
Abstract
With a generally warming global climate, the number of Tetranychus truncatus specimens in the Hexi region in China has been increasing. As ectotherms, the growth and development of T. truncatus are greatly affected by changes in environmental temperature. The effect of heatwaves on organisms depends on a delicate balance between damage and repair periods. Therefore, we simulated nine patterns of periodically recurring changes in the frequency of high-temperature days using an intraday gradual temperature change model to study and compare the effects on the development and reproduction of pyridaben-sensitive and -resistant strains of T. truncatus. The results showed that the influence of the frequency of high-temperature days on developmental stages, longevity and fecundity was different between the two strains. The egg and immature stages of the sensitive strain were all affected by hot days, whereas the adult stage was less affected by the frequency. The egg stage of the resistant strain was less affected; it was mainly affected in the immature and adult stages. Under the moderate condition of increasing the proportion of days at normal temperature, the longevity of the resistant strain gradually increased and reached a maximum at a 1:3 frequency, and then it decreased with the increase in high-temperature days. The longevity of the sensitive strain was less affected by frequency, and there was no significant difference between most treatment and control groups. In addition, both sensitive and resistant strains were able to complete growth and development under all nine frequencies of high-temperature days, but the reproductive rate was lower than it was at normal temperatures, indicating that both strains of T. truncatus adapted to high temperatures at the expense of reduced reproduction rates. This lays a key theoretical foundation for predicting the occurrence of agricultural pest populations under the background of climate warming and developing appropriate control strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Phytophagous mites comprise important harmful species that affect the production of many crops in greenhouses and fields worldwide. Tetranychus truncatus Ehara (Acari: Tetranychidae) is a harmful spider mite that infests various agricultural field and greenhouse crops, and its distribution is mostly limited to Asian countries, including Guam, Marianas, China, Indonesia, the Philippines, Taiwan, Bangladesh, Thailand, Vietnam, Iran, Japan, and Korea (Ullah et al. 2014; Alain and Franck 2019).
Tetranychus truncatus has a complex diet and has caused serious infestations on multiple plants, including soybean, corn (maize), and other food crops and vegetables; it even harms jujube, apple, and other fruit trees (Jin et al. 2018). Especially in recent years, T. truncatus has become increasingly common in maize-producing areas in Northern China, and its damage rate has reached 10–30% (Zheng et al. 2016; Wang et al. 2021). Thus, the control of T. truncatus and other phytophagous mites is mostly dependent on frequent acaricide applications. However, the high acaricide selection pressure on T. truncatus has led to the development of resistance to several commercially available acaricides (Thomas et al. 2010, 2014; Fariborz et al. 2019; Ma et al. 2020; Rimy et al. 2021). With the warming of the global climate, an increase in the frequency of herbivore attacks was recorded recently, and pest pressure is expected to increase in agriculture and forestry wrought by climate change (Pham and Hwang 2020). For example, the period during which T. truncatus can harm crops has been prolonged, and T. truncatus has even caused crop failure in some areas. However, higher temperatures or hot days also present major challenges for arthropod herbivores (Zhang et al. 2021). Under unfavorable environmental conditions, arthropods have undergone complex changes in physiological, biochemical (Karl et al. 2008, 2011; Zhang et al. 2016), and molecular repair mechanisms to regulate their individual growth, development, reproduction, and even stress tolerance to counteract the negative effects of high temperatures (Guillermo et al. 2011; Kuroki et al. 2019; Liu et al. 2021).
Many studies have focused on the effects of temperature fluctuations on the growth and development of spider mites and other arthropods (Bayu et al. 2017; Rismayani et al. 2021; Ullah and Lim 2015), and also studied the effects on animals of daily temperature range (DTR), the range between the highest and lowest temperatures (Tmax – Tmin) and the average daily temperature (Chen et al. 2015). However, the effect of heatwaves on organisms should depend on the delicate balance between damage and repair periods, such as the mutual variation in the maximum daily temperature and the nightly minimum temperature, as well as the interval between short periods of extremely hot days and normal days (Zhang et al. 2015). This will help us to understand and compare the potential implications of changing patterns in higher temperature events on arthropods due to global climate change (Ma et al. 2018).
The Hexi region of the Gansu province (Zhangye, Wuwei, and other cities) is located in the arid area of Northwest China. In recent years, the hybrid corn seed production industry has developed rapidly. The seed production area is about 1.5 × 109 m2, accounting for 60% of the total domestic output, and has become the largest hybrid corn seed production base in China (Yin et al. 2022). The maximum temperature of cropland in summer is 35–40 °C during the day and about 18–20 °C at night. The suitably low temperature at night counteracts the adverse effects of high temperature stress on mites, resulting in an increase in T. truncatus in this area; T. truncatus has become the main pest affecting corn. The mites have also become resistant to many acaricides, making control more difficult (Chen et al. 2014; Chang and Sun 2021).
