
Abstract
Despite a general lack of knowledge on the effects of different strategies, conversion of even-aged stands to uneven-aged forest is ongoing across Europe. Conversion of Bavarian Norway spruce stands under the present climate scenario was simulated using the individual tree simulator SILVA. Three conversion strategies initiated at two different stand ages, 30 and 60 years, were simulated to develop uneven-aged mixed stands of Norway spruce, silver fir and European beech: gap creation, shelterwood and passive conversion. The three conversion strategies were furthermore combined with different harvesting rates. These conversion scenarios were compared with maintaining the even-aged Norway spruce management as reference. Scenarios were evaluated in terms of mean annual increment and structural development over a 150-year conversion period as well as the expectation value (EV) for eternal future rotations. Compared to the reference scenario, conversion scenarios reduced mean annual increment (6–43%) and also generally EV (−5–78%), except for some scenarios when stand age at conversion was 60 and applying a 3% discount rate. Conversion by shelterwood always reduced EV (both compared to the reference and other conversion scenarios) when initiated at age 30. With passive conversion, the effect on EV was dependent on the assumptions regarding regeneration costs. Gap conversion generally resulted in high EV and increased stand heterogeneity fastest among the different strategies. Other scenarios, especially passive conversion, were dependent on heavy thinning for developing heterogeneity faster (although still slower than with creation of gaps). Most conversion scenarios eventually resulted in similar structural heterogeneity, but the time it took to get to this stage varied greatly (50–120 years). Conversion by creation of smaller gaps in combination with a high rate of target diameter harvesting resulted in a favorable conversion in terms of economic returns and development of stand heterogeneity due to early income and differentiated regeneration.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In many regions of Europe, mono-specific and even-aged plantations are converted to uneven-aged forest because of anticipated benefits of uneven-aged forest systems with regard to, e.g., biodiversity protection (Hekhuis and Wieman 1999; Larsen 2012; Pretzsch 1998) and reduced risk of damage to the forest ecosystem, e.g., from wind throw (Pukkala et al. 2016). Conversion of pure Norway spruce (Picea abies) is a particular priority as 6–7 million ha have been planted with this species outside of its natural distribution in Europe (Brunner et al. 2006; Hanewinkel 2001; von Teuffel et al. 2004). Consequently, these Norway spruce stands are often vulnerable to damage from natural hazards, such as wind throw (Schütz et al. 2006), bark beetles (Christiansen and Bakke 1988) and root rot (Delatour 1998; Ronnberg et al. 1999). Furthermore, Norway spruce is predicted to have a poor resilience towards the future climate (Huang et al. 2017). As a consequence, it is estimated that at least 4–5 million ha of Norway spruce will have to be converted in the long run (von Teuffel et al. 2004).
Although not a novel concept, conversion of mono-specific, even-aged stands to uneven-aged, mixed forests is new to many regions of Europe (Vítková and Dhubháin 2013). Knowledge on adequate strategies for the conversion into uneven-aged forest as well as the results in terms of wood production, economic outcome and forest structure is therefore limited. Studies conducted so far mainly focused on the earlier steps in the conversion, while the longer time span has largely been omitted (Vítková and Dhubháin 2013). Different stand conditions make it complicated to give general recommendations and have, along with the long time span involved in the conversion, been an important reason why so few experimental plots on conversion have been established (Kenk and Guehne 2001). Those few plots that do exist have not yet been sufficiently long under observation to provide sufficient evidence on the consequences of conversion in terms of production, economic output and structural heterogeneity.
Some knowledge on conversion processes is, however, available, especially from Central Europe (von Lüpke et al. 2004), which has been formulated in a few regional guidelines for the conversion of plantations to uneven-aged forests (e.g., Bayerische Staatsforsten AöR 2018; Larsen and Skov- og Naturstyrelsen 2005). The guidelines cover both passive as well as active conversion methods. Active conversion refers to the active removal of large volumes in the overstorey, before the trees reach the target diameter (TD), to benefit regeneration. Passive conversion is likely to provide a better economic result but is also expected to result in slower conversion (Larsen and Skov- og Naturstyrelsen 2005). However, the guidelines are largely based on theoretical schemes that have not been tested scientifically because there are hardly any practical examples for conversion (Hilmers et al. 2020). Thus, there is a need for scientifically validated schemes that can aid decision making of practitioners.
Because only little, and mostly short term, data are available and since conversion is a long-term process, consequences of conversion at present can mainly be assessed through modelling or analysis of chronosequences producing artificial time series (Pretzsch 2009). Such conversion modelling can be done in two different ways. The first option is to optimize conversion for a variety of parameters (e.g., Messerer et al. 2017; Roessiger et al. 2011, 2013; Yousefpour and Hanewinkel 2009). While this approach may provide a set of beneficial conversion schemes, the exploration of the trade-offs between different alternatives is less clear. Also, while distance-dependent individual tree simulators could be used for optimization of the conversion to uneven-aged stands, no individual tree simulators in European forestry are to our knowledge capable of this at present. The second option is to define specific scenarios and investigate how these scenarios differ in the fulfillment of the management objectives, i.e., simulate how conversion will occur under a variety of conditions. While some studies have used a stand level approach, e.g., based on transition matrixes (e.g., Buongiorno 2001; Schou et al. 2012; Schou and Meilby 2013), a distance-dependent individual tree approach can incorporate spatial interactions among trees, which is important to produce realistic simulation of regeneration (Hyytiäinen & Haight 2012; Vanclay 2012). While, theoretically, distance-dependent individual tree simulators could be used for optimization of the conversion to uneven-aged stands, no available individual tree simulators in European forestry are to our knowledge at present capable of this (see Miina 1996 for optimization in an even-aged setting using individual tree distance-dependent model). Individual tree models have previously been used to simulate the conversion to uneven-aged forest in several studies using, e.g., the distance-independent Heureka (Drössler et al. 2015, 2014), BWINpro (Schröder et al. 2007) and the distance-dependent SILVA (Hanewinkel 2001; Hanewinkel and Pretzsch 2000; Knoke and Plusczyk 2001; Hilmers et al. 2020).
Due to its non-dependence on stand age, as well as its design for multi-criteria scenarios assessment, SILVA is well equipped for the simulation of uneven-aged stand development (Bravo et al. 2019). Since the early conversion studies in SILVA, the simulator has been equipped with a validated regeneration model (Biber and Herling 2002; Pretzsch et al. 2006) as well as calibrated with a large database on complex mixed uneven-aged stands (Hanewinkel and Pretzsch 2000). While Hanewinkel and Pretzsch (2000) looked at different conversion strategies and how these influenced the structural development, the study only did so for the simple case of converting even-aged Norway spruce to uneven-aged and did not include economic aspects. Hanewinkel (2001) and Knoke and Plusczyk (2001) both included an evaluation of the economic consequences of conversion, but they did not investigate different conversion strategies. In addition, no studies have so far quantified the effects of conducting conversion in stands of different age, which is an important factor for success in conversion (Mosandl and Küssner 1999; Schütz 2001). Finally, none of the aforementioned studies included a validation of the simulations with data from long-term mixed uneven-aged experimental plots. Such validation is important due to the complexity of modelling mixed uneven-aged stands (Bravo et al. 2019). Validation was undertaken in Hilmers et al. (2020), which simulated in SILVA the effect of different overall conversion strategies (e.g., using shelterwood and gaps). Hilmers et al. (2020) tested one stand age at conversion and different overall conversion strategies were defined without varying factors such as target diameter harvesting (TDH) rate.
From the synthesis of studies assessing the conversion of even-aged Norway spruce into uneven-aged mixed forests, it follows that the consequences of different conversion methods are not sufficiently investigated for a variety of conditions. It is especially meaningful to simulate the conversion of pure Norway spruce stands to mixed uneven-aged forests of Norway spruce, silver fir (Abies alba) and European beech (Fagus sylvatica), hereafter referred to as spruce, fir and beech. This mixture is of special interest since it combines stability, also towards the predicted climate change (Albrecht et al. 2012; Hilmers et al. 2019; Vitasse et al. 2019), applicability in uneven-aged systems across Europe, and availability of data to validate simulations with (Pretzsch 2019; Pretzsch et al. 2015). Such validation data are available from experimental plots mainly located in the elevational zone of 600–1400 m known as the mixed mountain forest zone (Pretzsch et al. 2015). Furthermore, this mixture would allow keeping a significant percentage of fast growing, valuable conifers, which due to their favorable wood properties will likely continue to be in high future demand, e.g., for construction timber (Ramage et al. 2017; European Commission 2018).
In this study, we explored an array of conversion scenarios from even-aged spruce to uneven-aged forest of spruce, fir and beech. The aim was mainly to analyze and compare the consequences of different combinations of overall conversion strategy, thinning regime, TDH rate and stand age at conversion but also to compare these conversion scenarios to continued even-aged spruce management. Scenarios were compared in terms of stand productivity, economic performance and structural heterogeneity.
Methods
The conversion from even-aged spruce to uneven-aged spruce, beech and fir was simulated with synthetic stands of Norway spruce using the distance-dependent, individual tree simulator SILVA version 2.3 created by the chair of Forest Growth and Yield Science, the Technical University of Munich (Pretzsch et al. 2006). Firstly, we explain how we constructed the initial simulation stands and how we defined the growing conditions. Secondly, we explain the different conversion scenarios, and the reference Norway spruce scenario, we applied to these initial simulation stands. Thirdly, we show the methodology for validating the growth simulations for the conversion scenarios before lastly explaining the response variables used to compare different scenarios.
