fish fasted for 24 h and were subsequently anesthetized with Eugenol (50 mg L−1) for biometry. With the biometric data, the zootechnical indexes were calculated for weight gain (WG) = (final weight—initial weight); length gain (LG) = (final length—initial length); apparent feed conversion (FCR) = (feed intake (g) / weight gain); protein efficiency ratio (PER) = ((weight gain / protein intake) × 100), specific growth rate (SGR) = ((ln Final weight/Initial weight/days) × 100). Survival rates (SR) (initial fish number—final fish number) were calculated after 45 days of dietary supplementation as well as on the seventh post-challenge day, according to Nunes et al. (2020). The allometric condition factors were calculated by the formula K = W/Lb where, where b is estimated by the weight-length equation (W = aLb; where W is the weight and L is the total length and a and b are estimates of the correlation parameters), after logarithmic transformation and adjustment by the method of least squares of the data (Lima-Junior et al. 2002).
Hemato-biochemicals analyses
Three fish from each experimental unit, 12 per treatment, were removed from the units and used in blood collections for complete blood count analysis before the bacterial challenge. The fish remaining in the experimental units were challenged with the bacterium A. hydroplila and 24 h post-challenge, aliquots of blood from three fish per experimental unit were collected again for complete blood count. Blood collections were performed by puncturing the caudal vessel with insulin syringes coated with anticoagulant solution HEMSTB (EDTA K2 15 g dL−1). Hematocrit (Ht) was determined by the microhematocrit method (Goldenfarb et al. 1971). Hemoglobin (Hb) levels were determined by spectrometry according to the cyanmethemoglobin method (Collier 1944). The number of erythrocytes (Er) was performed in in Neubauer's chamber after diluting in formalin citrate solution (1:200). From these data, the hematimetric indexes were calculated: mean corpuscular volume (MCV) = (Ht × 10/Er); mean corpuscular hemoglobin concentration (MCHC) = (Hb × 100/Ht); and mean corpuscular hemoglobin (MCH) = Hb/10 × Er (Wintrobe 1934). Blood smears were made in duplicate from each animal, the slides air-dried and stained with May Grünwald-Giemsa-Wright (Tavares-Dias and Moraes 2004). Differential leukocyte counts (lymphocytes, neutrophils, monocytes, eosinophils, granulocytic cells or granular leukocyte PAS (LG-PAS) and basophils) were performed under an optical microscope (1000 ×) with the aid of a digital differential counter.
For blood glucose analysis (glucose oxidase [GPO]) the roche accu chek active glucose measuring device was used. Leukocyte activities were determined according to the methodology described by Biller et al. (2013). This method consists of determining the reactive oxygen species (ROS) produced by the respiratory burst through a colorimetric assay based on the reduction of the nitroblue tetrazolium (NBT) reagent, which gives rise to precipitates of insoluble material with a dark blue color inside the phagocyte, called formazan granules (Klein et al. 1990). Fifty microliters of blood added to 50 μL of 0.2% nitroblue tetrazolium (NBT) solution were used. The NBT solution was prepared in buffer (PBS) and incubated for 30 min at 25 °C. After the incubation period, 50 μL of the solution were diluted in 1000 μL (1 mL) of N-dimethyl formamide (DMF) and centrifuged at 3000 g for 5 min. The reading of the samples was determined by spectrophotometry at a wavelength of 540 nm.
Intestinal and hepatic histomorphometric analyses
At the end of the dietary supplementation period, two fish from each experimental unit (n = 32) were anesthetized with Eugenol (50 mg L−1) and euthanized by spinal cord transection and submitted to a longitudinal ventral incision to expose the intestine and liver. Tissue fragments from the liver and posterior portion of the medial intestine of approximately three cm were fixed in 10% buffered formalin. The samples were then washed and dehydrated in increasing series of ethyl alcohol, clarified in xylol and embedded in paraffin at 60 °C to be cut in 4 μm sections and stained with Hematoxylin–Eosin (H & E) according to Humason (1972).
Subsequently, the slides were prepared in Entellan® media and the histological sections were photomicrographed using an opticam 14.0 camera and the variables were analyzed using the Motic Images Plus 2.0 ML program. For intestinal histomorphometry, the total villi height (µm), villi height (µm), villi width (µm), epithelium thickness (µm), lamina propria (µm), submucosa (µm), layer muscle (µm), serosa (µm) and number of goblet cells per villi (n = 40 per region). For liver histomorphometry, 50 hepatocytes were randomly sampled to measure the area (μm2) and perimeter of the cytoplasm (μm); area (μm2), perimeter (μm) and diameter (μm) of the nucleus; nucleus area/cytoplasm area ratio (Nacr = nuclear area/cytoplasm area × 100); nucleus perimeter/cytoplasm ratio (Npcr = nuclear perimeter/cytoplasm perimeter × 100); hepatocyte nuclear volume (NV (µm3) = 4/3.π.r3, where r3 = diameter/2); and the circularity of the hepatocyte nucleus (CN = p2 ̸4.π.a, where p2 = nuclear perimeter and a = nuclear area) according to Rodrigues et al. (2017).
