
Abstract
Predictions derived from species distribution models (SDMs) are strongly influenced by the spatial scale at which species and environmental data (e.g. climate) are gathered. SDMs of mountain birds usually build on large-scale temperature estimates. However, the topographic complexity of mountain areas could create microclimatic refuges which may alter species distributions at small spatial scales. To assess whether fine-scale data (temperature and/or topography) improve model performance when predicting species occurrence, we collected data on presence–absence of bird species, habitat and fine-scale temperature at survey points along an elevational gradient in the Alps (NW Italy). Large-scale temperature data, and both large- and fine-scale topography data, were extracted from online databases for each point. We compared species models (fine-scale vs large-scale) using an information-theoretic approach. Models including fine-scale temperature estimates performed better than corresponding large-scale models for all open habitat species, whereas most forest/ecotone species showed no difference between the two scales. Grassland birds such as Northern Wheatear Oenanthe oenanthe and Water Pipit Anthus spinoletta were positively associated with warmer microclimates. These results suggest that alpine grassland species are potentially more resistant to the impact of climate change than previously predicted, but that indirect effects of climate change such as habitat shifts (forest- and shrub encroachment at high elevations) pose a major threat. Therefore, active management of alpine grassland is needed to maintain open areas and to prevent potential habitat loss and fragmentation. SDMs based solely on large-scale temperatures for open habitat species in the Alps should be re-assessed.
Zusammenfassung
Mikroklima beeinflusst die Verbreitung von Offenlandarten jedoch nicht von Waldarten im Alpenraum
Vorhersagen, die auf Artverbreitungsmodellen basieren, können stark davon beeinflusst werden, auf welcher räumlichen Skala Art- und Umweltdaten (z.B. Klimadaten) erhoben werden. Die Artverbreitungsmodelle von Gebirgsvögeln stützen sich häufig auf großskalige Temperaturdaten. Allerdings könnten mikroklimatische Refugien, hervorgerufen durch die topografische Komplexität von Gebirgsregionen, die Verbreitung von Arten kleinräumig verändern. Um beurteilen zu können, ob feinskalige Daten (Temperatur und/oder Topografie) die Modellleistung bei der Vorhersage von Artvorkommen verbessern können, haben wir Präsenz/Absenz-Daten von Vogelarten, Habitatdaten und feinskalige Temperaturdaten an Erfassungspunkten entlang eines altitudinalen Gradienten in den Alpen (NW Italien) gesammelt. Für jeden dieser Punkte wurden zudem großskalige Temperaturdaten sowie klein- und großskalige topografische Daten aus Online-Datenbanken extrahiert. Basierend auf den unterschiedlichen Datenskalen (fein- vs. großskalig) wurden die jeweiligen Artmodelle mithilfe eines informationstheoretischen Ansatzes verglichen. Die Modelle aller Offenlandarten, welche feinskalige Temperaturdaten enthielten, waren besser als ihre entsprechenden großskaligen Varianten. Für Wald- und Ökotonarten zeigten sich keine Unterschiede zwischen Modellen unterschiedlicher Datenskalen. Offenlandarten, wie der Steinschmätzer und der Bergpieper, zeigten eine positive Beziehung zu wärmeren mikroklimatischen Bedingungen. Dies könnte darauf hinweisen, dass alpine Offenlandarten gegenüber den direkten Auswirkungen des Klimawandels widerstandsfähiger sind als bisher angenommen. Dennoch stellen indirekte Effekte des Klimawandels (Verbuschung, Verschiebung der Waldgrenze in größere Höhenlagen) noch immer eine der Hauptgefährdungsursachen für jene Arten dar. Um dem entgegen zu wirken, ist es notwendig, dass Almwiesen durch aktive Bewirtschaftung offen gehalten werden, sodass durch Verbuschung verursachter potenzieller Habitatverlust und Habitatfragmentation verhindert werden können. Zudem sollten Artverbreitungsmodelle für Offenlandarten der Alpen, welche nur auf großskaligen Temperaturdaten beruhen, neu bewertet werden.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Species distribution models (henceforth SDMs) are a widely used tool in conservation (Guisan and Thuiller 2005; Rodríguez et al. 2007; Franklin 2013; Engler et al. 2017) for a range of taxa (Ongaro et al. 2018; Lewthwaite et al. 2018; Hof and Allen 2019). In the face of climate change, SDMs have become particularly important in predicting current and/or future distributions of species under different climate change scenarios (Avalos and Hernández 2015; Jackson et al. 2015; Lehikoinen and Virkkala 2016). These studies usually rely on macroclimate data, which describe climatic conditions at a relatively large scale (approximately one square kilometre or more; Zellweger et al. 2019) derived from national networks, weather stations or online databases (e.g. Worldclim; Hijmans et al. 2005).
However, mountain environments are often poorly represented by conventional climate station data, and uncertainty for interpolated climatic values is high (Hijmans et al. 2005). Furthermore, local temperature can vary substantially due to the topographic complexity in mountain areas (Scherrer and Körner 2010; Gunton et al. 2015), thus creating a mosaic of microclimatic conditions over small spatial scales. Depending on discipline, microclimates have been defined in various ways. In this study, we adopt the definition by Bramer et al. (2018) who defined microclimate as fine-scale climate variations at spatial resolutions of < 100 m, which are influenced by fine-resolution biotic and abiotic variations (topography, soil type and vegetation). Topographic variables like aspect and slope can markedly alter microclimate by influencing the amount of incoming solar radiation between different exposed slopes. Between north- and south-exposed slopes, temperature can differ by approximately 1 °C if slopes are gentle (< 5° gradient) but can increase up to 5 °C if slopes are steep (40° gradient; Gubler et al. 2011). Moreover, these differences could subsequently influence snow accumulation processes and thus the rate of snow melt in spring (Gubler et al. 2011).