Construction of simulation stands
As basis for creating the synthetic spruce stands for the simulations, we used data from roughly 2,000 inventory plots (each 500 m2) in the forest district Wasserburg am Inn (approximately 20,000 ha) in Bavaria, Germany. To assess local site quality, we extracted the average height of the five tallest trees (corresponding to the 100 highest trees ha−1), which we used as a proxy for H100 (i.e., the average height of the 100 thickest trees ha−1) and the stand age for each plot. Based on these roughly 2,000 data points, we performed quantile regression (90%) using the nonlinear Chapman–Richards function (with parameter estimates A = 36.00, k = 0.035 and p = 2.034) (Richards 1959) to derive a site index curve for a favorable site. This curve roughly fitted the height increment for site index 36 (H100 = 36 m at index age 100) from the local yield table (Assmann and Franz 1963) and represents a common Bavarian site condition for spruce (Pretzsch 2002). Based on this local yield table, we obtained stem numbers (N), quadratic mean diameter (Dg), dominant height (H100) and height corresponding to quadratic mean diameter (Hg) at stand ages 30 and 60 years (Table 1). These two stand ages represent different starting conditions for the conversion (Fig. 1).

Diameter distribution of the simulation stands at a age 30 b age 60
Based on the stand variables in Table 1, we randomly created six synthetic spruce stands (three stands of age 30 and three stands of age 60) using the stand generator STRUGEN (a tool in the SILVA simulator, see Pretzsch 2002). Each of these six initial stands was subjected to all the treatments investigated in this study. This resulted in three replicates of each treatment, which incorporated the stochasticity from the different modules. This was especially important in regard to the modelling of mortality where the stochasticity can be problematic in individual tree models (Vanclay 2012). The created stands each had an area of 70 m × 70 m, but should be considered as part of a larger spruce forest with surroundings of similar characteristics. The simulated area of 70 m × 70 m was a compromise between having enough space for executing silvicultural operations and processing time.
Specifying growing conditions
In SILVA, species-specific site quality is modelled from a number of site variables resulting in a site specific potential height growth curve. Consequently, we converted the observations of height growth (H100) from Wasserburg am Inn, to potential height growth by multiplying with 1.138 (based on Kahn 1994). Then in SILVA, we manually chose the combination of site variables that most closely fitted this potential height growth curve. The resulting site specification was forest growth region “Isener Altmoräne und Hochterrasse”, at a latitude of 48°, an altitude of 525 m, soil moisture level of 6, soil nutrient level of 5, and no aspect or slope, a realistic site for a spruce plantations in the chosen forest district, especially Ebersberger Forst. Such sites typically have an average annual temperature of app. 7.5 °C, an annual precipitation of app. 900 mm and app. 500 mm of precipitation from May–September according to the 1960–1990 climatic norm that SILVA uses as standard.
After the site conditions were specified, the climatic parameters in SILVA were updated to a 2017 climate norm using the Global Climate model (AOGCMs) and emission scenario RCP 4.5 (IPCC 2013). Specifically, the update was conducted using ClimateEU v4.63 software, available at http://tinyurl.com/ClimateEU using methodology based on Wang et al. (2016). The result was an increase in temperature during May to September of 1.7 degrees and an increase in the vegetation period of 18 days, relative to the 1960–1990 climate norm. This reduced site conditions for all species to site index 31 (H100 = 31 m at index age 100) (reduction was much more substantial for spruce).
Scenario setup
Unless mentioned otherwise, standard SILVA settings were used. Management was assigned on a 10-year basis for simplicity (see Table 6 in Appendix for an example on how simulations were carried out), although SILVA operates on 5-year time intervals. Harvesting is conducted in the beginning of a time period – before mortality and growth. Species composition in natural regeneration was modelled using the tree species share of basal area of adult trees, assuming that seed fall was proportional to the species share of basal area of adult trees. However subsequently, from seed fall to trees grew large enough to enter the growth model (> 7 m height), ecological differences in competition were incorporated (the shade-tolerant beech and fir were able to tolerate higher levels of competition than spruce). All natural regeneration was assumed to occur at no cost.
A simulation period of 150 years was used for assessing consequences of different conversion methods (hereafter named the conversion period). However, to ensure unambiguous economic comparison of alternative scenarios, we employed an, in principle, eternal cash flow of the different management alternatives.
Reference scenarios
The reference scenario (maintaining pure spruce, hereafter referred to as Reference) for the different conversion strategies was simulated in accordance with the guideline for spruce management in Bavaria (BMFAF 1993). In short, the guideline is based on natural regeneration initiated at age 105 and gradual harvesting of the regenerated stand using a TDH criteria within a 20-year period. Reference was slightly altered compared to the guideline (Table 2) because following the guideline resulted in too many trees and a too long retention of the overstorey – likely due to the effect of climate change on spruce growth. In order to complete the harvest of the remaining trees within 20 years, the target diameter (TD) is reduced to 42 cm in the second harvest (10 years after TDH was initiated) and all remaining overstorey trees are removed after 20 years.
Conversion scenarios
Overall, three different conversion strategies were investigated in this study:
- Passive conversion::
-
A passive conversion using a combination of selective thinning and TDH
- Shelterwood conversion::
-
Using a combination of selective thinning and TDH, but conversion is initiated actively with a very heavy, future tree oriented thinning reducing stand density to 300 and 100 trees ha−1 for stands converted at age 30 and 60, respectively
- Gap conversion::
-
Using a combination of selective thinning and TDH, but conversion is also initiated actively by creating a 30 m diameter gap at 5, 25 and 45 years after conversion initiation. The gaps were installed in the stand with centers at these coordinates; (Gap 1: 18.3 m, 18.3 m), (Gap2: 51.67 m, 18.3 m) and (Gap3: 35.0 m, 51.7 m). Gap cutting were programmed in R (R Core Team 2017) and imported into SILVA
For the remainder of the study, these strategies are referred to as Passive, Shelterwood and Gaps. Shelterwood and Gaps are both examples of active conversion strategies.
Within these three overall conversion strategies, TDH cutting rate (how many percent of the trees that have reached TD are harvested) and thinning intensity (using selective thinning) were varied systematically to provide a range of different conversion scenarios (Table 3). Selective thinning specifies thinning around elite trees according to the A-value concept (Johann 1982). Only neighbor trees located beyond a certain critical distance (dependent on the size of the neighbor tree) from the elite tree will not be removed in thinning. This critical distance is given by
where Dist is the distance between the elite center tree C and the neighbor tree N that is evaluated whether it should be thinned or not, HC is the height of the elite tree, DBHC and DBHN is the diameter of the central and neighbor tree, respectively, and A is the A-value. Thus, lower A-value equals heavier thinning. The algorithm keeps selecting elite trees, one by one, and removing that selected tree’s competitors according to the A-value, and continuing to select elite trees until no more competitors can be removed according to the selected A-value. For Passive and Gaps, the effect of only conducting selective thinning until reaching TD and hereafter only conducting TDH was tested (selective thinning was halted once D100 > 40.0 cm). This combination was not meaningful to test in Shelterwood, because preliminary results showed that an even-aged stand would emerge in the next generation unless the understorey was thinned.
Parameters tested included three overall conversion strategies, two different stand ages at conversion and three combinations of selective thinning and TDH cutting rate. Adding to these 3 × 2x3 = 18 scenarios was four scenarios where selective thinning ceased at the initiation of TDH (only for Gaps and Passive for both tested stand ages at conversion). Including Reference at both ages, 24 scenarios emerged.
We assumed all conversion scenarios required artificial regeneration (underplanting) to introduce fir and beech, either in gaps or under shelter/canopy. The regeneration module in SILVA was adjusted to provide 50% fir and beech in a gap when the small trees entered the growth model (> 7 m height). This species share in the underplanting was also used for Shelterwood and Passive in juvenile stands. However, since the underplanting in Shelterwood and Passive was not restricted to a smaller section of the stand like in Gaps, our opinion was that it was less realistic to simulate a complete underplanting of fir and beech below mature spruce stands, since the natural regeneration of spruce could be put to use under such conditions. Thus, here the goal was to have 50% of spruce and 50% fir/beech when the small trees entered the growth model (> 7 m height).
In Shelterwood and Gaps, underplanting was only simulated in the 5-year period when the shelter and gaps were created, while in Passive, underplanting to supplement natural spruce regeneration was continued until H100 ≥ 19 m for one of the underplanted species (either fir or beech). This was to ensure establishment of the underplanting below the denser overstorey in Passive.
Evaluation of growth model
The ability of SILVA to make accurate prediction for uneven-aged spruce fir and beech stands was evaluated with data from experiments in Bavaria collected during the period 1953–2015 (Table 4). The collection included experiments with a long history of selection forest structure (FRY129, see Pretzsch 1985, 2019) and experiments where stands had more recently been converted to a higher structural heterogeneity (MUE145, KRE120-126, RUH110).
Firstly, the ability of SILVA to reflect the effect of growing stock on stand volume production was evaluated by visual comparison of species-specific actual and predicted growth in experiments where stand density reduction was included in the experimental design (experiments FRY129, RUH110 and KRE120-126). Specifically, using data from experiments with rather similar growing conditions to those specified in the simulations of the present study, we plotted species-specific volume production against stand volume for both experimental and simulation data. By this procedure, we aimed to reflect effects of differences in stand density between experiments and simulations in the visual assessment. Mixing proportions were calculated based on the stand density index (SDI, Reineke 1933) and adjusted according to the species-specific growing area requirements by using equivalence coefficients (Pretzsch and Biber 2016). Based on these mixing proportions, individual species growing stock and volume production were scaled to the one ha level. In this analysis, species constituting less than 20% of stand density were excluded from the analysis to avoid scaling error. Similarly, species making up more than 80% of stand density were excluded to compare only stands that had a certain level of mixing.
