
Abstract
Temperate short-rotation agroforestry systems are promising solutions for environmentally and economically sound production of crops and woody biomass. Accurate estimates of microclimatic conditions and tree-crop interactions in these systems are clearly needed to improve their adoption and management, especially in northeastern North America. The objective of this study was to determine the effects of a windbreak and an alley cropping system that were planted with strips of short-rotation willow on microclimate and forage crop yields. Microclimatic conditions (wind speed, air temperature, air relative humidity, light availability, soil moisture, snow depth) and forage yields were measured at different distances from the willow strips in these two agroforestry systems and in agricultural control plots between the third (2020) and fourth years (2021) after experiment establishment. Willow strips significantly reduced wind speed in both experiments, despite their relatively short stature (260 cm, June 2021). Wind speed reduction was greatest (50 and 58% in alley cropping and windbreak systems, respectively) close to the willow strips (5 m), intermediate at 20 m (22% in both systems), and negligible at 50 m (windbreak system). A significant increase in daytime air temperature that reached almost 1 °C was measured close to the willow strips in the agroforestry systems, compared to control plots. Higher soil moisture was measured close to the windbreak. In both agroforestry systems, snow depth was significantly greater near the willow strips. In both systems, forage yield close to the willow strips was not different from that at any other measured distances and in the control plots, which suggests that no competition for resources occurred between the willows and the forage crop during the study. In July 2021, forage yield within the windbreak (0–50 m from the willow strip) was 44% greater than that in the control plots. The alley cropping system had significant negative (-12%, June 2020) and positive (+ 13%, September 2021) effects on forage yield, possibly as a result of temporal variation in climatic conditions. Results of this study stress the importance of better understanding of intra- and inter-annual forage crop variability within agroforestry systems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Windbreaks are the most important, widespread and recognizable temperate agroforestry system in North America and many other regions worldwide. There is unequivocal evidence that these systems, when designed properly, can improve crop and forage yield (Smith et al. 2021). Windbreaks can moderate microclimate conditions, for example, through wind speed reduction, thereby increasing the efficient use of soil water by reducing evaporation from the soil surface and crop transpiration, and leaving more water for crop growth (Brandle et al. 2022). In northern agricultural landscapes, windbreaks can also trap snow and reduce the amount and distance of snow movement, which may help to protect winter cereals and perennials forage crops against frost damage (Kort et al. 2012). In the face of climate anomalies, particularly those favouring summer water stress and reduction in snow loading, windbreaks are increasingly viewed as an effective means of adaptation (Schoeneberger et al. 2012; Hernández-Morcillo et al. 2018).
Alley cropping is another temperate agroforestry system that has attracted growing interest in its potential to aid the agricultural sector in adapting to climate change, while substantially improving soil, water and biodiversity conservation (Wolz et al. 2018). However, alley cropping systems have yet to be widely implemented in temperate regions compared to windbreaks. Major barriers to wider adoption are long-term delays that are required to recoup economic returns from timber harvesting, together with the unsure and variable nature of agricultural productivity in alley cropping, especially in the zone of tree-crop competition (Rois-Díaz et al. 2018; Laroche et al. 2020). Several alley cropping trials experimenting with relatively narrow spacing between tree rows (e.g., 8–15 m) permitted rapid documentation of tree-crop interactions, but they were shown not to be optimal from the viewpoint of agricultural productivity (Garrett et al. 2022). The few studies that have experimented with alley cropping systems where wider spacing was maintained between tree rows (e.g., 30–50 m) have shown satisfactory agricultural yields, generally equivalent to those that have been measured in control fields without trees (Pardon et al. 2018; Carrier et al. 2019). In these studies, forage crops were less sensitive than annual crops to water and light competition in the vicinity of the trees. Many forage species are shade-tolerant with performances that are similar or even better under moderate light shading than in full sunlight (Pang et al. 2019).