Aeromonas hydrophila challenge
The A. hydrophila strain (Strain KJ561021) used for the challenge was provided by the Laboratory of Microbiology and Parasitology of Aquatic Organisms at CAUNESP/Jaboticabal-SP. After sterilization of the culture medium (BHI, agar) in an autoclave at 121 °C for 20 min, the A. hydrophila strains were plated and incubated for 24 h in a bacteriological incubator at 28 °C. After the growth of bacterial colonies, the washing procedure was performed with phosphate-bufferid saline (PBS) in a centrifuge at 4000 g for 20 min, 3 times. Then the bacteria were resuspended in 20 ml PBS and the reading performed in a spectrophotometer (625 nm). The established concentration for the challenge was 0.8 × 106 CFU and 0.1 mL of the solution were inoculated intraperitoneally in each fish (n = 40 per treatment) according to Rodrigues et al. (2021).
Statistical analyses
Data were analysed for normality and homogeneity using the Shapiro–Wilk and Bartlett tests, respectively. Growth performance and histomorphometric data were submitted to analysis of variance (one-way ANOVA). A Two-way ANOVA was used on blood parameters with two independent variables in a 4 × 2 factorial scheme, (4 treatments × 2 collection times). The parameters immature leukocytes, monocytes, neutrophils, leukocytes, thrombocytes and respiratory activity of leukocytes were transformed into log and square root because they did not present the assumptions of normality and homogeneity. Means were compared by Duncan's test (p < 0.05). All tests were performed using the statistical program R version 3.4.3 (Packages ExpDes.pf).
Results
Zootechnical performance
As in the control group, supplementation had no effects on selection or rejection of feed. After 45 days of supplementation, survival was 100% in all treatments. The feed intake was significantly lower (p < 0.05) for fish in the probiotic group when compared to the other groups; however, feed intakes were equal between the control and synbiotic groups. The allomeric condition factors were significantly higher (p < 0.05) in fish from the paraprobiotic and synbiotic groups, while the lowest condition factor was found in fish from the probiotic group. In the other zootechnical indexes, no significant differences were found between the groups (Table 1).
Hemato-biochemical parameters
There were no interactions between the factors in the hematological analyses. Separately, no significant differences (p > 0.05) were observed in the blood count between the supplemented groups and the control group. However, significant differences (p < 0.05) were observed in the blood count between the pre-challenge and post-challenge periods. Hemoglobin, hematocrit, number of neutrophils, lymphocytes, leukocytes, immature leukocytes, monocytes and thrombocytes decreased significantly (p < 0.05) after exposure to A. hydrophila (Table 2). The other hematimetric indexes (MCH, MCV and MCHC) showed no significant difference (p > 0.05) neither between treatments nor between collection times (Fig. 1).

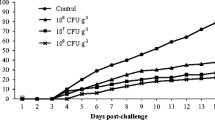
In (A) Hematimetric indexes of mean corpuscular hemoglobin concentration (MCHC), mean corpuscular hemoglobin (MCH) and mean corpuscular volume (MCV); in (B) plasma glucose levels; in (C) leukocyte respiratory activity (nitroblue tetrazolium (NBT) reduction) of dourado (S. brasiliensis) fed for 45 days with a diet containing 0.02% probiotic (L. rhamnosus), 2.0% paraprobiotic (inactive L. rhamnosus), synbiotic (0.02% probiotic + 2.0% paraprobiotic), and control group (diet without additive). In (D) survival rate of juvenile dourado (S. brasiliensis) up to 7 days after infection by A. hydrophila. Values are expressed as mean ± standard deviation. Different capital letters represent significant differences (p < 0.05) between the sampling periods (pre-and post-challenge) by Duncan's test
Blood glucose levels did not show significant differences (p > 0.05) between the treatments at the end of the supplementation period. However, glucose levels reduced significantly (p < 0.05) in the post-challenge period (Fig. 1). Likewise, the respiratory activity of leukocytes showed a significant increase (p < 0.05) only after exposure to A. hydrophila (Fig. 1). Survival rates decreased after bacterial infection, but no significant differences (p > 0.05) were observed between groups (Fig. 1).
Hepatic and intestinal histomorphometric analysis
Significant differences (p < 0.05) were observed in the hepatic and intestinal histomorphometric indexes. Fish in the paraprobiotic and synbiotic groups had higher total villus height and villus height when compared to fish in the probiotic group. Serosa were significantly (p < 0.05) thicker in the intestines of fish from the probiotic and synbiotic groups; however, the paraprobiotic and control group were the same as the synbiotic group. The number of goblet cells was significantly higher (p < 0.05) in fish from the synbiotics group.
Regarding liver tissue, the cytoplasmic areas of hepatocytes in fish from the paraprobiotic group were significantly larger (p < 0.05) than in the other groups. Cytoplasmic perimeters were significantly smaller (p > 0.05) in fish in the synbiotic group when compared to fish in the probiotic and paraprobiotic groups, and equal to the fish in the control group. Hepatocyte perimeter/nucleus/cytoplasm ratio was significantly higher (p < 0.05) in fish from the synbiotic group. The other variables showed no difference (p > 0.05) (Table 3).
Discussion
Nutrition based on functional diets has been shown to be fundamental for improvements in the aquaculture production sector, where the use of prebiotics, probiotics, paraprobiotics and synbiotics help to improve the health status of animals, the aquatic environment, and consequently reduce the incidence of disease outbreaks (Amenyogbe et al. 2020; El-Saadony et al. 2021).