Secondly, we evaluated the ability of SILVA to replicate individual tree growth in uneven-aged forest using data from the experiment MUE145 (see Utschig 1997 and Schmitt 1994 for detailed description) because of its similarity to our synthetic stands regarding growing conditions, stand history and forest management. In the analysis, spatially explicit, individual tree data from MUE145 were fed into SILVA and predicted growth was compared to actual growth using the lm function in R. For the statistical analysis, we modelled residual dbh and height growth (observed growth on MUE145 plot 1 between 2006 and 2015 minus predicted growth from SILVA) with tree size (dbh and height, respectively) as the predictor variable. We hypothesized that zero slope of the regression line would reflect the ability of SILVA to replicate relative growth across tree sizes, while zero intercept would reflect our ability to replicate overall growing conditions. Since we merely adjusted the site conditions in SILVA according to site information on MUE145 from Table 4, we only assumed zero slope of the regression as our null hypothesis, recognizing that the settings in SILVA were unlikely to match actual site conditions perfectly. While the experimental plot MUE145, along with most of the other mentioned Bavarian experimental plots, was used as database for the parameterization of SILVA in the late 1990s (Kahn and Pretzsch 1997), the growth and yield characteristics in the 20 years afterwards were so far not exploited for model parameterization.
Reference was not validated as the conversion scenarios because the simulator was well calibrated with experimental plot data from even-aged monoculture stands of spruce and can predict the growth of these in Bavaria with a sufficient accuracy (Pretzsch et al. 2002, 2006) (for a general validation of SILVA, see Pretzsch 2002 and Pretzsch et al. 2002). Such a validation was also performed in Hilmers et al. (2020).
Analyses
The different conversion scenarios were compared in terms of growing stock, mean annual increment (MAI) (of wood > 7 cm top diameter limit), mean structural heterogeneity during the 150-year conversion period, and expectation value (EV) (including later periods for the calculation of the eternal rotations part of EV). The growing stock is included in the analysis because it is an important determinant of the other variables used to evaluate conversion, but is itself not used as an evaluation criterion.
Economic yield
SILVA has inbuilt functions for calculating costs and revenues from timber harvesting. Grading for all species were set to mid-diameter grading, calculated with the log grading program BDAT (see Kublin and Scharnagl 1988) integrated into SILVA. Average timber prices for the different assortments were provided by the Bavarian State Institute of Forestry (LWF) for 2010–2014 (see Table 7 in Appendix).
Harvesting costs were calculated using Extended Forest Worker’s Tariff (German: EST, for more information see KWF 2017). The costs are first of all based on time tables that describe the time spend per harvested cubic meter, dependent on species and log dimension (for more information see KWF n.d.). The other important components are the hourly salary (set to 30 €) and the general efficiency of the harvesting (set to 150%), these values are commonly used based on time studies (KWF n.d.). Skidding costs were set to 7.00 € m−3. Regenerations costs for planting were derived from Härtl (2015), which represent typical values for Bavaria. Planting density was set to 4000 trees ha−1 (1.6 m x 1.6 m), amounting to the following values for fir and beech in the different scenarios; 2000 ha−1 in Shelterwood when stand age at conversion was 30 years, 1333 ha−1 in Shelterwood when stand age at conversion was 60 years (not 2000 ha−1 because as mentioned, natural regeneration of spruce is also present), and a total for all three gaps of 867 ha−1 in Gaps. In an attempt to reflect the likely higher costs in small-scale planting (such as underplanting in a gap), the efficiency of underplanting in Gaps was set to only 50% of the efficiency in Härtl (2015). Due to the uncertainty in assessing the regeneration costs of Passive, a sensitivity analysis was used here to construct a range. In the high-cost scenario, planting costs were equal to the most expensive scenario—Shelterwood conversion at age 30 (2000 fir and 2000 beech ha−1), while in the low-cost scenario the costs were based on a Shelterwood conversion using only 750 fir and 750 beech per ha−1.
Having calculated the net return for each period, considering planting costs and net return for timber harvesting, EV for each scenario was calculated as the sum of discounted net returns. We reported the economic results using discount rates of 1 and 3%. In contrast to the other evaluation criteria, EV was calculated for the eternal future time periods. EV of Reference (Eq. 2) was calculated as the discounted net returns for the remainder of the first rotation (depending on whether the stand age at conversion was 30 or 60 years) and (in square brackets) subsequent eternal rotations of the cyclic regime (Faustmann’s “land expectation value”) initiated when the overstorey trees from the first rotation have been harvested and discounted with the length of the first rotation:
where i is the discount rate, s is the stand age at conversion (30 or 60 years), t is the age of the stand during the first rotation, NR(t-s) is the net return in the first rotation obtained at time t-s (years from the initiation of the simulation) and Rr1 is the rotation age of spruce (125 years) in the first rotation. Rotation age, net return and time of subsequent rotations are denoted with an asterisk. The rotation age is slightly longer in the subsequent rotations \({(R}_{r}^{*}=130 years\)) because of the reduction in site index in the present day climate norm (site index 31) compared to the climate under the former historic steady state conditions (site index 36). \({R}_{r}^{*}\) cover the time period between harvesting the last overstorey trees in each rotation. We used the harvesting of the last overstorey trees in our definition of rotation (instead of initiation of regeneration) since the majority of the income was generated during the later regeneration cuttings.
For the conversion scenarios, EV was calculated as the discounted net returns for the 150-year conversion period plus the discounted net returns from eternal rotations of the converted system (square brackets) assumed to have some cyclic nature (Eq. 3). The eternal rotations part of the equation is finally discounted with the length of the conversion period:
where t is the time since the conversion was initiated plus s, NR(tc-s) is the net return obtained at time t-s, i.e., years since the initiation of the conversion, during the 150-year conversion period C. An asterisk denotes the net returns and time of the converted system with a total conversion period of C and a rotation length of R to calculate the cyclic incomes.
Preliminary investigations showed that not all of the conversion scenarios reached a steady state within the 150-year conversion period. Also, the cyclic incomes, assumed to occur when the stand had reached steady state, were not constant for harvestings on a ten-year basis—even when stands had a structure that resembled selection forest. Thus, it was decided that it was more realistic to use the net returns from the period 150–240 years to calculate the cyclic incomes (NR(t*)’s) for the different scenarios (240 years in the future was the technical limit for SILVA). In practice, NR(t*)’s were calculated using only one of the three created stands (as it was rather similar for the three different stands). It is important to emphasize that the period 150–240 years was solely used to calculate a more realistic cyclic income for the eternal rotations. The assessment was generally performed on the 150-year conversion period, and the period 150–240 years was therefore not used in the evaluation of MAI and SPI.
It is also important to remember that the two different stand ages at the time of conversion represent different stands at different age. It should not be understood as a single stand that is either converted at age 30 years, or converted after having waited another 30 years for the stand to become 60 years. Thus, the EV’s from the two different stand ages at conversion are not comparable.
Structural heterogeneity
The structural heterogeneity of the simulated forest was evaluated from the species profile index (SPI) (Pretzsch 1998); a modified version of the Shannon-index, which can also take vertical differentiation within the stand into account:
where S is the number of species in the stand, Z is the number of canopy layers in the stand, and pij is the species proportion (in terms of stem number) of species i in the canopy layer j. The number of canopy layers in the stands is assumed to be three, extending from 0–50%, 50–80% and 80–100% of the maximum height of the stand (Pretzsch 1998). While a highly structured stand, such as a selection forest, will have high SPI approaching 2, very uniform stands will have low SPI approaching 0. Scenarios are compared in terms of the average SPI for all time periods during the 150-year conversion period.
Apart from SPI, the scenarios were also evaluated by comparing the diameter distributions at the end of the conversion period, but this was merely a visual comparison.
Results
Evaluation of growth model
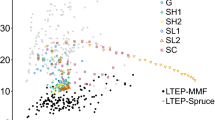
Figure 2 revealed that the volume production of beech was generally less on the experimental plots compared to the simulation while the reverse was true for fir. The slope of the trend between growing stock and volume production for fir was also steeper for the experimental plot data. The simulated volume production was closer to the experimental plot data for Norway spruce than for the other species.

Species-specific relation between growing stock and periodic annual volume production for conversion simulations and experimental plot data (illustrated with simulation stand 1). Results are scaled according to species-specific growing area requirements and scaled up to one hectare. Common y-axis label for each species-specific graph and each point from the conversion simulations represents a simulated period. See Pretzsch (1985, 2019), Pretzsch et al. (2015, 2020) and Hilmers et al. (2019, 2020) for the experimental plot data
For MUE145 plot 1, SILVA was able to correctly specify relative DBH growth between 2006 and 2015 for trees no matter their size (P = 0.8). However, the simulator was not able to predict the corresponding relative height growth (P < 0.01) across tree sizes. Trees below 25 m were generally predicted to grow relatively less than larger trees in the simulation compared to the observation on MUE145.
Effect on growing stock and volume production
The growing stock development illustrates the temporal differences in wood extraction among the different scenarios (Fig. 3). Reference results in a build-up of wood to more than 600 m3/ha at around age 105, when regeneration and harvesting by TD is initiated. This is followed by a sharp decline when the remaining overstorey trees are harvested during the following 20 years. The growing stock in the conversion scenarios generally increases until the stand approaches TD; hereafter, it drops for about 50 years, after which it slowly starts to rise, illustrating the shifting number of trees that reach TD in the different periods. Gaps with only TDH after D100 ≥ 40 cm deviates from the other gap scenarios by having a larger growing stock towards the end of the conversion period. The species-specific graphs in the supplementary materials show the slow build of beech and fir, which is especially slow for fir.

