
Abstract
Cannibalism is commonly found among phytoseiids at almost all mobile life stages. The avoidance of kin cannibalism in the presence of non-kin has been observed in a limited number of phytoseiid species. However, studies examining kin recognition by individuals at different life stages are limited. Early experience with conspecifics can alter individuals’ social behaviour at later stages. In this study, we examined the influence of ontogenetic isolation on cannibalism and kin recognition by the predatory mite Amblyseius herbicolus (Chant) (Acari: Phytoseiidae) at different life stages. In a choice test, A. herbicolus protonymphs, deutonymphs, pre-ovipositional adults, and ovipositional adults were used as predators, and larvae (siblings vs. non-siblings) were given as prey. Two rearing treatments were applied to the predators to generate different social experience during ontogeny: reared in isolation or in pairs with ad libitum mixed stages of Carpoglyphus lactis (Acari: Carpoglyphidae). Social experience with siblings during ontogeny significantly reduced the cannibalism rate. Despite the social experience during ontogeny, protonymphs, deutonymphs, and pre-ovipositional adults did not discriminate siblings from non-siblings during cannibalism. The discrimination behaviour between siblings and non-siblings of A. herbicolus occurred after adults became ovipositional. We hypothesize that A. herbicolus adults could learn the characteristics of their larvae through the contact with and assessment of their eggs during oviposition. The result of this study shows that kin discrimination could be phase-specific. Future studies can investigate if contact with non-kin (or non-sibling) eggs and subsequently hatching larvae can influence the discrimination ability of ovipositional adults.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cannibalism has been widely observed among animal taxa, where individuals predate on conspecifics mostly to survive food scarcity (Fox 1975; Pfennig et al. 1993; Schausberger 2003; Khodaei and Long 2020). The cannibals acquire nutrients and reduce the intraspecific competition for resources and mates (Schausberger 2003). Despite the benefits individuals gain, the major concern of cannibalism is its influence on the inclusive fitness of cannibals (Schausberger 2003; Khodaei and Long 2020). If other potential food including non-kin is present, the cannibals can reduce their inclusive fitness if close relatives are cannibalised (Hamilton 1964; Schausberger 2003). Thus, the inclusive fitness of cannibals will depend on whether kin recognition is present, and mechanisms that allow the avoidance of kin cannibalism would be favoured by selection (Schausberger 2003; Rudolf et al. 2010).
Many species can discriminate conspecifics according to their genetic relatedness, leading to differential treatment of kin and non-kin (Holmes and Sherman 1983; Tang-Martinez 2001; Penn and Frommen 2010; Breed 2014). Besides cannibalism, kin recognition is also observed in many other social interactions such as mating, competition, foraging, habitat selection, dispersal, and cooperation (Pfennig and Frankino 1997; Pfennig et al. 1998; West et al. 2007; Johnstone and Cant 2008; Penn and Frommen 2010; Bitume et al. 2013). The driving forces of kin recognition are thought to be inbreeding avoidance and increased inclusive fitness (Mateo 2004; Penn and Frommen 2010). Animals may use contextual cues, or possess alleles coding for kin recognition, or prior association, and phenotype matching to recognise kin; the last two mechanisms are most often reported (Mateo 2004; Penn and Frommen 2010).
The mechanisms used by animals in kin recognition are not mutually exclusive, and can vary intra-specifically, depending on the condition of individuals (Mateo 2004; Penn and Frommen 2010). Kin recognition was found to vary during ontogenetic development of each individual (Mott et al. 2019). For example, whereas worker ants (Ectatomma tuberculatum) aged between 2 and 10 weeks discriminated nestmate larvae from non-nestmate larvae when they were responsible for nursing (Fénéron and Jaisson 1995), younger and older workers did not. The third instars of ladybirds, Propylea dissecta and Coccinella transversalis, avoided cannibalising kin, whereas the fourth instars of both species showed no discrimination towards kin and non-kin (Pervez et al. 2005). The neighbour recognition based on familiarity of the marbled salamander Ambystoma opacum was only observed in individuals after metamorphorsis (Walls 1991). Young ground squirrels (Urocitellus beldingi) used familiarisation to recognise kin, whereas older individuals used phenotype matching (Mateo 2017).
