
Abstract
Florida citrus and vegetable crops generate billions of dollars in revenue every year. However, wind, freezing temperatures, hurricanes, and diseases negatively impact production. Windbreaks located perpendicular to the prevailing wind can increase farm production simply by reducing wind and modifying microclimate. Windbreaks can also help in managing pathogens such as citrus canker (Xanthomonas campestris pv. citri). To study the modification of wind speed, temperature, and relative humidity on the leeside of single-row tree windbreaks in southern Florida, automated weather stations were installed in 2007/2008 at 2 m above the ground along transects perpendicular to a eastern redcedar (Juniperus virginiana) and three cadaghi (Corymbia torelliana) (WB1–WB3) windbreaks. All windbreaks reduced wind speed, with minimum wind speed (~5% of the open wind speed) at two times the distance of windbreak height (2H, where H = windbreak height in m) on the leeside of a E. redcedar (~17% porosity) and at 4H (~3–30% of the open wind speed) and 6H (<50% of the open wind speed) on the leeside of cadaghi windbreaks WB1 (~22% porosity) and WB2 (~36% porosity), respectively, when the wind direction was nearly perpendicular to the windbreaks. Wind speed reduction was observed up to 31 times the windbreak height (31H). Cadaghi windbreaks reduced wind speed on the leeside even during a tropical storm event. Temperatures on the leeside of the windbreaks were warmer during the day and cooler near the windbreaks at night compared to temperature in the open fields. This study demonstrates that single-row tree windbreaks can reduce wind and modify the microclimate to enhance crop production for Florida growers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Physical damage to crops, and nutrient loss through soil erosion are problematic to growers worldwide especially in agricultural systems where strong winds are common. This is a serious problem in areas where the soil is sandy and the top soil contains nutrients. It has been shown that wind transported soil contains more nutrients than the field soil from which it was removed (Nuberg 1998; Sudmeyer and Scott 2002). Strong winds may also reduce flowering, increase flower shedding, reduce pollination and increase endosperm abortion in some fruit trees such as cherry (Prunus avium) and can decrease the number of pollinators such as bees (Peri and Bloomberg 2002). Factors such as these can lead to a reduction in productivity and pose a serious threat to the economic viability of growers throughout the US and in particular Florida.
Florida is the major citrus producer and one of the leading vegetable producers in the US. Both productions are profitable, generating ~$20 billion annually. Agriculture products from Florida were distributed throughout the US and exported to 140 countries (FDACS 2007). However, increase in disease incidences in citrus such as citrus canker (Xanthomonas campestris pv. citri), impact of high winds (including tropical storms and hurricanes), and freezes during cold fronts threaten these industries. Wind scarring, caused by sand abrasion and rubbing of plant parts primarily on young tender fruits, is another factor that reduces the quality of Florida agricultural products for fresh market (Albrigo 1976; Miller et al. 1990; Miller and Burns 1992; Morales and Davies 2000; Núñez-Elisea and Crane 2000; Stover et al. 2004).
Citrus canker is caused by bacteria and spores spread through wind and rain splashes. Its spread intensifies during catastrophic events such as tropical storms and hurricanes. A canker eradication program was active until the end of 2005. Infected trees were removed, and all trees within a 579 m (1,900-foot) radius of an infected tree were mandatorily destroyed. Because of the extensive canker spread in the 2004/2005 hurricane seasons, eradication of canker became unfeasible, and the United States Department of Agriculture (USDA) deactivated the rule in January 2006. As a result of this change in policy, canker control efforts have shifted to canker management leaving Florida growers with the challenge of needing to develop effective tools for reducing the spread of the spores.
Windbreaks are commonly used in South America for canker management (Leite and Mohan 1990; Gottwald and Timmer 1995; Behlau et al. 2008) and to reduce the impact of high winds throughout the globe (Jones and Sudmeyer 2002; Peri and Bloomberg 2002; Sudmeyer and Scott 2002; Brandle et al. 2004; Sudmeyer and Flugge 2005). They are also an important component of integrated citrus management (Leite 2000; Behlau et al. 2008), and citrus growers in Florida are beginning to introduce tree windbreaks for canker management.