Windbreaks and alley cropping systems that are composed of short-rotation woody coppices show promise for an environmentally and economically sound production of agricultural crops and woody biomass of fast-growing trees, such as willow (Salix spp.) (Quinkenstein et al. 2017). These arrangements result in novel short-rotation agroforestry (SRAF) systems, where trees are quickly established and easily propagated within high-density strips (e.g., 10 000 to 15 000 plants per ha). Tree strips are harvested many times before replanting at intervals of 3 to 6 years, forming a temporally and spatially dynamic growing environment for both agricultural crops and woody components (Gamble et al. 2019). Due to short rotation periods, tree height (e.g., 1–6 m at commercial maturity) in these agroforestry systems remains relatively low comparatively to “conventional” agroforestry systems that are devoted to timber production (e.g., 15–20 m). Yet, previous studies in different temperate SRAF systems have suggested that the strip of trees, despite their short stature compared to conventional agroforestry systems, may quickly reduce wind velocity and alter other microclimatic conditions, which can benefit crop yield in a large “sheltered zone” (Foereid et al. 2002; Böhm et al. 2014). At the same time, these systems can rapidly create a narrow zone of potential competition for light and soil moisture near the tree strips (Gamble et al. 2016; Quinkenstein et al. 2017). The consequences of these biophysical interactions on overall crop yield at the field scale remain relatively poorly documented. Previous studies focusing on SRAF systems, mostly in Europe, have shown positive, neutral, and negative effects of tree strips in crop yields (Foereid et al. 2002; Ehret et al. 2018; Kanzler et al. 2019; Gamble et al. 2019; Swieter et al. 2022). Crop yields in SRAF systems appear to be strongly dependent upon the edaphic and climatic contexts of the study and to vary widely with respect to distance from the woody strip, crop species and system design.
More realistic estimates of microclimatic conditions and tree-crop interactions in SRAF systems are clearly needed to help improve decision-making with respect to their implementation, especially in northeastern North America. The objective of this study was to determine the effects of a windbreak and an alley cropping system that were planted with strips of short-rotation willow on microclimate and forage crop yields. We evaluated variation in microclimatic conditions (wind speed, air temperature, relative humidity, light availability, soil moisture and snow depth) and forage yield at different distances from the willow strip in these two SRAF systems and in agricultural control plots. We hypothesized that the competitive effects of willow strips, which were limited to a small area at their interface (yield decrease), would be offset by facilitative effects (yield increase) that extend into the large crop area that is sheltered from winds, thereby resulting in overall yields in SRAF systems that were equal to or greater than those in agricultural control plots.
Materials and methods
Experimental site
The experimental site was located near the town of Wakefield (45o69’ N, 75o97’ W; ca. 155 m above sea level), in southern Québec. Thirty-year mean annual precipitation in the region is 981 mm, which includes 187 cm of snowfall (Chelsea weather station, Environment and Climate Change Canada 2022). Mean annual average temperature is 5.9 °C, with the lowest monthly average in January (-11.0 C) and the highest monthly average in July (20.6 °C). Between January and September inclusive, the frequency of winds in the study area originating from the West (between 220 and 320°) was 38.2% (NOAA 2022). The soil is classified as a Humic-Gleysol (Dalhousie soil series, Lajoie 1962; Soil Classification Working Group 1998) with a silty loam texture (30% clay, 69% silt, 1% sand), Mehlich-III-extractable P of 31.1 kg ha−1, cation exchange capacity (CEC) of 23.0 cmolc kg−1, organic matter content of 6.2%, and bulk pHwater of 5.8 in the uppermost 20 cm.
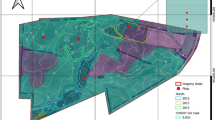
A forage crop field (ca. 200 m long × 300 m wide) that was composed of a mix of perennial grasses (Phleum pratense L., Dactylis glomerata L., Bromus inermis Leyss., Poa pratensis L.) and legumes (Trifolium pratense L., Vicia cracca L., Lotus corniculatus L.) was divided into two independent experiments that tested two short-rotation agroforestry (SRAF) systems: windbreak and alley cropping (Fig. 1). The SRAF systems consists of one (windbreak) or three (alley cropping) willow strips (2.5 m wide × 100 m long) that were planted in May 2018 with two species (Salix viminalis L. and Salix miyabeana Seemen) for wood energy production. In the alley cropping system, willow strips were spaced 40 m apart. The willow strip in the windbreak experiment and the closest willow strip composing the alley cropping experiment were spaced 160 m apart (Fig. 1). Prior to willow planting, the soil in each strip was prepared to a depth of 15 cm and over a width of 2.5 m using a rototiller. In each strip, 30-cm long rooted willow cuttings were planted to a depth of 20 cm in two rows at a density of 13 333 plants ha−1 (100 cm between rows, 75 cm between plants within a row) in a North–South orientation, which is perpendicular to the main wind direction (i.e., from the West). Mean height of willows in mid-June 2020 and 2021 was 190 and 260 cm, respectively. Repression of herbaceous vegetation in each willow strip was assured by a continuous application of hardwood wood chip mulch (ca. 10 cm thickness) after planting and manual weeding in June 2019. During this study (2018 to 2021), the forage crop field was unfertilized. In each experiment, in two experimental blocks perpendicular to the direction of the slope, the SRAF system was compared to an adjacent agricultural control (without willows) that included the same forage composition and the same agricultural history and management (Fig. 1). Within each block, soil physico-chemical properties at the beginning of the experiment (July 2019) were similar between SRAF systems and controls (Table 1).