Learning and imprinting can happen at prenatal and postnatal stages, and during critical or sensitive periods in most animals (Hepper and Cleland 1998). Early learning can influence an individual’s behaviour at later life stages (Signorotti et al. 2014). In particular, social isolation can negatively influence individuals’ recognition ability with long-lasting effects (Schausberger 2004, 2007; Lesne et al. 2016; Mateo 2017; Schausberger et al. 2017). For example, compared with socially experienced individuals, social isolation of the predaceous mite Phytoseiulus persimilis led to changes in their life-history traits and behaviour (Schausberger et al. 2017). Socially isolated P. persimilis protonymphs became more cannibalistic towards larval siblings (Schausberger 2004).
Kin recognition in many species requires learning through social interaction (Fénéron and Jaisson 1995; Penn and Frommen 2010). For instance, zebrafish larvae (Danio rerio) imprint on kin (siblings) at days 5 and 6 post-fertilisation using visual and olfactory cues (Biechl et al. 2016). Those zebrafish larvae that experienced non-kin cues during this period did not initiate imprinting, and no discrimination between kin and non-kin was observed later in life. Worker ants of E. tubercalutum isolated from the brood after emergence showed no preference between nest and non-nest mate larvae (Fénéron and Jaisson 1995).
Predatory mites of the family Phytoseiidae (Acari: Mesostigmata) are excellent subjects to test the ontogenetic influence on cannibalism and kin recognition, as most of them were reported to be cannibalistic, individuals at each mobile life stage can be potential cannibals, and they have relatively short developmental time (Schausberger 2003; Li et al. 2021). The thelytokous mite Amblyseius herbicolus (Chant) is a Type-III generalist predator feeding on various phytophagous mites, insects, and pollen (Moraes & Mesa, 1988; McMurtry et al., 2013; Cavalcante et al., 2017). In a choice experiment, the adult A. herbicolus females avoided cannibalising kin (daughters) and preferred non-kin larvae (Zhang and Zhang 2022a). Additionally, adult A. herbicolus females discriminated larvae from their first and second cousins by preferentially eating the more distantly related kin (Zhang and Zhang 2022b). In addition to cannibalism, A. herbicolus also showed a preference to oviposit near eggs of non-kin rather than their own eggs in the absence of prey interference (Zhang and Zhang 2022c). Studies examining kin recognition at different life stages are limited (Mateo 2017). Hence, in this study, we examined the influence of ontogenetic development on cannibalism and kin recognition of A. herbicolus. The influence of social experience during development on kin recognition and cannibalism is also examined to see if kin discrimination in A. herbicolus requires learning from social interaction. Due to the potential for cannibalism of phytoseiids at different life stages, we predict that A. herbicolus at nymphal stages are also cannibalistic. We hypothesise that cannibals will avoid cannibalising siblings in the presence of non-siblings, and previous social interactions with siblings will reduce the occurrence of cannibalistic behaviour and strengthen the avoidance of sibling cannibalism in A. herbicolus.
Materials and methods
Mite rearing
The culture of A. herbicolus has been fed with ad libitum dried fruit mite Carpoglyphus lactis (Acari: Carpoglyphidae) and maintained under laboratory conditions (see Zhang and Zhang 2021 for details). Both the predator and prey species were obtained in early 2021. Adult A. herbicolus females were initially collected from naturally infested nightshade Solanum nigrum leaves at Manaaki Whenua – Landcare Research, St Johns, Auckland, and from a garden in Epsom, Auckland. Carpoglyphus lactis was obtained from Bioforce Limited (Karaka, Auckland). Cultures and experimental units were kept in Plexiglas chambers at 25 ± 1 °C, 80 ± 5% relative humidity, and a L16:D8 h photoperiod.
Two cultures of A. herbicolus from the same strain with identical rearing conditions were maintained separately for more than 2 months before the start of the experiment. Individuals from different cultures were considered non-siblings.
Experimental procedure
The choice test was conducted according to the methods of Schausberger and Croft (2001) and Zhang and Zhang (2022a), with modifications of choice cells used and treatments of testing subjects. Nylon threads (about 2 cm long) were used to harvest eggs from the culture where adults of A. herbicolus preferred to oviposit (Fig. 1). Amblyseius herbicolus reproduced asexually, without males (Moraes and Mesa 1988). To prepare for the ovipositional adults, eggs were reared singly using the modified Munger cells (see Zhang & Zhang 2021 for details) and fed with ad libitum mixed stages of C. lactis. Dry yeast was placed into the cells to feed C. lactis and replenished every second day or when required.