Development of growing stock for simulated scenarios (illustrated with simulation stand 1), split up into overall strategies and stand age at conversion. TDH = target diameter harvesting rate (number in subscript refers to the cutting rate, e.g., 30%). A-value = the A-value component in the selective thinning approach by Johann (1982)
MAI of Reference was 9.0–9.6 m3ha−1 during the conversion period dependent on stand age at conversion (Table 5). All conversion scenarios reduced MAI relative to Reference (6–43%). Reductions were largest when initiating the conversion at age 60 (18–43%) rather than at age 30 (6–36%). Scenarios with heavy thinning produced a lower MAI than the same overall conversion strategy with moderate thinning (5–22%). Adjusting TDH cutting rate only influenced MAI to a smaller degree (≤ 3%). Excluding thinning operations after the trees reached their TD increased MAI (2–23%). Gaps resulted in roughly the same MAI as Shelterwood and Passive.
Effect on expectation value
Using a discount rate of 1%, Reference resulted in higher EV relative to all conversion scenarios no matter if conversion was initiated at age 30 or 60 (Table 5). Similarly, when stand age at conversion was 30 years and at a discount rate of 3%, Reference resulted in higher mean EV than all conversion scenarios. Oppositely, when stand age at conversion was 60 years and at a discount rate of 3%, Shelterwood and Gaps (and some Passive scenarios when low regeneration costs were assumed) resulted in higher EV than Reference.
Compared to the other conversion strategies, EV was generally higher for Gaps, unless low regeneration costs were assumed in Passive. Shelterwood generally resulted in lowest EV, except when stand age at conversion was 60 years, evaluated with 3% discount rate, where Passive resulted in the lowest EV (even when assuming low regeneration costs) and Shelterwood were similar to Gaps.
At a discount rate of 1%, early, heavy thinning decreased EV for all conversion strategies and both stand ages at conversion, while at a discount rate of 3%, the results were more mixed: 1) When stand age at conversion was 60 years, heavy thinning resulted in similar or higher EV, 2) when stand age at conversion was 30, heavy thinning decreased EV for all treatments compared to lighter thinning scenarios, except for Shelterwood. Scenarios without thinning after the trees had reached TD typically had a slightly lower EV, apart from in Gaps when stand age at conversion was 60 (and discount rate 1%). Differences between unthinned and thinned scenarios were however usually rather small, except in Passive when stand age at conversion was 60 years.
Increased TDH cutting rate always resulted in an increase in EV. The effect was typically largest when using 1% discount rate. EV of Gaps was generally not much affected by increasing TDH cutting rates compared to Shelterwood. For Passive, EV was not substantially affected when stand age at conversion was 30 years, but more so when stand age at conversion was 60 years.
Effects on structural heterogeneity
Reference resulted in a low SPI compared to all the conversion scenarios. Among the conversion scenarios, Gaps generally resulted in a higher SPI compared to Shelterwood and Passive. This difference was especially apparent when stand age at conversion was 30 years.
Heavily thinned scenarios generally had higher SPI than moderately thinned scenarios, except for Gaps when stand age at conversion was 60 years. TDH cutting rates did not generally influence SPI, although a slight positive effect was observed in Passive. Excluding thinning after stands had reached TD decreased SPI in Passive and seemed to slightly increase SPI in Gaps. However, this increase in structural heterogeneity in Gaps was not confirmed when looking at the diameter distributions (Fig. 4a and b). SPI was generally higher when stand age at conversion was 60 years.

Diameter distribution after 150 years when stand age was 30 years at conversion, using the overall conversion strategy of gaps and target diameter harvesting was 50% a) with selective thinning b) without selective thinning
The structural development followed very different patterns in the three overall conversion strategies (Fig. 5) and were influenced by stand age at conversion. When stand age at conversion was 30 years, Gaps and Shelterwood resulted in a rapid initial increase in SPI, just 20 years after conversion was initiated. The structure in Gaps became increasingly more heterogeneous during the first 90 years after the start of the conversion, while large increases in heterogeneity started after 90 years in Shelterwood and in Passive. After 120 years of conversion, SPI was roughly similar in the different overall conversion strategies. When stand age at conversion was 60 years, the time before high SPI was reached in Shelterwood, and especially in Passive, was shorted considerably. Generally, we observed a strong increase in the beech proportion of the growing stock towards the end of the conversion period. While the stand percentage composed of beech and fir were practically similar during the early stages (as well as very low compared to spruce), stands were on average composed of 48% beech and 14% fir after 150 years at the end of the conversion period. The proportion of beech continued to increase during the 90-year interval after the end of the conversion period (between 150 and 240 years after conversion was initiated), accompanied by a decline in the proportion of conifers, driven by a decrease in the proportion of spruce.

Average species profile index in the different conversion scenarios, when target diameter harvesting rate was 50% and selective thinning degree was according to A-value = 6 (see Johann (1982)). For stand age at conversion a 30 years, b 60 years
Discussion
Conversion effects on volume production
Due to the heavy initial stand density reduction, all conversion scenarios reduced MAI relative to Reference. This was also generally observed in other simulation studies using dedicated gap cutting in SILVA (Hanewinkel and Pretzsch 2000) or heavy stand density reduction to initiate conversion in the simulator Heureka (Drössler et al. 2014). Both studies simulated conversion from even to uneven-aged spruce. Seemingly, our results were quite similar to these studies because the altered climatic variables in our study reduced spruce growth to be rather similar to fir and beech growth. Drössler et al. (2014) found similar growth for conversion scenarios with the same growing stock whether thinning were differentiated spatially across the stand or applied homogeneously, but this could have been due to the use of the distance-independent Heureka simulator. This highlights the benefit of distant dependent individual tree models for conversion simulation, especially in our situation where quite contrasting conversion treatments were applied, and especially gap cutting and TDH created spatial heterogeneity.
Using SILVA, Hanewinkel and Pretzsch (2000) found that passive conversion by TDH did not have any practical influence on MAI, relative to traditional spruce management, while scenarios involving harvesting of gaps resulted in a considerable reduction in MAI. Contrasting this finding, Passive conversion using only TDH after D100 ≥ 40 cm (the scenario that most closely mimicked the only TDH scenario in Hanewinkel and Pretzsch 2000) resulted in reduced MAI compared to Reference. Our analysis indicated that the longer simulation period in our study (150 years) compared to the 100-year period used in Hanewinkel and Pretzsch (2000) reduced growth since overstorey trees retained to old age had slower growth. Furthermore, in Hanewinkel and Pretzsch (2000), gaps had to be very large to accommodate spruce regeneration establishment. This resulted in large growth reduction compared to gap creation with fir and beech in our study, which required smaller gaps to ensure successful establishment in accordance with differences in light demand between spruce and fir/beech. This is in line with Hilmers et al. (2020) who, based on modelling in SILVA, demonstrated that smaller gap sizes are sufficient if shade-tolerant species are involved in the transformation process by planting or from natural regeneration if potential mast trees are located nearby. It is also in line with Brunner et al. (2006), who simulated a transformation of spruce stands in SILVA by under-planting with beech in a gradually opened stand of spruce. Thus, of interest for practice, our results indicate that minor gaps do not necessarily result in a substantial growth reduction. Actually, Gaps generally produced a larger MAI than Passive in the simulations.
Although gap scenarios also achieved relatively high MAI in Hilmers et al. (2020) and Knoke and Plusczyk (2001), the relative production of Passive and Gaps is surprising, since the early cutting of gaps to benefit regeneration is normally expected to reduce production of overstorey trees. Contrasting the simulations in SILVA, results from mixed mountain forest experiment such as KRE120-126 and RUH110 suggested that cutting of gaps strongly decrease productivity, although such experiments have only been performed in old stands (> 100 years old).
How substantial MAI losses will be from cutting Gaps in younger stands is unknown. The effect of gap creation on stand productivity has, in comparison with the effect of species diversity, only been little investigated (Bourdier et al. 2016). Our results might partly be due to the favorable conditions for regeneration in small gaps (Brunner and Huss 1994). The reduction in MAI relative to Reference was more pronounced for strategies that involved heavy and late stand density reductions than for scenarios with early and more moderate interventions. We did not find any substantial effects on MAI when varying the TDH harvesting rates in the conversion scenarios. In comparison, Brunner et al. (2006) compared TDH using different TDH limits following shelterwood and found substantial differences in overstorey growth but these results are incomparable to our study because Brunner et al. (2006) did not include understorey growth.
Few studies have compared growth across different conversion scenarios, and none have involved different ages for the onset of conversion. However, the observed mechanisms are biologically meaningful (e.g., Bryndum 1980 finds that thinning reduces growth more at older age).
The transferability of our results for conversion compared to Reference to other site conditions is especially affected by the site-specific relative productivity between spruce, beech and fir. Under warmer and drier conditions, Norway spruce will be less competitive relative to beech and fir. The relative performance of the different conversion scenarios is less affected by site conditions, or at least in terms of how site conditions are specified in SILVA. Nevertheless, a warmer and drier climate will result in more reduced growth at the later development stages (Pretzsch 2009), pointing to lower optimal TD as well as a higher rate of TDH. In our case, spruce growth was already substantially reduced from our climate adjustment compared with Huang et al. (2017) and Pretzsch et al. (2014).