Experimental layouts (not to scale) showing the locations of the blocks within each experiment and plots (open squares) where snow depth, soil moisture and forage yield were repeatedly measured at different distances from the centre of the willow strips. Open circles represent the locations where photosynthetically active radiation, wind speed, air temperature and relative humidity were measured with data loggers. Dotted lines represent North–South oriented willow rows
Measured variables
Sampling of the variables (soil moisture, snow depth, forage yield) that were repeatedly measured in the SRAF plots was conducted along transects that ran perpendicular to the willow strips (Fig. 1). In the windbreak experiment, the variables were measured at six distances from the centre of the willow strip, i.e., 5, 10, 20, 30, 50 and 100 m. In the alley cropping experiment, the variables were measured at three distances from the centre of the willow strips: 5, 10 and 20 m (Fig. 1). In the control, the variables were measured in 12 (windbreak) to 14 (alley cropping) plots in each block. During 2021, snow depth was measured in each plot on three dates (January 31, February 13 and March 6). In each plot, snow depth was measured at two points (spaced ca. 5 m apart) and the average of the two values was retained for statistical analyses. Soil volumetric water content (hereafter, referred to as soil moisture) was measured in each plot during 2021 at three dates (May 31, July 23 and September 9), more than 3 days after the last rain, with a portable moisture meter (FieldScout TDR100 equipped with 8 cm probe rods, Spectrum Technologies, Aurora, IL). In each plot, soil moisture was measured at three points (spaced ca. 3 m apart) and the average of the three values was retained for statistical analyses.
In each plot, the ground biomass production of forage crop was determined on two dates in 2020 (June 15 and August 19) and 2021 (July 6 and September 13), which were a few days prior to mechanical harvesting of the entire field. Herbaceous plants were clipped 1 cm above the soil surface within a 0.25 m−2 (0.5 m × 0.5 m) quadrat that was located in each of the plots (total of 48 and 56 plots in the windbreak and alley cropping experiments, respectively). The fresh mass of each sample was determined in the field immediately after harvest. Fresh subsamples (about 250 g) were dried to constant mass at 60 °C to calculate forage yield on a dry-mass basis.
Wind speed and photosynthetically active radiation (PAR) were measured by weather stations at 5-min intervals in 2021 between August 21 and September 6. We installed a total of seven WatchDog® data loggers (3910S and 3902, Spectrum Technologies, Aurora, IL) 1.5 m above the ground surface; four were placed in the windbreak experiment (5, 20 and 50 m from the willow strip, and in the control; block 2) and three were installed in the alley cropping experiment (5 and 20 m from the eastern side of the central willow strip, and in the control; block 2) (Fig. 1). The stations were equipped with a digital anemometer (item 3305ADD) and a quantum light sensor (item 3668I) that measured PAR between 400 and 700 nm. Each station was also equipped with one datalogger with sensors for air temperature and relative humidity (HOBO MX2301, Onset Computer Corp., Bourne, MA). These sensors were installed at 1 m from the ground under a two-layer radiation shield made with two 30 cm squares of plywood with a white corrugated plastic on the top plywood to reflect as much heat as possible.
Statistical analyses
In each experiment, we estimated an integrated forage yield value for the whole cultivated areas to contrast with their respective controls. To do so, a weighted average of the forage yield at different distances for each transect was calculated. Within the windbreak experiment, these transect averages were calculated from 1.25 to 50 m, given that windbreak effects on airflow, microclimate and crop yield generally do not exceed 20H (Cleugh 1998) and willow heights during the study were in the range of ca.1.5 to 3 m. Within the alley cropping experiment, the transect averages were calculated from 1.25 to 20 m. The weight for each measurement was obtained with the proportion of the transect that it represented, divided by the number of measurements at this distance, halving the distance between adjacent measurements. For each experiment, ANOVA was conducted using data from the four harvests. An interaction was added to verify if the difference between the agroforestry system and its control differed from one harvest to another, and block was added as a random effect. If the interaction was significant, an ANOVA for each harvest was performed, with Bonferroni correction applied to the significance level.