Illustration of the experimental procedure. (1) Eggs of Amblyseius herbicolus were harvested from the culture using nylon threads (ca. 2 cm long) and reared until oviposition using modified Munger cells with ad libitum mixed stage Carpoglyphus lactis. (2) Eggs laid by the same mother (siblings) were reared in isolation or in pairs using the Munger cells with ad libitum mixed stage C. lactis until reaching the designated life stages as cannibals (protonymphs, deutonymphs, pre-ovipositional adults, and ovipositional adults). Prey (at larval stage) hatched individually without food (C. lactis). (3) One cannibal (either socially isolated or experienced protonymph, deutonymph, pre-ovipositional adult, or ovipositional adult) and two prey (sibling and non-sibling at the larval stage) were placed into an enclosed Munger cell (choice cell) for the choice test
Under laboratory conditions, A. herbicolus could oviposit for about 2 weeks and lay 1–2 eggs per day (Zhang and Zhang 2021). Thus, those adults reared in the ovipositional cells were allowed to oviposit over 2 weeks, and the eggs laid were used as testing subjects. The ovipositional cells were checked twice a day and eggs laid were randomly assigned into socially isolated (1 egg) or socially experienced (2 eggs laid by the same adult) cells for hatching. Two eggs reared in pairs were placed together with physical contact to enhance social interaction. The earlier hatched larvae could have contacted the eggs of conspecifics. Food cues can interfere with the early learning of kin in the phytoseiid N. californicus (Christiansen and Schausberger 2017). Thus, in the socially experienced group, the hatched larvae were allowed to interact in the cell for at least 2 h without food. Individuals in the socially isolated group were starved for at least 2 h to ensure a similar treatment. After 2 h, ad libitum mixed stages of C. lactis were provided, and A. herbicolus larvae, both in isolation or in pairs, were reared until reaching the designated life stages. No cannibalism between A. herbicolus larvae reared in pairs was seen during the study. The life stages (i.e., protonymph, deutonymph, pre-ovipositional adult, and ovipositional adult) of cannibals needed for the experiment were randomly assigned for choice tests. In the socially experienced group, if the hatching time between two eggs was greater than 6 h, the hatched larva was removed to avoid extreme starvation. For consistency, the removed larva was not used in the experiment as a predator because it may have contacted the sibling’s egg shell.
All cannibals (protonymphs, deutonymphs, pre-ovipositional adults, and ovipositional adults) used in the choice test were at day 1 (within 24 h of moulting or laying the first egg). Larvae were used as their prey because they are the most vulnerable life stages of phytoseiids (Ghazy et al. 2016). Cannibals were not starved before the choice test. Prey (larvae) were reared in isolation, hatched within 15 h, and not fed to ensure a similar size. No contact between predator and prey were allowed before the choice test.
One socially isolated or experienced cannibal (either protonymph, deutonymph, pre-ovipositional adult, or ovipositional adult), one larva (sibling of the cannibal), and one non-sibling larva were randomly selected and placed into a choice test cell. Food (C. lactis) was not given to individuals in the choice cells to increase cannibalism rate. However, water was supplied to the testing subjects twice daily to prolong their survival (see Zhang and Zhang 2022a for details). The dorsal shield of each prey was marked with a randomly chosen colour for separation. The choice test cells were checked every 30 min during the day (6–8 h) and unmonitored overnight until one of the prey was cannibalised. Cannibalised larvae had shrunk bodies, because the cannibals drained almost all body fluid. Cannibalism choice and latency (time from the start of the choice test to when one prey was cannibalised) were recorded. All cannibals and prey were used only once in the choice test.
More than 50 replications were done for each predatory life stage and social experience. Due to the different cannibalism rates between treatments, and the sample size required for a significant comparison in the binomial test, some life stages had a higher number of replicates than the others. The number of eggs laid by the ovipositional adults (i.e., before and during oviposition) was noted. Individuals reared in pairs were given ad libitum mixed stages C. lactis.
