
Abstract
Almonds (Prunus dulcis Mill.) are generally relative drought-resistant crops. In the present study, morphological and pomological characteristics of 84 non-irrigated seedling-originated trees were evaluated to identify the genotypes best adapted to drought. Based on the characters recorded, significant differences were observed among the genotypes. Spring frost resistance was high in 17 out of 84 genotypes due late-blooming time. Nut weight ranged between 1.40 and 12.90 g with an average of 5.76, while kernels weighted between 0.10 and 3.10 g with an average of 1.35. Principal component analysis (PCA) revealed that 74.76% of total variances were explained by 10 components and confirmed the appropriateness of applying pomological traits to select the superior genotypes. Unweighted pair group method of arithmetic average (UPGMA) method and Euclidean distances-based analysis on all the recorded characters divided the genotypes into two major clusters. Based on ideal values of commercially important characteristics such as fruit yield, nut weight, shell hardness, kernel shape, kernel weight, and kernel taste, 11 genotypes were promising and are recommended for cultivation under drought condition.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Current climate change will lead to more frequent and severe drought events and mainly increase water scarcity (Collins et al. 2009). Moreover, as the world’s population increases, leading to the rapid expansion of urban, touristic and increases industrial activities, tensions and conflicts between water users and pressures on the environment will intensify. Because precipitation has been low in recent years, scarce water supplies necessitate careful management of agricultural crop irrigation in many regions of the world. Thus, the cultivation of drought-resistant fruit crops with a suitable performance at low water availability may decrease the requirements of irrigation. Some fruit tree species indeed own the ability to morphological acclimate to long-term drought, e.g., by variations (Egea et al. 2010).
Almonds (Prunus dulcis Mill.) are generally relative drought-resistant crops (Romero et al. 2004). They can withstand frequent periods of low soil moisture accompanied by high evaporative demand and high air temperature during the growing season (De Herralde et al. 2003). The tolerance of almond trees to water deficit stress is presumably related to adaptive mechanisms present in their leaves or roots. Such mechanisms are probably related to physiological mechanisms such as osmotic adjustment, long-term reduction stomatal conductance, and leaf shedding, which both decrease transpiration and increase in root density and rooting depth. Thus, almonds may be suitable for deficit irrigation programs, maintaining its commercial value by retaining a proper kernel size (Egea et al. 2010).
Water availability is an essential factor affecting plant growth and yield, mainly in arid and semi-arid regions, where plants are often subjected to periods of drought. The occurrence of morphological and physiological responses, which lead to some adaptation to drought stress, may vary considerably among species. In general, strategies of drought-avoidance or drought tolerance can be recognized, both involving diverse plant mechanisms that provide the plants the ability to respond and survive drought (Souza et al. 2004; Rouhi et al. 2007).
Drought stress significantly affects photosynthesis, and thus reduces plant productivity and growth, also depending on rapidity, severity, and duration of drought events (Rouhi et al. 2007). Some plant species can control stomatal morphology and density to acclimate to changes in the environment (Camposeo et al. 2011). Also, the stomatal limitation of photosynthesis during drought stress has been well documented. Under drought, prolonged stomatal closure may decrease CO2 uptake, thus reducing leaf internal CO2 concentrations and imposing an imbalance between energy provision by the photosynthetic light absorption systems and energy requirement by photosynthetic CO2 assimilation (Souza et al. 2004).
The selection of genotypes more resistant to drought is essential to optimize production in dry environments. Native germplasms of almond are a valuable genetic source for important physiological characteristics such as drought tolerance that can be identified and used for breeding programs. Identification of drought-tolerant genotypes is essential in arid and semi-arid regions, and it is one of the essential breeding purposes in applied researches for most of the crops. Therefore, the objective of the current study was to evaluate morphological and pomological characteristics of some non-irrigated seedling-originated almond trees to identify the genotypes best adapted to drought.
Material and Methods
Plant Material
Morphological and pomological characteristics of 84 non-irrigated seedling-originated trees were evaluated to identify the genotypes best adapted to drought from Sabzevar and Ghorveh regions in Iran. The Sabzevar region is located in 36º12′32″N, 57º43′10″E, and 977 m above sea level with 17.10 ºC mean annual temperature and 190 mm annual rainfall. Also, the Ghorveh region is located in 35º15′22″N, 47º34′13″E, and 1900 m above sea level with 10.60 ºC mean annual temperature and 418 mm annual rainfall. The selected mature seedling origin trees were named and labeled based on their region area.