The effects of the distance from the willow strips were also analyzed using ANOVA in the windbreak (5, 10, 20, 30, 50, 100 m and control) and alley cropping (5, 10, 20 m and control) experiments for three variables, i.e., forage yield, soil moisture and snow depth. These three variables were log-transformed prior to model fitting to obtain normal distributions. In each ANOVA, an interaction accounted for differences in the effect of the distance from one sampling period to the next, and block was added as a random effect. If the distance effect was significant, multiple pairwise comparisons were performed with Tukey's honestly significant difference (HSD) tests. If the interaction was significant, Tukey's HSD tests were also performed for each sampling separately.
For each experiment, meteorological station data were analyzed using multiple ANOVA, with the distances and the control as the only independent categorical variable. These ANOVAs were performed for each day separately to compare wind speed, and they were done for each hour separately to compare air temperature, air relative humidity, and PAR. Confidence intervals at 95% were used to graphically evaluate the difference between two distances.
All analyses were performed in R version 3.5.3 (R Core Team 2018) with the package lme4 for fitting the mixed-effect models and the package car for the ANOVA, i.e., analyses of deviance using Wald's χ2 tests.
Results
Microclimatic conditions
Within the windbreak experiment, wind speed at 5 m from the willow strip was almost invariably lower than that in the control (Fig. 2a). For half of the days, wind speed at 20 m was significantly lower than in the control. Weak differences were measured between 50 m in the windbreak experiment and the control. Similarly, within the alley cropping experiment, wind speed at 5 m from the willow strips was almost invariably lower than that in the control (Fig. 2b). At 20 m, significant wind speed reductions happened more frequently compared to the same distance within the windbreak system.

Daily average of wind speed at different distances from the willow strips in the windbreak a and alley cropping b systems and in their controls. Multiple ANOVAs revealed at least one difference between two distances every day in A (P < 0.001) and in B (at least P < 0.01). Error bars are 95% confidence intervals
Within the windbreak experiment, the air was warmer from 10:00 to 16:00 h at 5 m compared to the other sampling positions, while it was cooler from 18:00 to 23:00 h (Fig. 3a). Relative humidity at 5 m was significantly higher than in the control (from 16:00 to 10:00 h) and at 20 and 50 m (from 21:00 to 10:00 h). Within the alley cropping experiment, air temperature at 5 and 10 m was generally higher than that in the control from 8:00 to 16:00 h (Fig. 3b). Relative humidity at 5 m was significantly higher than that at all other sampling positions from 16:00 to 6:00 h. Relative humidity at 10 m was significantly higher than in the control from 18:00 to 3:00 h.

Hourly average of air temperature (solid lines) and relative humidity (dashed lines) at different distances from the willow strips in the windbreak a and alley cropping b systems and in their controls. In A, for air temperature, ANOVAs were significant from 9:00 to 15:00 h and from 18:00 to 23:00 h (at least P < 0.05); and they were significant from 16:00 to 10:00 h for relative humidity (at least P < 0.05). In B, ANOVAs were significant from 8:00 to 18:00 h for air temperature (at least P < 0.01); and they were significant from 16:00 to 7:00 h for relative humidity (P < 0.001). Error bars are 95% confidence intervals
PAR was reduced only for a short period of time at 5 m from the willow strips. Within the windbreak experiment, there was significantly less light at 5 m compared to the control near 18:00 h (Fig. S1). During the same period within the alley cropping experiment, PAR at 5 m was significantly lower than at 10 and 20 m and in the control (Fig. S2).
The distance from the willow strip affected significantly soil moisture within the windbreak system only (Table 2). Soil moisture at 5 and 10 m was significantly higher than that in the control (Fig. 4). Within both experiments, the distance from the willow strips affected significantly snow depth, and these effects differed from one sampling to another (Table 2, interaction only significant at P < 0.1 within the windbreak experiment). Regardless of sampling date, snow depth at 5 m from the windbreak and in the control was significantly greater compared to that at 30 m (Fig. 5a). In March 2021, snow depth within the windbreak experiment at 5 m (55 cm) was significantly greater than that at 10 to 50 m (45 cm) (data not shown). Within the alley cropping experiment, snow depth at 5 m was significantly greater than that at all other sampling positions (Fig. 5b). In each sampling date, this pattern repeated itself except in February when snow depth was not significantly different between 5 and 10 m (data not shown).