Zhang et al. (2021) revealed the negative effect of high temperature on arthropod-bacterial symbiosis, and the decrease of bacterial symbiosis led to the increase of its sensitivity to pesticides. Is there any other effect of high temperature variations on the resistance of arthropods to pesticides? Given the potential for changes in both resistance competence and life-history traits when temperatures fluctuate around different means, which could lead to altered population dynamics, we characterized responses by pyridaben-sensitive and -resistant strains of T. truncatus from the Hexi region in China to various frequencies of high-temperature days in variable temperature modes. These data provide a necessary basis for exploring the adaptability of the two strains of T. truncatus to high temperatures, predicting the occurrence of agricultural pest populations under the background of climate warming, and developing appropriate control strategies.
Materials and methods
Mite source and mite strain isolation
Tetranychus truncatus were collected from a corn field that had been maintained without any pesticide application in a suburb of Zhangye (38.93°N, 100.45°E), Gansu, China, in August 2018. We identified the mites based on a combination of morphology and molecular technology (DNA sequences) described in Matsuda et al. (2013) and Xin (1988). The mites were reared on young corn leaves in a controlled climatic chamber leaves at 25 ± 0.5 °C, 70 ± 5% RH, and L16:D8 photoperiod.
After being reared under these conditions for 2 years, these T. truncatus were considered a sensitive strain (SS). Some of the sensitive strains were then isolated and cultured with 15% pyridaben EC (Jiangsu Lanfeng Biochemical, Xuzhou, China), and the concentration of acaricide remained unchanged at the dose that killed 70% of the population. After a year of administration, the population was tested using a toxicity assay, and the resistance index (RI) was calculated by comparing with the sensitive strain as follows: RI = LC50 for the cultured population/LC50 for the susceptible strain. RI was classified as follows: RI < 5 indicated susceptibility/low resistance, 5 < RR < 10 indicated moderate resistance, and RI > 10 indicated high resistance (World Health Organization 2020; Hafez and Abbas 2021).
The pyridaben toxicity to T. truncatus is shown in Table 1. The results show that the resistance index reached 19.15, a more than 10-fold increase. This confirms that the Py-R strain may be regarded as a highly resistant strain.
Temperature regimes
In order to test how the diurnal variation in temperature and different high-temperature days affected the growth, development, and fecundity of T. truncatus, the design of the intervals of hot days (1–3 days) and normal days (1–3 days) was based on the method of Ma et al. (2018). There were three groups of high-temperature-day frequencies: (1) heatwaves with different successive hot days but the same number of normal days (Fig. 1, columns 1, 2, and 3); (2) heatwaves with the same successive hot days but a different number of normal days (Fig. 1, rows 1, 2, and 3); and (3) heatwaves with the same number of hot and normal days but differing in the absolute number of hot and normal days (Fig. 1, top-left to bottom-right diagonal).

The alternation patterns of successive hot and normal days. Different panels show different combinations of continuous hot and normal days separating high-temperature periods: (1) heatwaves with different successive hot days but the same number of normal days (columns 1, 2, and 3), (2) heatwaves with the same successive hot days but a different number of normal days (rows 1, 2, and 3), and (3) heatwaves with the same number of hot and normal days but differing in the absolute number of hot and normal days (top-left to bottom-right diagonal)
To simulate diurnal fluctuations in temperature in heatwaves, daily mean temperature and daily maximum and minimum temperature in the summer records for August in the Hexi region (100°26′E, 38°55′N) from 2014 to 2020 were downloaded from the China Meteorological Data Sharing Service System (http://data.cma.cn/). The mites are most common in August in corn-producing areas of China. In our study regions, heatwaves during that period usually last for 1–3 days with an interval of 1–3 days of normal temperatures between heatwaves. Mites were exposed under this cyclic alternating pattern until they died.
According to the data, the daily maximum and minimum temperatures were set to 38 and 18 °C, which represents a hot day. The maximum and minimum daily temperatures for normal days were set to 25 and 18 °C. A sinusoidal curve was used to simulate hourly temperature changes during the hot weather and normal weather. These daily temperature changes of 18–38 and 18–25 °C were mimicked in two growth chambers (RXZ-280B; Jiangnan, Ningbo, China), and temperature data were displayed on a screen in these chambers.