Statistical analysis
Statistical analysis was done using R Development Core Team (2021), graphs were produced with package ggplot2 (Wickham 2016), and models were computed with packages lme4 (Bates et al. 2015) and survival (Therneau 2020). Cox regression was used to determine the influence of previous social experience and life stages on the cannibalism propensity (combining occurrence and latency data). The cannibalistic preference of A. herbicolus was determined using the generalised linear model (GLM) with a binomial distribution. A generalised linear mixed model (GLMM) with a binomial distribution was used to determine the influence of the number of eggs laid before cannibalism, life stages and social experience (reared in isolation or in pairs) of ovipositional A. herbicolus adults on their prey choice (kin or non-kin). The two cultures, where A. herbicolus was harvested, and test IDs were used as random effects in GLMM. An α level of 0.05 was applied.
Results
Social context and cannibalism
The protonymphs, deutonymphs, and adults of A. herbicolus tested in this study were cannibalistic (Fig. 2). Cox regression showed that the social experience affected the propensity to cannibalise: individuals with prior experience with conspecifics were less cannibalistic (Wald χ² = 14.881, df = 1, p < 0.001). Life stages had a bordering significance influence the propensity to cannibalise (Wald χ² = 7.503, df = 3, p = 0.057). The interaction of life stages and social experience had a marginally significant influence on cannibalism propensity (Wald χ² = 7.568, df = 3, p = 0.056). Protonymphs, deutonymphs, and pre-ovipositional adults of A. herbicolus reared in pairs had a reduced cannibalism rate compared to those reared in isolation (protonymphs: Wald χ² = 9.645, p = 0.001; deutonymphs: Wald χ² = 8.055, p = 0.005; pre-ovipositional adults: Wald χ² = 4.330, p = 0.037, all df = 1). Ovipositional adults reared in isolation and in pairs had similar cannibalism rates (Wald χ² = 0.101, df = 1, p = 0.75).

The survival rates of Amblyseius herbicolus prey larvae (either siblings or non-siblings) in the presence of conspecific predators at different life stages: protonymphs (P), deutonymphs (D), pre-ovipositional adults (A), and ovipositional adults (OA). The social experiences of the predators are given in parentheses, where they are reared either in isolation (I) or in pairs (P)
Social context and kin recognition
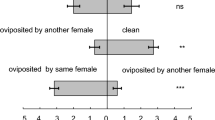
The social experience did not influence the cannibalism preference of A. herbicolus (GLMM: Wald χ² = 0.001, df = 1, p = 0.98). Avoidance of sibling cannibalism did not differ among life stages and early social experience except for deutonymphs (Fig. 3; GLMM: Wald χ² = 7.941, df = 1, p = 0.047). The protonymphs, deutonymphs, and pre-ovipositional adults of A. herbicolus had no cannibalism preferences toward siblings and non-siblings (Fig. 3). In contrast, the ovipositional adults of A. herbicolus both reared in isolation and in pairs preferred cannibalising non-siblings, and avoided sibling cannibalism. The number of eggs laid by the ovipositional adults of A. herbicolus varied, with a total of 1–7 eggs before commencing cannibalism. Nevertheless, their cannibalism choice between kin and non-kin was not affected by the number of eggs laid (GLMM: Wald χ² = 0.157, df = 1, p = 0.69).

