
Abstract
High-elevation species are expected to be vulnerable to climate warming and to experience dramatic range contractions in the coming decades. Indeed, climate change in high-altitude biota has proceeded at a faster pace compared to lowlands. Understanding basic ecological features of mountain species, such as their foraging ecology, may be useful to further our understanding of the mechanisms dictating species distributions and their responses to global warming, ultimately improving conservation strategies. In this study, we investigated foraging habitat selection of the poorly studied Alpine White-Winged Snowfinch Montifringilla nivalis during the nestling rearing period (June–July) in the Italian Alps. Pair members from 18 different nests were visually followed for 1 day or until we obtained 10 foraging locations. At foraging and control plots (equal numbers per breeding pair; control plots within 300 m of the nest, the average exploited radius according to literature) we recorded habitat variables (habitat types/heterogeneity, sward height, slope, solar radiation). We built models of foraging habitat selection and evaluated whether the selection of climate-related variables varied with temperature and season progression. Snowfinches preferred to forage at colder (low solar radiation) sites, with snow patches and short grasses, some boulders and bare ground, and shifted towards sites with increasingly lower solar radiation after the first week of July. Snow patches are likely to provide both arthropod fallout and suitable sites for invertebrates at their melting margins. Short herbaceous layers likely improved invertebrate detectability in addition to their abundance. These results suggested that climate change may impact on the foraging ecology of this species: warming may reduce the availability of snow patches and favor a denser and taller sward, and may reduce the time frame within which melting snow patches are highly profitable. Hence, the fine-scale habitat requirements of foraging Snowfinches highlight the species’ potential high sensitivity to anthropogenic climate warming.
Zusammenfassung
Futterhabitatwahl bei Schneefinken während der Nestlingsaufzucht Es ist zu erwarten, dass Arten der Hochlagen anfällig sind gegenüber Klimaerwärmung, und dass sie in den kommenden Jahrzehnten eine dramatische Verkleinerung ihres Lebensraumes erfahren. In der Tat ist der Klimawandel in hochgelegenen Biota schneller fortgeschritten als in niedriger gelegenen Gebieten. Das Verständnis grundlegender ökologischer Merkmale von Gebirgs-Arten, wie ihre Futtersuche, könnte nützlich sein, um unser Verständnis für die Mechanismen zu erweitern, die die Verbreitung einer Art und ihre Reaktion auf globale Erwärmung bedingen, um schließlich die Strategien im Artenschutz zu verbessern. In dieser Studie untersuchten wir die Auswahl von Futterhabitaten beim wenig erforschten Schneefinken (Montifringilla nivalis) in den italienischen Alpen während der Zeit der Nestlingsaufzucht (Juni-Juli). Die Individuen der Paare aus 18 verschiedenen Nestern wurden über einen Tag, oder bis wir 10 Futterplätze aufgenommen hatten, verfolgt. An Futter- und Kontrollplätzen (gleiche Anzahl bei Brutpaaren; Kontrollplätze innerhalb von 300 m vom Nest, laut Literatur der mittlere ausgenutzte Radius) nahmen wir Habitat-Variablen auf (Habitattyp/-heterogenität, Höhe der Grasnarbe, Neigung, Sonneneinstrahlung). Wir erstellten Modelle für eine Futterhabitat-Auswahl und überprüften, ob sich die Auswahl von klimabezogenen Variablen mit der Temperatur und fortschreitender Saison änderte. Schneefinken zogen es vor, an kälteren Stellen (geringere Sonneneinstrahlung) mit Schneeflecken und kurzem Gras, einigen Steinen und offenem Boden auf Futtersuche zu gehen, und wechselten nach der ersten Juli-Woche an Stellen mit zunehmend geringerer Sonneneinstrahlung. Schneeflecken stellen vermutlich sowohl Arthropoden-Fallout zur Verfügung als auch geeignete Stellen für Invertebraten an ihren Rändern. Kurze krautige Bewüchse erleichterten das Auffinden der Invertebraten und vergrößerten deren Abundanz. Diese Ergebnisse legen nahe, dass der Klimawandel einen Einfluss haben könnte auf die Futtersuche dieser Art: eine Erwärmung könnte die Verfügbarkeit von Schneeflecken reduzieren und eine dichtere und höhere Grasnarbe bevorzugen, und sie könnte das Zeitfenster verkleinern, in der schmelzende Schneeflächen hochprofitabel sind. Daher betonen die kleinräumigen Bedürfnisse von Schneefinken an ihr Futterhabitat die potentiell hohe Empfindlichkeit dieser Art gegenüber anthropogener Klimaerwärmung.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Species living in harsh environments often display specific adaptations to extreme environmental conditions, and this makes them particularly susceptible to environmental changes (e.g., Lu et al. 2009). A case in point is represented by high-elevation species, which are generally well adapted to cold climates and/or sudden changes in resource availability (Muscio et al. 2005; Rosvold 2016). These species are expected to be particularly vulnerable to anthropogenic climate warming (Sekercioglu et al. 2008; Dirnböck et al. 2011; Chamberlain et al. 2012). At high altitudes, climate warming has been progressing at a greater rate than the global average over the past two centuries, a trend which is expected to continue (IPCC 2013; Gobiet et al. 2014). Identification of key habitat resources for such potentially threatened species is therefore important if management strategies are to be developed which may ameliorate negative impacts of climate change (e.g., Braunisch et al. 2014).