Experimental protocol
We estimated the response of pyridaben-sensitive and -resistant strains of T. truncatus at different instars to various types of heatwaves in terms of the development time, longevity, and fecundity. Through a preliminary high-temperature test, it was found that some larvae had developed to the nymph stage before 3 days of high-temperature treatment. Therefore, in this experiment, instead of inspecting individual stages, the larval and nymph stages were combined and considered the immature stage. The sensitive and resistant strains were able to survive in this mild high-temperature mode. Almost all individuals were able to reach their physiological life span at a high temperature. Thus, the survival rate is not shown in the results. In the control group (CK), each mite was only placed in the climatic chamber that simulated the diurnal temperature fluctuation of normal days (18–25 °C), and no high-temperature frequency treatment was performed.
Egg stage treatment
A mature female mite was placed on each same-sized corn leaf in a Petri dish (3 cm diameter, a cover with drilled air holes, with the gap between the dish and cover sealed with tape to prevent the mite from escaping) to lay eggs for 12 h and was then removed. All eggs except one were also removed; the Petri dishes with one egg each were then transferred to the climatic chambers for treatment. According to this method, 20 Petri dish with eggs were placed in a chamber that simulated the diurnal temperature fluctuations of hot and normal days. Each treatment was repeated 3×, totaling 60 Petri dishes.
Immature stage treatment
The rate of egg hatching, development time, and survival rate from the larval to nymph stage were checked once a day at 20:00 h. Different instars were identified by their exuviae.
Adult stage treatment
Mating was performed after mites entered the adult stage, and the reproduction of adults was then recorded once a day. New eggs were removed after each observation. The experiment ended after all tested individuals had died.
Statistical analysis
All statistical tests were performed using IBM-SPSS v.22 software (IBM, Armonk, NY, USA). For longevity, fecundity, and egg-to-adult development time, we used a multivariate analysis of variance (MANOVA) to test the effects of frequencies, strains, and frequencies × strains. Subsequently we compared life-history traits affected by different frequencies in the same strain using one-way ANOVA and Tukey’s post hoc tests (α = 0.05). The differences of development duration, longevity and fecundity between the two strains were compared by independent-sample t test. Prior to the MANOVA, Box’s M test was used to test whether the variance-covariance matrix of the dependent variable of each observation group is equal. The Shapiro–Wilk method was used to test whether the three dependent variables of egg stage, immature stage and adult stage were normally distributed (p > 0.05), and Levene’s method was used to test whether the variance of cause variables in each group were equal (p > 0.05). Graphs were plotted using OriginPro 2018 (OriginLab, Northampton, MA, USA).
Results
Impact of high-temperature day frequency on developmental stages
The Shapiro–Wilk test showed that the three dependent variables (egg, immature and adult stage) were normally distributed (p > 0.05); Levene’s test showed that the variances of cause variables in each group were equal (p > 0.05). There were significant interaction effects of frequency and strain on the developmental stages of T. truncatus (MANOVA: F3,27 = 107.108, p < 0.05; Wilk’s Λ = 0.001; partial η2 = 0.959).
The interaction between frequency and strain had a significant effect on all stages (egg: F = 217.742, p < 0.05, partial η2 = 0.980; immature: F = 80.097, p < 0.05, partial η2 = 0.947; adult: F = 266.819, p < 0.05, partial η2 = 0.984).
Pairwise comparisons
The growth of sensitive and resistant strains differed significantly at all stages (Fig. 2). Compared with the normal temperature, the resistant strain showed little change, but the sensitive strain showed a strong response: the incubation time of the eggs was prolonged after experiencing any of the various frequencies of high-temperature days. Development from larva to nymph of the resistant strain was affected by frequency, its development stage increased, and the significant difference between the resistant strain and the sensitive strain decreased. In the adult stage, due to the influence of frequency, there were significant differences in the survival time between the two strains. The adult stage of resistant strains was prolonged at the 1:3 and 2:2 frequencies, whereas that of sensitive strains was shortened. Under the frequency of the maximum high-temperature days and the minimum normal-temperature days (3:1), the adult period of the two strains was the shortest.

Mean (± SE) developmental time (days) of three stages of a pyridaben-sensitive strain (SS) and a pyridaben-resistant strain (Py-R) of Tetranychus truncatus at different high-temperature day frequencies. ‘Hot day : normal day’ indicates the number of consecutive hot days between consecutive normal days. Asterisks indicate significant pairwise differences (independent-sample t test: *p < 0.05)
Multiple comparisons
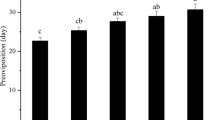
The developmental times of the egg, immature (larva to nymph), and adult stages of the sensitive and resistant strains were significantly affected by different frequencies of high-temperature days (one-way ANOVA; egg, SS strain: F9,20 = 345.793, Py-R strain: F9,20 = 41.216; immature, SS strain: F9,20 = 111.645, Py-R strain: F9,20 = 329.460; adult, SS strain: F9,20 = 580.231, Py-R strain: F9,20 = 470.404, all p < 0.01) (Fig. 3).