Morphological and Pomological Evaluations
The survey was made using 34 morphological and pomological characters according to almond descriptors developed by the International Plant Genetic Resources Institute (IPGRI) (Gulcan 1985). Measurements of nut and kernel traits were based on 50 replicates, and the mean values were used. Dimensions of leaf, nut, and kernel were measured with a digital caliper. Nut and kernel weight was measured with an electronic balance with 0.01 g precision. The traits, including tree growth habit, tree growth vigor, tree height, trunk color intensity, trunk diameter, canopy density, branching, branch density, branch flexibility, leaf density, leaf serration shape, leaf upper surface color, leaf lower surface color, location of flower bud, fruit yield, nut apex shape, nut shape, shell hardness, shell color intensity, marking of the outer shell, kernel shriveling, kernel pubescence, and kernel taste were qualitatively estimated based on rating and coding described by the almond descriptor (IPGRI) (Gulcan 1985).
Data Analysis
Significant differences among the genotypes based on the measured characters were determined using one-way analysis of variance (ANOVA) with SAS software (SAS Institute, Cary, NC, USA, 1990). Coefficients of variation (CV%; SD/mean × 100) were calculated as a variation index. Pearson correlation coefficients were used to determine correlations between the traits using SPSS® software version 16 (SPSS Inc., Chicago, IL, USA, Norusis 1998). Principal component analysis (PCA) was applied to investigate the relationships among the genotypes using SPSS® software. To better understand the patterns of variations among the genotypes, the distance matrix of morphological data as input data was used for cluster analysis using the unweighted pair group method of arithmetic average (UPGMA) method with PAST statistics software (Hammer et al. 2001). Also, a scatter plot was created according to the first and second principal components (PC1/PC2), using PAST statistics software.
Results and Discussion
Morphological and Pomological Characterizations
In total, 29 out of 34 characters measured showed CVs more than 20.00%, indicating high variability among the genotypes. The highest CV was observed for branch flexibility (76.10%) and followed by fruit yield (69.10%) and canopy density (65.27%). In contrast, nut diameter showed the least CV (16.11%) and then nut width (18.40%), trunk color intensity (18.68%), nut length (18.90%), and kernel length (19.36%) (Table 1).
Spring frost resistance was low (in 35 genotypes), intermediate (32), and high (17) due to blooming time of genotypes. Late-blooming is an essential factor to protect damages caused by spring frosts in continental climates (Khadivi-Khub and Etemadi-Khah 2015). Therefore, finding late-blooming trees is one of the main goals of almond breeding program.
The genotypes showed four types of growth habit, including drooping (2), spreading (10), spreading to upright (32), and upright (40). Upright genotypes are the main characteristics required for high and super high-density systems, and the current tendency towards intensification in many species such as olive may affect almond shortly, especially given that the first results have been so promising (Miarnau et al. 2013; Rius et al. 2013). Intermediate status was predominant for six vegetative variables, including growth vigor (65 genotypes), tree height (58), trunk diameter (69), branching (57), branch density (54), and leaf density (64) (Table 2). Leaf length ranged from 16.33 to 111.50 mm, while leaf width varied from 7.00 to 40.26 mm. Also, the range of petiole length was 6.70–32.05 mm (Table 1). The small leaves likely indicate adaptation to the xerophytic conditions (rainfall less than 150 mm per year) (Lansari et al. 1994).
The location of flower buds was mostly on the spur (57 genotypes). Fruit yield was very low in 37, low in 11, intermediate in 26, and high in 10 genotypes. The genotypes formed four groups based on nut shape, including round (17), ovate (31), cordate (14), and oblong (22). Nut length ranged from 21.73 to 51.50 mm, and nut width varied from 9.97 to 31.52 mm, while the range of nut diameter was 9.95–21.15 mm. Nut weight ranged between 1.40 and 12.90 g, with an average of 5.76 (Table 1). Shell hardness was hard and then semi-hard in most of the genotypes (34 and 29 genotypes, respectively) (Table 2).