Bird assemblages from high elevation zones often include a relatively low number of cold-adapted species that occur almost exclusively in these areas. The White-Winged Snowfinch Montifringilla nivalis (henceforth Snowfinch) is among the species most tied to high-altitude areas (Muscio et al. 2005) and is predicted to undergo a dramatic range contraction over the course of the twenty-first century in the Alps because of a warming climate (Maggini et al. 2014; Brambilla et al. 2016b). In spite of these worrisome future predictions of the species’ distributional changes in relation to climate change, there is a knowledge gap in the basic ecology of this species, including its foraging habits, a feature shared with other high-altitude taxa (Chamberlain et al. 2012). Indeed, the limited available information mostly comes from non-Alpine populations (e.g., Antor 1995; Strinella et al. 2007), notwithstanding that the Alps host ca. 12 % of the continental population of the species and 70–80 % of the nominal subspecies’ population (population size estimates according to BirdLife International 2015).
Assessing the fine-scale foraging habitat selection of the Snowfinch may be helpful in understanding the mechanisms dictating its current distribution, and therefore in improving predictions of the response of the species to future global warming. Moreover, it will be helpful in formulating management strategies to ameliorate the potential negative effects of environmental change. In fact, even if distribution modelling techniques suggest a dramatic impact of climate warming on Snowfinch distribution (Brambilla et al. 2016b), it is possible that ecological features at a finer scale may to some extent modulate the impact of climate change at a larger scale, in both negative and positive ways. For instance, fine-scale habitat selection could buffer the impact of climate warming if a species mostly uses habitats and resources likely to be unaffected by increasing temperatures. On the other hand, if a species largely relies on resources that are dependent on climate the potential impact of warming may be even stronger than suggested by distribution modelling analyses.
The few studies carried out in non-Alpine populations suggest that snow patches (especially melting borders) and grassland may constitute crucial foraging habitats for Snowfinches, especially during the nestling rearing period (Catzeflis 1975; Cramp and Perrins 1994; Antor 1995; Strinella et al. 2007; Grangé 2008). Melting snow patches harbor abundant dipteran larvae (especially Tipulidae; Muscio et al. 2005), a fundamental component of the Snowfinch diet during nestling rearing (Cramp and Perrins 1994; Grangé 2008). Alpine grasslands may harbor the highest densities of high-elevation arthropods, but prey detectability may be much lower than in snowfields (Antor 1995). Sward height (and its heterogeneity), together with the occurrence of patches of bare ground within vegetated areas, may thus be important in driving foraging habitat selection: lower sward height, higher heterogeneity, and occurrence of bare ground patches are all factors increasing prey detectability (Arlettaz et al. 2012; Vickery and Arlettaz 2012; Brambilla et al. 2013).
In addition, habitat selection patterns may vary during the course of the breeding season and in relation to variation in daily temperature: hence, seasonal changes in habitat selection patterns may further help in understanding the potential effect of climate change on the Alpine Snowfinch’s foraging ecology.
In this study, we aimed to investigate foraging habitat selection in Snowfinches from the Italian Alps by analyzing foraging trips of breeding pairs raising nestlings, and concurrently recording habitat variables at foraging and control sites on a daily basis throughout the breeding season. We paid special attention to the recording of habitat variables that undergo dramatic changes during the spring season in the Alps, such as vegetation height (grass height increases during the spring) and snow cover (progressively disappearing during the season), and which could be particularly important for foraging Snowfinches.