Mean (± SE) developmental time (days) of three stages of a pyridaben-sensitive strain (SS) and a pyridaben-resistant strain (Py-R) of Tetranychus truncatus in nine high-temperature day frequencies. ‘Hot day : normal day’ indicates the number of consecutive hot days between consecutive normal days. Means within a stage and within a strain capped with a different letter are significantly different (Tukey’s test: p < 0.05)
The influence of frequency on the developmental duration of the resistant strain at the egg stage was significantly lower than that at other stages (Fig. 3). Under the 1:2, 1:3, and 2:2 frequencies of high-temperature days, the development times of the immature and adult stages of the resistant strain were significantly higher than those of the control. The sensitive strain did not show this in the adult stage, but in the immature and egg stages, the development times under the influence of all nine frequencies were significantly higher than those of the control. The immature stage of the sensitive strain had the longest development time at the frequencies of 1:1 and 1:2, and the development time of the egg stage was longest at the frequencies of 1:2 and 1:3.
Impact of high-temperature frequency on longevity and fecundity
The Shapiro–Wilk test showed that the two dependent variables (longevity and fecundity) were normally distributed (p > 0.05); Box’s M test showed that the variance–covariance matrix of the two dependent variables in each group of independent variables was equal (p = 0.85); Levene’s test showed that the variances of cause variables in each group were equal (p > 0.05).
The interaction between the frequencies of high-temperature days and the strain had a significant effect on the dependent variables (F2,18 = 12.921, p < 0.05; Wilk’s Λ = 0.063; partial η2 = 0.749); that is, the influence of the frequency of high-temperature days on longevity and fecundity was different between the two strains. As can be seen in Table 2, the interaction between frequency and strain had a significant effect on the longevity and fecundity of T. truncatus.
Main effect of frequency and strains
The main effect of frequency on longevity and fecundity was significant (F2,18 = 73.137, p < 0.01; Wilk’s Λ = 0.003; partial η2 = 0.944). The effect of the strains was also significant (F2,18 = 44.426, p < 0.01; Wilk’s Λ = 0.305; partial η2 = 0.695).
Pairwise comparisons
The pairwise comparison results of the average longevity and fecundity of the two strains under different frequencies are shown in Fig. 4.
Except for 1:2 and 3:1 frequencies, the influence of the other high-temperature day frequencies on longevity showed highly significant differences (1:1, mean difference (MD) = 6.718, p < 0.01; 1:3, MD = 2.822, p < 0.05; 2:1, MD = 5.814, p < 0.01; 2:2, MD = 4.873, p < 0.01; 2:3, MD = 3.956, p < 0.05; 3:2, MD = 6.815, p < 0.01; 3:3, MD = 3.012, p < 0.05). Figure 4 shows that the longevity of resistant strains changed greatly, and was significantly different from that of sensitive strains under the influence of various frequencies of high-temperature days. For example, at the frequency of 1:3, the longevity of the resistant strain increased rapidly and was significantly higher than that of the sensitive strain (at room temperature, the longevity of the sensitive strain was significantly higher than that of the resistant strain).
The influence of the three high-temperature day frequencies on fecundity showed large differences at 1:1, 1:3, 2:1, 2:2, and 2:3 frequencies (1:1, MD = 18.103, p < 0.01; 1:3, MD = 22.902, p < 0.01; 2:1, MD = 4.794, p < 0.05; 2:2, MD = 8.374, p < 0.01; 2:3, MD = 4.274, p < 0.05). Under the influence of different high-temperature day frequencies, the fecundities of the sensitive and resistant strains experienced a decrease compared with normal-temperature days. There was no significant difference of fecundity between sensitivity vs. resistance due to 3 days of high temperature and various days of normal temperature interval (3:1, 3:2, and 3:3). However, the fecundity of resistant strains was significantly higher than that of sensitive strains with different high-temperature days and a maximum interval of normal-temperature days (1:3 and 2:3).