As suggested windbreaks are used commonly in other parts of the world and their effectiveness is well documented. However, in most cases the research has focused on windbreaks composed of multiple rows of trees (Sun and Dickinson 1997; Michels et al. 1998; Cleugh 2002; Peri and Bloomberg 2002; Sudmeyer and Scott 2002). Most of the windbreaks planted around citrus groves in Florida are composed of a single-row of trees versus multiple rows due to limited space for planting and because of the already established citrus tree. Therefore, the function of single-row windbreaks needs to be established if Florida growers are to plant windbreaks on large scale.
Established windbreaks of native tree species such as eastern redcedar (Juniperus virginiana) and pines (Pinus spp.) are present in some areas of Florida. But due to the urgency caused by the spread of the citrus canker, citrus growers are also using non-native, fast-growing species such as cadaghi (also known as Eucalyptus torelliana in earlier literature and now Corymbia torelliana) and other eucalypts. Cadaghi is highly suitable for windbreaks (Sun and Dickinson 1997) and preferred by growers because of its fast growth, dense canopy, branch retention, and evergreen nature. Field observations suggest that cadaghi can produce effective windbreaks within 2–3 years. It is now planted for citrus windbreaks in Florida (Rockwood et al. 2008) and is increasing in use every year.
This case study was conducted to examine the spatial and temporal patterns of wind speed, temperature and relative humidity on the leeside of single-row E. redcedar and cadaghi windbreaks at the Southwest Florida Research and Education Center/University of Florida (SWFREC/UF) and C&B Farms.
Materials and methods
Study areas
The study was conducted at SWFREC/UF (26°27′46″N, 81°26′04″W) at Immokalee and C&B Farms (26°27′30″N, 80°58′46″W) near Clewiston, Florida. The sites are ~48 km apart, but both have similar climates and poorly drained sandy soils. In 2008, the absolute minimum temperature in the area was −0.4°C in January, and absolute maximum was 35.1°C in June recorded at Florida Automated Weather Network (FAWN) station at SWFREC/UF. Prevailing wind direction is from the east.
SWFREC/UF had a single-row E. redcedar windbreak on its northern boundary (Table 1; Fig. 1a). Single-row cadaghi windbreaks at C&B Farms were planted along irrigation channels. East–west oriented windbreaks were planted approximately every 300 m, while the distances between north–south oriented windbreaks ranged between 480–680 m. Sugarcane windbreaks were planted parallel to north–south oriented windbreaks (between two east–west oriented windbreaks) and bordered ~0.9 ha blocks. The oldest cadaghi windbreak was planted in 1988, while the rest were planted in subsequent years. Some windbreaks were established and functional while others were in the early stages of growth. Three established cadaghi windbreaks (WB1–WB3) were selected for the study (Table 1; Fig. 1b). Orientation of WB1 was true east–west azimuth and was located on the northern boundary of C&B Farms. WB2 was ~300 m south of WB1 oriented along true east–west azimuth (Fig. 1b). Orientation of WB3, the easternmost windbreak, was true north–south azimuth. Porosities of E. redcedar, WB1, and WB3 were uniform throughout the height of the windbreak. However, pruning of lower branches had created large gaps near the ground in WB2.

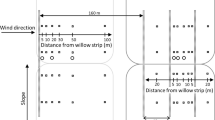
Location of measurement transects (only one transect shown) and automated weather stations along transects relative to study windbreaks at (a) Southwest Florida Research and Education Center/University of Florida and (b) C&B Farms
Methods
Five windbreak sections (~25 m long each in E. redcedar and ~45 m long each in cadaghi windbreaks) were randomly selected in each windbreak. Total height, diameter at breast height (DBH), live crown length (length from the lowest live branch to the top of the tree) and spacing between trees were measured for each tree in the windbreak sections (Table 1). Digital images of randomly selected windbreak sections were used to estimate porosity following Kenny’s (1987) digital analysis method. For this method, digital color images are converted to black and white, then black and white pixels are counted and porosity is calculated as the ratio of white to total pixels in the picture.