Feed intake in fish can be regulated by several factors related to the rearing environment, and the better use of feed ensures better growth performance with less waste generation (Houlihan et al. 2001). Herein, although no significant differences were observed in weight gain and final weight, we found that dietary supplementation with L. rhamnosus in different forms (probiotic, paraprobiotic and synbiotic) were able to reduce feed intake in juvenile dourado S. brasiliensis, where the fish in the probiotic and paraprobiotic groups, even ingesting smaller amounts of feed, reached the same productive indexes as the fish in the control group. This benefit may have been achieved by the presence of probiotic bacteria in the intestine, which can improve the balance of the intestinal microbiota and improve the absorption and utilization of nutrients. (Amenyogbe et al. 2020; Naiel et al. 2021).
Likewise, the condition factor is seen as an important productive indicator (Owatari et al. 2022a) and in the paraprobiotic and synbiotic groups, the highest condition factor index can indicate the condition of the fish, which were in an accelerated growth phase, directing all the nutritional benefits of ingested feed for body building (Gomiero et al. 2010). Considering that the largest inputs resulting from fish farming are related to feed expenses (Castilho-Barros et al. 2020), we can infer that L. rhamnosus can be a viable alternative to generate savings in the productive sector.
Recently, Noshair et al. (2023) carried out an evaluation of the dietary supplementation of the probiotic L. rhamnosus on the growth performance of O. niloticus and found that the probiotic was able to significantly improve weight gain, protein efficiency rate, specific growth rate and condition factor in all probiotics-supplemented groups. On the occasion Noshair et al. (2023) conducted the experiment for 12 weeks, which brings us a huge suspicion that these indexes can also be achieved with dourado S. brasiliensis in trials with a longer duration.
Hematological assessments of fish in experimental aquaculture environments are usually related to diagnostics to decipher the health status of fish in response to nutritional changes or exposure to pathogens (Fazio 2019). Dawood et al. (2016) evaluated the effects of isolated or combined dietary supplementation of L. rhamnosus and L. lactis on the immune responses of sea bream Pagrus major and found that, compared to the control group, fish fed with probiotics in both forms, presented a significant increase in the hematocrit, guaranteeing a better state of health of the fish.
According to Satake et al. (2009) the hematocrit values of S. brasiliensis under experimental conditions can vary between 36 and 45% and despite that, in the present study the values remained below this reference, both before and after the bacterial infection. Likewise, reduced values for hematocrit were observed by Koch et al. (2018) in juvenile S. brasiliensis supplemented with different doses of vitamin A and exposed to A. hydrophila. The decrease in hematocrit was related to the reduction in erythrocytes after the bacterial challenge, but this reduction observed by the authors was not below the reference values (1.95 ~ 2.33 × 106 μL−1) described by Satake et al. (2009). In the present study the number of monocytes, immature leukocytes, thrombocytes and leukocytes also decreased, corroborating the findings of Claudiano et al. (2019) who observed a reduction in the number of leukocytes in Piaractus mesopotamicus between one hour and nine hours after infection by A. hydrophila.
Blood glucose and cortisol levels are used as good physiological indicators in response to stress in fish (Sopinka et al. 2016). In aquaculture, fish are often subjected to different stressors, including routine handling (Portz et al. 2006), which cause changes in plasma glucose concentrations and have a multifactorial impact on the organism (Sopinka et al. 2016; Bartoňková et al. 2017). Such glycemic changes occur within minutes, hours or even days after suffering an adverse stimulus (Langiano and Martínez 2009).
According to Sopinka et al. (2016) knowing the basal levels of plasma glucose of the species studied is of great relevance, as the values obtained establish parameters to be used as a reference in future research. In the present study, fish exposed to A. hydrophila showed different blood glucose levels before and after infection, remaining higher in the post-challenge period, regardless of treatment. Cortisol is one of the hyperglycemic hormones reported as the main responsible, during the period of infection, for deregulating glycemia (Johar et al. 2021). When a pathological agent invades the organism, it automatically activates the immune system, triggering an innate response that includes the action of phagocytic cells (neutrophils, monocytes and eosinophils) (Mahoney and Macnulty 1992; Iwama and Nakanishi 1996; Roberts 2012) causing hyperglycemia. This information explains the increase in glucose during the infectious process caused by A. hydrophila.
The increase of respiratory activity of leukocytes was observed by Román et al. (2012) in Sparus aurata and Dicentrarchus labrax after dietary supplementation with inactivated probiotic (paraprobiotic) Vagococcus fluvialis; and by Muñoz-Atienza et al. (2015) in Scophthalmus maximus fed a mix of viable and inactivated lactic acid bacteria. However, in the present study, no change in the respiratory activity of leukocytes related to dietary supplementation was observed. On the other hand, during the bacterial challenge, the respiratory activity of leukocytes was increased, suggesting a physiological response in fish related to the stress suffered. Reactive oxygen species (ROS) are normally produced during respiratory metabolism; however, production is controlled by an antioxidant defense system that inhibits and/or reduces damage caused by the action of free radicals (Biller and Takahashi 2018), establishing the respiratory activity of leukocytes as an indicator of the activation of the innate immune response after infection by pathogens (Biller et al. 2013).
The mechanisms of action of paraprobiotics are still not well understood, however, studies reinforce the ability of this additive to modulate the immune system (Almada et al. 2016; Barros et al. 2019) and inhibit the growth of pathogens through the adhesion in the intestinal epithelium (Grześkowiak et al. 2014). Such information is corroborated by the findings of the present study, where the paraprobiotic potentiated the effects of the probiotic on intestinal histomorphometry, and on the number of goblet cells.