Mean (± SE) longevity (days) and fecundity (no. eggs/female) of a pyridaben-sensitive strain (SS) and a pyridaben-resistant strain (Py-R) of Tetranychus truncatus in nine high-temperature day frequencies. ‘Hot day : normal day’ indicates the number of consecutive hot days between consecutive normal days Asterisks indicate significant pairwise differences between strains (independent-sample t test: *0.01 < p < 0.05, **p < 0.01)
Multiple comparisons
The longevity and fecundity of the sensitive (SS) and resistant (Py-R) strain were significantly affected by different high-temperature day frequencies (one-way ANOVA; longevity, SS strain: F9,20 = 9.185, Py-R strain: F9,20 = 23.970; fecundity, SS strain: F9,20 = 178.536, F9,20 = 106.674, all p < 0.01) (Fig. 5). Under different high-temperature day frequency treatments, the fecundity of the sensitive strain was lower than at normal temperatures, but longevity was little influenced. Except for the significant increase or decrease at the 2:2 and 3:1 frequencies, there was no significant difference between the longevity of the other frequency treatments and the control.
The fecundity of the resistant strain was also obviously lower than that of the control, but the fecundity was significantly higher than that of the control at the 1:3 frequency. In addition, unlike the sensitive strain, the longevity of the resistant strain was significantly affected by high temperatures. Under the moderate condition of increasing the proportion of days at normal temperature, the longevity of the resistant strain gradually increased and reached a maximum at 1:3 frequency, which was significantly higher than that of the control (normal temperature). Then, the longevity of the resistant strain decreased with the increase in high-temperature days.

Mean (± SE) fecundity (no. eggs/female) and longevity (days) of a pyridaben-sensitive strain (SS) and a pyridaben-resistant strain (Py-R) of Tetranychus truncatus in nine high-temperature day frequencies. ‘Hot day : normal day’ indicates the number of consecutive hot days between consecutive normal days. Means within a strain capped with a different capital letter (fecundity) or lowercase letter (longevity) are significantly different (Tukey’s test: p < 0.05)
Discussion
Natural temperatures exhibit highly regular changes in time and space, such as circadian changes in temperature and seasonal periodic changes (Wang and Ma 2013). There are repetitive changes in the temperature of a natural environmental over a period of several days, which are specifically reflected in the occurrence of successive high- or low-temperature days, and the occurrence of high- and low-temperature days alternates periodically (Tank et al. 2002). Therefore, periodically recurring high-temperature weather in the natural environment has an inevitable impact on the growth and development of arthropods. For example, after experiencing the same high temperature 4× in succession, adult Drosophila melanogaster laid significantly fewer eggs than it did at room temperature (Krebs 1994). Robinet et al. (2013) found that, after repeated high-temperature days, the egg and larval stages of Thaumetopoea pityocampa were both affected.
We are the first to study how periodically recurring changes in high temperatures affect the development and reproduction of T. truncatus. We found that there were interaction effects between frequency and strain, and their effects on the longevity, fecundity, and developmental stages of T. truncatus were significant. Bowler and Terblanche (2008) postulated that the heat resistance of arthropods decreases with the maturity of individuals. However, in some studies, the survival rate of arthropods in each life stage did not show obvious regularity (Ju et al. 2011; Bahrndorff et al. 2021; Zhao et al. 2022). Our experimental results showed that the egg and immature stages of the sensitive strain were all affected by hot days, whereas the adult stage was less affected by frequency. The egg stage of the resistant strain was less affected; it was mainly affected in the immature and adult stages. Therefore, the sensitivity of arthropods to high temperature in different life-history stages cannot be summarized simply as increasing with the age of arthropods, but seems to vary according to species and experimental conditions.
There is evidence that repeated daily stress causes a significant decrease in both fertility and longevity in D. melanogaster females exposed to short-term heat stress. However, stressing insects weekly, only in the first 2 weeks after eclosion, caused a significant increase in the total level of fertility (Gruntenko et al. 2021). Ma et al. (2004) found that the closer the stage of periodic repeated high temperature action is to the adult stage, the more easily the reproduction is affected. We found that the reproductive rate of both sensitive and resistant strains were lower than those at normal temperatures under all nine frequencies of high-temperature days, but the fecundity of resistant strains was significantly affected by interval of the highest normal temperature days (1:3, 2:3).
Many studies have shown that, although high temperatures affect the longevity of arthropods, a low night temperature may delay the early death of adults to a certain extent. A low nighttime temperature phase after a high temperature provides sufficient energy in time for the synthesis of such substances as polyol compounds, cytoprotective agents or heat shock proteins; thus, the heat tolerance of arthropods may be effectively restored, continuous daily high-temperature stress may be avoided, and arthropods may continue to survive (Krebs and Loeschcke 1994; Terblanche et al. 2010; Tank et al. 2002; Yan et al. 2013; Wang et al. 2019; Miao et al. 2020). This could be related to why the damage caused by mites in the summer was serious, as the diurnal temperature range in the Hexi area in China in our experiment was large.