Effects of conversion on the expectation value
Conversion generally resulted in reductions of EV, most likely mainly as a result of premature harvest of trees at the onset of the conversion and reduced growth of the remaining stand. The exception was when stand age at conversion was 60 and using a discount rate of 3%.
Several studies have found that increasing the discount rate increases the profitability of conversion scenarios relative to traditional management as a consequence of increased harvesting during the early conversion phase (Hanewinkel 2001; Knoke and Plusczyk 2001; Price and Price 2006). In line with these results, our study showed that active scenarios were relatively more profitable than Passive and Reference, when stand age at conversion was 60 years and using a discount rate of 3%. When stand age at conversion was 30 years, the revenue from early heavy harvesting to initiate conversion was apparently not enough to compensate for the expensive underplanting and loss of growth and later harvesting opportunities.
A general concern when initiating conversion at later stages is that the proportion of trees that are harvested later than their financially optimal time of cutting may be relatively large (Knoke 2009) rendering the conversion at old age less profitable. However, in our study the stumpage price curve obtained from the Bavarian State forest (see Table 7 in Appendix) did not dictate a significant increase in the price per unit for increasing diameter beyond 25 cm. Also, MAI during the individual rotations in Reference reached maximum when Dg reached app. 34 cm. Consequently, the optimal DBH at harvesting would likely be less than the TD of 45 cm used for spruce, especially for large discount rates (Meilby and Nord-Larsen 2012). When initiating conversion at age 60 years (see Table 1), a large part of the trees felled would therefore be at or close to the optimal harvesting diameter increasing the relatively profitability of active conversion at high discount rates. It can be argued that a smaller TD should have been used in all scenarios (both for Reference and conversion) given the underlying stumpage prices. This would have resulted in earlier initiation of TDH, which would have benefited Reference more than the conversion scenarios because of the higher growing stock in Reference when TD is reached. This illustrates the importance of stumpage prices when comparing conversion and traditional management (Knoke et al. 2001). It also illustrates an issue in economic evaluation of traditional management vs. conversion that has previously been raised (Price 2003): is it meaningful to consider conversion at high discount rates as superior when it is merely caused by overly long rotation periods resulting from silvicultural tradition? This point seems to be valid for Bavarian conditions. This also highlights that a secondary TD for spruce, e.g., 35 cm, which would have resulted in a significant reduction in rotation length, should be considered in studies analyzing the advantageousness of conversion.
A number of practical assumptions made in our study may have affected the relative performance of the Reference and conversion scenarios. In general, we assumed that wood quality resulting from the different scenarios would be similar. Although this may not be true, the combined effect of time required for this to manifest itself in the economic calculations and the discounting, likely render this of minor economic importance (Schou et al. 2012). We also made assumptions about the regeneration costs and as could be seen from the sensitivity analysis in Passive, regeneration costs could have a considerable effect on the results. However, our assumptions likely did not heavily influence the relative results between scenarios. Gaps was penalized in terms of planting costs due to the assumption of no use of natural regeneration in the initial plantings when stand age at conversion was 60 and twice as expensive planting costs per plant due to the small scale planting. Still, this scenario came out with an overall better EV result than Shelterwood. The relative ranking between Passive and Gaps was to some degree affected, but the regeneration costs in Passive were in any case highly uncertain.
More importantly, we also considered harvesting costs fixed across different thinning types and stand structures similarly to other studies (e.g., Hanewinkel 2001 and Knoke and Plusczyk 2001). Studies have shown that this is probably not a realistic assumption and have found that harvesting costs differ in the scale of roughly 2.5–5 €/m3 across different harvesting regimes and generally find that clearcut is the most efficient harvesting technique (Pausch 2005; Price and Price 2008, 2006). To illustrate the effect of differences in harvesting costs, consider a conversion scenario with high EV (Gaps, A-value6, TDH50). For this scenario, penalizing harvest costs in uneven-aged forests would likely reduce EV 1,600- 3,300 € using 1% discount rate and 400–1000 € using 3% discount rate (almost similar for both stand ages at conversion). Such an assumption could be realistic across most conversion scenarios since harvesting costs between different uneven-aged forest systems (group selection and single tree selection) have been found to be rather similar (Suadicani and Fjeld 2001). This would not have a large impact on the results, although if assuming the highest penalty for harvesting in uneven-aged forest, Reference would be more at level with the most profitable conversion scenarios for stand ages at conversion of 60 years and using a discount rate of 3%.
The effect of different natural hazards (especially wind throw) on the economic output in practice often exert large influence on the choice of conversion method. Nonetheless, longevity of overstorey trees and risk of windthrow was not included in this study despite being a severe constraint to conversion efforts on many sites in Europe. Comparison to MUE145 and regional survival probability curves in Neuner et al. (2015) shows that the predicted longevity of overstorey trees during a conversion scenario could be realistic for the simulated site. This is especially because spruce in mixed stands have higher survival probability, compared to pure stands, at older age (see also Griess et al. 2012), which could have implications during the later stages of conversion. However, the longevity of overstorey spruce would be much less on more shallow rooted clayey sites in many regions of Europe (Henriksen 1988), especially in windy climates. Wind throw is often the cause of stand destruction following shelterwood thinning in spruce (Löf et al. 2010; Reventlow 2020). Experiments have also found that heavier shelterwood thinning down to 10 m2 ha−1 further increases the risk of wind throw (Reventlow 2020). In relation to our study, including the risk of windthrow may reduce the expectation value of the conversion scenarios substantially. However, large parts of central Europe experience significantly lower wind speeds compared to more oceanic areas (Troen and Petersen 1989). This explains why conversion efforts can take place at older age in such regions, e.g., in the Munich area as described in Matthes and Ammer (2000). While shelterwood thinning in stands taller than 18 m is considered very risky in oceanic Europe (Larsen and Skov- og Naturstyrelsen 2005), such operations are recommended in stands higher than 25 m for areas situated in 500–1400 m altitude in Bavaria (Bayerische Staatsforsten AöR 2018). At present, a wind risk model for use in complex forest (based on the work of Hale et al. 2012, 2015) is undergoing testing. At the time when this study was conducted, the model was still under development, but this model has a large potential to make simulation studies more realistic, especially for more wind exposed areas.
Recent studies have criticized the use of only net present value (and supposedly EV), for the estimation of the economic value of different land use alternatives (Knoke et al. 2020). The argument is especially that flexibility (e.g., of liquidity) is not given any weight, although studies have shown flexibility to be of high importance for landowners in decision making (Yemshanov et al. 2015). Uneven-aged stands is often considered to have higher cash flow flexibility than even-aged stands (Nord-Larsen et al. 2003), unless forest owners manage to create a normal forest, which is rarely achieved (Knoke 2012). However, conversion to uneven-aged stands also represents a long-term investment decision, especially when expensive underplanting is carried out, which can be considered a flexibility constrain on the forest owner.
Structural heterogeneity
Our results demonstrated large differences in the time required to convert even-aged spruce to uneven-aged spruce, fir and beech depending on the applied strategy, especially when initiating conversion at age 30. Similar results were derived for conversion of spruce plantations to uneven-aged spruce, fir and beech when stand age at conversion was 30 years in Hilmers et al. (2020) and to uneven-aged spruce in Hanewinkel and Pretzsch (2000), although the latter study did not include a shelterwood scenario. The faster structural development in Shelterwood when initiating conversion at age 60 may, at least partly, be attributed to our defined scenarios. For conversion at age 60, we allowed natural regeneration of spruce to be incorporated in the underplanting below the Shelterwood. Thus, according to the definition of SPI, this raises the structural development.
The uniform shelterwood is a common outset for conversion in spruce (Brunner et al. 2005; Schütz 2001) and has been recommended for a range of stand conditions (e.g., Larsen and Skov- og Naturstyrelsen 2005 and Nyland 2003). Our results question the use of uniform shelterwood thinning as a method to achieve large structural heterogeneity rapidly, at least in young stands. Here, Shelterwood had a tendency to create quite homogeneous regeneration, subsequently resulting in a homogeneous two-storied stand for a long time (roughly 100 years). This observation is in line with observations made in practical forestry and the lack of differentiation in many shelterwood systems is often the reason why conversion to uneven-aged stands fails in practice (Matthes and Ammer 2000; Schütz 2001). However, shelterwood conversion may be advantageous in situations where conditions for regeneration are harsh, especially due to late spring frost, high browsing pressure or heavy competition from weeds. Under such difficult conditions, observations show that gaps of 30 m diameter can cause high mortality among underplanting of susceptible species (Reventlow 2020). Furthermore, Shelterwood allows for a fast change in species composition compared to Passive and Gaps, which can be beneficial when converting spruce stands that are unstable due to, e.g., bark beetle attacks.
The slow development of SPI in Passive was caused by the time required for the regeneration to establish. It took shorter time before TDH was initiated when stand age at conversion was 60 years, increasing mean heterogeneity. Despite our results, it seems possible to achieve acceptable underplanting establishment without having to reduce the shelter density to the degree visualized in Shelterwood (Pretzsch 2019; Schmitt 1994; Utschig 1997) and hence to achieve larger heterogeneity earlier in Passive. Conversion using a passive approach actually has the potential to provide faster structural heterogeneity than uniform shelterwood, since thinning is more spatially differentiated and results in a wider diameter distribution in the overstorey. This is also illustrated from the high SPI in Passive when used in combination with heavy thinning. But this potential is however dependent on a spatially heterogeneous thinning and continuous regeneration, which both somewhat lacked in our study. It should also be mentioned that denser overstorey (which is typically the case for passive conversion) reduces height growth of the underplanting, slowing conversion (Reventlow 2020). This is also clear from MUE145 where conversion is clearly progressing faster in the plot where the most intensive shelter density reduction was undertaken (Pretzsch 2019).