Goblet cells play an important role in maintaining homeostasis and act as the front line of innate host defense (Kim and Ho 2010; Bevins and Salzman 2011). They are responsible for various digestive enzymes and mucus secretion, predominantly mucins, which act as a bactericidal barrier against pathogenic microorganisms (McGuckin et al. 2015). In view of the facts, the synbiotic (viable and inactive L. rhamnosus) provided intestinal immunomodulation, with an increase in the length of the villi, and consequent increase in the amount of goblet cells. On the other hand, the probiotic L. rhamnosus promoted a thicker serosa, which probably secrete more serous fluid, promoting greater organ integrity (Veggetti et al. 1999). The modulation of intestinal morphology can consequently contribute to the improvement of zootechnical performance, explaining the best performance indexes in feed intake and condition factor in the paraprobiotic and synbiotic groups.
Finally, regarding hepatocytes, liver tissue morphology can be used as biomarkers of nutritional responses, metabolic conditions and fish health (Rašković et al. 2011; Rodrigues et al. 2017), and alterations such as glycogen depletion, inflammation and neoplasms can be interpreted as a response to stress (Köhler 1992; Teh et al. 1997). Herein, the histomorphometric parameters of hepatocytes showed significant differences between treatments. According to Watanabe & Tanaka (1982) size of hepatocytes is linked to the volume of stored lipid droplets, and according to Schott et al. (2019) in hepatocytes, lipid droplets have diameters ranging from 60 nm to more than 5 µm and may play an important role in cellular catabolic processes. Thus, the presence of lipid in the liver should not be directly related to physiological anomalies or diseases (steatosis), as these droplets are common in up to 5% of liver cells (Scorletti and Carr 2022). However, increased lipid droplets within hepatocytes, constant and for a long time, can be caused by inflammation (Owatari et al. 2022b).
Conclusion
In conclusion, in the present study, the use of 0.02% of probiotic (L. rhamnosus 108 CFU) and 2.0% of paraprobiotic (inactive strains of L. rhamnosus 1010 CFU) proved to be a promising practice due to their action on productive performance, enabling weight gain similar to that of control group with lower feed intake, as well as improving the condition factor with 2.0% paraprobiotic and symbiotic (0.02% probiotic + 2.0% paraprobiotic). In addition to improving intestinal immunohistomorphometric parameters (goblet cells/villi), possibly promoting an improved condition in facing intestinal pathogens.
Data Availability
The data related to this research are available upon prior request.
References
Abd El-Hack ME, El-Saadony MT, Shafi ME, Qattan SYA, Batiha GE, Khafaga AF, Abdel-Moneim AME, Alagawany M (2020) Probiotics in poultry feed: A comprehensive review. J Anim Physiol Anim Nutr 104(6):1835–1850. https://doi.org/10.1111/jpn.13454
Akhter N, Wu B, Memon AM, Mohsin M (2015) Probiotics and prebiotics associated with aquaculture: a review. Fish Shellfish Immunol 45(2):733–741. https://doi.org/10.1016/j.fsi.2015.05.038
Almada CN, Almada CN, Martinez RC, Sant’Ana AS (2016) Paraprobiotics: Evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci Technol 58:96–114. https://doi.org/10.1016/j.tifs.2016.09.011
Amenyogbe E, Chen G, Wang Z, Huang J, Huang B, Li H (2020) The exploitation of probiotics, prebiotics and synbiotics in aquaculture: present study, limitations and future directions.: a review. Aquacult Int 28:1017–1041. https://doi.org/10.1007/s10499-020-00509-0
Arowolo MA, He J (2018) Use of probiotics and botanical extracts to improve ruminant production in the tropics: A review. Animal Nutrition 4(3):241–249. https://doi.org/10.1016/j.aninu.2018.04.010
Austin B, Austin DA (2016) Bacterial fish pathogens: disease of farmed and wild fish. Springer Cham, Springer International, Publishing Switzerland, 6th edition, p. 732. https://doi.org/10.1007/978-3-319-32674-0
Barros CP, Guimarães JT, Esmerino EA, Duarte MCKH, Silva MC, Silva R, Sant’Ana FBMAS, Freitas MQ, Cruz AG (2019) Paraprobiotics and prostbiotics: concepts and potential applications in dairy products. Currente Opinion in Food Science 32:1–8. https://doi.org/10.1016/j.cofs.2019.12.003
Bartoňková J, Hyršl P, Vojtek L (2017) Glucose determination in fish plasma by two different moderate methods. Acta Vet Brno 85(4):349–353. https://doi.org/10.2754/avb201685040349
Bevins CL, Salzman NH (2011) Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol 9(5):356–368. https://doi.org/10.1038/nrmicro2546