We thus found that the relationship between high-temperature day frequency and phytophagous mites is of significance for predicting the occurrence of agricultural pest populations and formulating appropriate control strategies against the background of climate warming. However, the temperature patterns under study are not sufficiently systematic or comprehensive. Moreover, climate warming not only has a profound impact on the growth, development, reproduction, survival and reproduction of T. truncatus, but also on the competition among species, predation and parasitism of natural enemies, etc. The main direction in the next stage of research will involve an experimental design that more closely mimics the effect of actual high-temperature modes on mites in the field, a study of the high adaptive response of resistant mites to environmental changes, and the prediction of their behavioral, physiological, and adaptive molecular mechanisms. To expand the research object, also changes at the three trophic levels of plants, herbivores and natural enemies under the influence of high temperature will be studied.
Conclusions
We found that the frequency of high-temperature days significantly affected the development of different stages, longevity, and reproduction of T. truncatus. These characteristics also showed significant differences between susceptible and resistant strains. The higher the number of normal-temperature days (vs. high-temperature days), the more the negative impact of high temperature can be offset. Moreover, the large diurnal temperature difference in the Hexi area simulated in our experiment also results in the low night temperature repairing the adverse impact of daily high-temperature stress on mites to a certain extent. However, the adaptation of T. truncatus to high temperatures comes at the cost of reduced reproductive rates.
Data availability
The datasets generated during and analysed during the current study are available from the corresponding author on reasonable request.
References
Alain M, Franck D (2019) A comprehensive database for the Tetranychidae. Spider Mites Web: http://www1.montpellier.inra.fr/CBGP/spmweb. Accessed 20 July 2021
Bahrndorff S, Alemu T, Kristensen TN, Sørensen MH, Høye TT, Holmstrup M (2021) Thermal adaptations of adults and eggs in the Arctic seed bug Nysius groenlandicus (Insecta: Hemiptera) from South Greenland. Polar Biol 44(3):491–498. https://doi.org/10.1007/s00300-021-02807-6
Bayu MSYI, Ullah MS, Takano Y, Gotoh T (2017) Impact of constant versus fluctuating temperatures on the development and life history parameters of Tetranychus urticae (Acari: Tetranychidae). Exp Appl Acarol 72(3):205–227. https://doi.org/10.1007/s10493-017-0151-9
Bowler K, Terblanche JS (2008) Insect thermal tolerance:what is the role of ontogeny, ageing and senescence? Biol Rev 83(3):339–355. https://doi.org/10.1111/j.1469-185X.2008.00046.x
Chang ZZ, Sun JT (2021) Susceptibility of Tetranychus Truncatus to three acaricides. J Maize Sci 29(4):110–114
Chen ZW, Zhu H, Cai JY, Zhang J (2014) Investigation of leaf mite species of corn seed production field in Hexi Corridor. Chin Hortic Abstracts 30(2):218–220
Chen S, Fleischer SJ, Saunders MC, Thomas MB (2015) The influence of diurnal temperature variation on degree-day accumulation and insect life history. PLoS ONE 10(3):e0120772. https://doi.org/10.1371/journal.pone.0120772
Fariborz R, Jahangir K, Ralf N, Wannes D, Thomas VL (2019) Characterization of abamectin resistance in Iranian populations of European red mite, Panonychus Ulmi Koch (Acari: Tetranychidae). Crop Prot 125:104903. https://doi.org/10.1016/j.cropro.2019.104903
Gruntenko NE, Karpova EK, Babenko VN, Vasiliev GV, Andreenkova OV, Bobrovskikh MA, Menshanov PN, Babenko RO, Rauschenbach IY (2021) Fitness analysis and transcriptome profiling following repeated mild heat stress of varying frequency in Drosophila melanogaster females. Biology 10(12):1323. https://doi.org/10.3390/biology10121323
Guillermo F, Daniel AB, Jelle C, José MR, Maria-Dolors P, Xavier B, Francisco B (2011) An experimental test of the role of environmental temperature variability on ectotherm molecular, physiological and life-history traits: implications for global warming. Comp Biochem Phys A 159:242–246. https://doi.org/10.1016/j.cbpa.2011.03.002