High kernel shriveling was predominant (45 genotypes). Kernel taste was sweet in the majority of genotypes (63) (Table 2). Kernel length ranged from 11.75 to 31.52 mm, and kernel width varied from 7.08 to 19.53 mm, while the range of kernel thickness was 1.00–13.33 mm. Kernel weight ranged between 0.10 and 3.10 g, with an average of 1.35 (Table 1). In many European and American cultivars, kernel weight average with approximately 1.00 g is common, and such weight is a desired trait in breeding programs (Gradziel and Kester 1998). In the current investigation, the value of kernel weight in 53 out of 84 studied non-irrigated genotypes was more than 1.00 g. The values of the most important fruit-related traits for the superior genotypes are presented in Table 3.
Correlations Between Characters
Simple correlation coefficient analysis revealed significant correlations between the variables measured (Table 4). Tree growth habit was negatively and significantly correlated with trunk diameter (r = −0.23). Tree growth vigor showed significant and positive correlations with tree height (r = 0.38), canopy density (r = 0.47), branching (r = 0.65), branch density (r = 0.41), leaf density (r = 0.26), and leaf length (r = 0.29). Leaf length was positively and significantly correlated with leaf width (r = 0.79) and petiole length (r = 0.71) and corresponded with the previous results in almond (Talhouk et al. 2000; Khadivi-Khub and Etemadi-Khah 2015).
Fruit yield showed significant and positive correlations with canopy density (r = 0.25), leaf density (r = 0.22), and leaf lower surface color (r = 0.34). Nut length was positively and significantly correlated with tree growth vigor (r = 0.40), tree height (r = 0.27), leaf length (r = 0.38), leaf width (r = 0.21), petiole length (r = 0.29), nut width (r = 0.75), and nut diameter (r = 0.47) and agreed with the previous results in almond (Khadivi-Khub and Etemadi-Khah 2015; Sepahvand et al. 2015; Khadivi-Khub and Osati 2015). In addition, leaf width showed similar positive correlation with above variables.
Nut weight showed significant and positive correlations with tree growth vigor (r = 0.32), leaf length (r = 0.38), leaf width (r = 0.27), petiole length (r = 0.22), nut length (r = 0.82), nut width (r = 0.87), and nut diameter (r = 0.70), while it was negatively and significantly correlated with fruit yield (r = −0.22). Kernel weight showed significant and positive correlations with leaf length (r = 0.30), leaf width (r = 0.29), nut length (r = 0.65), nut width (r = 0.69), nut diameter (r = 0.69), nut weight (r = 0.85), kernel length (r = 0.72), kernel width (r = 0.70), and kernel thickness (r = 0.73), while it was negatively and significantly correlated with fruit yield (r = −0.26). Kernel taste showed negative and significant correlation with marking of outer shell (r = −0.28) and agreed with the previous results in almond (Khadivi-Khub and Etemadi-Khah 2015; Sepahvand et al. 2015; Khadivi-Khub and Osati 2015).
PCA
The PCA revealed that 74.76% of total variances were explained by 10 components (Table 5). The PC1 was positively and significantly correlated with nut length, nut width, nut diameter, nut weight, kernel length, kernel width, kernel thickness, and kernel weight and explained 17.83% of total variance. This situation confirms the appropriateness of applying the pomological traits to select the superior genotypes. The characteristics related to fruit size are reported to be important for differentiating the almond genotypes (Talhouk et al. 2000; Khadivi-Khub and Etemadi-Khah 2015). Five traits, including tree height, canopy density, leaf density, leaf lower surface color, and fruit yield showed the most significant weight on PC2 that accounted for 10.39%. The PC3, with explaining 7.81% of total variance, showed positive correlations with leaf length, leaf width, and petiole length. Relationships established among the genotypes using PCA may inform genetic linkages between loci controlling the observed traits or indicate pleiotropic effects (Iezzoni and Pritts 1991).
Cluster Analysis and Scatter Plot
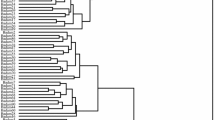
The dendrogram created using the UPGMA method and Euclidean distances based on all the recorded data placed the genotypes studied into two major clusters (Fig. 1). The first major cluster contained 54 genotypes that formed two sub-clusters. The first sub-cluster consisted of 14 genotypes, while the second sub-cluster included 40 genotypes. Also, the second major cluster contained 30 genotypes.