On the basis of the limited anecdotal information available, we expected cover of snow and/or grassland, the habitats hosting the highest prey abundance (grassland and melting margins of snow patches), to rank among the most important predictors of foraging habitat selection.
Given that the Snowfinch is a cold-adapted species, it is also possible that the species’ preferred habitat could shift to colder sites as the season progresses, or when the temperature increases: we thus expected a differential selection of foraging sites according to solar radiation, with less exposed sites preferred later in the season and/or on the warmest days, whereas snow cover could be of greater importance during the melting phase (i.e., at intermediate or increasing temperatures).
We thus aimed to investigate (1) whether the preferred foraging habitat includes features known to affect prey abundance and/or detectability, and in particular grassland and snow; and (2) whether habitat selection shifts during the season or with increasing temperature, with particular reference to the pattern of association with solar radiation and snow cover.
Methods
Snowfinch biology and ecology
The Snowfinch lays one or two broods per season. In the Alps, egg laying begins mostly in the second half of May (Cramp and Perrins 1994). The nest is placed in cavities such as rock crevices or holes in buildings. Females lay and brood clutches of 3–6 eggs that are incubated for 12–16 days. Nestlings fledge at 18–22 days and are fed by both parents (Cramp and Perrins 1994; Grangé 2008; Strinella et al. 2011). Fledging occurs mostly between June and July for first broods and between July and August for second broods (Cramp and Perrins 1994; Grangé 2008). The species is largely resident in the Alps, where it feeds on both seeds and invertebrates, with the former predominating in winter, while insects and spiders are preferred in spring-summer (Cramp and Perrins 1994). Nestlings are fed almost exclusively with invertebrates (Cramp and Perrins 1994).
Study area and field protocols
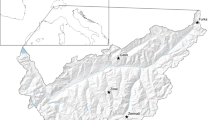
We investigated Snowfinch foraging habitat selection in the central Italian Alps (latitude 46.27°–46.54° N, longitude 9.33°–11.84° E; provinces of Sondrio, Brescia and Trento) (Fig. 1). Fieldwork was undertaken during spring-summer 2015 by carrying out field observations at nest sites in eight study locations (Table 1) located at different altitudes (1900–2825 m a.s.l.).

Location of study locations (white dots) in the central Alps (northern Italy). The black line is the Italian boundary; the mountain relief is shown for Lombardy and Trento Province. The inset shows the location of the study area within the Alps
At all sites, we only considered Snowfinch pairs raising nestlings, as nestling rearing is a crucial phase of the avian life cycle and adults behave as central-place foragers. After locating occupied nests, we followed each pair (individuals moving from the nest) for one entire day or until we obtained 10 foraging locations.
We carefully recorded the exact location of each foraging event on detailed aerial photographs (scale 1:2000). After a bird landed, the position where it collected the food item(s) was taken as the exact position for habitat measurement (i.e., a foraging patch). In the few cases where it was not possible to observe a prey capture event (e.g., when the bird was partially obscured, or when it was too far away), the final position observed before flying back to the nest was taken as the location of the foraging patch, as Snowfinches typically return to the nest immediately after a successful capture (pers. obs.). Every hour we recorded air temperature (at a fixed position per each breeding pair, corresponding to our watch point) by means of a mercury thermometer held in a box 1 m above the ground.
Each breeding pair was surveyed once, and each study location was visited 1–5 times, according to the number of active breeding pairs.
Habitat recording
At each foraging patch, and in an equal number of control plots (see below), we recorded a number of predefined habitat variables, which were measured within a 10-m radius centered on the patch (Table 2). Habitat variables referred to habitat types, habitat heterogeneity, sward height, and topographic factors. Habitat types were measured as their proportional cover at a very fine spatial resolution (mapping all features with a linear extent ≥50 cm). Habitat heterogeneity was expressed as the number of different habitat patches (i.e., single units of the habitat types measured). Sward height was measured in centimeters (taken as 0 in the case of no grassland cover), and calculated as the average of five measurements at each plot (at the center and at the four cardinal points). The standard deviation was also calculated as a measure of the heterogeneity of grass height, which could potentially affect invertebrate abundance and detectability. Topographic variables included slope and solar radiation (both computed using GRASS 6.4.4 GIS software, commands r.slope.aspect and r.sun, respectively), calculated by means of a 10-m-resolution digital elevation model (Tarquini et al. 2012). Control plots were established on the basis of a predefined set of randomly scattered points within a 300-m radius of each nest, which corresponds to the main food searching area around the nest during nestling rearing (Strinella et al. 2007; Grangé 2008; personal observations). If random points coincided with foraging plots, they were replaced by other predefined points according to a previously established (random) order. In that way, we avoided any bias due to observers’ non-random selection of control plots.