Hafez AM, Abbas N (2021) Insecticide resistance to insect growth regulators, avermectins, spinosyns and diamides in Culex quinquefasciatus in Saudi Arabia. Parasites Vector 14:558. https://doi.org/10.1186/s13071-021-05068-8
Jin PY, Chen L, Lu T, Hong XY (2018) Spider mites of agricultural importance in China, with focus on species composition during the last decade(2008–2017). Syst Appl Acarol 23(11):2087–2099
Ju RT, Chen GB, Wang F, Li B (2011) Effects of heat shock, heat exposure pattern, and heat hardening on survival of the sycamore lace bug, Corythucha ciliate. Entomol Exp Appl 141(2):168–177. https://doi.org/10.1111/j.1570-7458.2011.01180.x
Karl I, Janowitz SA, Fischer K (2008) Altitudinal life-history variation and thermal adaptation in the copper butterfly Lycaena Tityrus. Oikos 117:778–788. https://doi.org/10.1111/j.0030-1299.2008.16522.x
Karl I, Stoks R, De Block M, Janowitz SA, Fischer K (2011) Temperature extremes and butterfly fitness: conflicting evidence from life history and immune function. Global Change Biol 17:676–687. https://doi.org/10.1111/j.1365-2486.2010.02277.x
Krebs RA, Loeschcke V (1994) Costs and benefits of activation of the heat-shock response in Drosophila melanogaster. Funct Ecol 8(6):730–737
Kuroki I, Nakamura K, Tagawa J (2019) Geographic variation of temperature effects on initial colony development of Lasius japonicus (Hymenoptera: Formicidae). Appl Entomol Zool 54:175–183. https://doi.org/10.1007/s13355-019-00610-8
Liu JP, Wang C, Nicolas D, Lu YH (2021) Impact of temperature on survival rate, fecundity, and feeding behavior of two aphids, Aphis gossypii and Acyrthosiphon Gossypii, when reared on cotton. Insects 12:565. https://doi.org/10.3390/insects12060565
Ma CS, Hau B, Poehling HM (2004) Effects of pattern and timing of high temperature exposure on reproduction of the rose grain aphid, Metopolophium Dirhodum. Entomol Exp Appl 110:65–71. https://doi.org/10.1111/j.0013-8703.2004.00123.x
Ma CS, Wang L, Zhang W, Rudolf Volker HW (2018) Resolving biological impacts of multiple heat waves: interaction of hot and recovery days. Oikos 127:622–633. https://doi.org/10.1111/oik.04699
Ma YH, Zhang AX, Jin DY (2020) Efficacy tests of nine insecticides on Tetranychus Truncatus Ehara. Agricultural Biotechnol 9(3):34–36
Matsuda T, Fukumoto C, Hinomoto N, Gotoh T (2013) DNA-based identification of spider mites: molecular evidence for cryptic species of the genus Tetranychus (Acari: Tetranychidae). J Econ Entomol 106(1):463–472
Miao ZQ, Tu YQ, Guo PY, He W, Jing TX, Wang JJ, Wei DD (2020) Antioxidant enzymes and heat shock protein genes from Liposcelis bostrychophila are involved in stress defense upon heat shock. Insects 11:839. https://doi.org/10.3390/insects11120839
Pham TA, Hwang SY (2020) High temperatures reduce nutrients and defense compounds against generalist Spodoptera litura F in Rorippa dubia. Arthropod-plant Inte 14(3):333–344. https://doi.org/10.1007/s11829-020-09750-z
Rimy SJ, Das G, Gotoh T, Ullah MS (2021) Lethal and sublethal effects of bifenazate on the biological parameters of Tetranychus Truncatus Ehara (Acari: Tetranychidae). Syst Appl Acarol-UK 26(11):2118–2132. https://doi.org/10.11158/saa.26.11.12
Rismayani, Ullah MS, Chi H, Gotoh T (2021) Impact of constant and fluctuating temperatures on population characteristics of Tetranychus pacificus (Acri: Tetranychidae). J Econ Entomol 114(2):638–651. https://doi.org/10.1093/jee/toaa327
Robinet C, Rousselet J, Pineau P, Miard F, Roques A (2013) Are heat waves susceptible to mitigate the expansion of a species progressing with global warming? Ecol Evol 3(9):2947–2957. https://doi.org/10.1002/ece3.690
Tank A, Wijngaard JB, Können GP, Boehm R, Demaree G, Gocheva A, Mileta M, Pashiardis S, Hejkrlik L, Kern-Hansen C, Heino R, Bessemoulin P, Mueller-Westermeier G, Tzanakou M (2002) Daily dataset of 20th-century surface air temperature and precipitation series for the European Climate Assessment. Int J Climatol 22(12):1441–1453. https://doi.org/10.1002/joc.773