Automated weather stations (AWS) were installed at two times the windbreak height (2H, where H = windbreak height in m), 6, 10, and 14H along two separate transects perpendicular to the windbreak on the south side of E. redcedar windbreak at SWFREC between January and June 2008 (Fig. 1a). At C&B Farms, AWS were installed in November 2007 between WB1 and WB2 in transects perpendicular to both windbreaks and measurements were taken until December 2008. AWS were installed at 2, 4, 6, and 10H from WB1 (Fig. 1b). Another series of AWS were installed at 2, 4, 6, and 8H from WB2 in the same transects. Stations at 10, 6, 4, and 2H from WB1 were approximately at 15, 23, 27, and 31H from WB2. Similarly, stations at 8, 6, 4, and 2H from WB2 were approximately at 13, 14, 15, and 16H from WB1. AWS were also installed at 4, 8, 12, and 16H on the west side of WB3 along two transects from July to December 2008 (Fig. 1b).
At each station, wind speed, temperature, and relative humidity were measured at a height of 2 m above the ground. Wind speed was measured using HOBO® wind speed smart sensors (S-WSA-M003) and temperature and relative humidity were measured using HOBO® temperature and relative humidity sensors (S-THA-M002). Automatic measurements were taken every 30 s until July and every 1-min after that. Measurements were recorded in HOBO® Micro Station Data Loggers (H21-002). Control stations located at 266 m (38H from E. redcedar windbreak) and 531 m (59H from WB2) from the windbreaks at SWFREC and C&B Farms, respectively, measured wind speed, temperature, relative humidity and wind direction in the open at 2 m above the ground. Wind speed and direction were measured using HOBO® wind speed and direction smart sensors (S-WCA-M003).
Data analysis
Hourly averages were computed from the recorded data. Readings from transects were also averaged for each location. Data were filtered by wind direction and used in the analysis only when the incident angle of the wind was between 0 and 180 degrees to the windbreak on the windward side. Therefore, only the measurements when wind direction was between 270 and 90 degrees azimuth (between west and east cardinal directions) were considered for E. redcedar and cadaghi WB1 windbreaks since AWS were located on the south side of both windbreaks, and the orientation of both windbreaks was true east–west azimuth (Fig. 1). For cadaghi windbreaks WB2 and WB3, only the measurements between wind directions of 90 and 270 (between east and west cardinal directions), and 0 and 180 (between north and south cardinal directions) degrees azimuth, respectively, were considered. Relative values were calculated for wind speed, temperature and relative humidity measured at each station on the leeside to normalize to the conditions at the control station as
where RV is the relative values of wind speed, temperature and relative humidity; U the variable measured on the leeside of the windbreaks and U o the same variable measured at the same time interval at the control station.
To examine the patterns of wind speed, temperature and relative humidity change on the leeside of the windbreaks during different weather events, these relative values were plotted against time (Figs. 2, 3, 4, 5, 8, 9, 10, 11, 12, 13, 14) (Cleugh 2002; Sudmeyer and Scott 2002). Values less than 100 indicate reduction in the measured variables on the leeside compared to the control station and greater than 100 indicate an increase.

Relative wind speed on the leeside at different distances from the eastern redcedar windbreak when the wind direction was between 45 and 135 degrees (90 ± 45 degrees) to the windbreak and open wind speed was >1 m/s

Relative wind speed at 2H and 14H on the leeside of the eastern redcedar windbreak when the wind direction was less than 45 and greater than 135 degrees to the windbreak and open wind speed was >1 m/s

Relative wind speed on the leeside at different distances from WB1 at C&B Farms when wind direction was nearly perpendicular (90 ± 15 degrees) to the windbreak and open wind speed was >2 m/s

Relative wind speed on the leeside at different distances from WB2 at C&B Farms when wind direction was between 60 and 120 degrees (90 ± 30 degrees) to the windbreak and when open wind speed was >3 m/s
Two weather events were considered to look at the temperature modification. Weather was considered normal when the temperature in the open was greater than 10°C. For cold front events, temperature below 10°C in the open was considered. For temporal variation in temperature and relative humidity, time between 0700 and 1900 h was considered day and between 1900 and 0700 night. In some cases, only the measurements from stations which had the minimum and maximum observations, respectively, were presented to make the plots legible. As sugarcane windbreaks were fully grown and influenced measurements in September 2008, only data collected prior to August 31, 2008 were used in the analysis.