Biller JD, Takahashi LS (2018) Oxidative stress and fish immune system: phagocytosis and leukocyte respiratory burst activity. An Acad Bras Ciênc 90:3403–3414. https://doi.org/10.1590/0001-3765201820170730
Biller JD, Takahashi LS, Saita MV, Gimbo RY, Urbinati EC (2013) Leukocytes respiratory burst activity as indicator of innate immunity of pacu Piaractus mesopotamicus. Braz J Biol 73:425–429. https://doi.org/10.1590/s1519-69842013000200026
Braun N, Lima RL, Baldisserotto B, Dafre AL, Nuñer APO (2010) Growth, biochemical and physiological responses of Salminus brasiliensis with different stocking densities and handling. Aquaculture 301(1–4):22–30
Carraschi SP, Cruz C, Machado Neto JG, Castro MP, Bortoluzzi NL, Gírio ACF (2011) Efficacy of the florfenicol and of the oxytetracycline in the control in Aeromonas hydrophila in pacu (Piaractus mesopotamicus). Arquivo Brasileiro De Medicina Veterinária e Zootecnia 63:579–583. https://doi.org/10.1590/S0102-09352011000300007
Castilho-Barros L, Owatari MS, Mouriño JLP, Silva BC, Seiffert WQ (2020) Economic feasibility of tilapia culture in southern Brazil: A small-scale farm model. Aquaculture 515:734551. https://doi.org/10.1016/j.aquaculture.2019.734551
Choudhury TG, Kamilya D (2019) Paraprobiotics: An aquaculture perspective. Rev Aquac 11(4):1258–1270. https://doi.org/10.1111/raq.12290
Claudiano GS, Yunis-Aguinaga J, Marinho-Neto FA, Miranda RL, Martins IM, Otani FS, Mundim AV, Marzocchi-Machado CM, Moraes JRE, Moraes FR (2019) Hematological and immune changes in Piaractus mesopotamicus in the sepsis induced by Aeromonas hydrophila. Fish Shellfish Immunol 88:259–265. https://doi.org/10.1016/j.fsi.2019.01.044
Collier HB (1944) The standardization of blood haemoglobin determinations. Can Med Assoc J 50:550–552
Cuevas-González PF, Liceaga AM, Aguilar-Toalá JE (2020) Postbiotics and paraprobiotics: From concepts to applications. Food Res Int 136:109502. https://doi.org/10.1016/j.foodres.2020.109502
Dawood MA, Koshio S, Ishikawa M, Yokoyama S, El Basuini MF, Hossain MS, Nhu TH, Dossou S, Moss AS (2016) Effects of dietary supplementation of Lactobacillus rhamnosus or/and Lactococcus lactis on the growth, gut microbiota and immune responses of red sea bream, Pagrus major. Fish Shellfish Immunol 49:275–285. https://doi.org/10.1016/j.fsi.2015.12.047
Dawood MAO, Koshio S, Ishikawa M, El-Sabagh M, Yokoyama S, Wang WL, Yukun Z, Olivier A (2017) Physiological response, blood chemistry profile and mucus secretion of red sea bream (Pagrus major) fed diets supplemented with Lactobacillus rhamnosus under low salinity stress. Fish Physiol Biochem 43:179–192. https://doi.org/10.1007/s10695-016-0277-4
Della Flora MA, Maschke F, Ferreira CC, Pedron FDA (2010) Biologia e cultivo do dourado (Salminus brasiliensis). Acta Veterinaria Brasilica 4:7–14. https://doi.org/10.21708/avb.2010.4.1.1492
El-Saadony MT, Alagawany M, Patra AK, Kar I, Tiwari R, Dawood MAO, Dhama K, Abdel-Latif HMR (2021) The functionality of probiotics in aquaculture: An overview. Fish Shellfish Immunol 117:36–52. https://doi.org/10.1016/j.fsi.2021.07.007
Elumalai P, Kurian A, Lakshmi S, Faggio C, Esteban MA, Ringø E (2020) Herbal immunomodulators in aquaculture. Reviews in Fisheries Science & Aquaculture 29(1):33–57. https://doi.org/10.1080/23308249.2020.1779651
Fazio F (2019) Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 500:237–242. https://doi.org/10.1016/j.aquaculture.2018.10.030
Goldenfarb PB, Bowyer FP, Hall E, Brosious E (1971) Reproducibility in the hematology laboratory: the microhematocrit determinations. Am J Clin Pathol 56(1):35–39
Gomiero LM, Villares Junior GA, Braga FMDS (2010) Relação peso-comprimento e fator de condição de Oligosarcus hepsetus (Cuvier, 1829) no Parque Estadual da Serra do Mar-Núcleo Santa Virgínia, Mata Atlântica, estado de São Paulo, Brasil. Biota Neotrop 10:101–105. https://doi.org/10.1590/S1676-06032010000100009
Grześkowiak Ł, Collado MC, Beasley S, Salminen S (2014) Pathogen exclusion properties of canine probiotics are influenced by the growth media and physical treatments simulating industrial processes. J Appl Microbiol 116(5):1308–1314. https://doi.org/10.1111/jam.12477
Hoseini SM, Rajabiesterabadi H, Khalili M, Yousefi M, Hoseinifar SH, Van Doan H (2020) Antioxidant and immune responses of common carp (Cyprinus carpio) anesthetized by cineole: Effects of anesthetic concentration. Aquaculture 520:734680. https://doi.org/10.1016/j.aquaculture.2019.734680
Hoseinifar SH, Sun YZ, Wang A, Zhou Z (2018) Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front Microbiol 9:2429. https://doi.org/10.3389/fmicb.2018.02429
Houlihan D, Boujard T, Jobling M (eds) (2001) Food intake in fish. Blackwell Science, Oxford
Humason GL (1972) Animal Tissue Techniques. 3rd edn. San Francisco: W. H. Freeman and Company. p 641