Soil volumetric water content at different distances from the willow strip in the windbreak system and in the control. ANOVA revealed a general effect of distance. Different letters represent a significant difference (Tukey HSD for the multiple comparisons, P < 0.05). Error bars are standard errors of the means

Snow depth at different distances from the willow strips in the windbreak a and alley cropping b systems and in their controls. In both experiments, ANOVA revealed a general effect of distance. Different letters represent a significant difference (Tukey's HSD tests, P < 0.05). Error bars are standard errors of the means
Forage yield
When averaging forage yield for each transect within the agroforestry systems and their controls, it was only in the windbreak system that a significant difference between agroforestry and control was found (Table 2). Yet, there was a significant interaction in both experiments, suggesting that effects occurred during some of the harvests. Overall, forage yield within the windbreak system was significantly greater than what was observed in its controls, which was due to the positive effect on production in July 2021 (Fig. 6a). The overall neutral effect of alley cropping system on forage yield resulted from a yield decreases in June 2020 and a yield increase in September 2021 (Fig. 6b).

Forage yield for the whole cultivated area (weighted average per transect) in the windbreak (A) and alley cropping (B) systems (AF) compared to their controls. For each experiment, an ANOVA was conducted for all harvests together and revealed a significant interaction with harvest; thus, ANOVAs were also conducted separately for each harvest (Bonferroni correction of significance levels). Different letters represent a significant difference (at least P < 0.01). Error bars are standard errors of the means
Within the windbreak experiment, a significant effect of sampling position on forage yield was measured, and this effect differed from one harvest to the next (Table 2). In July 2021, forage yield was significantly greater at 10 m from the willow strip compared to the control (Fig. 7). Within the alley cropping experiment, forage yield did not vary among the different distances from the willow strip.

Forage yield at different distances from the centre of the willow strip in the windbreak system and in the control. ANOVA revealed no general effect of distance, but a significant interaction between harvest and distance. Different letters represent significant differences for the harvest of July 2021 (Tukey's HSD tests for the multiple comparisons, P < 0.05). Error bars are standard errors of the means
Discussion
Changes in microclimatic conditions within the agroforestry systems
As expected, willow strips significantly reduced wind speed in both experiments, despite their relatively low stature (260 cm in June 2021). Wind speed reduction was highest (50 to 58% in alley cropping and windbreak systems, respectively) at the nearest measured distance from the willow strips (5 m), remains important at 20 m (22% in both experiments), and was negligible at 50 m (in the windbreak system). Similar wind speed patterns on the leeward side of short-rotation woody hedgerows were found in other studies that were related to young SRAF systems in temperate climates (Foereid et al. 2002; Böhm et al. 2014; Kanzler et al. 2019). In SRAF systems, the typical structure of hedgerows (i.e., bush-shaped trees in multiple rows) results in lower permeability than classical agroforestry systems with taller trees and greater tree spacing. Decreasing permeability is generally associated with a smaller, but highly sheltered area that is close to the hedgerow (Wang and Takle 1996; Böhm et al. 2014). At 20 m from the willow strip, wind speed reduction was more frequent within the alley cropping than within the windbreak system. Böhm et al. (2014) argued that multiple arranged tree strips within short-rotation alley cropping systems may enhance wind speed protection and result in a larger scale reduction of wind speed compared to a single standing short rotation windbreak.
During a large portion of the day, air temperature (from 8:00 h to 16:00 h) and relative humidity (from 16:00 to 10:00 h) significantly increased close to the willow strips in both experiments. Similar observations were made in comparable SRAF systems (Kanzler et al. 2019; Swieter et al. 2022). Higher daytime air temperature in SRAF systems likely results from a decrease in wind speed, turbulent mixing of air masses with different temperatures, and heat loss (Campi et al. 2009; Brandle et al. 2022). The reduction in turbulent mixing generally reduces the amount of water vapour that is transported away from surfaces in the sheltered area. Therefore, relative humidity and vapour pressure gradients are generally higher in sheltered zones, both during the day and at night (McNaughton 1988). The cooler air temperature in the vicinity of the willow strip from 18:00 to 23:00 h in the windbreak experiment may be a reflection of the shade effect of the willow strip that was the greatest near 18:00 h. Yet, results that are related to spatio-temporal variation of PAR revealed that willow shading was generally negligible at 5 m from the centre of the willow strips in both experiments. Hedgerows of relatively low height that are oriented in a North–South direction in SRAF systems established in temperate latitudes (40–50°) could have little effect on radiation reaching the crop because any shading in the morning or evening could be partly compensated by light reflection off the hedgerow (Jackson and Palmer 1972; Kanzler et al. 2019).