Terblanche JS, Nyamukondiwa C, Kleynhans E (2010) Thermal variability alters climatic stress resistance and plastic responses in a globally invasive pest, the Mediterranean fruit fly (Ceratitis capitata). Entomol Exp Appl 137(3):304–315. https://doi.org/10.1111/j.1570-7458.2010.01067.x
Thomas VL, John V, Anastasia T, Wannes D, Luc T (2010) Acaricide resistance mechanisms in the two-spotted spider mite tetranychus urticae and other important Acari: a review. Insect Biochem Molec 40:563–572. https://doi.org/10.1016/j.ibmb.2010.05.008
Thomas VL, Atsushi Y, Luc T, Ralf N, Wannes D (2014) The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic Biochem Phys 121:12–21. https://doi.org/10.1016/j.pestbp.2014.12.009
Ullah MS, Lim UT (2015) Life history characteristics of Frankliniella occidentalis and F. intonsa (Thysanoptera: Thripidae) in constant and fluctuating temperature. J Econ Entomol 108(3):1000–1009. https://doi.org/10.1093/jee/tov035
Ullah MS, Gotoh T, Lim UT (2014) Life history parameters of three phytophagous spider mites, Tetranychus piercei, T. truncatus and T. bambusae (Acari: Tetranychidae). J Asia-Pac Entomol 17:767–773. https://doi.org/10.1016/j.aspen.2014.07.008
Wang L, Ma CS (2013) Impacts of periodic repeated heat events on ecological performance in insects. Chin J Applie Entomol 50(6):149–150
Wang XR, Wang C, Ban FX, Zhu DT, Liu SS, Wang XW (2019) Genome-wide identification and characterization of HSP gene superfamily in whitefly (Bemisia tabaci) and expression profiling analysis under temperature stress. Insect Sci 26:44–57. https://doi.org/10.1111/1744-7917.12505
Wang CM, Zhang JW, Zhou TW, Jing JJ, Zhang XJ, Guo C, Jia CH (2021) Pest infestation, status of pesticides application and strategies on reducing their input in irrigated maize field in the Hexi corridor region. J Maize Sci 29(4):88–96
World Health Organization (2016) Monitoring and managing insecticide resistance in Aedes mosquito populations: interim guidance for entomologists. WHO. https://apps.who.int/iris/handle/10665/204588. Accessed 1 Jan 2020
Xin JL (1988) Tetranychoidea. Agricultural Acarology. China Agricultural Press, Beijing, p 247
Yan HR, Jia HH, Wang XL, Gao HR, Guo XQ, Xu BH (2013) Identification and characterization of an Apis cerana Cerana Delta class glutathione S-transferase gene (AccGSTD) in response to thermal stress. Naturwissenschaften 100:153–163. https://doi.org/10.1007/s00114-012-1006-1
Yin XL, Chu C, Bai QH, Guo PP (2022) Seed maize in Zhangye: Meteorological conditions in growth period and climate quality certification. Chin Agric Sci Bull 38(2):71–77
Zhang W, Rudolf VHW, Ma CS (2015) Stage-specific heat effects: timing and duration of heat waves alter demographic rates of a global insect pest. Oecologia 179:947–957. https://doi.org/10.1007/s00442-015-3409-0
Zhang B, Zhao F, Hoffmann A, Ma G, Ding HM, Ma CS (2016) Warming accelerates carbohydrate consumption in the diapausing overwintering peach fruit moth Carposina Sasakii (Lepidoptera: Carposinidae). Environ Entomol 45:1287–1293
Zhang YH, Cai TW, Ren ZJ, Liu Y, Yuan MJ, Cai YF, Yu C, Shu RH, He S, Li JH, Wong Adam CN, Wan H (2021) Decline in symbiont-dependent host detoxification metabolism contributes to increased insecticide susceptibility of insects under high temperature. ISME J 15(12):3693–3703. https://doi.org/10.1038/s41396-021-01046-1
Zheng DH, Li Z, Jin DY (2016) Occurrence and control of Tetranychus Truncatus and T.urticae on corns of Yanbian Area. Agr Sci Tech 17(2):394–397
Zhao CC, Chen HS, Guo JY, Zhou ZS (2022) Effects of Fluctuating thermal regimes on life history parameters and body size of Ophraella communa. Insects 13(9):821. https://doi.org/10.3390/insects13090821
Funding
This work was supported by [National Natural Science Funds of China] (Grant numbers [31860503]).
Author information
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Song, L., Fu, W., Li, W. et al. The influence of high-temperature frequency variation on the life-history traits of pyridaben-sensitive and -resistant strains of Tetranychus truncatus. Exp Appl Acarol 92, 109–122 (2024). https://doi.org/10.1007/s10493-023-00873-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-023-00873-5