In general, thinning was needed to ensure long-term heterogeneity—even in Gaps. Using only TDH, the growing stock builds up as beech and fir grows without being thinned until they finally reach the TD after a long time (after 180 years). The same growing stock build-up pattern as in Gaps was not observed in Passive because the establishment of fir and beech was much less complete and the regeneration had not had a long time to develop at the end of the conversion period (but SPI was still low due to the long time before regeneration was established). While Gaps using only TDH had a high SPI, comparison with the diameter distributions showed very limited ingrowth.
This also illustrates a flaw of SPI, since all tree heights below 50% of max tree height in the stand were treated similarly. Still, the problem with other structural indices such as the Gini index (Gini 1912) and the diameter differentiation index (Gadow and Füldner 1995) was that they give weight to extremes more than they give to variation. As an example, a stand with recent shelterwood thinning would have a higher diameter differentiation index and Gini index compared to a selection forest (since the indices responds mostly to observations that are far apart, such as a small tree in the understorey and a large tree in the overstorey), although the selection forest is clearly more heterogeneous.
The marked increase in beech share of the growing stock towards the end of the conversion period (and beyond) was due to the complete dominance of beech in the predicted natural regeneration (after the underplanting had reached maturity and started bearing seeds). The beech dominance in the simulated natural regeneration seemed excessive at first. However, comparison with experimental plot data from KRE120 and FRY129 showed that such dominance can be expected. This is despite that these experiments are located at higher elevation having higher water supply and lower temperature than the simulated sites, meaning that spruce has a competitive advantage (Pretzsch 2009). Practical guidelines for the Bavarian alps up to 1200 m altitude also suggest that frequent pre-commercial thinning is needed to sustain both species of conifers because of the strong competitiveness of beech regeneration (Bayerische Staatsforsten AöR 2018). Thus, for most sites and many types of silvicultural systems, it will be a struggle against nature to introduce and keep fir in a mixed forest system with beech.
Evaluation of SILVA simulations and procedure
Some deviations were expected for the comparison of species-specific volume production across different growing stocks since both treatments and growing conditions differed somewhat between experimental and simulated stands. The slightly colder conditions (Table 4) lead to lower growth of beech on the experimental plots at a certain growing stock. Conversely, silver fir growth seems to be under-predicted by SILVA, as previously reported (Mette et al. 2009), likely due to the smaller database used for the parameterization of this species. In contrast, spruce volume growth was well predicted, probably because of the large database used for parameterization (Pretzsch et al. 2002). These results imply that the simulated growth dynamics could be realistic for beech and spruce but with some deviations for silver fir.
We made no attempt to conduct a formal validation of the regeneration and mortality modules in Silva due to the erratic nature of seedling establishment and mortality (Vanclay 2012) as well as the sparse data available on regeneration and mortality in such development stages. However, we observed that when shelter basal area exceeded roughly 12 m2ha−1, predicted survival of the underplanting was almost none. This is in contrast to high survival rates observed for beech and fir planted below spruce shelter having basal areas exceeding 20 m2ha−1 (Ammer and Weber 1999; Pretzsch et al. 2015; Reventlow 2020). We suspect that this relation between stand density and regeneration survival could be one reason why productivity is relatively high in Gaps relative to Passive due to the lack of regeneration in Passive, resulting in less growth.
The predicted regeneration mortality in SILVA at higher basal areas had implications for the design of the conversion scenarios, which had to accommodate the successful establishment of regeneration. Hence, the stand density reduction applied in some of the conversion scenarios (especially Shelterwood) exceeded what would be necessary in practice (see, e.g., Utschig 1997 and Pretzsch 2019).
Whether a 150-year conversion period can be considered sufficient to incorporate the effect of different conversion treatments is an important discussion. Firstly, the investigated treatments were initiated early following conversion. Treatments were initiated no later than 65 years after initiation of conversion (TDH, when stand age at conversion was 30 years). Furthermore, SPI generally reached steady state within the 150-year conversion period. What was not reached was, as already mentioned, equilibrium in terms of species distribution. From that perspective, a longer conversion period would have been preferable. Our approach is relatively long compared to other studies (Hanewinkel 2001; Hanewinkel and Pretzsch 2000; Hilmers et al. 2020; Knoke and Plusczyk 2001), especially considering that we also used some data from 150–240 years for calculating EV.
Conclusion
Simulation is by definition a simplification of the real world in terms of variables that affects stand development. Hence, the challenge of this study was to produce consistent, scientific but also relevant and realistic simulations across a variety of treatments when converting plantations to heterogeneous forest. Despite limitations, the conducted simulations represent an overview on how different factors influence the expected trajectories of conversion. The results indicated that especially creation of several gaps combined with moderately heavy thinning, and a high rate of target diameter harvesting resulted in a favorable combination of high productivity and economic yield (across tested discount rates) and fast development of high structural heterogeneity. On the other hand, shelterwood conversion generally resulted in inferior structural development and economic results, especially when conducted at an early stage. Thus, shelterwood conversion should mainly be used in conversion situations where a fast change of tree species is a necessity. Using a more passive conversion was harder to evaluate, because the used simulators prediction of regeneration was sensitive to overstorey density. But the economic result was generally not better than in gaps, and structural development would be slower.
In order to simulate scenarios more realistically, especially on sites with larger wind exposure than Central Europe, growth models and risk tools for natural hazards should be linked together more closely than was possible in our study. Furthermore, it is important that more research is devoted to increase the accuracy of regeneration models since regeneration is a crucial part of the conversion process.
Data availability
Readers may contact the corresponding author for getting access to data.
Code availability
Readers may contact the corresponding author for getting access to simulator and code.
References
Albrecht A, Hanewinkel M, Bauhus J, Kohnle U (2012) How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modeling based on long-term observations. Eur J For Res 131:229–247
Ammer C, Weber M (1999) Impact of silvicultural treatments on natural regeneration of a mixed mountain forest in the Bavarian alps. In: Olsthoorn AFM, Bartelink HH, Gardiner JJ, Pretzsch H, Hekhuis HJ, Franc A, Wall S (ed.) Management of mixed species forest: silviculture and economics. DLO Institute for Forestry and Nature research (IBN-DLO). IBM Scientific contributions 15, Wageningen, pp 68–78
Assmann E, Franz F (1963) Vorläufige Fichten—Ertragstafel für Bayern in Bayerische Staatsministerium pur Ernahrung. Landwirtschaft und Forsten (Hrsg) 19901
Bayerische Staatsforsten AöR (2018) Waldbauhandbuch Bayerische Staatsforsten Richtlinie für die Waldbewirtschaftung im Hochgebirge. https://www.baysf.de/fileadmin/user_upload/04-wald_verstehen/Publikationen/WNJF-RL-006_Bergwaldrichtlinie.pdf
Biber P, Herling H (2002) Modellierung der Verjünungsdynamik als Bestandteil von einzelbaumorientierten Waldwachstumssimulatoren. Sekton Ertragskunde DVFF, Schwarzenbach, pp 194–216
BMFAF (Bavarian Ministry for Food, Agriculture and Forests) (1993) Grundsätze für die waldbauliche Behandlung der Fichte im Bayerischen Staatswald. LMS F 4-W
Bourdier T, Cordonnier T, Kunstler G, Piedallu C, Lagarrigues G, Courbaud B (2016) Tree size inequality reduces forest productivity: an analysis combining inventory data for ten european species and a light competition model. PLoS ONE. https://doi.org/10.1371/journal.pone.0151852
Bravo F et al (2019) Modelling approaches for mixed forests dynamics prognosis. Forest Systems, Research gaps and opportunities. https://doi.org/10.3929/ethz-b-000349182
Brunner A, Hahn K, Biber P, Skovsgaard JP (2006) Conversion of Norway Spruce: A Case Study in Denmark Based on Silvicultural Scenario Modelling. In: Hasenauer H (ed) Sustainable Forest Management: Growth Models for Europe. Springer, Berlin, Heidelberg, pp 343–371
Brunner A, Huss J (1994) Die Entwicklung von Bergmischwaldkulturen in den Chiemgauer Alpen. Forstwissenschaftliches Centralblatt vereinigt mit Tharandter forstliches Jahrbuch 113(1):194–203
Brunner A, Sørensen IH, Skovsgaard JP (2005) Skærmstilling og underplantning af rødgran i Gludsted plantage. Forsøg nr. 1512. Anlægsrapport nr. 603 Skov & Landskab, Hørsholm Arbejdsrapport nr. 13–2005
Bryndum H (1980) Bøgehugstforsøget i Totterup skov. Det Forstlige Forsøgsvæsen i Danmark 38:1–76
Buongiorno J (2001) Quantifying the implications of transformation from even to uneven-aged forest stands. For Ecol Manage 151:121–132. https://doi.org/10.1016/S0378-1127(00)00702-7
Christiansen E, Bakke A (1988) The spruce bark beetle of Eurasia. In: Alan A. Berryman (Ed.) Dynamics of forest insect populations. Springer, pp 479–503
Delatour C, Weissenberg, K von,Dimitri, L (1998) Host Resistance. In: Woodward S, Stenlid J, Karjalainen R., Hüttermann, A (ed.) Heterobasidion annosum: Biology, ecology, impact, and control. CABI, pp 143–166
Drössler L, Overgaard R, Eko PM, Gemmel P, Bohlenius H (2015) Early development of pure and mixed tree species plantations in Snogeholm, southern Sweden. Scand J For Res 30:304–316. https://doi.org/10.1080/02827581.2015.1005127
Drössler L, Nilsson U, Lundqvist L (2014) Simulated transformation of even-aged Norway spruce stands to multi-layered forests: an experiment to explore the potential of tree size differentiation. Forestry: An International Journal of Forest Research, 87:239–248.