Statistical analyses
All analyses were performed by means of the software R 3.2.1 (R Development Core Team 2015). We adopted a two-step approach, according to the two aims of our study. First, we built models of foraging habitat selection. Then, we evaluated whether the selection of specific habitat factors potentially related to climate varied according to temperature and season progression in order to have an indication of the potential impact of climate change on the foraging ecology of the species.
We modelled the probability of occurrence of foraging Snowfinches at a given plot (foraging vs. control) by means of multivariate adaptive regression splines (MARS, Friedman 1991), a machine-learning, non-parametric regression technique frequently adopted in autoecological studies (e.g., Leathwick et al. 2005; Jedlikowski et al. 2014). It is particularly suited to investigating habitat selection in species inhabiting or using heterogeneous habitats owing to its flexibility in modelling non-linear and interaction effects (Brambilla et al. 2016a).
We omitted from the analyses those habitat types which were less frequently recorded (mean cover lower than or equal to 2 %: cover of sand, garbage, shrub, dirt tracks, paved roads, urbanized land) and the cover of scree, which led to convergence problems. Notably, the mean scree cover was nearly exactly the same in foraging and control plots. We therefore tested the following predictors: slope, solar radiation, snow, water, rock, boulders, bare ground, grassland, number of patches, sward height, standard deviation of sward height. We included four two-way interactions that we deemed ecologically plausible. For instance, we speculated that Snowfinches forage preferentially (1) on snow patches close to water or vice versa (snow patch × water cover); (2) in compact mosaics of snow and grass (snow × grass cover); (3) at the interface between water and grass (water × grass cover). Close proximity of these habitats at a fine scale may indeed lead to the margins of melting snow patches, pools, and other contexts as being identified as important foraging habitats, where some invertebrates (and in particular their larvae) may be particularly frequent and/or abundant (Muscio et al. 2005). Furthermore, it is also possible that grassland cover and sward height might jointly affect prey availability and detectability: prey can be abundant in grassland, but may be difficult to detect over large grassland patches with a (relatively) tall sward. We therefore tested also (4) the grass cover × sward height interaction.
We adopted the following settings for model selection: threshold = 0.001, degree of interactions = 2, penalty = 3. Notably, adopting a penalty value of 4 instead of 3 (the latter is the value commonly adopted when degree of interactions is 2) led to exactly the same model, removing the biologically meaningless drop in occurrence probability at intermediate levels of number of patches (see Fig. 2), but resulted also in the exclusion of cover of bare ground. We carried out a fivefold model cross-validation, from which we estimated model performance, calculated over different data subsets.

Predicted probability of occurrence from MARS models of foraging habitat selection in Snowfinches. The plots with three axes illustrate interactions. The values on the x-axes represent the percentage cover of a given habitat within the plot (all variables) or height in cm (sward height)
We used the evimp command to estimate variable importance (Milborrow 2011a) in three different ways: (1) the frequency that a given variable is present in the different model subsets generated by the pruning pass; (2) the sum of the decreases (rescaled to a percentage) in the residual sum of squares (RSS) for model subsets including a given variable relative to the previous subset; and (3) the generalized cross-validation (GCV) of the model, computed by means of the penalty argument, under the same approach as the RSS criterion (Milborrow 2011a).
The earth package was used to run the models and to estimate variable importance (Milborrow 2011a), and model fitted functions were plotted using the plotmo package (Milborrow 2011b).
We also ran conditional logistic regression analyses (Hosmer and Lemeshow 1989), accounting for the paired nature of the dataset, whereby each set of foraging plots for a given pair was matched to the set of control plots for that pair, thus controlling for pair-based sampling. In these analyses, pair identity was specified as the stratum variable. In that way, we checked for potential effects of non-independent data. We started by fitting the same variables entered in the MARS model (VIF < 2 for all variables), adding the quadratic term for snow (as an association with intermediate snow cover is likely; see above), and testing the interaction terms between snow and grassland cover, and between grassland cover and sward height (highlighted as important by MARS analyses; see “Results”). Then, we removed non-significant (p > 0.05) interactions in a single step and assessed the coefficients and p values for all remaining predictors. We also ran univariate analyses testing the effect of each predictor individually, to assess how collinearity among predictors affected the outcome of the multivariate analysis.