Results
Wind reduction
On the leeside of the E. redcedar windbreak, minimum wind was always recorded at 2H and maximum at 14H regardless of wind speed and direction. Maximum wind reduction was obtained when wind direction was perpendicular to the windbreak. Relative wind speed at 14H was less than 80% of the open wind speed when the wind direction was between 45 and 135 degrees (90 ± 45 degrees) to the windbreak (Fig. 2) and at 2H was always lower than 18% of the open wind speed. When the wind direction was nearly perpendicular to the windbreak, it was less than 5% of the open wind speed. When wind direction was less than 45 and greater than 135 degrees to the windbreak, relative wind speed at 14H remained more or less the same as when wind direction was nearly perpendicular to the windbreak, but there was an increase in wind speed at all other locations (Fig. 3). Regardless of wind direction, wind generally was not detected at 2H when the open wind speed was less than 2.5 m/s.
At C&B farms, minimum wind speed on the leeside of WB1 was recorded at 2H when wind direction was less than 20 and greater than 160 degrees to the windbreak. When wind direction was between 75 and 105 degrees (90 ± 15 degrees) to the windbreak, wind speed at all measurement locations on the leeside was lower than in the open and minimum wind speed was recorded at 4H (Fig. 4). Wind speed gradually increased up to 15H and then decreased at 16H as it approached WB2. Wind was detected at 2H even when the open wind speed was less than 1.5 m/s and wind direction was nearly perpendicular (90 ± 15 degrees) to the windbreak.
On the leeside of WB2, wind speed at 23 and 27H occasionally reached the open wind speed when the direction was between 0 and 180 degrees to the windbreak. When wind direction was less than 45 and greater than 135 degrees to the windbreak, minimum wind speed was observed at 2H. When wind direction was between 60 and 120 degrees to the windbreak, 6H had the minimum wind speed (Fig. 5). As observed on the leeside of WB1, wind speed gradually increased at locations further away from the windbreak and decreased at 31H as it approached WB1.
Cadaghi windbreaks reduced wind on the leeside even during a tropical storm. On August 19, 2008, tropical storm Fay passed through the Florida peninsula. Wind direction ranged between East–Northeast in the morning to West by midnight. Wind speed at C&B Farms ranged between 2.8 and 13 m/s. Average wind speed on the leeside of WB2 and WB3 was less than 90 and 80% of the open wind speed, respectively, when the wind direction was between 0 and 180 degrees to the windbreak (Figs. 6, 7). Average wind speed was the minimum at 6H on the leeside of WB2 and at 4H on the leeside of WB3. Due to WB1 at about 33H from WB2 and sugarcane windbreak at about 20H from WB3, wind speed at the last locations was lower than at preceding locations. Wind was forced through the gaps in the lower portion of WB2 causing occasional higher wind speed at 2H compared to the open when the wind direction was perpendicular the windbreak.

Average relative wind speed on the leeside of WB2 at C&B Farms during the tropical storm Fay on August 19, 2008 when wind direction was between 0 and 180 degrees to the windbreak

Average relative wind speed on the leeside of WB3 at C&B Farms during tropical storm Fay on August 19, 2008 when wind direction was between 0 and 180 degrees to the windbreak
Temperature modification
During normal weather conditions, regardless of the wind speed, nighttime temperature on the leeside of the E. redcedar windbreak was cooler at 2H (up to 1.5°C) and warmer at 14H (up to 1.3°C) compared to temperature in the open (Fig. 8). Temperature patterns on the leeside of the windbreak at all locations were almost similar to temperature in the open when the open wind speed exceeded 3 m/s. During the day, temperature at all locations on the leeside of the windbreak was usually higher than in the open (Fig. 9). Temperature at 2H was up to 2.5°C warmer while at 14H was only 0.6°C warmer than in the open. Temperatures at all locations were either similar or higher than in the open when the wind direction was perpendicular to the windbreak.