There is mounting evidence of the importance of microclimate in influencing habitat selection. For example, Bramblings Fringilla montifringilla tend to rest in higher densities in areas with warm microclimatic conditions (Zabala et al. 2012). In Mountain Chickadees Poecile gambeli, microclimates influence the selection of foraging sites (Wachob 1996). Microclimates can also act as thermal refuges, which enable individuals to persist despite unfavourable ambient conditions (Wilson et al. 2015). This has been shown in Northern Bobwhites Colinus virginianus, which mitigated thermal stress by seeking thermally buffered microclimatic sites during hot days (Carroll et al. 2015). Furthermore, Northern Bobwhite nest site selection was proven to be influenced by microclimate: Individuals nested in cooler and moister microclimatic conditions compared to surrounding non-nesting locations (Tomecek et al. 2017; Carroll et al. 2018).
Only a few studies have investigated the role of microclimate within a mountain context. Frey et al. (2016) showed that fine-scale temperature metrics were strong predictors of bird distributions, with temperature effects being larger than vegetation effects on occupancy dynamics in mountain forests (but see Viterbi et al. 2013). In the Alps, the habitat of the alpine Rock Ptarmigan Lagopus muta helvetica is characterised by a wide variety of microclimates over small spatial scales with individuals choosing colder sites in summer (Visinoni et al. 2015).
Beside the direct impact on birds, microclimate also plays a crucial role in habitat selection in insects. It has been demonstrated that in Parnassius apollo, a mountain specialist butterfly, larval habitat selection is related to ambient temperature. Larvae selected warm microclimates when ambient temperatures fell below a threshold of 27 °C, whereas cold microclimates were selected when this threshold was exceeded (Ashton et al. 2009). Microclimate can further influence oviposition (Stuhldreher et al. 2012), and the precise microclimatic conditions for thermoregulation are actively sought by montane species of the genus Erebia (Kleckova et al. 2014). In this respect, microclimate would not only shape the distributions of these butterfly species, but it will also indirectly influence bird species which rely on caterpillars as a food source for chick rearing.
Microclimate thus has the potential to influence many aspects of an organism’s life cycle. It could help to buffer or to compound the effects of climate change (Spasojevic et al. 2013). To assess the impact of climate change on current or future distributions of species it is crucial to gather climate data at the most appropriate scale in order to increase model accuracy (Barton et al. 2018; Randin et al. 2009). However, predictions for future geographic distributions of mountain birds under a range of climate change scenarios have thus far been based on models which have considered climate variables measured at large scales, usually ca. 1 km2 (Chamberlain et al. 2013, 2016; Brambilla et al. 2016, 2017a). Given the potential for bird responses to microclimatic conditions in mountains (Frey et al. 2016; Visinoni et al. 2015), it may be more appropriate to consider the role of climate measured at finer spatial resolutions in determining mountain bird distributions. This is particularly important given that environmental conditions in mountains typically change over very small spatial scales thanks to steep elevation gradients (Scherrer and Körner 2010; Gunton et al. 2015).
In this study, we investigated the role of microclimate for a range of Alpine ecotone and open habitat species. There were two specific aims. First, to evaluate if models including a microclimatic variable (in this case temperature) show better performance than models using large-scale climate estimates. This will inform future modelling studies, and should help to improve predictions of future impacts of climate change on Alpine birds where microclimatic effects are evident. Second, to assess if models including topographic variables (slope and aspect) in combination with climatic variables (fine and large scale) increase model performance. This will assess the extent to which topographic variables should be included in SDMs of alpine bird species. Based on previous studies, which showed that microclimate can influence bird distributions within mountain habitats (Frey et al. 2016; Visinoni et al. 2015), we hypothesise that models using fine-scale temperature estimates will show better model performance than models using large-scale temperature estimates.
Methods
Study area and point selection
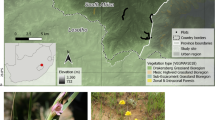
The study was carried out in Val Troncea Natural Park (44° 57′ 28″ N; 6° 56′ 28″ E) in the western Italian Alps. At lower elevations, the area is dominated by larch Larix decidua. The natural treeline is typically found at around 2200 m asl, but varies depending on local conditions. Typical shrub species are Juniperus nana (henceforth Juniper) and Rhododendron ferrugineum (henceforth Rhododendron) which rapidly encroached wide areas of grasslands after the decline of agro-pastoral activities. Grasslands are mainly dominated by Festuca curvula, Carex sempervirens, and Trifolium alpinum. Scree and rocky areas occur predominantly at higher elevations, above approximately 2700 m asl.
Point counts were carried out along an elevational gradient ranging from 1750 to 2820 m encompassing forest, ecotone and open habitats. Point count locations coincided with the centroids of a pre-existing grid at a scale of approximately 100 × 100 m (there was some variation, due to access constraints for example; Probo et al. 2014) along the south-western facing slope of the valley. All points were spaced a minimum of 200 m apart.
Bird surveys
Point counts (n = 221) were carried out from mid-May to mid-July 2017 following the methods of Bibby et al. (2000), using a 10-min count period. At each point count location, all individual birds seen or heard were recorded within a 100-m radius (estimated with the aid of a laser range finder). Point counts commenced 1- to 1.5 h after sunrise and continued until 1200 h. Surveys did not take place in excessively wet or windy conditions. Each point count location was visited once. Point counts were performed by a team of four field workers. Fieldwork was proceeded by at least one day of training for each of the field workers in order to minimise potential observer bias.
Habitat data collection
At each point count location, habitat data were collected through the visual estimation of the percentage cover of canopy (i.e. vegetation above head height), the dominant shrub species, open grassland and bare rock (including scree and unvegetated areas) within a 100-m radius of the point’s centre. The dominant shrub species were defined into four groups: Rhododendron, Juniper, bilberry (Vaccinium myrtillus and V. gaultherioides) and other (e.g. Green Alder Alnus viridis, Willow Salix spp, and also including young trees less than two meters in height, mostly European Larch Larix decidua). Furthermore, the number of mature trees (greater than c. 20 cm in diameter at breast height) within a 50-m radius of a point count location was counted. These estimates have been shown to correlate well with estimates of land cover derived from remote sensing and have been used as the basis of predictive models for several species considered here (Chamberlain et al. 2013, 2016; Jähnig et al. 2018).