In the windbreak experiment, soil volumetric water content at 5 and 10 m was significantly higher compared to that in the control. Others have written that water evaporation from the soil is generally reduced in the sheltered zone of a windbreak due to reduction in wind speed and transfer of water vapour away from the surface (Brandle et al. 2022). For example, Kanzler et al. (2019) found that short-rotation poplar hedgerows reduced evaporation by an average 27% within 24 m along the tree-crop interface, with the highest reductions at 3 and 9 m from the tree rows. Kanzler et al. (2019) interpreted this result as an indication that microclimate conditions (e.g., lower wind speed and higher relative humidity) in the sheltered zone of woody hedgerows led to lower atmospheric evaporative demand. They argued that lower wind speeds are more likely to limit the replacement of saturated air by unsaturated air, which usually allows favourable conditions for continued evaporation if energy and water are available.
In the windbreak system, our results suggested that the willow strip contributed to snow retention in its vicinity (5 m), which resulted in slightly lower snow depth at 30 m. Similarly, in the alley experiment, snow depth was significantly greatest at 5 m from the willow strips. Very dense and few permeable hedgerows such as those in this study would lead to relatively short, deep snowdrifts on both the windward and leeward sides (Brandle et al. 2022). Deep snow drifts near the tree row may be a concern as they delay snowmelt, which can lead to increased snow mold diseases on forage and winter cereal crops under prolonged snow cover (Gaudet and Laroche 1997). This potential drawback of SRAF systems in cold temperate climates deserves more research attention.
Forage yield response to short rotation agroforestry systems
In July 2021, forage yield within the windbreak (0–50 m from the willow strip) was 44% significantly higher than that in the control plots. Different literature syntheses have highlighted a global positive net effect of temperate zone windbreaks on yield of different crops, including forage crops (Smith et al. 2021), although a closer reading of several individual studies reporting these beneficial effects has revealed great variability in yield results. Higher air temperature in the shelter zone can be an advantage for crop growth, especially in cooler regions (Brandle et al. 2022). Even a small increase in temperature may have substantial positive effects on the rate of crop cellular processes and physiology (Grace 1988). In contrast, lower nighttime temperatures in the shelter zone may reduce crop respiration, resulting in higher rates of net photosynthesis and more vegetative growth.
In September 2021, overall forage yield within the alley cropping system was 13% significantly higher than that in the control plots. In a short rotation poplar alley cropping system, Kanzler et al. (2019) found that winter wheat grain yield in the alleys was 16% higher than that in an open control field. Kanzler et al. (2019) attributed this increased yield to increased temperatures and better water status due to a decrease in potential evaporation. However, in June 2020, forage yield within the alley cropping system was 12% significantly lower than that in the control plots. This discrepancy in forage yield between cutting times could be due to variation in climatic conditions that prevailed during each forage growth period. The period between the beginning of May and mid-June in 2020 was abnormally dry (ca. 40 mm vs. 130 mm that routinely falls; Environment and Climate Change Canada 2022). The increase in sheltered air temperature within the alleys could have accentuated possible crop water stress during this period.
Contrary to expectation, in both experiments, forage yield close to the willow strips (5 m) did not significantly decrease, compared to the control plots. Consequently, our results suggest no competition for light and belowground resources between the willows and the forage crop. The absence of a competition zone at the tree-crop interface was also exemplified by PAR and soil moisture results at 5 m from the willow strips, which were similar to those at the other distances in the alley and in the open field plots. In a short-rotation willow alley cropping system, Ehret et al. (2018) also observed a weak effect of woody strips on forage yield in their vicinity during the first three years of the system establishment (willow height of ca. 4 m after year three).
The experimental agroforestry systems were in their establishment phase (3- to 4 years-old), although willows had reached almost half of their rotation, which was anticipated to be 6 years for the first rotation. The weak effects of trees upon crop yields that were found near the tree rows would possibly strengthen, becoming negative as the trees mature and compete with crops for light and water. For example, in a short rotation willow alley cropping system with a comparable north–south woody strip orientation and willows twice as tall as those in this study (ca. 6 m), Gamble et al. (2019) found that much lower availability in light (57% reduction in PAR at 2.4 m) and soil water at the willow-crop interface was associated with a significant reduction in forage yield. Gamble et al. (2019) estimated that forage crop yield increased by 623 kg DM ha−1 for every 100 µmol m−2 s−1 increase in PAR and by 1038 kg DM ha−1 for every 20 kPa increase in average daily water potential. Similarly, in short-rotation poplar alley cropping systems, reductions in yield of different crops (wheat, oilseed rape and silage maize) were found up to 7 m from the woody strips (tree heights were between 4.8 and 7.5 m) in the alleys (48 and 96 m width) due to the presence of poplar roots, tree shading and tree leaf litter coverage (Swieter et al. 2019, 2022). However, these yield reductions did not negatively influence the average long-term crop yields within the entire alleys of these SRAF systems.