Nighttime temperature at 2H and 14H on the leeside of the eastern redcedar windbreak during normal weather conditions when the wind direction was between 0 and 180 degrees to the windbreak. Month above the time axis indicates the beginning of the month

Daytime temperature at 2H and 14H on the leeside of the eastern redcedar windbreak during normal weather conditions when the wind direction was between 0 and 180 degrees to the windbreak. Month above the time axis indicates the beginning of the month
At C&B Farms, nighttime temperature on the leeside at locations near WB1 (2H and 6H) did not show any specific trend until the end of June, after which the measurements were lower than in the open (Fig. 10). Temperatures at 10, 14 and 16H were usually higher (up to 2%) than in the open and slightly greater than at 2H and 6H. Beginning July, temperatures at all locations were lower than in the open. Similar patterns were observed on the leeside of WB2 when wind direction was from the south, except that the temperature at all locations was relatively higher compared to open (not shown here). Daytime temperature on the leeside of WB1 and WB2 was also higher than in the open. On average relative temperatures at 2 and 6H from WB1 were 2% higher than in the open, where as it was only 1% higher at 10, 14 and 16H.

Nighttime temperature at 6H and 16H on the leeside of WB1 during normal weather conditions when the wind direction was between 0 and 180 degrees to the windbreak. Month above the time axis indicates the beginning of the month
During cold fronts, temperature near the windbreaks was cooler than in the open. Temperatures at 2H and 6H on the leeside of E. redcedar windbreaks were cooler up to 1 and 1.3°C than in the open, respectively, when the open wind speed was less than 2 m/s. At C&B Farms, temperatures at 2 and 6H on the leeside of WB1 were up to 1.7 and 1.9°C cooler than in the open, respectively (Fig. 11). Temperature at all measurement locations was either equal or similar to the open when the open wind speed was greater than 2 m/s.

Temperature at 2H and 16H on the leeside of WB1 at C&B Farms during cold fronts when the wind direction was between 0 and 180 degrees to the windbreak
Relative humidity modification
Nighttime relative humidity on the leeside of the E. redcedar windbreak was higher than in the open beginning February (Fig. 12). Relative humidity gradually increased from February until peaking at the end of May. On average, nighttime relative humidity at all locations was ~4% higher than in the open. Daytime relative humidity also showed the same pattern with ~9% higher than in the open at all locations, except that relative humidity at 2 and 6H was slightly lower and at 10 and 14H was slightly higher than in the open until mid February (Fig. 13).

Nighttime relative humidity at 2H and 14H on the leeside of the eastern redcedar windbreak. U is the relative humidity on the leeside of the windbreak. U o is the relative humidity at the control station. Month above the time axis indicates the beginning of the month

Daytime relative humidity at 2H and 14H on the leeside of the eastern redcedar windbreak. U is the relative humidity on the leeside of the windbreak. U o is the relative humidity at the control station. Month above the time axis indicates the beginning of the month
On the leeside of WB1 at C&B Farms, 6H had the maximum and 16H had the minimum nighttime relative humidity, among all locations until the end of May when the wind direction was from the north (Fig. 14). On average, nighttime relative humidity was 3.8% higher at 6H and 1% lower at 16H compared to the open. When the wind direction was from the south, maximum relative humidity was recorded at 6H among all locations on the leeside of WB2 (not shown here). On the leeside of WB1, average daytime relative humidity at 16H was 1% higher and at 6H was 2% lower than in the open during the study. When the wind was from the south, 6H on the leeside of WB2 had the maximum and 2H had the minimum daytime relative humidity among all locations (not shown here).

Nighttime relative humidity at 6H and 16H on the leeside of WB1 at C&B Farms when the wind direction was from north. U is the relative humidity on the leeside of the windbreak. U o is the relative humidity at the control station. Month above the time axis indicates the beginning of the month
Discussion
Wind speed on the leeside of the E. redcedar and cadaghi windbreaks was lower than in the open most of the time but extent of wind reduction varied with windbreak porosity and wind direction. Maximum wind reductions were obtained on the leeside when wind direction approached perpendicular to the windbreak. Regardless of the wind direction, the lowest wind speeds were recorded at 2H on the leeside of the E. redcedar windbreak. At C&B Farms, the lowest wind speeds were recorded at 2H only when the wind direction was oblique and at 4 or 6H on the leeside of WB1 and WB2, respectively, when the wind direction was perpendicular to the windbreaks. The location of maximum wind reduction ranged between 2 and 6H (Loeffler et al. 1992) and the location moved closer to the windbreak when windbreak porosity decreased (Cleugh 1998). Therefore, it is not unusual to see minimum wind speed at 2H on the leeside of E. redcedar windbreak which had ~17% porosity and at 4H on the leeside of WB1 with ~22% porosity. However, large gaps (due to branch pruning) in WB2 allowed more wind to pass through and the minimum wind speed was recorded further away at 6H.