Temperature measurements
At each point count location, temperature was measured with hygro buttons (Plug & Track™), using methods based on Frey et al. (2016). Each button was stuck on the bottom of a small plastic cup, which was attached upside down to a bamboo stick to protect the button against wind, direct sunlight and water. Mean button height was 40.89 cm (min = 28 cm, max = 47 cm). Hygro buttons were programmed to record temperature every 5 min. They were placed 24 h before a point count commenced and were collected 24 h after the point count ended, which resulted in a total recording time of 48 h. At every hygro button location, button height, distance to slope, substrate and canopy presence/absence were recorded.
Statistical analysis
Temperature modelling
For each point count location, minimum, maximum and mean temperatures were derived over the 48-h recording period. All temperature measurements were checked for collinearity by calculating Pearson’s correlation coefficient. Mean temperature was strongly correlated with both minimum (r = 0.80) and maximum temperature (r = 0.73) over the recording period. Therefore, temperature modelling was undertaken with mean temperature values. The same procedure was repeated for night-time temperatures. Minimum, maximum and mean night-time temperatures were obtained for the time period between 23:00 p.m. and 03:00 a.m. over the same recording period at each point. There was a strong positive correlation between mean night-time temperature and both minimum (r = 0.97) and maximum night-time temperature (r = 0.89).
The objective of the first analysis was to model temperature in relation to date and elevation. This model was then used to predict a standardised temperature at each point count location, set at a fixed date, which was representative of the fine-scale temperature at that point controlling for seasonal effects. This procedure provided data which were analogous to the larger scale temperature data (see below). This standardised temperature was then used subsequently as a variable in species distribution models. Note that all subsequent modelling steps were performed separately for mean temperature and mean night-time temperature. However, models with night-time temperature were very similar to those using mean temperature, so we focus on the latter. Further details on night-time temperature models are given in the Electronic Supplementary Material (ESM) Table S1.
First, to investigate if temperature recording was influenced by characteristics of the hygro button’s position, it was analysed using a generalised linear model in relation to button height, distance to slope, substrate underneath the button and canopy presence/absence, specifying a normal error distribution. None of the variables showed a significant effect on mean temperature (p > 0.05); therefore, they were not considered further in the analysis.
In the next modelling step, standardised temperature estimates were derived separately for open Alpine grassland and forest/ecotone habitat, i.e. models were used to estimate temperature for a given elevation whilst accounting for seasonal variation. Points were classified as Alpine grassland if there was no canopy within 100 m radius of the point count centre (following Chamberlain et al. 2013). For open habitat points (n = 93), temperature was modelled in relation to date and elevation. Date were described as the number of days passed since the start of the field season, where day 1 = 27-May-2017. Canopy cover was added to the model structure for points located in forest and ecotone habitat (n = 128). In both cases, a normal distribution was specified. Prior to modelling, all variables were scaled and centred using the scale function in R version 3.5.2 (R Development Core Team 2018). Collinearity was assessed using Variance inflation factors (VIFs), calculated using the ‘corvif’ function (package ‘AED’, Zuur et al. 2009), and by considering Spearman correlations between continuous variables. All variables had VIF < 3, and no pair of variables showed a correlation > 0.7, indicating low levels of inter-correlation. These models were used to derive a standardised temperature for each point, based on the elevation at that point, the canopy cover (for forest/ecotone habitat) and for a date fixed at 15th June.
Species distribution models
Birds detected within a 100-m radius of a point count location were used to analyse species distribution (presence/absence of individual species). Bird species were considered in the modelling process if they were present on at least 15% of the points; below this threshold model performance is consistently poor (Chamberlain et al. 2013) (Table 1).
The commonest species were modelled in relation to four different variable sets: (1) habitat (HABITAT), (2) habitat + temperature (TEMP), (3) habitat + topography (TOPO), (4) habitat + temperature + topography (COMB; Table 2). Temperature and topographic variables were used at two different scales (large-scale/ fine-scale). Fine-scale temperature estimates were derived from the temperature modelling approach described above, whereas large-scale temperature data for each point were extracted from the Worldclim database (Hijmans et al. 2005) by calculating the average temperature within a 1000-m radius of the point count centre. Topographic variables (aspect and slope) were derived from a Digital Elevation Model (DEM) at a spatial resolution of 10 m. Aspect was transformed as x = − 1 × cos[Ø(π/180)], where Ø is measured in degrees. Values ranged from 1 where solar insolation was higher (south-facing slopes) to − 1 (north-facing slopes) where it was lower.
The mean aspect (transformed values) and slope was calculated within a 100-m (fine-scale) and a 1000-m (large-scale) radius of the point count centre for the analysis. Habitat variables were kept at a constant scale in the models (as the objective was to test scale effects in temperature and topography). Habitat models of Lesser Whitethroat Sylvia curruca and Dunnock Prunella modularis were tested for non-linear relationships with Rhododendron and Juniper cover as suggested by previous work (Jähnig et al. 2018). Habitat models with and without quadratic terms for shrub species cover were compared using AIC. Lesser Whitethroat models showed lower AIC values for the habitat model without quadratic terms. Therefore, these were omitted in further modelling steps. The addition of the quadratic term for Rhododendron cover reduced the AIC of the habitat model for Dunnock by ΔAIC > 2; hence it was included in the next modelling steps.
The occurrence probability of each species was modelled in relation to the different variable sets using a binomial logistic regression, after controlling for potential collinearity (as above). In the case of open habitat species, we found high VIFs for the variables rock and grass cover. After the removal of rock cover, all VIFs were below the threshold of three. As a result, rock cover was removed from all models for open habitat species. The correlation between rock and grass cover occurred because of the landscape characteristics above the treeline, where alpine grassland is often interspersed with rocks. As those two habitat characteristics are not separable, we used grass cover as proxy to describe this kind of habitat.