Iwama G, Nakanishi T (1996) The Fish Immune System. Academic Press, San Diego
Johar H, Spieler D, Bidlingmaier M, Herder C, Rathmann W, Koenig W, Peters A, Kruse J, Ladwig KH (2021) Chronic inflammation mediates the association between cortisol and hyperglycemia: findings from the cross-sectional population-based KORA age study. J Clin Med 10(13):2751
Kim YS, Ho SB (2010) Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr Gastroenterol Rep 12(5):319–330. https://doi.org/10.1007/s11894-010-0131-2
Klein JB, Payne V, Schepers TM, McLeish KR (1990) Bacterial lipopolysaccharide enhances polymorphonuclear leukocyte function independent of changes in intracellular calcium. Inflammation 14:599–611. https://doi.org/10.1007/BF00914279
Koch JFA, Sabioni RE, Aguilar AA, Lorenz EK, Cyrino JEP (2018) Vitamin A requirements of dourado (Salminus brasiliensis): Growth performance and immunological parameters. Aquaculture 491:86–93. https://doi.org/10.1016/j.aquaculture.2018.03.017
Köhler TP (1992) Molecular and Biochemical Responses of Fish to Stressors and Their Potential Use in Environmental Monitoring. Am Fish Soc Symp 8:9–28
Langiano VC, Martínez CBR (2009) Toxicity and effects of a glyphosphate-based herbicide on the neotropical fish Prochilodus lineatus. Comparative Biochemistry and Physiology Part C 147(2):22–231
Lima-Junior, SE, Cardone, IB, Goitein, R (2002) Determination of a method for calculation of Allometric Condition Factor of fish. Acta Scientiarum: Biological and Health Sciences, 24(2), 397–400. Available at: http://hdl.handle.net/11449/67090. Accessed Nov 2023
Lorenz EK, Sabioni RE, Volkoff H, Cyrino JEP (2022) Growth performance, health, and gene expression of appetite-regulating hormones in Dourado Salminus brasiliensis, fed vegetable-based diets supplemented with swine liver hydrolysate. Aquaculture 548:737640. https://doi.org/10.1016/j.aquaculture.2021.737640
Mahesh MS, Mohanta RK, Patra AK (2021) Probiotics in livestock and poultry nutrition and health. Adv Probiotics Sustain Food Med, 149–179. https://doi.org/10.1007/978-981-15-6795-7_7
Mahoney JB, Macnulty JK (1992) Diseases- associated blood changes and normal seasonal hematological variation in winter flounder in the Hudson- Raritan estuary. Trans Am Fish Soc 121:261–268
McGuckin MA, Thornton DJ, Whitsett JA (2015) Mucins and mucus. In Mucosal Immunology (pp. 231–250). Academic Press. https://doi.org/10.1016/B978-0-12-415847-4.00014-8
Muñoz-Atienza E, Araújo C, Lluch N, Hernández PE, Herranz C, Cintas LM, Magadán S (2015) Different impact of heat-inactivated and viable lactic acid bacteria of aquatic origin on turbot (Scophthalmus maximus L.) head-kidney leucocytes. Fish Shellfish Immunol 44:214–223. https://doi.org/10.1016/j.fsi.2015.02.021
Naiel MA, Farag MR, Gewida AG, Elnakeeb MA, Amer MS, Alagawany M (2021) Using lactic acid bacteria as an immunostimulants in cultured shrimp with special reference to Lactobacillus spp. Aquacult Int 29:219–231. https://doi.org/10.1007/s10499-020-00620-2
Nakandakare IB, IwashitaMKP D, de Carla DIAS, Tachibana L, Ranzani-PaivaMJT RE (2018) Incorporação de probióticos na dieta para juvenis de tilapias-do-Nilo: parâmetros hematológicos, imunológicos e microbiológicos. Bol Inst Pesca 39(2):121–135
Navarrete P, Tovar-Ramírez D (2014) Use of yeasts as probiotics in fish aquaculture. Sustainable Aquaculture Techniques 1:135–172. https://doi.org/10.5772/57196
Nikoskelainen S, Ouwehand AC, Bylund G, Salminen S, Lilius EM (2003) Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol 15(5):443–452. https://doi.org/10.1016/S1050-4648(03)00023-8
Noshair I, Kanwal Z, Jabeen G, Arshad M, Yunus FUN, Hafeez R, Mairaj R, Haider I, Ahmad M, Alomar SY (2023) Assessment of Dietary Supplementation of Lactobacillus rhamnosus Probiotic on Growth Performance and Disease Resistance in Oreochromis niloticus. Microorganisms 11(6):1423. https://doi.org/10.3390/microorganisms11061423
Nunes AL, Owatari MS, Rodrigues RA, Fantini LE, Kasai RYD, Martins ML, Mouriño JPL, Campos CM (2020) Effects of Bacillus subtilis C-3102-supplemented diet on growth, non-specific immunity, intestinal morphometry and resistance of hybrid juvenile Pseudoplatystoma sp. challenged with Aeromonas hydrophila. Aquacult Int 28:2345–2361. https://doi.org/10.1007/s10499-020-00586-1
Oboh A (2022) Diversification of farmed fish species: A means to increase aquaculture production in Nigeria. Rev Aquac 14(4):2089–2098. https://doi.org/10.1111/raq.12690
Oliveira FC, Kasai RYD, Fernandes CE, da Silva WS, Campos CM (2022) Probiotic, prebiotic and synbiotics supplementation on growth performance and intestinal histomorphometry Pseudoplatystoma reticulatum larvae. J Appl Aquac 34(2):279–293. https://doi.org/10.1080/10454438.2020.1841060