UPGMA cluster analysis of the studied non-irrigated almond genotypes based on morphological traits using Euclidean distance coefficients
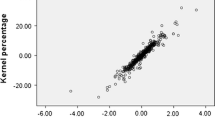
Also, the scatter plot generated using PC1 and PC2 determined similarities and dissimilarities among genotypes and showed that genotypes were distributed into four sides of the plot (Fig. 2). From negative to positive values of PC1, the genotypes showed gradual increases in nut length, nut width, nut diameter, nut weight, kernel length, kernel width, kernel thickness, and kernel weight. Starting from negative towards positive values of PC2, the genotypes showed gradual increases in tree height, canopy density, leaf density, leaf lower surface color, and fruit yield. The majority of genotypes were distributed in central parts of the plot, while Sabzevar‑55 and Sabzevar-51 genotype were arranged outside the inner ellipse characterized by the lowest values for nut length, nut width, nut weight, kernel width, and kernel length.

Biplot for the studied non-irrigated almond genotypes based on PC1/PC2. The symbols represent the genotypes of each site, including Ghorveh (G) and Sabzevar (S)
The obtained data revealed high morphological diversity among the studied genotypes. Some of the non-irrigated genotypes produced high yield and also kernels with high quality. Now, it is of paramount importance to protect water resources and their integrity for future use (Katerji et al. 2008). In this sense, to overcome the problems associated to a boost in water prices, as the discouragement of farmers and ultimately land abandonment, Garcia-Tejero et al. (2014) indicated that an alternative could be to provide correct incentives for farmers to adopt changes in their irrigation methods by implementing strategies and tools for sustainable water saving. Among the strategies that can be applied to attain water saving are the use of improved, innovative, and precise deficit irrigation (DI) management practices able to minimize the impact on crop yield and quality (Fernandez and Torrecillas 2012). Also, to contribute to water saving, fruit culture should be directed towards the use of plant materials that are less water-demanding or able to withstand deficit irrigation with minimum impact on yield and fruit quality. The development of drought-resistant almond production systems using native germplasm would make more sustainable production possible, particularly in marginal areas with harsh climate conditions (Gouta et al. 2019).
Conclusions
Almond orchards have been established traditionally in most parts of Iran with seedlings. Therefore, there is a valuable genetic diversity in these orchards that can be used in breeding programs for some characters such as resistance to drought. To improve not only biodiversity but also to save water and hence protect the integrity of water resources for the future, the diversification of production and consumption habits is necessary, including the use of a broader range of plant species, in particular those currently identified as underutilized and needing a low input of synthetic fertilizers, pesticides, and water. This option must be compatible with the consolidation of the cultivation of some crops, such as olive, almond, or grapevine, which are low water demanding and profitable crops. In this sense, in some countries, during recent decades, there has been a specific interest in diversifying fruit tree production by cultivating species with under-exploited potential. Among these emerging crops, most of them are characterized by their attractive fruits and health-related qualities, so that they may attract consumer attention and contribute to producer profitability.
Results of the current study showed that some of the genotypes can produce high kernel quantity and quality under non-irrigated conditions. Other suitable characteristics for an almond tree in rainfed conditions are compact growth habit, leaves with thick cuticle layer, and osmotic adjustment ability in leaves. Based on ideal values of the important and commercial characters of almond such as fruit yield, nut weight, shell hardness, kernel shape, kernel weight, and kernel taste, 11 genotypes including Sabzevar-46, Sabzevar-27, Sabzevar‑5, Sabzevar-13, Ghorveh‑7, Sabzevar-17, Sabzevar-29, Ghorveh‑5, Ghorveh‑6, Sabzevar-12, and Sabzevar-62 were promising and are recommended for cultivation in orchards.