Data were analysed using an information theoretic approach with the MuMIn package in R (Bartón 2013). This entailed deriving full models for each variable set at each scale (except habitat which was kept constant in all models) using generalised linear models (R package lme4; Bates et al. 2015). This approach served two goals. First, model-averaged parameter estimates were derived for all combinations of variables in each full model set in order to identify variables that were most closely associated with bird distribution. p values derived from the model-averaged parameter estimates and their SEs were considered to represent significant effects when p < 0.05. Second, the Akaike information criterion corrected for small sample size (AICc) was determined for each individual model and was used to assess model performance for different variable combinations at different scales in the full model. In this way it was possible to assess which combination of the four different variable sets produced the best models, and at which scale.
At each scale, the residuals for all full models were extracted and tested for spatial autocorrelation using Moran’s I (Moran 1950). Significant spatial autocorrelation was found for models of Eurasian Skylark Alauda arvensis, Tree Pipit Anthus trivialis and Water Pipit. For these species, spatial effects were incorporated by modelling their distributions using Generalized Additive Models (GAMs) from the mgcv package (Wood 2011) by fitting smoothed terms for latitude and longitude in the model, following Wood (2017). However, for Skylark, these models failed to converge, so a GLM was used (as for the other species). Interpretation of the Skylark model outputs, therefore, needs some caution, although the level of spatial autocorrelation for the best model was not especially high (12%).
Results
In total, 862 individuals of 40 species were recorded in 221 point counts over an elevational range of 1750–2800 m a.s.l. There were seven species that were recorded on at least 15% of the points within forest and ecotone habitat: Dunnock, Lesser Whitethroat, Chaffinch Fringilla coelebs, Mistle Thrush Turdus viscivorus, Coal Tit Parus ater, Rock Bunting Emberiza cia, Tree Pipit and three species within open habitat: Eurasian Skylark, Water Pipit and Northern Wheatear.
The best model to predict Rock Bunting occurrence was always the null model for each model set at each scale, with no model-averaged parameter estimates being significant. Therefore, this species was not considered further in the analysis.
Forest and ecotone species
Habitat variables such as trees and shrubs were the variables most commonly associated with species occurrence within the HABITAT model for forest and ecotone species. In general, the results of the HABITAT models were in line with previous findings by Jähnig et al. (2018). Juniper showed a positive relationship with Coal Tit, Dunnock and Lesser Whitethroat, but was negatively related to Tree Pipit presence. Rhododendron was positively associated with Mistle Thrush and Lesser Whitethroat presence, whereas it showed a non-linear relationship with Dunnock presence. The number of mature trees showed a positive relationship with forest species (Chaffinch, Mistle Thrush and Coal Tit). Habitat associations among the species remained mostly constant in TEMP, TOPO and COMB models (for full details see ESM Table S2, S4).
Each variable set at each scale performed equally well for Lesser Whitethroat, Mistle Thrush and Coal Tit (Table 3). (Note that full details of all models are given in ESM Table S3). Large-scale temperature and topographic variables were included in the best performing model for Dunnock, temperature being negatively associated with Dunnock presence (Table 4, Fig. 1). In contrast, large-scale temperature showed a positive relationship with Chaffinch presence in models including only large-scale temperature (Table 4, Fig. 1), or in models including a combination of large-scale temperature and topographic variables. In both species, large-scale model sets performed better than their fine-scale equivalents. Large-scale models for TOPO and COMB were the best performing models for Tree Pipit, whose presence was more closely associated with large-scale topographic variables such as aspect, for which it showed a strong negative relationship indicating a preference for westerly over southerly slopes (Fig. 2). Beside Tree Pipit, only Mistle Thrush showed a negative association with aspect. No other species showed any association with slope or aspect. Furthermore, Tree Pipit was the only species that showed better model performance (ΔAICc ≤ 2) for the large-scale TOPO model compared to all fine-scale models and the large-scale TEMP model. All other species showed better (Chaffinch) or equal model performance of TEMP models compared to TOPO models at both scales.

Relationship between large-scale temperature and the probability of occurrence of Dunnock and Chaffinch based on the large-scale COMB model. Shading indicates the 95% confidence interval

Relationship between aspect and the probability of occurrence for Tree Pipit and Northern Wheatear for the large-scale TOPO and the fine-scale COMB model, respectively. Note that aspect was modelled as an index from 1 (south-facing) to − 1 (north facing), but here we present the axis as the equivalent cardinal direction for ease of interpretation. Shading indicates the 95% confidence interval
Open habitat species
The HABITAT model for each open species did not show any habitat associations among the recorded variables. However, all fine-scale models (TEMP, TOPO and COMB) showed a positive association between grass cover and Skylark presence while Juniper cover was only positively associated in the TEMP and COMP models.
Models including fine-scale temperature and topography performed best (ΔAICc ≤ 2) for Northern Wheatear. The best performing models of Skylark and Water Pipit included both fine-scale TEMP and COMB models. Fine-scale temperature was positively associated with Water Pipit and Northern Wheatear presence, whereas Eurasian Skylark presence was negatively associated (Table 4, Fig. 3).

Relationship between fine-scale temperature and probability of occurrence for open habitat species for the fine-scale COMB model. Shading indicates the 95% confidence interval
At a fine scale, TEMP models showed better model performance than TOPO models for Northern Wheatear and Water Pipit, whereas on a large scale, model sets for TEMP and TOPO were overlapping (Northern Wheatear, Water Pipit). The large-scale TOPO model showed equal model performance compared to the large-scale TEMP model for Skylark, but AICc was still higher compared to fine-scale COMB. In addition, aspect showed a significant relationship with Northern Wheatear (Fig. 2, fine-scale COMB model) and Skylark presence (large-scale TOPO model) while slope was positively related to Skylark presence in the fine-scale TOPO model.
Discussion
Models including fine-scale temperature estimates (TEMP, COMB) showed better model performance (ΔAICc < 2) than corresponding large-scale models for all three open habitat species. Northern Wheatear and Water Pipit were both positively associated with warm microclimates while Skylark presence was negatively associated with fine-scale temperature. These results contrast with previous findings from the same region of the Alps (Chamberlain et al. 2013, 2016), where model predictions were based on large-scale climatic variables. In these studies, SDMs (based on temperature change and assuming no change in habitat) suggested that under warmer conditions, Skylark and Northern Wheatear would show an increase in their distribution, whereas Water Pipit distribution would decrease. Therefore, for Water Pipit and Skylark distributions, our findings suggest opposite associations between fine-scale and large-scale temperature.