Larvae (siblings or non-siblings) cannibalised in the choice test by Amblyseius herbicolus at different life stages. Adults used are at two stages: pre-ovipositional adults (Adult) and ovipositional adults (Adult-O). Amblyseius herbicolus reared individually (I) or in pairs (P) are denoted in parentheses. Sample sizes are in parentheses for each life stage. Asterisks denote a significant difference in the adults’ choice from GLM (p < 0.05). GLM test results: protonymph (I), p = 0.41; protonymph (P), p = 0.091; deutonymph (I), p = 0.75; deutonymph (P), p = 1.0; adult (I), p = 0.53; adult (P), p = 0.75; Adult-O (I), p = 0.007; adult-O (P), p = 0.014. Treatments marked with different letters are significantly different (Tukey’s test: p < 0.05)
Discussion
Phytoseiid mites are blind and use tactile and chemical cues in foraging and social interactions (van Wijk et al. 2006; Saitoh et al. 2020). Individuals of A. herbicolus might acquire kin-related information through contacting egg shells, shed skins, self-inspection, metabolic waste products, and other conspecifics. In this study, previous social contacts of A. herbicolus during its ontogeny could influence their tendency to cannibalism: socially isolated protonymphs, deutonymphs, and pre-ovipositional adults had a higher cannibalism rate than socially experienced individuals. Similarly, the spider Anelosimus studiosus was behaviourally affected by previous social experience (Jones et al. 2010). Social females were less aggressive than solitary females towards conspecifics and other prey, and were more frightened of predators (Jones et al. 2010). Although the spiderlings of Agelena labyrinthica generally avoided cannibalism and social isolation had an insignificant influence on their cannibalism behaviour (Lesne et al. 2016), solitary adults, after dispersal, became more aggressive towards conspecifics (Chiara et al. 2019). Therefore, based on the results of this study and previous studies, the previous social experience can influence individuals’ aggressiveness in intraspecific social behaviours. However, the previous social experience with siblings only affected the cannibalism rate of A. herbicolus protonymphs, deutonymphs, and pre-ovipositional adults, but not ovipositional adults. Thus, oviposition and contacting of own offspring can also reduce the cannibalism propensity of A. herbicolus adults.
Body size and weight difference are major factors in cannibalism, where larger individuals predate smaller conspecifics (Dobler and Kölliker 2011). For phytoseiids, individuals at the later stages are more voracious than larvae (Schausberger 2003). However, the propensity for cannibalism in A. herbicolus, as observed in this study, was inversely affected by ontogenetic development. The ovipositional adults of the individually reared A. herbicolus had a reduced cannibalism rate compared with the other life stages tested. A previous study found that the mated males and reproductive females of T. confusum had a reduced propensity for cannibalism compared with unmated individuals (Parsons et al. 2013).
Ovipositional females of A. herbicolus showed the most accurate avoidance of sibling cannibalism. Depending on the ecological circumstances, the benefit and cost of being discriminative among conspecifics can vary (Breed 2014). The benefit for ovipositional A. herbicolus adults to avoid cannibalising their offspring might be higher than individuals at the other life stages prior to oviposition to avoid cannibalising siblings. Our previous studies used ovipositional adults (gravid females) to discriminate their daughters from non-kin and more distant kin (Zhang and Zhang 2022a, b). The contact with eggs laid during oviposition may allow A. herbicolus adults to learn the cues of their offspring or to generalise a template for later kin discrimination. Additionally, oviposition may cause physiological and behavioural changes in A. herbicolus adults, encouraging kin discrimination in cannibalism.
Phenotype matching and familiarisation are both possible mechanisms used by the ovipositional adults of A. herbicolus. Familiarisation may be more important to phytoseiids in kin recognition than the other recognition mechanisms (Schausberger 2007; Muleta and Schausberger 2013): the protonymphs of P. persimilis avoided cannibalising familiar larvae irrespective of the relatedness (kin or non-kin) (Schausberger 2007). Our previous study found that A. herbicolus adults did not discriminate against daughters and nieces, suggesting offspring and siblings may share great similarities in their recognisable characteristics (Zhang and Zhang 2022b). Therefore, the level of corresponding characteristics between siblings and offspring of A. herbicolus must be above the threshold to recognise them as kin (offspring and siblings) (Breed 2010), and adults avoid preying on them in the presence of non-kin or non-siblings.
The kin discrimination mechanism of A. herbicolus is less likely to be innate, and direct learning (familiarisation) during oviposition may be required. As a non-eusocial arthropod, learning from conspecifics encountered during development can be risky for A. herbicolus due to the likelihood of imprinting on non-kin (Breed 2010; Penn and Frommen 2010). Alternatively, learning from eggs (e.g., physical and chemical characteristics) laid during oviposition as a recognition template may minimise recognition errors and maximise the survival of own offspring. Although not statistically significant, most life stages of A. herbicolus tested in this study were slightly biased towards non-siblings, especially protonymphs reared in groups. Thus, we cannot exclude the possibility of kin recognition ability in A. herbicolus nymphs, pre-ovipositional adults, and the likelihood of learning from contacting conspecifics. Additionally, the interference of food cues can also influence the learning of A. herbicolus reared in pairs based on the results of Christiansen and Schausberger (2017). Although we have experimentally ensured individuals reared in pairs would have at least 2 h together before the introduction of C. lactis, some individuals (hatched overnight) might have a longer time than others (hatched during the day). Individuals who had more time together are likely to have more contacts for learning and later be discriminative. Furthermore, a 2-h period may not be sufficient for imprinting as the earlier hatched larva might have contacted the unhatched egg, whereas the later hatched larva might not have a chance to contact the earlier hatched larva. Further studies could investigate the learning of A. herbicolus by increasing their time together in the absence of prey.