To evaluate whether selection of habitat traits varied with temperature and season progression, we first calculated, for all the variables deemed as important by the habitat selection models (solar radiation, number of patches, sward height, and cover of snow, boulders, bare ground, and grassland; see “Results”), the log10-ratio of the mean values of foraging and control points (values equal to 0 were adjusted to 0.1) for each pair, and took this proportion as an index of habitat use. Hence, these analyses included a single datum for each breeding pair for each habitat variable (sample size for the analyses, n = 18). Log-ratio values greater than 0 indicate positive selection for a given habitat variable, whereas values less than 0 indicate avoidance. We then built further MARS models to evaluate the variation in the index of habitat use according to date (day of year; days since 4 June, which was the first day of fieldwork) and temperature (entered as the mean value of all hourly records at a given site/day). We adopted the following setting: degree = 2, penalty = 3. We did not carry out any cross-validation in this case, given that we wanted to explore a pattern (on a rather small dataset, N = 18 pairs), rather than to produce a habitat selection model. To check for consistency with the MARS results, we also carried out a simple correlation analysis by means of the Spearman’s rank order correlation test, relating the log-ratios of different habitat variables with season or temperature. We acknowledge that the scope of this analysis was mostly exploratory because of the limited sample size.
Results
Foraging habitat selection
Overall, we surveyed 18 breeding pairs and obtained data for 134 foraging plots and an equal number of control plots. The model for foraging habitat selection suggested that Snowfinches preferentially searched for food at sites characterized by lower solar radiation and a higher number of habitat patches (Fig. 2). The occurrence of foraging individuals was generally positively associated with the cover of snow, grass, boulders, and bare ground. There was also evidence for threshold effects, in that the probability of occurrence of foraging Snowfinches reached an asymptote at ca. 40 % snow cover and at ca. 40 % boulder cover, and was rather constant up to 15 habitat patches (the slight drop associated with 6–14 patches is likely a statistical artefact and not a biologically relevant effect), after which it increased in a mostly linear fashion (Fig. 2). Two interaction effects emerged: Snowfinch occurrence probability was particularly low at sites with no or very low grassland cover and low snow cover, while it was highest at sites with high grassland cover and low sward height.
The performance of the MARS model was not particularly high, with R 2 equal to 0.30, decreasing to 0.15 ± 0.09 according to the fivefold cross-validation, which also had an AUC equal to 0.75 ± 0.06. The evimp command confirmed the validity of the model, and suggested that grassland and sward height were the most important factors affecting habitat choice, followed by the number of patches and the cover of snow (Table 2).
The univariate conditional logistic regression analyses (Table 3) suggested that a significant (p < 0.05) positive effect on Snowfinch occurrence was exerted by cover of bare ground and number of habitat patches. Sward height and its standard deviation had a highly significant (p < 0.01) negative effect. The two interaction terms (snow × grassland cover and grassland cover × sward height) were also statistically significant (Table 3). The effect of snow cover was highly significant and non-linear (Table 3), a pattern similar to that highlighted by the MARS analyses. Furthermore, a marginally significant (0.05 < p < 0.10) negative effect of rock cover was found. The multiple conditional logistic regression model did not confirm the univariate effects of standard deviation of sward height and snow cover, whereas it suggested a marginally significant positive effect of boulder cover and a negative effect of solar radiation (Table 3). On the whole, the multiple conditional regression model supported the conclusions drawn from the MARS model, showing statistically significant positive effects of grassland cover, bare ground and habitat heterogeneity (number of habitat patches) and negative effects of sward height (Table 3). Moreover, the effect of snow cover varied according to grassland cover, with sites with more grassland and snow cover being preferred over sites with low grassland and snow cover (Table 3).
Seasonal and temperature-driven patterns of habitat selection
The analysis of the potential effect of date and temperature on the index of habitat use revealed significant variation only for solar radiation in relation to season progression (MARS model R 2 = 0.42). While solar radiation did not appear to affect habitat use up to 7 July, Snowfinches showed a strong shift to foraging at sites with lower solar radiation thereafter (Fig. 3). For all the other variables, the MARS analyses provided no evidence for different selection patterns along seasonal and temperature gradients. Indeed, the null MARS model (no effect of season and mean daily temperature on the index of habitat selection) had the highest support for the index of habitat use of number of patches, sward height, cover of snow, boulders, bare ground, and grassland. Simple correlation analyses of log-ratios of different habitat variables with season or temperature gave some support to the results of the MARS analyses (seasonal changes in the index of habitat use with season: solar radiation, r s = −0.41, p = 0.087; all other variables, season: p always >0.50; all variables, temperature: p > 0.22; n = 18 pairs).