Differences in model predictions at different spatial scales have been reported for a range of studies, and thus identifying the appropriate scale represents a major problem when forecasting suitable habitat in order to inform conservation planning (Elith and Leathwick 2009; Randin et al. 2009; Franklin et al. 2013; Logan et al. 2013; Scridel et al. 2018). To improve SDMs, it is therefore necessary to carefully select predictors (e.g. temperature variables) and their spatial resolution. In the case of microclimate, local topography could create areas with suitable climatic conditions under which it would still be possible for a species to persist under the impact of climate change. Through the use of large-scale climate data, these areas might not be recognised by SDMs (Austin and Niel 2011). Besides affecting the future distribution of a species, microclimate can also influence many other aspects of a species’ life cycle.
There is evidence that microclimate can be important in influencing habitat selection in mountain birds which may explain our findings. For example, it has been shown that Horned Larks Eremophila alpestris adjusted the amount of incubation time in response to microclimatic conditions (Camfield and Martin 2009) by spending less time on the nest as temperatures in the nest surrounding increased, which may imply energy savings in warmer microclimates. Furthermore, microclimate and aspect strongly influenced nestling survival in Water Pipits (Rauter et al. 2002). Nests which were located at ENE-facing slopes (temperature maximum in the morning) had more fledglings than those on WSW-facing slopes (temperature maximum in the afternoon). In contrast, foraging habitat selection by alpine White-winged Snowfinches Montifringilla nivalis, a high altitude specialist, was influenced by solar radiation (Brambilla et al. 2017b). Snowfinches preferred to forage at colder sites (low solar radiation) throughout the season. These studies illustrate that behaviour, foraging habitat selection and choice of nest sites could be driven by microclimatic conditions thereby affecting bird species distributions. Therefore, we would strongly recommend considering microclimate as a predictor in future SDMs for open habitat Alpine species.
In contrast to the open habitat species considered, forest and ecotone species showed no association with fine-scale temperature. One possible reason might be the buffering effect of vegetation. Körner et al. (2007) showed that temperature can vary strongly between forest and open alpine grassland along the elevation gradient with intermediate values at the treeline ecotone. Furthermore, canopies can buffer the diurnal amplitude of air temperature in the forest (Chen et al. 1999).
For two species (Dunnock and Chaffinch) large-scale models including temperature (TEMP, COMB) performed better than fine-scale models. The probability of occurrence of Chaffinch was positively associated with large-scale temperature, whereas the probability of Dunnock presence was negatively affected. A future increase in temperature could, therefore, affect the distribution of Chaffinches by expanding its range towards higher elevations. In contrast, the distribution of Dunnocks might be severely limited. Bani et al. (2019) demonstrated that Dunnock distribution experienced a lower range contraction along the elevational gradient during the past 35 years, but a simple dispersal into higher elevations as a response to environmental change might not be possible because its preferred nesting habitat in our study area, Rhododendron, has a slow rate of colonisation to the extent that treeline shifts towards higher elevations are likely to be more rapid than upwards shifts in this species (Komac et al. 2016).
The mismatch between temperature and available future habitat can also affect open habitat species considered in this study. Due to increasing temperatures, shifts in major habitat types (i.e. forest and shrub encroachment; Harsch et al. 2009) may lead to habitat fragmentation and/or loss of open alpine grassland at higher elevations. This process might even be exacerbated by the abandonment of pastoral activities which formerly have maintained the forest limit at lower elevations than would be possible under climatic constraints only (Gehrig-Fasel et al. 2007).
We have considered the associations between bird species distributions and temperature estimated at different scales. In doing so, one of our goals was to compare our results with previous work that has focussed on temperature as a likely driver of species distributions. There seems little doubt that temperature has a direct effect on the distribution and behaviour of organisms along elevation gradients. Nevertheless, we must acknowledge that there are potentially other important climate variables that vary along the gradient, including light intensity, humidity, wind speed and precipitation that may affect bird species distributions and their invertebrate prey (e.g. Hodkinson 2005; Tingley et al. 2012) which would be worth considering in future research.
Topography
For the majority of species, COMB models performed equally well in comparison with TEMP models at both spatial scales. Combining temperature with topographic variables increased model performance only for Northern Wheatear at a fine scale, whose occurrence was more closely related with south-facing slopes. At a large scale, the probability of Tree Pipit presence was higher on westerly slopes. However, general topographic variables were rarely associated with species occurrence. The influence of aspect on the occurrence of some species could be explained by its effect on snow melt patterns during spring. Thermal differences among slopes with different exposition, which are caused by the amount of received solar radiation, could lead to an early snow melt on south-exposed slopes whereas north-exposed slopes might stay snow covered for a longer period (Keller et al. 2005). These early snow free areas could potentially benefit Northern Wheatears by making suitable nesting sites available earlier. Furthermore, it has been shown that differences in temperature among slopes can influence plant species diversity in temperate mountains (Winkler et al. 2016) with south-exposed slopes favouring a higher degree of species richness and diversity which may in turn influence insect availability. However, a potential caveat of this study might be that not all aspect directions were equally represented as we were tied to the south-western facing side of our study area due to frequent snow and rock avalanches and large inaccessible areas on the north-east facing side of Val Troncea Natural Park (ESM Figure S1). Therefore, aspect varied only from 123° to 311°, which might limit the transferability of our results. To complement our findings, further research is needed to fully understand how bird species distributions might be influenced by topographic variables in mountain areas of different exposition.