Microclimatic conditions were measured only for one year and within a short period. Likewise, soil moisture was measured on few dates. These measurements, therefore, must be interpreted with caution. Also, crop yields were measured within the first willow rotation only. Yet, the yield of the willow strips in the second rotation is usually higher than that in the first rotation, which may increase competitive effects of willow on crop yields (Dzene et al. 2021). Future research should be guided towards investigating how microclimatic conditions, moisture dynamics and crop yields vary in SRAF systems at larger temporal scales during the growing seasons, within a willow rotation and among different willow rotations.
Conclusions
This study assessed the variation in microclimatic conditions and forage yield at different distances from willow strips that were established in a windbreak and an alley cropping system, and in agricultural control plots. A significant decrease in wind speed and an increase in daytime air temperature were measured in the agroforestry systems, especially close to the willow strips. Significant higher soil moisture was measured close to the windbreak. Snow depth was significantly greater in the vicinity of the willow strips in both agroforestry systems. Overall yields (four cutting times and all distances combined) in the studied agroforestry systems were similar (alley cropping) or significantly higher (windbreak) than those in agricultural control plots. Yet, the effect of short-rotation agroforestry systems on patterns of forage yield varied considerably across cutting times, likely as a result of temporal variation in climatic conditions that prevailed during each cycle of forage growth. Our study highlights the importance of measuring intra- and inter-annual forage crop variability within agroforestry systems.
References
Böhm C, Kanzler M, Freese D (2014) Wind speed reductions as influenced by woody hedgerows grown for biomass in short rotation alley cropping systems in Germany. Agrofor Syst 88:579–591
Brandle JR, Takle E, Zhou X (2022) Windbreak practices. In: Garrett HE, Jose S, Gold MA (eds) North american agroforestry. John & Wiley Sons Inc., Hoboken, pp 91–125
Campi P, Palumbo AD, Mastrorilli M (2009) Effects of tree windbreak on microclimate and wheat productivity in a Mediterranean environment. Eur J Agron 30:220–227
Carrier M, Rhéaume Gonzalez FA, Cogliastro A, Olivier A, Vanasse A, Rivest D (2019) Light availability, weed cover and crop yields in second generation of temperate tree-based intercropping systems. Field Crops Res 239:30–37
Cleugh HA (1998) Effects of windbreaks on airflow, microclimates and crop yields. Agrofor Syst 41:55–84
Dzene I, Hensgen F, Graß R, Wachendorf M (2021) Net energy balance and fuel quality of an alley cropping system combining grassland and willow: results of the 2nd rotation. Agronomy 11:1272
Ehret M, Graß R, Wachendorf M (2018) Productivity at the tree-crop interface of a young willow-grassland alley cropping system. Agrofor Syst 92:71–83
Environment and climate change Canada (2022) 1981–2010 Climate normals and averages. https://weather.gc.ca. Accessed 21 March 2022
Foereid B, Bro R, Mogensen VO, Porter JR (2002) Effects of windbreak strips of willow coppice—modelling and field experiment on barley in Denmark. Agric Ecosyst Environ 93:25–32
Gamble JD, Johnson G, Current DA, Wyse DL, Sheaffer CC (2016) Species pairing and edge effects on biomass yield and nutrient uptake in perennial alley cropping systems. Agron J 108:1020–1029
Gamble JD, Johnson G, Current DA, Wyse DL, Zamora D, Sheaffer CC (2019) Biophysical interactions in perennial biomass alley cropping systems. Agrofor Syst 93:901–914
Garrett HE, Wolz KJ, Walter WD, Godsey LD, McGraw RL (2022) Alley cropping practices. In: Garrett HE, Jose S, Gold MA (eds) North American agroforestry. John & Wiley Sons Inc., Hoboken, pp 163–203
Gaudet DA, Laroche A (1997) Winter survival of cereals parasitized by snow mold. In: Li H, Chen THH (eds) Plant cold hardiness. Springer, Boston, MA, pp 331–342
Grace J (1988) Plant response to wind. Agric Ecosyst Environ 22–23:71–88
Hernández-Morcillo M, Burgess P, Mirck J, Pantera A, Plieninger T (2018) Scanning agroforestry-based solutions for climate change mitigation and adaptation in Europe. Environ Sci Policy 80:44–52
Jackson JE, Palmer JW (1972) Interception of light by model hedgerow orchards in relation to latitude, time of year and hedgerow configuration and orientation. J Appl Ecol 9:341–357
Kanzler M, Böhm C, Mirck J, Schmitt D, Veste M (2019) Microclimate effects on evaporation and winter wheat (Triticum aestivum L.) yield within a temperate agroforestry system. Agrofor Syst 93:1821–1841
Kort J, Bank G, Pomeroy J, Fang X (2012) Effects of shelterbelts on snow distribution and sublimation. Agrofor Syst 86:335–344
Lajoie PG (1962) Étude pédologique des comtés de Gatineau et Pontiac, Québec. Service de recherches, Ministère de l'agriculture du Canada, Ministère de l'agriculture du Québec et Collège Macdonald, Université McGill.