Season-dependent variation in the index of habitat use for solar radiation by foraging pairs of Snowfinches according to the specific MARS model. Black dots represent observed values for the 18 pairs
Discussion
High-elevation species such as the Snowfinch are well adapted to extreme environmental conditions, and this makes them particularly vulnerable to perturbation (as already reported for other Montifringilla species; Lu et al. 2009). Because of the challenging environment they occupy, the ecology and population dynamics of high-altitude species are little studied (Chamberlain et al. 2012). Here, we provide the first quantitative description of foraging habitat selection in Snowfinches breeding in the Alps during the critical nestling rearing period.
Our results strongly suggest that Snowfinches tend to forage at colder sites with snow patches and with short swards. Such results confirm the general importance of both habitat and climate for alpine bird species (e.g., Muscio et al. 2005; Chamberlain et al. 2016a, b), which indeed are highly sensitive to environmental, and in particular climatic and climate-induced, changes (e.g., Chamberlain et al. 2013; Maggini et al. 2014; Brambilla et al. 2015). Both the availability of snow patches and grass development are largely influenced by climate, and in particular by the length of the snow-cover period (Muscio et al. 2005). Snow patches probably offer both arthropod fallout over their surface (Antor 1995; Rosvold 2016), and suitable sites for dipteran larvae at their melting margins (Muscio et al. 2005). The latter source of food has been reported as particularly important for Snowfinches (Muscio et al. 2005; Strinella et al. 2007), and indeed we mostly observed Snowfinches foraging at the margins of snow patches. Arthropods that alight on the snow surface are highly detectable and accessible (Rosvold 2016), whereas alpine grasslands generally harbor more abundant and larger invertebrate prey, which are however less detectable (Antor 1995). Snowfinches made use of both snow patches and grassland, favoring short herbaceous layers, a condition that is expected to favor invertebrate detectability in addition to their abundance. Snow patches are particularly important for several bird and mammal species adapted to cold climates, which may find, e.g., food or resting sites on or around snowfields and glacier areas; all those species are likely to be highly threatened by climate change and its consequences on Alpine ecosystems (Rosvold 2016).
The index of habitat use revealed a consistent selection for less exposed sites as the season progressed, reflecting a tendency to rely on cooler sites. Hence, birds increasingly selected cooler locations in which to forage as the overall ambient temperature increased with season, as expected for a cold-adapted organism. However, no seasonal effect was found for snow cover. It is possible that our study design did not allow us to detect significant changes in the use of snow patches in relation to date and temperature simply because snow cover obviously tended to disappear at later dates and higher temperatures, implying that no use was possible under those conditions. Nevertheless, we cannot exclude that the lack of seasonal effect was due to the fact that snow patches were rather constantly used by the species (as long as they were available) (see, e.g., Strinella et al. 2007).
The particular pattern of foraging habitat selection by breeding Snowfinches confirmed how this species could be highly sensitive to climate change throughout its Alpine range (Maggini et al. 2014; Brambilla et al. 2016b). Climate warming in particular may strongly affect the suitability of foraging habitats by reducing the availability of snow patches, by favoring a denser and taller sward because of a shorter period of snow cover, and by reducing the time frame within which melting snow patches and the associated pools are highly profitable. Phenological changes in the timing of birds’ breeding seasons and in the timing of key resources have been a recent major topic in climate change research (Charmantier and Gienapp 2014). However, there are few studies that have considered such effects in a mountain context, yet the harsh environment and steep environmental gradients over small spatial scales may mean that such effects are important (e.g., Inouye et al. 2000). Specifically, the likely effects of climate warming on the extent and duration of snow patches in spring and summer, and the consequences for invertebrate populations, should be research priorities for alpine birds.