Conservation implications
Previous studies from the Italian Alps have indicated that increasing temperatures could have detrimental effects for certain Alpine species in the future (Chamberlain et al. 2013), with some species being potentially impacted by both temperature and habitat shifts (Water Pipit), while for others, loss of habitat due to forest and shrub encroachment will likely be more important (Northern Wheatear, Skylark). Indeed, Water Pipit in particular has shown declines at their lower elevation range margin in some areas that are apparently consistent with these predictions (e.g. Ebenhöh 2003; Bani et al. 2019). However, our results have shown that species such as Water Pipit and Northern Wheatear are positively associated with warm microclimates which could indicate that both species are potentially more resistant to the impact of a warming climate than previously emphasised by large-scale temperature modelling (e.g. Chamberlain et al. 2013). As a consequence, our results imply that changes in habitat in the form of advancing treelines and the encroachment of formerly open areas by shrubs and trees (Gehrig-Fasel et al. 2007; Leonelli et al. 2011) are currently the major threat to those Alpine species, rather than direct effects of temperature. Therefore, it becomes particularly important to actively manage open areas within mountain environments. This could be achieved by targeted grazing techniques such as mineral mix supplements (Pittarello et al. 2016) or temporary night camp areas (Tocco et al. 2013). Both techniques lead to the mechanical damage of shrubs (including saplings) and eventually result in a reduction of shrub cover (Probo et al. 2013, 2014).
References
Ashton S, Gutierrez D, Wilson RJ (2009) Effects of temperature and elevation on habitat use by a rare mountain butterfly: implications for species responses to climate change. Ecol Entomol 34:437–446. https://doi.org/10.1111/j.1365-2311.2008.01068.x
Austin MP, Van Niel KP (2011) Improving species distribution models for climate change studies: variable selection and scale. J Biogeogr 38:1–8. https://doi.org/10.1111/j.1365-2699.2010.02416.x
Avalos VD, Hernandez J (2015) Projected distribution shifts and protected area coverage of range-restricted Andean birds under climate change. Glob Ecol Conserv 4:459–469. https://doi.org/10.1016/j.gecco.2015.08.004
Bani L, Luppi M, Rocchia E, Dondina O, Orioli V (2019) Winners and losers: how the elevational range of breeding birds on Alps has varied over the past four decades due to climate and habitat changes. Ecol Evol 9:1289–1305. https://doi.org/10.1002/ece3.4838
Bartoń K (2013) MuMIn: Multi-model inference. R package version 1.9.0 ed
Barton MG, Clusella-Trullas S, Terblanche JS (2018) Spatial scale, topography and thermoregulatory behaviour interact when modelling species’ thermal niches. Ecography 41:1–14. https://doi.org/10.1111/ecog.03655
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2000) Bird census techniques. Academic Press, London
Brambilla M, Caprio E, Assandri G, Scridel D, Bassi E, Bionda R, Celada C, Falco R, Bogliani G, Pedrini P, Rolando A, Chamberlain D (2017a) A spatially explicit definition of conservation priorities according to population resistance and resilience, species importance and level of threat in a changing climate. Divers Distrib 23:727–738. https://doi.org/10.1111/ddi.12572
Brambilla M, Cortesi M, Capelli F, Chamberlain D, Pedrini P, Rubolini D (2017b) Foraging habitat selection by Alpine White-winged Snowfinches Montifringilla nivalis during the nestling rearing period. J Ornithol 158:277–286. https://doi.org/10.1007/s10336-016-1392-9
Brambilla M, Pedrini P, Rolando A, Chamberlain DE (2016) Climate change will increase the potential conflict between skiing and high-elevation bird species in the Alps. J Biogeogr 43:2299–2309. https://doi.org/10.1111/jbi.12796
Bramer I, Anderson BJ, Bennie J, Bladon AJ, De Frenne P, Hemming D, Hill RA, Kearney MR, Körner C, Korstjens AH, Lenoir J, Maclean IMD, Marsh CD, Morecroft MD, Ohlemüller R, Slater HD, Suggitt AJ, Zellweger F, Gillingham PK (2018) Advances in monitoring and modelling climate at ecologically relevant scales. Adv Ecol Res 58:101–161. https://doi.org/10.1016/bs.aecr.2017.12.005
Camfield AF, Martin K (2009) The influence of ambient temperature on horned lark incubation behaviour in an alpine environment. Behaviour 146:1615–1633. https://doi.org/10.1163/156853909x463335
Carroll JM, Davis CA, Elmore RD, Fuhlendorf SD, Thacker ET (2015) Thermal patterns constrain diurnal behavior of a ground-dwelling bird. Ecosphere. https://doi.org/10.1890/es15-00163.1
Carroll RL, Davis CA, Fuhlendorf SD, Elmore RD, DuRant SE, Carroll JM (2018) Avian parental behavior and nest success influenced by temperature fluctuations. J Therm Biol 74:140–148. https://doi.org/10.1016/j.jtherbio.2018.03.020
Chamberlain D, Brambilla M, Caprio E, Pedrini P, Rolando A (2016) Alpine bird distributions along elevation gradients: the consistency of climate and habitat effects across geographic regions. Oecologia 181:1139–1150. https://doi.org/10.1007/s00442-016-3637-y
Chamberlain DE, Negro M, Caprio E, Rolando A (2013) Assessing the sensitivity of alpine birds to potential future changes in habitat and climate to inform management strategies. Biol Conserv 167:127–135. https://doi.org/10.1016/j.biocon.2013.07.036
Chen JQ, Saunders SC, Crow TR, Naiman RJ, Brosofske KD, Mroz GD, Brookshire BL, Franklin JF (1999) Microclimate in forest ecosystem and landscape ecology—variations in local climate can be used to monitor and compare the effects of different management regimes. Bioscience 49:288–297. https://doi.org/10.2307/1313612