Laroche G, Domon G, Olivier A (2020) Exploring the social coherence of rural landscapes featuring agroforestry intercropping systems using locals’ visual assessments and perceptions. Sustain Sci 15:1337–1355
McNaughton KG (1988) Effects of windbreaks on turbulent transport and microclimate. Agric Ecosyst Environ 22–23:17–39
NOAA (2022) NOAA archive. http://windhistory.com/about.html. Accessed 21 March 2022
Pang K, Van Sambeek JW, Navarrete-Tindall NE, Lin CH, Jose S, Garrett HE (2019) Responses of legumes and grasses to non-, moderate, and dense shade in Missouri, USA. I. Forage yield and its species-level plasticity. Agrofor Syst 93:11–24
Pardon P, Reubens B, Mertens J, Verheyen K, De Frenne P, De Smet G, Van Waes C, Reheul D (2018) Effects of temperate agroforestry on yield and quality of different arable intercrops. Agricult Syst 166:135–151
Quinkenstein A, Tsonkova P, Freese D (2017) Alley cropping with short rotation coppices in the temperate region: a land-use strategy for optimizing microclimate, soil organic carbon and ecosystem service provision of agricultural landscapes. In: Dagar JC, Tewari VP (eds) Agroforestry. Springer, Berlin, pp 263–297
Rois-Díaz M, Lovric N, Lovric M, Ferreiro-Domínguez N, Mosquera-Losada MR, Den Herder M, Graves A, Palma JHN, Paulo JA, Pisanelli A, Smith J, Moreno G, Garcia S, Varga A, Pantera A, Burgess P (2018) Farmers’ reasoning behind the uptake of agroforestry practices: evidence from multiple case-studies across Europe. Agrofor Syst 92:811–828
Schoeneberger M, Bentrup G, De Gooijer H, Soolanayakanahally R, Sauer T, Brandle J, Zhou X, Current D (2012) Branching out: agroforestry as a climate change mitigation and adaptation tool for agriculture. J Soil Water Conserv 67:128A-136A
Smith MM, Bentrup G, Kellerman T, MacFarland K, Straight R, Ameyaw L (2021) Windbreaks in the United States: A systematic review of producer-reported benefits, challenges, management activities and drivers of adoption. Agricult Syst 187:103032
Soil classification working group (1998) The Canadian system of soil classification, 3rd ed. Agriculture and Agri-Food Canada Publication 1646.
Swieter A, Langhof M, Lamerre J, Greef JM (2019) Long-term yields of oilseed rape and winter wheat in a short rotation alley cropping agroforestry system. Agrofor Syst 93:1853–1864
Swieter A, Langhof M, Lamerre J (2022) Competition, stress and benefits: Trees and crops in the transition zone of a temperate short rotation alley cropping agroforestry system. J Agron Crop Sci 208:209–224
Wang H, Takle ES (1996) On three-dimensionality of shelterbelt structure and its influences on shelter effects. Bound-Layer Meteorol 79:83–105
Wolz KJ, Lovell ST, Branham BE, Eddy WC, Keeley K, Revord RS, Wander MM, Yang WH, DeLucia EH (2018) Frontiers in alley cropping: transformative solutions for temperate agriculture. Global Change Biol 24:883–894
Acknowledgements
We extend our sincere thanks to Ian McClatchy, owner of the property where the experiment was installed. This study was supported by a grant from the Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ, Prime-Vert program). We are grateful to Nathalie Magnan and Madelaine Rouleau for administrative assistance. We also thank Dr. W.F.J. Parsons for improving the language of this paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Rivest, D., Martin-Guay, MO. & Cossette, C. Willows rapidly affect microclimatic conditions and forage yield in two temperate short-rotation agroforestry systems. Agroforest Syst 96, 1009–1021 (2022). https://doi.org/10.1007/s10457-022-00758-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-022-00758-3