In conclusion, we showed that social experience with siblings could reduce the propensity of cannibalism in A. herbicolus. Furthermore, the discrimination against siblings and non-siblings in cannibalism of A. herbicolus was phase-specific: ovipositional A. herbicolus females showed the most accurate avoidance of sibling cannibalism compared to protonymphs, deutonymphs, and pre-ovipositional adults. Thus, the ability of an individual to discriminate kin or sibling might depend on the context, and oviposition might be an important factor in A. herbicolus. This study is the first to examine kin recognition of a phytoseiid at different life stages. The absence of kin discrimination in A. herbicolus protonymphs compared to other reported phytoseiid species may be due to their different lifestyles and genetic systems. Future studies can examine whether contact with non-kin eggs and subsequent exposure to the hatched larvae can influence their cannibalism choice.
References
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Biechl D, Tietje K, Gerlach G, Wullimann MF (2016) Crypt cells are involved in kin recognition in larval zebrafish. Sci Rep 6:24590. https://doi.org/10.1038/srep24590
Bitume EV, Bonte D, Ronce O, Bach F, Flaven E, Olivieri I, Nieberding CM (2013) Density and genetic relatedness increase dispersal distance in a subsocial organism. Ecol Lett 16:430–437. https://doi.org/10.1111/ele.12057
Breed MD (2010) Social Recognition. In: Breed MD, Moore J (eds) Encyclopedia of Animal Behavior. Academic Press, pp 267–272
Breed MD (2014) Kin and nestmate recognition: The influence of W. D. Hamilton on 50 years of research. Anim Behav 92:271–279. https://doi.org/10.1016/j.anbehav.2014.02.030
Chiara V, Portugal FR, Jeanson R (2019) Social intolerance is a consequence, not a cause, of dispersal in spiders. PLoS Biol 17:e3000319. https://doi.org/10.1371/journal.pbio.3000319
Christiansen IC, Schausberger P (2017) Interference in early dual-task learning by predatory mites. Anim Behav 133:21–28. https://doi.org/10.1016/j.anbehav.2017.09.005
Dobler R, Kölliker M (2011) Influence of weight asymmetry and kinship on siblicidal and cannibalistic behaviour in earwigs. Anim Behav 82:667–672. https://doi.org/10.1016/j.anbehav.2011.06.017
Fénéron R, Jaisson P (1995) Ontogeny of nestmate brood recognition in a primitive ant, Ectatomma tubercalutum Olivier (Ponerinae). Anim Behav 50:9–14. https://doi.org/10.1006/anbe.1995.0215
Fox LR (1975) Cannibalism in natural populations. Annu Rev Ecol Evol Syst 6:87–106. https://doi.org/10.1146/annurev.es.06.110175.000511
Ghazy NA, Osakabe M, Negm MW, Schausberger P, Gotoh T, Amano H (2016) Phytoseiid mites under environmental stress. Biol Control 96:120–134. https://doi.org/10.1016/j.biocontrol.2016.02.017
Hamilton WD (1964) The genetical evolution of social behaviour. I. J Theor Biol 7:1–16. https://doi.org/10.1016/0022-5193(64)90038-4
Hepper PG, Cleland J (1998) Developmental aspects of kin recognition. Genetica 104:199–205. https://doi.org/10.1023/A:1026477724836
Holmes WG, Sherman PW (1983) Kin recognition in animals: The prevalence of nepotism among animals raises basic questions about how and why they distinguish relatives from unrelated individuals. Am Sci 71:46–55
Johnstone RA, Cant MA (2008) Sex differences in dispersal and the evolution of helping and harming. Am Nat 172:318–330. https://doi.org/10.1086/589899
Jones TC, Pruitt JN, Riechert SE (2010) Reproductive success in a socially polymorphic spider: Social individuals experience depressed reproductive success in isolation. Ecol Entomol 35:684–690. https://doi.org/10.1111/j.1365-2311.2010.01227.x