Ebenhöh H (2003) Zur Bestandsentwicklung von Berg- und Wiesenpieper (Anthus spinoletta und A. pratensis) am Feldberg im Schwarzwald. Naturschutz südl Oberrhein 4:11–19
Elith J, Leathwick JR (2009) Species distribution models: ecological explanation and prediction across space and time. Annu Rev Ecol Evol S 40:677–697. https://doi.org/10.1146/annurev.ecolsys.110308.120159
Engler JO, Stiels D, Schidelko K, Strubbe D, Quillfeldt P, Brambilla M (2017) Avian SDMs: current state, challenges and opportunities. J Avian Biol 48:1483–1504. https://doi.org/10.1111/jav.01248
Franklin J (2013) Species distribution models in conservation biogeography: developments and challenges. Divers Distrib 19:1217–1223. https://doi.org/10.1111/ddi.12125
Frey SJK, Hadley AS, Betts MG (2016) Microclimate predicts within-season distribution dynamics of montane forest birds. Divers Distrib 22:944–959. https://doi.org/10.1111/ddi.12456
Gehrig-Fasel J, Guisan A, Zimmermann NE (2007) Tree line shifts in the Swiss Alps: climate change or land abandonment? J Veg Sci 18:571–582. https://doi.org/10.1111/j.1654-1103.2007.tb02571.x
Gubler S, Fiddes J, Keller M, Gruber S (2011) Scale-dependent measurement and analysis of ground surface temperature variability in alpine terrain. Cryosphere 5:431–443. https://doi.org/10.5194/tc-5-431-2011
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8:993–1009. https://doi.org/10.1111/j.1461-0248.2005.00792.x
Gunton RM, Polce C, Kunin WE (2015) Predicting ground temperatures across European landscapes. Methods Ecol Evol 6:532–542. https://doi.org/10.1111/2041-210x.12355
Harsch MA, Hulme PE, McGlone MS, Duncan RP (2009) Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol Lett 12:1040–1049. https://doi.org/10.1111/j.1461-0248.2009.01355.x
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978. https://doi.org/10.1002/joc.1276
Hodkinson ID (2005) Terrestrial insects along elevational gradients: species and community responses to altitude. Biol Rev 80:489–513. https://doi.org/10.1017/S1464793105006767
Hof AR, Allen AM (2019) An uncertain future for the endemic Galliformes of the Caucasus. Sci Total Environ 651:725–735. https://doi.org/10.1016/j.scitotenv.2018.09.227
Jackson MM, Gergel SE, Martin K (2015) Effects of climate change on habitat availability and configuration for an Endemic Coastal Alpine Bird. PLoS ONE 10(11):e0142110. https://doi.org/10.1371/journal.pone.0142110
Jähnig S, Alba R, Vallino C, Rosselli D, Pittarello M, Rolando A, Chamberlain D (2018) The contribution of broadscale and finescale habitat structure to the distribution and diversity of birds in an Alpine forest-shrub ecotone. J Ornithol 159:747–759. https://doi.org/10.1007/s10336-018-1549-9
Keller F, Goyette S, Beniston M (2005) Sensitivity analysis of snow cover to climate change scenarios and their impact on plant habitats in alpine terrain. Clim Change 72:299–319. https://doi.org/10.1007/s10584-005-5360-2
Kleckova I, Konvicka M, Klecka J (2014) Thermoregulation and microhabitat use in mountain butterflies of the genus Erebia: importance of fine-scale habitat heterogeneity. J Therm Biol 41:50–58. https://doi.org/10.1016/j.jtherbio.2014.02.002
Komac B, Esteban P, Trapero L, Caritg R (2016) Modelization of the current and future habitat suitability of rhododendron ferrugineum using potential snow accumulation. PLoS ONE 11(1):e0147324. https://doi.org/10.1371/journal.pone.0147324
Körner C (2007) Climatic treelines: conventions, global patterns, causes. Erdkunde 61:316–324. https://doi.org/10.3112/erdkunde.2007.04.02
Lehikoinen A, Virkkala R (2016) North by north-west: climate change and directions of density shifts in birds. Glob Change Biol 22:1121–1129. https://doi.org/10.1111/gcb.13150
Leonelli G, Pelfini M, di Cella UM, Garavaglia V (2011) Climate warming and the recent Treeline shift in the European Alps: the role of geomorphological factors in high-altitude sites. Ambio 40:264–273. https://doi.org/10.1007/s13280-010-0096-2
Lewthwaite JMM, Angert AL, Kembel SW, Goring SJ, Davies TJ, Mooers AO, Sperling FAH, Vamosi SM, Vamosi JC, Kerr JT (2018) Canadian butterfly climate debt is significant and correlated with range size. Ecography 41:2005–2015. https://doi.org/10.1111/ecog.03534
Logan ML, Huynh RK, Precious RA, Calsbeek RG (2013) The impact of climate change measured at relevant spatial scales: new hope for tropical lizards. Glob Change Biol 19:3093–3102. https://doi.org/10.1111/gcb.12253
Moran PAP (1950) Notes on continuous stochastic phenomena. Biometrika 37:17–23. https://doi.org/10.2307/2332142
Ongaro S, Martellos S, Bacarol G, De Agostini A, Cogoni A, Cortis P (2018) Distributional pattern of Sardinian orchids under a climate change scenario. Community Ecol 19:223–232. https://doi.org/10.1556/168.2018.19.3.3
Pittarello M, Probo M, Lonati M, Bailey DW, Lombardi G (2016) Effects of traditional salt placement and strategically placed mineral mix supplements on cattle distribution in the Western Italian Alps. Grass Forage Sci 71:529–539. https://doi.org/10.1111/gfs.12196
Probo M, Lonati M, Pittarello M, Bailey DW, Garbarino M, Gorlier A, Lombardi G (2014) Implementation of a rotational grazing system with large paddocks changes the distribution of grazing cattle in the south-western Italian Alps. Rangeland J 36:445–458. https://doi.org/10.1071/rj14043
Probo M, Massolo A, Lonati M, Bailey DW, Gorlier A, Maurino L, Lombardi G (2013) Use of mineral mix supplements to modify the grazing patterns by cattle for the restoration of sub-alpine and alpine shrub-encroached grasslands. Rangeland J 35:85–93. https://doi.org/10.1071/rj12108