European Commission (2018) A sustainable Bioeconomy for Europe: strengthening the connection between economy, society and the environment. Updated Bioeconomy Strategy. https://ec. europa.eu/research/bioeconomy
Gini C (1912) Variabilità e mutabilità. Reprinted in Memorie di metodologica statistica (Ed Pizetti E, Salvemini, T) Rome: Libreria Eredi Virgilio Veschi
Griess VC, Acevedo R, Härtl F, Staupendahl K, Knoke T, (2012) Does mixing tree species enhance stand resistance against natural hazards? A case study for spruce. Forest Ecology and Management 267: 284–296
Hale SE, Gardiner BA, Wellpott A, Nicoll BC, Achim A (2012) Wind loading of trees: influence of tree size and competition. Eur J Forest Res 131(1):203–217. https://doi.org/10.1007/s10342-010-0448-2
Hale SE, Gardiner BA, Peace A, Nicoll BC, Taylor P, Pizzirani S (2015) Comparison and validation of three versions of a forest wind risk model. Environ Model Softw 68:27–41.
Hanewinkel M (2001) Economic aspects of the transformation from even-aged pure stands of Norway spruce to uneven-aged mixed stands of Norway spruce and beech. For Ecol Manage 151:181–193. https://doi.org/10.1016/s0378-1127(00)00707-6
Hanewinkel M, Pretzsch H (2000) Modelling the conversion from even-aged to uneven-aged stands of Norway spruce (Picea abies L. Karst.) with a distance-dependent growth simulator. For Ecol Manage 134:55–70. https://doi.org/10.1016/S0378-1127(99)00245-5
Hekhuis H, Wieman E (1999) Economics and management: costs, revenues and function fulfilment of nature conservation and recreation values of mixed, uneven-aged forests in The Netherlands. IBN Scientific Contributions (Netherlands)
Henriksen H (1988) Skoven og dens dyrkning. Dansk Skovforening, Nyt Nordisk Forlag Arnold Busck, Copenhagen
Hilmers T et al. (2019), The productivity of mixed mountain forests comprised of Fagus sylvatica, Picea abies, and Abies alba across Europe. Forestry: An International Journal of Forest Research, 92:512–522.
Hilmers T, Biber P, Knoke T, Pretzsch H (2020) Assessing transformation scenarios from pure Norway spruce to mixed uneven-aged forests in mountain areas. Eur J Forest Res. https://doi.org/10.1007/s10342-020-01270-y
Huang W, Fonti P, Larsen JB, Ræbild A, Callesen I, Pedersen NB, Hansen JK (2017) Projecting tree-growth responses into future climate: A study case from a Danish-wide common garden. Agric For Meteorol 247:240–251. https://doi.org/10.1016/j.agrformet.2017.07.016
Hyytiäinen K, Haight RG (2012) Optimizing continuous cover forest management. In: Pukkala T, von Gadow K (eds) Continuous Cover Forestry. Springer, Netherlands, Dordrecht, pp 195–227
Härtl FH (2015) Der Einfluss des Holzpreises auf die Konkurrenz zwischen stofflicher und thermischer Holzverwertung. Technische Universität München
Messerer et al., 2017IPCC (2013) Climate change 2013: the physical science basis. Working Group I: Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate change. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9781107415324
Johann K (1982) Der A-Wert ein objektiver Parameter zur Bestimmung der Freistellungsstärke von Zentralbäumen. Deutscher Verband Forstlicher Forschungsanstalten–Sektion Ertragskunde 25:27
Kahn M (1994) Modellierung der Höhenentwicklung ausgewählter Baumarten in Abhängigkeit vom Standort. Forstlische Forschungsberichte München 141
Kahn M, Pretzsch H (1997) Das Wuchsmodell SILVA-Parametrisierung der Version 2.1 für Rein-und Mischbestände aus Fichte und Buche. Allgemeine Forst-und Jagdzeitung 168:115–123
Kenk G, Guehne S (2001) Management of transformation in central Europe. For Ecol Manage 151:107–119. https://doi.org/10.1016/s0378-1127(00)00701-5
Knoke T (2009) Zur finanziellen Attraktivität von Dauerwaldwirtschaft und Überführung: eine Literaturanalyse. Schweiz Z Forstwes 160:152–161
Knoke T (2012) The Economics of Continuous Cover Forestry. In: Pukkala T, von Gadow K (eds) Continuous Cover Forestry. Springer, Netherlands, Dordrecht, pp 167–193
Knoke T, Gosling E, Paul C (2020) Use and misuse of the net present value in environmental studies. Ecol Econ 174:106664. https://doi.org/10.1016/j.ecolecon.2020.106664
Knoke T, Moog M, Plusczyk N (2001) On the effect of volatile stumpage prices on the economic attractiveness of a silvicultural transformation strategy. Forest Policy Econ 2:229–240. https://doi.org/10.1016/S1389-9341(01)00030-2
Knoke T, Plusczyk N (2001) On economic consequences of transformation of a spruce (Picea abies (L.) Karst.) dominated stand from regular into irregular age structure. For Ecol Manage 151:163–179. https://doi.org/10.1016/S0378-1127(00)00706-4
Kublin E, Scharnagl G (1988) Verfahrens-und Programmbeschreibung zum BWI-Unterprogramm BDAT. Forstliche Versuchs-und Forschungsanstalt Baden-Württemberg
KWF (Kuratorium für Waldarbeit und Forsttechnik e.V.) (2017) Geldtafeln zum EST 2017. https://www.kwf-online.de/index.php/wissenstransfer/waldarbeit/311-fortschreibung-der-est-geldfaktoren. Accessed 11 june 2018
KWF (Kuratorium für Waldarbeit und Forsttechnik e.V.) (n.d.) Zeitbedarfstafeln für die motormanuelle Holzernte. http://www.kwf-online.org/fileadmin/dokumente/Mensch_Arbeit/TdL/Lohnentwicklung/MoFz/zeitbedarfstafeln.pdf. Accessed 11 june 2018
Larsen JB (2012) Close-to-nature forest management: the Danish approach to sustainable forestry. In: Casero JJD (ed) García JM. Sustainable Forest Management, Current Research. Intech, pp 199–218
Larsen JB, Skov- og Naturstyrelsen, (2005) Naturnær skovdrift – idekatalog til konvertering. Mijøministreriet, København
Löf M, Bergquist J, Brunet J, Karlsson M, Welander NT (2010) Conversion of Norway spruce stands to broadleaved woodland-regeneration systems, fencing and performance of planted seedlings. Ecological bulletins:165–174
Matthes U, Ammer U (2000) Conversion of Norway spruce (Picea abies L.) stands into mixed stands with Norway spruce and beech (Fagus sylvatica L.)–Effects on the stand structure in two different test areas. In: Klimo E, Hager H, Kulhavý (eds) Spruce Monocultures in Central Europe Problems and Prospects. European Forest Institute, Joensuu, pp 71–80
Meilby H, Nord-Larsen T (2012) Spatially explicit determination of individual tree target diameters in beech. For Ecol Manage 270:291–301. https://doi.org/10.1016/j.foreco.2011.08.037
Messerer K, Pretzsch H, Knoke T (2017) A non-stochastic portfolio model for optimizing the transformation of an even-aged forest stand to continuous cover forestry when information about return fluctuation is incomplete. Annals For Sci. https://doi.org/10.1007/s13595-017-0643-0
Mette T, Albrecht A, Ammer C, Biber P, Kohnle U, Pretzsch H (2009) Evaluation of the forest growth simulator SILVA on dominant trees in mature mixed Silver fir–Norway spruce stands in South-West Germany. Ecol Model 220(13–14):1670–1680
Miina J (1996) Optimizing Thinning and Rotation in a Stand of Pinus Sylvestris on a Drained Peatland Site. Scand J For Res 11:182–192. https://doi.org/10.1080/02827589609382927
Mosandl R, Küssner R (1999) Silviculture: conversion of pure pine and spruce forests into mixed forests in eastern Germany: some aspects of silvicultural strategy. IBN Scientific Contributions (Netherlands)
Neuner S et al (2015) Survival of Norway spruce remains higher in mixed stands under a dryer and warmer climate. Glob Change Biol 21:935–946. https://doi.org/10.1111/gcb.12751
Nord-Larsen T, Bechsgaard A, Holm M, Holten-Andersen P (2003) Economic analysis of near-natural beech stand management in Northern Germany. For Ecol Manage 184:149–165. https://doi.org/10.1016/S0378-1127(03)00212-3
Nyland RD (2003) Even- to uneven-aged: the challenges of conversion. For Ecol Manage 172:291–300. https://doi.org/10.1016/S0378-1127(01)00797-6
Pausch R (2005) Ein System-Ansatz zur Darstellung des Zusammenhangs zwischen Waldstruktur. Arbeitsvolumen und Kosten in naturnahen Wäldern Bayerns, Forstliche Forschungsberichte München, Heft, p 199
Pretzsch H (1985) Die Fichten-Tannen-Buchen-Plenterwaldversuche in den ostbayerischen Forstämtern Freyung und Bodenmais. Forstarchiv 56(1):3–9
Pretzsch H (1998) Structural diversity as a result of silvicultural operations. LesnictvÍ-Forestry 44(10):429–439
Pretzsch H (2002) Application and evaluation of the growth simulator SILVA 2.2 for forest stands, forest estates and large regions. Forstwissenschaftliches Centralblatt 121:28–51
Pretzsch H (2009) Forest dynamics, growth, and yield. Springer, Berlin, Heidelberg
Pretzsch H (2019) Transitioning monocultures to complex forest stands in Central Europe: principles and practice. In: Stanturf JA (ed) Achieving sustainable management of boreal and temperate forests. Burleigh Dodds Science Publishing, Cambridge, pp 1–42