Khodaei L, Long TAF (2020) Kin recognition and egg cannibalism by Drosophila melanogaster larvae. J Insect Behav 33:20–29. https://doi.org/10.1007/s10905-020-09742-0
Lesne P, Cazalé-Debat L, Portugal FR, Trabalon M, Jeanson R (2016) Early experience and social performance in spiderlings. Anim Behav 122:9–16. https://doi.org/10.1016/j.anbehav.2016.09.007
Li G, Pattison N, Zhang Z (2021) Immature development and survival of Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae) on eggs of Tyrophagus curvipenis (Fain & Fauvel) (Acari: Acaridae). Acarologia 61:84–93. https://doi.org/10.24349/acarologia/20214415
Mateo JM (2004) Recognition systems and biological organization: The perception component of social recognition. Ann Zool Fenn 41:729–745
Mateo JM (2017) The ontogeny of kin-recognition mechanisms in belding’s ground squirrels. Physiol Behav 173:279–284. https://doi.org/10.1016/j.physbeh.2017.02.024
McMurtry JA, De Moraes GJ, Sourassou NF (2013) Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst Appl Acarol 18:297–320. https://doi.org/10.11158/saa.18.4.1
Moraes G, Mesa N (1988) Mites of the family Phytoseiidae (Acari) in Colombia, with descriptions of three new species. Int J Acarol 14:71–88. https://doi.org/10.1080/01647958808683790
Mott CL, Dzaferbegovic H, Timm SR, Whiteman HH (2019) Influences of facultative paedomorphosis on kin selection in a larval salamander, Ambystoma talpoideum. Behaviour 156:287–306. https://doi.org/10.1163/1568539X-00003542
Muleta MG, Schausberger P (2013) Smells familiar: Group-joining decisions of predatory mites are mediated by olfactory cues of social familiarity. Anim Behav 86:507–512. https://doi.org/10.1016/j.anbehav.2013.05.040
Parsons W, Zhong W, Rudolf VHW (2013) Mating status and kin recognition influence the strength of cannibalism. Anim Behav 85:365–369. https://doi.org/10.1016/j.anbehav.2012.11.006
Penn D, Frommen J (2010) Kin recognition: An overview of conceptual issues, mechanisms and evolutionary theory. In: Kappeler P (ed) Animal behaviour: Evolution and mechanisms. Springer, Berlin, Heidelberg, pp 55–85
Pervez A, Gupta AK, Omkar (2005) Kin recognition and avoidance of kin cannibalism by the larvae of co-occurring ladybirds: A laboratory study. Eur J Entomol 102:513–518. https://doi.org/10.14411/eje.2005.073
Pfennig DW, Frankino WA (1997) Kin-mediated morphogenesis in facultatively cannibalistic tadpoles. Evolution 51:1993–1999. https://doi.org/10.1111/j.1558-5646.1997.tb05120.x
Pfennig DW, Reeve HK, Sherman PW (1993) Kin recognition and cannibalism in spadefoot toad tadpoles. Anim Behav 46:87–94. https://doi.org/10.1006/anbe.1993.1164
Pfennig DW, Ho SG, Hoffman EA (1998) Pathogen transmission as a selective force against cannibalism. Anim Behav 55:1255–1261. https://doi.org/10.1006/anbe.1997.9996
R Development Core Team (2021) R: A language and environment for statistical computing [computer software]. R Foundation for Statistical Computing, Vienna, Austria
Rudolf VHW, Kamo M, Boots M (2010) Cannibals in space: The coevolution of cannibalism and dispersal in spatially structured populations. Am Nat 175:513–524. https://doi.org/10.1086/651616
Saitoh F, Janssen A, Choh Y (2020) The use of volatile cues in recognition of kin eggs by predatory mites. Ecol Entomol 45:1220–1223. https://doi.org/10.1111/een.12872
Schausberger P (2003) Cannibalism among phytoseiid mites: A review. Exp Appl Acarol 29:173–191. https://doi.org/10.1023/a:1025839206394
Schausberger P (2004) Ontogenetic isolation favours sibling cannibalism in mites. Anim Behav 67:1031–1035. https://doi.org/10.1016/j.anbehav.2003.10.006