R Development Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing 3.5.2. R Foundation for Statistical Computing, Vienna
Randin CF, Engler R, Normand S, Zappa M, Zimmermann NE, Pearman PB, Vittoz P, Thuiller W, Guisan A (2009) Climate change and plant distribution: local models predict high-elevation persistence. Glob Change Biol 15:1557–1569. https://doi.org/10.1111/j.1365-2486.2008.01766.x
Rauter CM, Reyer HU, Bollmann K (2002) Selection through predation, snowfall and microclimate on nest-site preferences in the Water Pipit Anthus spinoletta. Ibis 144:433–444. https://doi.org/10.1046/j.1474-919X.2002.00013.x
Rodríguez JP, Brotons L, Bustamante J, Seoane J (2007) The application of predictive modelling of species distribution to biodiversity conservation. Divers Distrib 13:243–251. https://doi.org/10.1111/j.1472-4642.2007.00356.x
Scherrer D, Körner C (2010) Infra-red thermometry of alpine landscapes challenges climatic warming projections. Glob Change Biol 16:2602–2613. https://doi.org/10.1111/j.1365-2486.2009.02122.x
Scridel D, Brambilla M, Martin K, Lehikoinen A, Iemma A, Matteo A, Jähnig S, Caprio E, Bogliani G, Pedrini P, Rolando A, Arlettaz R, Chamberlain D (2018) A review and meta-analysis of the effects of climate change on Holarctic mountain and upland bird populations. Ibis 160:489–515. https://doi.org/10.1111/ibi.12585
Spasojevic MJ, Bowman WD, Humphries HC, Seastedt TR, Suding KN (2013) Changes in alpine vegetation over 21 years: Are patterns across a heterogeneous landscape consistent with predictions? Ecosphere. https://doi.org/10.1890/es13-00133.1
Stuhldreher G, Villar L, Fartmann T (2012) Inhabiting warm microhabitats and risk-spreading as strategies for survival of a phytophagous insect living in common pastures in the Pyrenees. Eur J Entomol 109:527–534. https://doi.org/10.14411/eje.2012.066
Tingley MW, Koo MS, Moritz C, Rush AC, Beissinger SR (2012) The push and pull of climate change causes heterogeneous shifts in avian elevational ranges. Glob Change Biol 18:3279–3290. https://doi.org/10.1111/j.1365-2486.2012.02784.x
Tocco C, Probo M, Lonati M, Lombardi G, Negro M, Nervo B, Rolando A, Palestrini C (2013) Pastoral practices to reverse shrub encroachment of Sub-Alpine Grasslands: dung beetles (Coleoptera, Scarabaeoidea) respond more quickly than vegetation. PLoS ONE 8(12):e83344. https://doi.org/10.1371/journal.pone.0083344
Tomecek JM, Pierce BL, Reyna KS, Peterson MJ (2017) Inadequate thermal refuge constrains landscape habitability for a grassland bird species. Peer J. https://doi.org/10.7717/peerj.3709
Visinoni L, Pernollet CA, Desmet JF, Korner-Nievergelt F, Jenni L (2015) Microclimate and microhabitat selection by the Alpine Rock Ptarmigan (Lagopus muta helvetica) during summer. J Ornithol 156:407–417. https://doi.org/10.1007/s10336-014-1138-5
Viterbi R, Cerrato C, Bassano B, Bionda R, von Hardenberg A, Provenzale A, Bogliani G (2013) Patterns of biodiversity in the northwestern Italian Alps: a multi-taxa approach. Community Ecol 14:18–30. https://doi.org/10.1556/ComEc.14.2013.1.3
Wachob DG (1996) The effect of thermal microclimate on foraging site selection by wintering Mountain Chickadees. Condor 98:114–122. https://doi.org/10.2307/1369514
Wilson RJ, Bennie J, Lawson CR, Pearson D, Ortuzar-Ugarte G, Gutierrez D (2015) Population turnover, habitat use and microclimate at the contracting range margin of a butterfly. J Insect Conserv 19:205–216. https://doi.org/10.1007/s10841-014-9710-0
Winkler M, Lamprecht A, Steinbauer K, Hulber K, Theurillat JP, Breiner F, Choler P, Ertl S, Giron AG, Rossi G, Vittoz P, Akhalkatsi M, Bay C, Alonso JLB, Bergstrom T, Carranza ML, Corcket E, Dick J, Erschbamer B, Calzado RF, Fosaa AM, Gavilan RG, Ghosn D, Gigauri K, Huber D, Kanka R, Kazakis G, Klipp M, Kollar J, Kudernatsch T, Larson P, Mallaun M, Michelsen O, Moiseev P, Moiseev D, Molau U, Mesa JM, di Cella UM, Nagy L, Petey M, Puscas M, Rixen C, Stanisci A, Suen M, Syverhuset AO, Tomaselli M, Unterluggauer P, Ursu T, Villar L, Gottfried M, Pauli H (2016) The rich sides of mountain summits—a pan-European view on aspect preferences of alpine plants. J Biogeogr 43:2261–2273. https://doi.org/10.1111/jbi.12835
Wood SN (2011) Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J R Stat Soc B 73:3–36. https://doi.org/10.1111/j.1467-9868.2010.00749.x
Wood SN (2017) Generalized additive models: an introduction with R, 2nd edn. Chapman and Hall, CRC Press, New York. https://doi.org/10.1201/9781315370279
Zabala J, Zuberogoitia I, Belamendia G, Arizaga J (2012) Micro-habitat use by Bramblings Fringilla montifringilla within a winter roosting site: influence of microclimate and human disturbance. Acta Ornithol 47:179–184. https://doi.org/10.3161/000164512x662287
Zellweger F, De Frenne P, Lenoir J, Rocchini D, Coomes D (2019) Advances in microclimate ecology arising from remote sensing. Trends Ecol Evolut 34(4):327–341
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
We thank all rangers and staff of Val Troncea Natural Park for their invaluable assistence. We are grateful to Nadja Schäfer and Riccardo Alba for help with the field work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Gottschalk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Jähnig, S., Sander, M.M., Caprio, E. et al. Microclimate affects the distribution of grassland birds, but not forest birds, in an Alpine environment. J Ornithol 161, 677–689 (2020). https://doi.org/10.1007/s10336-020-01778-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-020-01778-5