References
Camposeo S, Palasciano M, Vivaldi GA, Godini A (2011) Effect of increasing climatic water deficit on some leaf and stomatal parameters of wild and cultivated almonds under Mediterranean conditions. Sci Hortic 127:234–241
Collins R, Kristensen P, Thyssen N (2009) Water resources across europe-confronting water scarcity and drought. European Environment Agency, Copenhague, p 57
De Herralde F, Biel C, Save R (2003) Leaf photosynthesis of eight almond tree cultivars. Biol Plantarum 46:557–561
Egea G, Nortes PA, Gonzalez-Real MM, Baille A, Domingo R (2010) Agronomic response and water productivity of almond trees under contrasted deficit irrigation regimes. Agri Water Manag 97: 171–181
Fernandez JE, Torrecillas A (2012) For a better use and distribution of water. An introduction. Agric Water Manag 114:1–3
Garcia-Tejero IF, Duran-Zuazo VH, Muriel-Fernandez JL (2014) Towards sustainable irrigated Mediterranean agriculture: implications for water conservation in semi-arid environments. Water Int 39:635–648
Gouta H, Ksia E, Ayachi MM, Martinez-Gomez P (2019) Agronomical evaluation of local Tunisian almond cultivars and their breeding prospects. Eur J Hortic Sci 84(2):73–84
Gradziel TM, Kester DE (1998) Breeding for self-fertility in California almond cultivars. Acta Hortic 470:109–117
Gulcan R (ed) (1985) Descriptor list for almond (Prunus amygdalus). International Board for Plant Genetic Resources, Rome
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1):9 (http://palaeoelectronica.org/2001_1/past/issue1_01.htm)
Iezzoni AF, Pritts MP (1991) Applications of principal components analysis to horticultural research. Hortic Sci 26:334–338
Katerji N, Mastrorilli M, Rana G (2008) Water use efficiency of crops cultivated in the Mediterranean region. Review and analysis. Eur J Agron 28:493–507
Khadivi-Khub A, Etemadi-Khah A (2015) Phenotypic diversity and relationships between morphological traits in selected almond (Prunus amygdalus) germplasm. Agrofor Syst 89:205–216
Khadivi-Khub A, Osati E (2015) Identification of superior almond (Prunus dulcis) genotypes from a germplasm field in Iran. Europ J Hort Sci 80(3):139–144
Lansari A, Iezzoni AF, Kester DE (1994) Morphological variations within collections of Moroccan almond clones and Mediterranean and North American cultivars. Euphytica 78:27–41
Miarnau X, Montserrat R, Battle I, Vargas FJ (2013) High density planting in almond orchards. VI Int. Symp. on Almonds and Pistachios, Murcia, 05.2013
Norusis MJ (1998) SPSS/PC advanced statistics. SPSS Inc, Chicago
Rius X, Bordas M, Torrents J (2013) Super high density almond orchards. VI Int. Symp. on Almonds and Pistachios, Murcia, 27–31 May
Romero P, Botia P, Garcia F (2004) Effects of regulated deficit irrigation under subsurface drip irrigation conditions on vegetative development and yield of mature almond trees. Plant Soil 260:169–181
Rouhi V, Samson R, Lemeur R, Van Damme P (2007) Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery. Environ Exp Bot 59:117–129
SAS® Procedures (1990) Version 6, 3rd edn. SAS Institute, Cary
Sepahvand E, Khadivi-Khub A, Momenpour A, Fallahi E (2015) Evaluation of an almond collection using morphological variables to choose superior trees. Fruits 70:53–59
Souza RP, Machado EC, Silva JAB, Lagoa AMMA, Silveira JAG (2004) Photosynthetic gas exchanges in cowpea (Vigna unguiculata) during water stress and recovery. Environ Exp Bot 51:45–56
Talhouk SN, Lubani RT, Baalbaki R, Zurayk R, Al Khatib A, Parmaksizian L, Jaradat AA (2000) Phenotypic diversity and morphological characterization of Amygdalus species in Lebanon. Genet Resour Crop Evol 47:93–104
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
P. Heidari, S. Sanaeizadeh, M. Rezaei, and A. Khadivi declare that they have no competing interests.
Rights and permissions
About this article

Cite this article
Heidari, P., Sanaeizadeh, S., Rezaei, M. et al. Phenotypical and Pomological Characterization of Non-irrigated Almond (Prunus dulcis Mill.) Trees to Select Superior Genotypes. Erwerbs-Obstbau 64, 333–343 (2022). https://doi.org/10.1007/s10341-022-00653-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10341-022-00653-2