Schausberger P (2007) Kin recognition by juvenile predatory mites: Prior association or phenotype matching? Behav Ecol Sociobiol 62:119–125. https://doi.org/10.1007/s00265-007-0444-9
Schausberger P, Croft B (2001) Kin recognition and larval cannibalism by adult females in specialist predaceous mites. Anim Behav 61:459–464. https://doi.org/10.1006/anbe.2000.1611
Schausberger P, Gratzer M, Strodl MA (2017) Early social isolation impairs development, mate choice and grouping behaviour of predatory mites. Anim Behav 127:15–21. https://doi.org/10.1016/j.anbehav.2017.02.024
Signorotti L, Cappa F, d’Ettorre P, Cervo R (2014) Novel insights into the ontogeny of nestmate recognition in polistes social wasps. PLoS ONE 9:e97024. https://doi.org/10.1371/journal.pone.0097024
Tang-Martinez Z (2001) The mechanisms of kin discrimination and the evolution of kin recognition in vertebrates: A critical re-evaluation. Behav Processes 53:21–40. https://doi.org/10.1016/s0376-6357(00)00148-0
Therneau TM (2020) A Package for Survival Analysis in R. R package version 3.4-0. https://CRAN.R-project.org/package=survival
van Wijk M, Wadman W, Sabelis M (2006) Morphology of the olfactory system in the predatory mite Phytoseiulus persimilis. Exp Appl Acarol 40:217–229. https://doi.org/10.1007/s10493-006-9038-x
Walls SC (1991) Ontogenetic shifts in the recognition of siblings and neighbours by juvenile salamanders. Anim Behav 42:423–434. https://doi.org/10.1016/S0003-3472(05)80041-7
West SA, Griffin AS, Gardner A (2007) Evolutionary explanations for cooperation. Curr Biol 17:R661–672. https://doi.org/10.1016/j.cub.2007.06.004
Wickham H (2016) Ggplot2: Elegant graphics for data analysis. Springer-Verlag, New York
Zhang K, Zhang Z-Q (2021) The dried fruit mite Carpoglyphus lactis (Acari: Carpoglyphidae) is a suitable alternative prey for Amblyseius herbicolus (Acari: Phytoseiidae). Syst Appl Acarol 26:2167–2176. https://doi.org/10.11158/saa.26.11.15
Zhang K, Zhang Z-Q (2022a) Kin recognition by cannibals is modulated by hunger level in a generalist predatory mite Amblyseius herbicolus (Chant) (Acari: Phytoseiidae). J Appl Entomol 146:579–585. https://doi.org/10.1111/jen.12973
Zhang K, Zhang Z-Q (2022b) A thelytokous predatory mite is more cannibalistic towards distant kin. https://doi.org/10.1093/cz/zoac074. Curr Zool zoac074
Zhang K, Zhang Z-Q (2022c) Amblyseius herbicolus mothers prefer to oviposit near eggs of non-kin in the absence of prey Carpoglyphus lactis (Acari: Phytoseiidae, Carpoglyphidae). Syst Appl Acarol 27:2347–2354. https://doi.org/10.11158/saa.27.11.16
Acknowledgements
We thank Bioforce Ltd. for providing the initial population of Carpoglyphus lactis. We thank Ms Anne Austin (Manaaki Whenua – Landcare Research) and two anonymous reviewers for constructive comments and suggestions that improved this manuscript.
Funding
This study was supported in part by New Zealand Government core funding for Crown Research Institutes from the Ministry of Business, Innovation and Employment’s Science and Innovation Group.
Author information
Authors and Affiliations
Contributions
KZ and ZQZ designed the study. KZ completed the experiment, analyzed the data, and wrote the paper draft. KZ and ZQZ reviewed the manuscript.
Corresponding author
Ethics declarations
The authors declare no competing interests. The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhang, K., Zhang, ZQ. Social context during ontogeny affects cannibalism and kin recognition of the predatory mite Amblyseius herbicolus at different life stages. Exp Appl Acarol 88, 317–328 (2022). https://doi.org/10.1007/s10493-022-00764-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-022-00764-1