Owatari MS, Silva LRD, Ferreira GB, Rodhermel JCB, Andrade JIAD, Dartora A, Jatobá A (2022a) Body yield, growth performance, and haematological evaluation of Nile tilapia fed a diet supplemented with Saccharomyces cerevisiae. Anim Feed Sci Technol 293:115453. https://doi.org/10.1016/j.anifeedsci.2022.115453
Owatari MS, Cardoso L, Pereira SA, Pereira UDP, Tachibana L, Martins ML, Mouriño JLP (2022b) Laboratory-controlled challenges of streptococcosis in Nile tilapia using the oral route (infected-feed) for infection. Fish Shellfish Immunol 120:295–303. https://doi.org/10.1016/j.fsi.2021.11.039
PeixeBR - Brazilian Association of Pisciculture (2023) Brazilian Pisciculture Yearbook. Edition 2023. São Paulo, Brazil, p 65. Available at: https://www.peixebr.com.br/anuario/. Accessed 20 Nov 2023
Pfeiler EA, Klaenhammer TR (2007) The genomics of lactic acid bacteria. Trends Microbiol 15(12):546–553. https://doi.org/10.1016/j.tim.2007.09.010
Piqué N, Berlanga M, Miñana-Galbis D (2019) Health benefits of heat-killed (Tyndallized) probiotics: an overview. Int J Mol Sci 20(10):2534. https://doi.org/10.3390/ijms20102534
Pirarat N, Pinpimai K, Endo M, Katagiri T, Ponpornpisit A, Chansue N, Maita M (2011) Modulation of intestinal morphology and immunity in nile tilapia (Oreochromis niloticus) by Lactobacillus rhamnosus GG. Res Vet Sci 91(3):e92–e97. https://doi.org/10.1016/j.rvsc.2011.02.014
Portz DE, Woodley CM, Cech JJ (2006) Stress-associated impacts of short-term holding on fishes. Rev Fish Biol Fisheries 16:125–170. https://doi.org/10.1007/s11160-006-9012-z
Raabis S, Li W, Cersosimo L (2019) Effects and immune responses of probiotic treatment in ruminants. Vet Immunol Immunopathol 208:58–66. https://doi.org/10.1016/j.vetimm.2018.12.006
Rašković B, Stanković M, Marković Z, Poleksić V (2011) Histological methods in the assessment of different feed effects on liver and intestine of fish. Journal of Agricultural Sciences (Belgrade) 56(1):87–100. https://doi.org/10.2298/JAS1101087R
Roberts RJ (2012) Fish pathology. 4th edition, John Wiley & Sons. p. 592. https://doi.org/10.1002/9781118222942
Rodrigues RA, Saturnino KC, Fernandes CE (2017) Liver histology and histomorphometry in hybrid sorubim (Pseudoplatystoma reticulatum× Pseudoplatystoma corruscans) reared on intensive fish farming. Aquac Res 48(9):5083–5093. https://doi.org/10.1111/are.13325
Rodrigues TAR, Owatari MS, Veiga PTDN, Povh JA, Kasai RYD, Pilarski F, Fernandes CES, Campos CM (2021) Bacillus subtilis improves non-specific immunity and survival of Pseudoplatystoma reticulatum challenged with Aeromonas hydrophila during the feeding training phase. Aquac Res 52(5):2348–2352. https://doi.org/10.1111/are.15055
Román L, Real F, Sorroza L, Padilla D, Acosta B, Grasso V, Bravo J, Acosta F (2012) The in vitro effect of probiotic Vagococcus fluvialis on the innate immune parameters of Sparus aurata and Dicentrarchus labrax. Fish Shellfish Immunol 33(5):1071–1075. https://doi.org/10.1016/j.fsi.2012.06.028
Ruiz ML, Owatari MS, Yamashita MM, Ferrarezi JVS, Garcia P, Cardoso L, Martins ML, Mouriño JLP (2020) Histological effects on the kidney, spleen, and liver of Nile tilapia Oreochromis niloticus fed different concentrations of probiotic Lactobacillus plantarum. Trop Anim Health Prod 52:167–176. https://doi.org/10.1007/s11250-019-02001-1
Saint-Paul U (2017) Native fish species boosting Brazilian’s aquaculture development. Acta of Fisheries and Aquatic Resources 5(1):1–9. https://doi.org/10.2312/Actafish.2017.5.1.1-9
Satake F, Ishikawa MM, Hisano H, De Pádua SB, Tavares-Dias M (2009) Boletim de Pesquisa e Desenvolvimento: Relação peso-comprimento, fator de condição e parâmentros hematológicos de dourado Salminus brasiliensis cultivado em condições experimentais. EMBRAPA, p 22. Available at: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/574003/1/BP200951.pdf. Accessed Nov 2023
Schott MB, Weller SG, Schulze RJ, Krueger EW, Drizyte-Miller K, Casey CA, McNiven MA (2019) Lipid droplet size directs lipolysis and lipophagy catabolism in hepatocytes. J Cell Biol 218(10):3320–3335. https://doi.org/10.1083/jcb.201803153
Scorletti E, Carr RM (2022) A new perspective on NAFLD: Focusing on lipid droplets. J Hepatol 76(4):934–945. https://doi.org/10.1016/j.jhep.2021.11.009
Semwal A, Kumar A, Kumar N (2023) A review on pathogenicity of Aeromonas hydrophila and their mitigation through medicinal herbs in aquaculture. Heliyon. https://doi.org/10.1016/j.heliyon.2023.e14088