There are further specific studies that are needed to more fully understand potential impacts of environmental change on the Snowfinch. First, the potential variation in snow-patch use along the season and at different temperatures needs to be further evaluated in other areas in the Alps and at other spatial scales (e.g., at an even finer level, with data gathered from radio-tracked individuals, and at a larger spatial extent in terms of both geographical coverage and patch area). Ultimately, such information could be used to assess the importance of the phenology of snowmelt to the Snowfinch, and hence to predict potential future consequences of warmer conditions on snow cover. Second, the potential benefits deriving from suitable grassland management (e.g., through grazing) need to be assessed in order to make specific management recommendations which could ultimately be used to compensate for the negative impacts of climate change. Such management could be targeted at controlling sward height while promoting small-scale heterogeneity, enhancing a fine-scale, compact mosaic of grass cover (with patches of different sward height) and bare ground, which is likely to prove particularly suitable for several bird species feeding on grassland arthropods (see, e.g., Arlettaz et al. 2012; Vickery and Arlettaz 2012; Brambilla et al. 2013, 2016a). Third, whilst being a high-altitude specialist, the species also shows some affinity to disturbed habitats, and commonly uses buildings and other structures for nesting, and is often seen foraging for (human) food remains at high-altitude ski resorts in winter (Cramp and Perrins 1994). The extent to which populations rely on such anthropomorphic “subsidies”, and the role this might play in the conservation of the species, is therefore worthy of further research.
In conclusion, our work has contributed to the knowledge of fine-scale habitat associations of a poorly studied species inhabiting high-elevation sites (Chamberlain et al. 2012). The fine-scale habitat requirements shown by foraging Snowfinches during the nestling period have further highlighted the potential high sensitivity of this species to anthropogenic climate warming.
References
Antor RJ (1995) The importance of arthropod fallout on snow patches for the foraging of high-alpine birds. J Avian Biol 26:81–85
Arlettaz R, Maurer ML, Mosimann-Kampe P, Nusslé S, Abadi F, Braunisch V, Schaub M (2012) New vineyard cultivation practices create patchy ground vegetation, favouring Woodlarks. J Ornithol 153:229–238
BirdLife International (2015) European red list of birds. Office for Official Publications of the European Communities, Luxembourg
Brambilla M, Fulco E, Gustin M, Celada C (2013) Habitat preferences of the threatened Black-eared Wheatear Oenanthe hispanica in southern Italy. Bird Study 60:432–435
Brambilla M, Bergero V, Bassi E, Falco R (2015) Current and future effectiveness of Natura 2000 network in the central Alps for the conservation of mountain forest owl species in a warming climate. Eur J Wildl Res 61:35–44
Brambilla M, Gustin M, Vitulano S, Negri I, Celada C (2016a) A territory scale analysis of habitat preferences of the declining Ortolan Bunting Emberiza hortulana. Bird Study 63:52–57
Brambilla M, Pedrini P, Rolando A, Chamberlain DE (2016b) Climate change will increase the potential conflict between skiing and high-elevation bird species in the Alps. J Biogeogr. doi:10.1111/jbi.12796
Braunisch V, Coppes J, Arlettaz R, Suchant R, Zellweger F, Bollmann K (2014) Temperate mountain forest biodiversity under climate change: compensating negative effects by increasing structural complexity. PLoS One 9(5):e97718
Catzeflis F (1975) Remarques sur la nidification rupestre de la Niverolle. Nos Oiseaux 33:64–65
Chamberlain D, Arlettaz R, Caprio E, Maggini R, Pedrini P, Rolando A, Zbinden N (2012) The altitudinal frontier in avian climate change research. Ibis 154:205–209
Chamberlain DE, Negro M, Caprio E, Rolando A (2013) Assessing the sensitivity of alpine birds to potential future changes in habitat and climate to inform management strategies. Biol Conserv 167:127–135
Chamberlain D, Brambilla M, Caprio E, Pedrini P, Rolando A (2016a) Alpine bird distributions along elevation gradients: the consistency of climate and habitat effects across geographic regions. Oecologia 181:1139–1150
Chamberlain DE, Pedrini P, Brambilla M, Rolando A, Girardello M (2016b) Identifying key conservation threats to Alpine birds through expert knowledge. Peer J 4:e1723