Pretzsch H, Biber P, Ďurský J (2002) The single tree-based stand simulator SILVA: construction, application and evaluation. For Ecol Manage 162:3–21. https://doi.org/10.1016/S0378-1127(02)00047-6
Pretzsch H, Biber P, Ïurský J, Sodtke R (2006) The Individual-Tree-Based Stand Simulator SILVA. In: Hasenauer H (ed) Sustainable Forest Management. Springer, Berlin Heidelberg, pp 78–84
Pretzsch H, Biber P, Schütze G, Uhl E, Rötzer T (2014) Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat Commun 5:4967
Pretzsch H, Biber P, Uhl E, Dauber E (2015) Long-term stand dynamics of managed spruce–fir–beech mountain forests in Central Europe: structure productivity and regeneration success. For Int J For Res 88:407–428
Pretzsch H, Biber P (2016) Tree species mixing can increase maximum stand density. Can J For Res. https://doi.org/10.1139/cjfr-2015-0413
Pretzsch H et al (2020) Evidence of elevation-specific growth changes of spruce, fir, and beech in European mixed-mountain forests during the last three centuries. Can J For Res. https://doi.org/10.1139/cjfr-2019-0368
Price C (2003) The economics of transformation from even-aged to uneven-aged forestry. In: Helles F, Strange N, Wichmann L (eds) Recent Accomplishments in Applied Forest Economics Research. Springer Science+Business Media, Dordrecht, pp 3–17
Price C, Price M (2008) Cost-benefit analysis of continuous cover forestry. In: Proceedings of the Biennial Meeting of the Scandinavian Society of Forest Economics Lom, Norway, 6th-9th April, 2008, pp 36–65
Price M, Price C (2006) Creaming the best, or creatively transforming? Might felling the biggest trees first be a win–win strategy? For Ecol Manage 224:297–303. https://doi.org/10.1016/j.foreco.2005.12.050
Pukkala T, Laiho O, Lähde E (2016) Continuous cover management reduces wind damage. For Ecol Manage 372:120–127. https://doi.org/10.1016/j.foreco.2016.04.014
R Core Team (2017) R: A language and environment for statistical computing. R foundation for Statistical Computing, Vienna, Austria
Ramage MH, Burridge H, Busse-Wicher M, Fereday G, Reynolds T, Shah DU, Wu G, Yu L, Fleming P, Densley-Tingley D, Allwood J, Dupree P, Linden PF, Scherman O (2017) The wood from the trees: the use of timber in construction. Renew Sustain Energy Rev 68:333–359
Reineke LH (1933) Perfecting a stand density index for even-aged forests. J Agric Res 46:627–638
Reventlow, DOJ (2020) Transitioning to near-natural forest management - effects on stand structure and wood production. Doctoral thesis. Department of Geosciences and Natural Resource Management, University of Copenhagen.
Richards FJ (1959) A flexible growth function for empirical use. J Exp Bot 10(2):290–301
Roessiger J, Griess VC, Härtl F, Clasen C, Knoke T (2013) How economic performance of a stand increases due to decreased failure risk associated with the admixing of species. Ecol Model 255:58–69. https://doi.org/10.1016/j.ecolmodel.2013.01.019
Roessiger J, Griess VC, Knoke T (2011) May risk aversion lead to near-natural forestry? A simulation study. Forestry 84:527–537
Ronnberg J, Vollbrecht G, Thomsen IM (1999) Incidence of butt rot in a tree species experiment in northern denmark. Scand J For Res 14:234–239. https://doi.org/10.1080/02827589950152755
Schmitt M (1994) Von faulen gesellen und spätberufenen: neue ergebnisse aus der waldwachstumsforschung zum thema „zielstärkennutzung und umsetzungsverhalten der fichte. Der Dauerwald 11:4–28
Schou E, Jacobsen JB, Kristensen KL (2012) An economic evaluation of strategies for transforming even-aged into near-natural forestry in a conifer-dominated forest in Denmark. Forest Policy Econ 20:89–98. https://doi.org/10.1016/j.forpol.2012.02.010
Schou E, Meilby H (2013) Transformation of even-aged European beech (Fagus sylvatica L.) to uneven-aged management under changing growth conditions caused by climate change. Eur J Forest Res 132:777–789. https://doi.org/10.1007/s10342-013-0702-5
Schröder J, Röhle H, Gerold D, Münder K (2007) Modeling individual-tree growth in stands under forest conversion in East Germany. Eur J Forest Res 126:459–472. https://doi.org/10.1007/s10342-006-0167-x
Schütz J-P (2001) Opportunities and strategies of transforming regular forests to irregular forests. For Ecol Manage 151:87–94. https://doi.org/10.1016/S0378-1127(00)00699-X
Schütz J-P, Götz M, Schmid W, Mandallaz D (2006) Vulnerability of spruce (Picea abies) and beech (Fagus sylvatica) forest stands to storms and consequences for silviculture. Eur J Forest Res 125:291–302. https://doi.org/10.1007/s10342-006-0111-0
Suadicani K, Fjeld D (2001) Single-tree and group selection in montane Norway spruce stands: factors influencing operational efficiency. Scand J For Res 16:79–87. https://doi.org/10.1080/028275801300004433a
Troen I, Petersen EL (1989) European wind atlas. Risø National Laboratory, Roskilde
Utschig H (1997) Reconversion of pure spruce stands into mixed forests; an ecological and economic valuation. In: Olsthoorn AFM, Bartelink HH, Gardiner JJ, Pretzsch H, Hekhuis HJ, Franc A, Wall S (eds) Management of mixed species forest: silviculture and economics. DLO Institute for Forestry and Nature research (IBN-DLO). IBM Scientific contributions 15, Wageningen, pp 319–330
Vanclay JK (2012) Modelling continuous cover forests. In: Pukkala T, von Gadow K (eds) Continuous cover forestry, 2nd edn. Springer, Netherlands, Dordrecht, pp 229–241
Vitasse Y et al (2019) Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob Change Biol 25:3781–3792. https://doi.org/10.1111/gcb.14803
Vítková L, Dhubháin A (2013) Transformation to continuous cover forestry: a review. Irish For 130:119–140
von Gadow K, Füldner K (1995) Zur Beschreibung forstlicher Eingriffe. Forstwissenschaftliches Centralblatt 114:151–159. https://doi.org/10.1007/bf02742221
von Lüpke B et al. (2004) Silvicultural strategies for conversion In: Spiecker H, Hansen J, Klimo E, Skovsgaard JP, Sterba H, Teuffel KV (eds) Norway spruce conversion—options and consequences. European Forest Institute Research Report 18. Brill, Leiden - Boston. XI, pp 121–164
von Teuffel K, Heinrich B, Baumgarten M (2004) Present distribution of secondary Norway spruce in Europe. In: Spiecker H, Hansen J, Klimo E, Skovsgaard JP, Sterba H, Teuffel KV (eds) Norway spruce conversion—options and consequences. European Forest Institute Research Report 18. Brill, Leiden - Boston. XI, pp 63–96
Wang T, Hamann A, Spittlehouse DL, Carroll C (2016) Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE. https://doi.org/10.1371/journal.pone.0156720
Yemshanov D et al (2015) A real options-net present value approach to assessing land use change: a case study of afforestation in Canada. Forest Policy Econ 50:327–336. https://doi.org/10.1016/j.forpol.2014.09.016
Yousefpour R, Hanewinkel M (2009) Modelling of forest conversion planning with an adaptive simulation-optimization approach and simultaneous consideration of the values of timber, carbon and biodiversity. Ecol Econ 68:1711–1722
Acknowledgements
The authors want to express their thanks to the Bavarian State Forest (Bayerische Staatsforsten AöR) for providing the data used for generating the simulation stands. We would also like to thank the Bavarian State Institute of Forestry (LWF) for providing the timber prices used in simulations. We would also like to express our gratitude towards Leonhard Steinacker and Martin Nickel for collecting the data used to validate the simulations. Finally, we wish to thank Thomas Knoke and Fabian Härtl for good discussion on the economic analysis, as well as the anonymous reviewers for many good suggestions for improvement of the manuscript.
Funding
The work has been funded by a framework contract between the Danish Ministry of Environment and the Department of Geosciences and Natural Resource Management, University of Copenhagen.
Author information
Authors and Affiliations
Contributions
D. O. J. Reventlow and H. Pretzsch came up with the original idea for the study. D. O. J. Reventlow adjusted and implemented the idea, carried out the simulations and wrote the manuscript. All authors contributed significantly to the study concept and improved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The author declares that they have no competing of interest.
Additional information
Communicated by Thomas Knoke.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Reventlow, D.O.J., Nord-Larsen, T., Biber, P. et al. Simulating conversion of even-aged Norway spruce into uneven-aged mixed forest: effects of different scenarios on production, economy and heterogeneity. Eur J Forest Res 140, 1005–1027 (2021). https://doi.org/10.1007/s10342-021-01381-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-021-01381-0