Wind reduction on the leeside in our study varied between 5 and 100% and E. redcedar windbreak which had the lowest porosity (~17%) reduced the maximum wind. Wind reduction in the our study was generally higher than reported by Foereid et al. (2002) where the wind speed was 37% of the open wind speed closer to single-row willow windbreak and 86% at 7H. Other studies reported wind speeds between 40 and 100% of the open wind speed on the leeside (Brenner et al. 1995; Zhang et al. 1995). Since extent of wind reduction is generally related to windbreak porosity (Cleugh et al. 2002), wind reduction in the current study is within the expected range. Distance of wind reduction on the leeside depends on the windbreak height and can extend up to 30H (Wang and Takle 1995; Cleugh and Hughes 2002; Vigiak et al. 2003) and sometimes up to 60H (Caborn 1957). Distance of wind reduction extended up to 31H which is within the reported ranges. Windbreaks also influenced wind up to a distance of 5H on the windward side of the windbreak (Cleugh 1998). At C&B Farms, stations at 16H, 31H and 16H from WB1, WB2 and WB3 were closer to WB2, WB1 and sugarcane windbreak, respectively. Therefore, more wind reductions occurred at these locations compared to preceding locations in transects, respectively. Measurements at these locations were lower than in the open also because of the compounded effects of multiple windbreaks.
Reduced wind speed on the leeside influenced temperature and relative humidity. At SWFREC, nighttime temperature near the windbreak on the leeside was generally lower than the temperature in the open and higher at locations further away from the windbreak. An opposite pattern was observed during the day. Similar results were observed behind WB1 and WB2 at C&B Farms. Temperature generally increases with reduction in wind speed, and the location of maximum temperature coincides with the location of minimum wind speed (Cleugh et al. 2002). The results of this study are in general agreement with the observations at other locations (McAneney et al. 1990; Cleugh 1998). Foereid et al. (2002) also observed increased temperature near a willow windbreak in Denmark during the day, but a decrease at night. Relative humidity also increased on the leeside during the day (Sudmeyer et al. 2002). However, compared to wind reduction, temperature and relative humidity modifications were insignificant and such modification extended up to 10–12H behind the windbreak (Cleugh et al. 2002). There was also an effect of season on temperature and relative humidity modification in the current study. Increase in temperature on the leeside gradually decreased after May, but relative humidity generally increased compared to open. Plant transpiration is a cooling process that increases humidity in the surrounding area. Increases in soil moisture lowered temperature and increased relative humidity in the later part of the study.
Result suggests that temperature reduction on the leeside near windbreaks at night can make freeze events worse. In Florida, this usually occurs at night or early in the morning when the temperature is the lowest. Temperature inversion takes place at night and stratification of air layers occurs on the leeside of the windbreak. Reduced wind on the leeside of windbreak causes less exchange of heat between air layers. During the process cold air, which is heavier settles near the ground. This can lead to formation of frost on the leeside during calm nights (Brandle et al. 2004). In January 2009, frost formed on the leeside closer to WB1 at night during a cold front. Crops near the windbreak on the leeside suffered more damage compared to crops away from the windbreak, but frost did not form in areas without windbreak. Cold sensitive crops may suffer more damage during such events. Such catastrophic events can be avoided by allowing some air to flow through the windbreak. This suggests that windbreaks must be designed to meet the requirements of the landowner (Brandle et al. 2004). For example, if the windbreak is needed for protection of crops, medium porosity (40–50%) windbreaks are ideal, but for visual barrier or the protection of structures, windbreaks with low porosity may be used.