Charmantier A, Gienapp P (2014) Climate change and timing of avian breeding and migration: evolutionary versus plastic changes. Evol Appl 7:15–28
Cramp S, Perrins CM (eds) (1994) The birds of the Western Palearctic, vol VIII. Oxford University Press, Oxford
Dirnböck T, Essl F, Babitsch W (2011) Disproportional risk for habitat loss of high-altitude endemic species under climate change. Global Change Biol 17:990–996
Friedman JH (1991) Multivariate adaptive regression splines. Ann Stat 19:1–67
Gobiet A, Kotlarski S, Beniston M, Heinrich G, Rajczak J, Stoffel M (2014) 21st century climate change in the European Alps—a review. Sci Tot Environ 493:1138–1151
Grangé J-L (2008) Biologie de reproduction de la Niverolle Alpine Montifringilla nivalis dans le Pyrénées occidentales. Nos Oiseaux 55:67–82
Hosmer DW, Lemeshow S (1989) Applied logistic regression. Wiley, New York
Inouye DW, Barr B, Armitage KB, Inouye BD (2000) Climate change is affecting altitudinal migrants and declining species. PNAS 97:1630–1633
IPCC (2013) Climate change 2013: the physical science basis. In: Stocker TF et al (eds) Working group I contribution to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, pp 1–14
Jedlikowski J, Brambilla M, Suska-Malawska M (2014) Fine-scale selection of nesting habitat in Little Crake Porzana parva and Water Rail Rallus aquaticus in small ponds. Bird Study 61:171–181
Leathwick JR, Rowe D, Richardson J, Elith J, Hastie T (2005) Using multivariate adaptive regression splines to predict the distributions of New Zealand’s freshwater diadromous fish. Freshw Biol 50:2034–2052
Lu X, Ke DH, Zeng XH, Yu TL (2009) Reproductive ecology of two sympatric Tibetan snowfinch species at the edge of their altitudinal range: response to more stressful environments. J Arid Environ 73:1103–1108
Maggini R, Lehmann A, Zbinden N, Zimmermann NE, Bolliger J, Schröder B, Foppen R, Schmid H, Beniston M, Jenni L (2014) Assessing species vulnerability to climate and land use change: the case of the Swiss breeding birds. Divers Distrib 20:708–719
Milborrow S (2011a) Earth: multivariate adaptive regression spline models. R package version 4.4.3. http://cran.r-project.org/web/packages/earth. Accessed 1 Dec 2015
Milborrow S (2011b) Plotmo: plot a model’s response while varying the values of the predictors. R package 3.1.4. http://cran.r-project.org/web/packages/plotmo. Accessed 1 Dec 2015
Muscio G, Pellegrini GB, Solari M, Tomaselli M, Vanin S, Zanetti A (2005) Ambienti nivali. La vita in un ambiente estremo. Quaderni Habitat N. 10. Ministero dell’Ambiente e della Tutela del Territorio, Museo Friulano di Storia Naturale, Comune di Udine
R Development Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rosvold J (2016) Perennial ice and snow-covered land as important ecosystems for birds and mammals. J Biogeogr 43:3–12
Sekercioglu CH, Schneider SH, Fay JP, Loarie SR (2008) Climate change, elevational range shifts and bird extinctions. Conserv Biol 22:140–150
Strinella E, Ricci F, Vianale P (2007) Uso dell’habitat nel Fringuello alpino (Montifringilla nivalis) in periodo riproduttivo in un’area sub-antropizzata: Campo Imperatore (Gran Sasso-Abruzzo). Alula 14:107–114
Strinella E, Vianale P, Pirrello S, Artese C (2011) Biologia riproduttiva del Fringuello alpino Montifringilla nivalis a Campo Imperatore nel Parco Nazionale del Gran Sasso e Monti della Laga (AQ). Alula 18:95–100
Tarquini S, Vinci S, Favalli M, Doumaz F, Fornaciai A, Nannipieri L (2012) Release of a 10-m-resolution DEM for the Italian territory: comparison with global-coverage DEMs and anaglyph-mode exploration via the web. Comput Geosci 38:168–170
Vickery J, Arlettaz R (2012) The importance of habitat heterogeneity at multiple scales for birds in European agricultural landscapes. In: Fuller RJ (ed) Birds and habitat. Relationships in changing landscapes. Cambridge University Press, Cambridge
Acknowledgments
We are very grateful to E. Strinella, G. Volcan, E. Bassi, P. Trotti for helpful advice, and to two anonymous reviewers for very useful comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Gottschalk.
Rights and permissions
About this article

Cite this article
Brambilla, M., Cortesi, M., Capelli, F. et al. Foraging habitat selection by Alpine White-winged Snowfinches Montifringilla nivalis during the nestling rearing period. J Ornithol 158, 277–286 (2017). https://doi.org/10.1007/s10336-016-1392-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-016-1392-9