Soomro RN, Abd El-Hack ME, Shah SS, Taha AE, Alagawany M, Swelum AA, Hussein EOS, Ba-Aawdh HA, Saadeldin I, El-Edel MA, Tufarelli V (2019) Impact of restricting feed and probiotic supplementation on growth performance, mortality and carcass traits of meat-type quails. Anim Sci J 90(10):1388–1395. https://doi.org/10.1111/asj.13290
Sopinka NM, Donaldson MR, O’Connor CM, Suski CD, Cooke SJ (2016) Stress indicators in fish. In Fish physiology (Vol. 35, pp. 405–462). Academic Press. https://doi.org/10.1016/B978-0-12-802728-8.00011-4
Tamang JP (2014). Biochemical and Modern Identification Techniques: Microfloras of Fermented Foods. https://doi.org/10.1016/B978-0-12-384730-0.00038-0
Tavares-Dias M, Moraes FR (2004) Hematologia de peixes teleósteos Ribeirão Preto. São Paulo, p 144
Tavares-Dias M, Martins ML (2017) An overall estimation of losses caused by diseases in the Brazilian fish farms. J Parasit Dis 41:913–918. https://doi.org/10.1007/s12639-017-0938-y
Taverniti V, Guglielmetti S (2011) The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: proposal of paraprobiotic concept). Genes Nutr 6(3):261–274. https://doi.org/10.1007/s12263-011-0218-x
Teh SJ, Adams SM, Hinton DE (1997) Histopathologic biomarkers in feral freshwater fish populations exposed to different types of contaminant stress. Aquat Toxicol 37(1):51–70. https://doi.org/10.1016/S0166-445X(96)00808-9
Valenti WC, Barros HP, Moraes-Valenti P, Bueno GW, Cavalli RO (2021) Aquaculture in Brazil: past, present and future. Aquac Reports 19:100611. https://doi.org/10.1016/j.aqrep.2021.100611
Vallejos-Vidal E, Reyes-López F, Teles M, MacKenzie S (2016) The response of fish to immunostimulant diets. Fish Shellfish Immunol 56:34–69. https://doi.org/10.1016/j.fsi.2016.06.028
Veggetti A, Rowlerson A, Radaelli G, Arrighi S, Domeneghini C (1999) Post-hatching development of the gut and lateral muscle in the sole. J Fish Biol 55(AS):44–65. https://doi.org/10.1111/j.1095-8649.1999.tb01045.x
Watanabe T, Tanaka Y (1982) Age-related alterations in the size of human hepatocytes: A study of mononuclear and binucieate cells. Virchows Archiv B 39:9–20. https://doi.org/10.1007/BF02892832
Wintrobe MM (1934) Variations in the size and hemoglobin content of erythrocytes in the blood of various vertebrates. Folia Haematol 51(32):32–49
Xia Y, Lu M, Chen G, Cao J, Gao F, Wang M, Liu Z, Zhang D, Zhu H, Yi M (2018) Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia. Oreochromis Niloticus Fish Shellfish Immunol 76:368–379. https://doi.org/10.1016/j.fsi.2018.03.020
Acknowledgements
The authors thank the Coordination for the Improvement of Higher Education Personnel (CAPES) for a Master’s scholarship to the first author and GABBIA Biotecnologia® for donating the additive. Projeto Pacu Aquicultura Ltda. is also acknowledged for donating the fish.
Funding
The Foundation for the Development of Education, Science and Technology of the State of Mato Grosso do Sul / FUNDECT-MS provide the Research Fund [Proc. nº 71/044.568/2022; Grant nº 884, SIAFEM 32421].
Author information
Authors and Affiliations
Contributions
F.C.O. Experimental execution, Writing – original draft, Methodology. P.R.D.A. Experimental execution. R.G.D.S.A. Experimental execution. M.S.O. Data curation, Writing – original draft, final writing. F.P. Experimental execution, Methodology, bacterial challenge strain. C.E.D.S.F. Experimental execution; C.M.D.C. Conceptualization, Methodology, Project administration, Resources and Supervision.
Corresponding author
Ethics declarations
Ethical Approval
The study was approved by the Animal Ethics Committee (CEUA) of the State University of Mato Grosso do Sul – UEMS, Aquidauana, MS, Brazil, under Protocol Nº 034/2022 and followed all ethical principles in animal experimentation. All fish used in biological analyzes were previously anesthetized with Eugenol (50 mg L−1) and euthanized by spinal cord transection.
Competing interests
The authors declare there are no competing interests.
Additional information
Handling Editor: Brian Austin
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Highlights
• Feed intake of Salminus brasiliensis was significantly lower in the probiotic group.
• Allomeric condition factors of S. brasiliensis were significantly higher in paraprobiotic and synbiotic groups.
• Symbiotic and probiotic groups showed intestinal improvements.
• Goblet cells were significantly higher in S. brasiliensis from synbiotic group.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Oliveira, F.C., de Almeida, P.R., dos Santos Almeida, R.G. et al. Lactobacillus rhamnosus improves feed intake, condition factors, hepatic and intestinal histomorphometric indexes of dourado Salminus brasiliensis. Aquacult Int 32, 4757–4772 (2024). https://doi.org/10.1007/s10499-024-01400-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-024-01400-y