One of the conditions required for citrus canker dispersal is wind-driven rain with speeds greater than 8 m/s. At this wind speed, bacteria are dispersed within trees and from tree to tree (Timmer et al. 2000), and wind speeds greater than 8 m/s are required to force canker bacteria through stomates and wounds (Graham et al. 2004). Wind scar in citrus occurs at wind speeds as low as 6.7 m/s (Metcalf 1936). Results suggest that well designed single-row windbreaks have the potential to reduce physical damage to crops by reducing wind speed and could potentially lower canker infection in citrus.
Conclusions
The patterns are clear, wind on the leeside of the windbreaks was reduced and was lower than in the open when wind direction was between 0 and 180 degrees to the windbreak. Maximum reductions were obtained on the leeside of less porous E. redcedar (~17% porosity) and WB1 (~22% porosity) windbreaks compared to WB2 (~36% porosity). Distance and extent of wind reduction increased between two windbreaks planted parallel to each other. Windbreaks also modified temperature and relative humidity on the leeside. However, temperature and relative humidity modifications were less compared to wind reduction. Temperature reduction near windbreaks on the leeside during cold fronts may be damaging to crops. These patterns suggest that single-row windbreaks can be an effective tool for growers to use to lower canker infection by reducing wind and minimizing damage to plant parts.
References
Albrigo LG (1976) Influence of prevailing winds and hedging on citrus fruit wind scar. Proc Fla State Hort Soc 89:55–59
Behlau F, Belasque J Jr, Bergamin-Filho A, Graham JH, Leite RP Jr, Gottwald TR (2008) Copper sprays and windbreaks for control of citrus canker on young orange trees in southern Brazil. Crop Prot 27:803–807
Brandle JR, Hodges L, Zhou XH (2004) Windbreaks in North American agricultural systems. Agrofor Syst 61:65–78
Brenner AJ, Jarvis PG, van den Beldt RJ (1995) Windbreak-crop interactions in the Sahel. 1. Dependence of shelter on field conditions. Agric For Meteorol 75:215–234
Caborn JM (1957) Shelterbelts and microclimate. Forestry commission bulletin No. 29. Edinburgh University, Edinburgh
Cleugh HA (1998) Effects of windbreaks on airflow, microclimates and crop yields. Agrofor Syst 41:55–84
Cleugh HA (2002) Field measurements of windbreak effects on airflow, turbulent exchanges and microlimates. Aust J Exp Agric 42:665–677
Cleugh HA, Hughes DE (2002) Impact of shelter on crop microclimates: a synthesis of results from wind tunnel and field experiments. Aust J Exp Agric 42:679–701
Cleugh H, Prinsley R, Bird RP, Brooks SJ, Carberry PS, Crawford MC, Jackson TT, Meinke H, Mylius SJ, Nuberg IK, Sudmeyer RA, Wright AJ (2002) The Australian national windbreaks program: overview and summary of results. Aust J Exp Agric 42:649–664
FDACS (2007) Florida agricultural statistical directory. Florida department of agriculture and consumer services (FDACS). http://www.florida-agriculture.com/pubs/pubform/pdf/Florida_Agricultural_Statistical_Directory.pdf Accessed 15 Jan 2009
Foereid B, Bro R, Mogensen VO, Porter JR (2002) Effects of windbreak strips of willow coppice-modelling and field experiment on barley in Denmark. Agric Ecosyst Environ 93:25–32
Gottwald TR, Timmer LW (1995) The efficacy of windbreaks in reducing the spread of citrus canker caused by Xanthomonas campestris pv. citri. Trop Agric 72:194–201
Graham JH, Gottwald TR, Cubero J, Achor DS (2004) Xanthomonas axonopodis pv. citri: factors affecting successful eradication of citrus canker. Mol Plant Pathol 5:1–15
Jones HK, Sudmeyer RA (2002) Economic assessment of windbreaks on the southeast coast of Western Australia. Aust J Exp Agric 42:751–762
Kenny WA (1987) A method for estimating windbreak porosity using digitized photographic silhouettes. Agric For Meteor 39:91–94
Leite RP Jr (2000) Surviving with citrus canker in Brazil. Proc Int Soc Citricult 2:890–896
Leite RP, Mohan SK (1990) Integrated management of citrus bacterial canker caused by Xanthomonas campestris pv. citri in the State of Parana, Brazil. Crop Prot 9:3–7
Loeffler AE, Gordon AM, Gillespie TJ (1992) Optical porosity and windspeed reduction by coniferous windbreaks in Southern Ontario. Agrofor Syst 17:119–133
McAneney KJ, Salinger MJ, Porteous AS, Barber RF (1990) Modifications to an orchard climate with increasing shelter height. Agric For Meteorol 50:211–227
Metcalf W (1936) The influence of windbreaks in protecting citrus orchards. J For 34:571–580
Michels K, Lamers JPA, Buerkert A (1998) Effects of windbreak species and mulching on wind erosion and millet yield in the Sahel. Expl Agric 34:464–499
Miller WM, Burns JK (1992) Grade lowering defects and grading practices for Indian River grapefruit. Proc Fla State Hort Soc 105:129–130
Miller WR, McDonald RE, Sharp JL (1990) Condition of Florida carambolas after hot-air treatment and storage. Proc Fla State Hort Soc 103:238–241
Morales P, Davies FS (2000) Pruning and skirting affect canopy microclimate, yields, and fruit quality of ‘Orlando’ tangelo. Hort Sci 35:30–35
Nuberg IK (1998) Effect of shelter on temperate crops: a review to define research for Australian conditions. Agrofor Syst 41:3–34
Núñez-Elisea R, Crane JH (2000) Selective pruning and crop removal increase early-season fruit production of carambola (Averrhoa carambola L.). Sci Hort 86:115–126
Peri PL, Bloomberg M (2002) Windbreaks in southern Patagoina, Argentina: a review of research on growth models, wind speed reduction, and effects on crops. Agrofor Syst 56:129–144
Rockwood DL, Rudie AW, Ralph SA, Zhu JY, Winandy JE (2008) Energy product options for Eucalyptus species grown as short rotation woody crops. Int J Mol Sci 9:1361–1378
Stover E, Hebb J, Sonoda R (2004) Airblast application of copper fungicide to grapefruit does not affect windscar. Hort Sci 39:516–519
Sudmeyer R, Flugge F (2005) The economics of managing tree–crop competition in windbreak and alley systems. Aust J Exp Agric 45:1403–1414
Sudmeyer RA, Scott PR (2002) Characterization of a windbreak system on the south cost of Western Australia. 1. Microclimate and wind erosion. Aust J Exp Agric 42:703–715
Sudmeyer RA, Crawford MC, Meinke H, Poulton PL, Robertson MJ (2002) Effect of artificial wind shelters on the growth and yield of rainfed crops. Aust J Exp Agric 42:841–858
Sun D, Dickinson GR (1997) Early growth of six native Australian tree species in windbreaks and their effect on potato growth in tropical northern Australia. For Ecol Manage 95:21–34
Timmer LW, Garnsey SM, Graham JH (2000) Compendium of citrus diseases. American Phytopathogical Society, St. Paul
Vigiak O, Sterk G, Warren A, Jagen LJ (2003) Spatial modeling of wind speed around windbreaks. Catena 52:273–288
Wang H, Takle ES (1995) Numerical simulations of shelterbelt effects on wind direction. J Appl Meteorol 34:2206–2219
Zhang H, Brandle JR, Meyer GE, Hodges L (1995) The relationship between open wind speed and wind speed reduction in shelter. Agrofor Syst 32:297–311
Acknowledgments
This research was financially supported by the Institute of Food and Agricultural Sciences (IFAS/UF), the College of Agriculture and Life Sciences (CALS/IFAS), and the Sustainable Agriculture Research and Education (Southern Region) Program. The authors would like to thank SFRC/UF, SWFREC/UF, and C&B Farms staff for their help during the study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tamang, B., Andreu, M.G. & Rockwood, D.L. Microclimate patterns on the leeside of single-row tree windbreaks during different weather conditions in Florida farms: implications for improved crop production. Agroforest Syst 79, 111–122 (2010). https://doi.org/10.1007/s10457-010-9280-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-010-9280-4