
Abstract
Arthropod hemocyanins (Hcs) are a family of large extracellular oxygen-transporting proteins with high molecular mass and hexameric or multi-hexameric molecular assembly. This study reports for the first time the isolation and characterization of the structure of an arthropod hemocyanin from crab Eriphia verrucosa (EvH) living in the Black Sea. Its oligomeric quaternary structure is based on different arrangements of a basic 6 × 75 kDa hexameric unit, and four of them (EvH1, EvH2, EvH3, and EvH4) were identified using ion-exchange chromatography. Subunit 3 (EvH3) shows high similarity scores (75.0, 87.5, 91.7, and 75.0 %, respectively) by comparison of the N-terminal sequence of subunit 1 from Cancer pagurus of the North Sea (Cp1), subunits 3 and 6 of Cancer magister (Cm3 and Cm6), and subunit 2 of Carcinus aestuarii (CaSS2), respectively. Moreover, a partial cDNA sequence (1309 bp) of E. verrucosa hemocyanin encoding a protein of 435 amino acids was isolated. The deduced amino acid sequence shows a high degree of similarity with subunits 3, 4, 5, and 6 of C. magister (81–84 %). Most of the hemocyanins are glycosylated, and three putative O-linkage sites were identified in the partial amino acid sequence of EvH at positions 444–446, 478–480, and 547–549, respectively. The higher stability of native Hc in comparison to its subunit EvH4 as determined by circular dichroism (CD) could be explained with the formation of a stabilizing quaternary structure.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Hemocyanins are copper-containing respiratory proteins, dissolved in concentrations up to 120 mg/ml in the hemolymph of molluscs and arthropods, including Crustacea, Chelicerata, Myriapoda, and some species of insects (Hagner-Holler et al. 2004; Sánchez et al. 1998; van Holde and Miller 1995; Chen et al. 2007; Mičetic et al. 2010). These molecules are composed of a number of subunits that assemble in an extremely large macro-molecular entity. These particles, which are similar in size to viruses or ribosomes, exhibit a complex allosteric behavior during oxygen binding.
Arthropod hemocyanins, hexamers, or oligo-hexameric assemblies with a molecular mass from 0.45 × 106 to 3.9 × 106 daltons are cubical molecules which in the electron microscope differ largely from cylindrical particles found in mollusks. Molluscan hemocyanins are decamers, didecamers, or multidecamers of polypeptides of Mr up to 450 kDa that carry seven or eight binuclear copper sites. In contrast, arthropod hemocyanins are hexamers (1 × 6) or oligohexamers (n × 6), and their aggregates are composed of 6 subunits (Herskovits 1988) with an Mr of 67–90 kDa, each containing only a single copper dioxygen-binding site (Herskovits 1988). Recently, the structures of several arthropod hemocyanins were very well described (Martin et al. 2007; Cong et al. 2009; Jaenicke et al. 2012; Kölsch et al. 2013).
Amino acid sequences of some isoforms of hemocyanins from several arthropod species have also been determined: subunits a, b, and c of Panulirus interruptus hemocyanin (Jekel et al. 1988; Neuteboom et al. 1992); d and e of Eurypelma californicum (Schartau et al. 1983), II of Limulus polyphemus (Martin et al. 2007); Androctonus australis (Buzy et al. 1995); Cancer magister (Carpenter and Van Holde 1973) or CaSS2 of Carcinus aestuarii hemocyanin (Dolashka-Angelova et al. 2005). Sequence similarity between these respiratory proteins was assessed by their comparisons. However, the primary structures of only few representatives of hemocyanins from the subphylum crustacea, namely Macrobrachium nipponense, Homarus americanus, P. interruptus, Palinurus vulgaris, C. magister, and CaSS2 of C. aestuarii, are known (Wang et al. 2012; Kusche and Burmester 2001). Structural comparison of crustacean hemocyanin polypeptide chains by sequence alignments (Burmester 2004; Marxen et al. 2013) and immunochemistry (Markl et al. 1984) showed a discrimination between different subunit types within infraorders. High homology was found between hemocyanins from infraorder Brachyura (Cancer magister, Callinectes sapidus, Eriocheir sinensis). Eriphia verrucosa, commonly known as warty crab or yellow crab, is a species of crab infraorder Brachyura living in the Black Sea, Mediterranean Sea, and Eastern Atlantic Ocean, (Di Giamberardino 1967). Until now, there is no information about the proteins in the hemolymph of crab E. verrucosa. Therefore, the structure of the hemocyanin from the crab E. verrucosa, living in the Black Sea, is determined for the first time and compared with those of other arthropod hemocyanins.
Materials and Methods
Animals and Sample Preparation
Adult edible female E. verrucosa crabs were obtained from the Black Sea, Bulgaria, and maintained in a recirculating sea water system at ambient light and temperature. E. verrucosa hemolymph was produced using a fine gauge needle, taking care not to dislodge the fat layer on the surface and homogenized in 50 mM bicarbonate buffer (pH 7.5) in presence of a protease inhibitor cocktail (Sigma) used at tenfold of the recommended concentration. The homogenate was centrifuged (15,000g, 4 °C, 30 min), the protein-containing supernatant carefully removed, placed into a clean tube, and re-centrifuged as described above. The protein content of the supernatant was determined by Bradford assay (Bio-Rad).
Isolation and Purification
For dissociation of the aggregates, hemocyanins were dialyzed versus 50 mM bicarbonate buffer, pH 9.2, containing 2 M urea, for 24 h. The mixture was chromatographed on a Mono Q HR 16/10 column (GE Healthcare Europe GmbH), equilibrated with the same buffer, and the subunits were eluted using the same buffer and a gradient from 0.0–1.0 M NaCl. The subunits were further purified by HPLC on a Nucleosil 7C18 column (250 × 10 mm; Machery–Nagel, Duren, Germany) using the following conditions: eluent A, 0.1 % trifluoroacetic acid; eluent B, 80 % acetonitrile in A; gradient program: 15 % B for 5 min followed by 15–100 % B in 55 min at a flow rate of 1 ml/min. A total of 10 % SDS-polyacrylamide gel electrophoresis was carried out to analyze the purity of the subunits and their molecular masses.
Amino Acid Sequence Determinations
Automated Edman degradation was performed using N-terminal sequence analysis (Procise 494A Pulsed Liquid Protein Sequencer, Applied Biosystems GmbH, Weiterstadt, Germany). Approximately 50–200 pmol of protein was applied on the cartridge filter, previously treated with polybrene. Alignments of the obtained N-terminal amino acid sequences of EvH subunits were created by the LALING program.
MALDI-TOF Mass Spectrometry
MALDI analyses were performed using a MALDI-TOF Ultraflex II instrument (Bruker Daltonics, Bremen, Germany) operating in linear positive ion mode. Ions were formed by a pulsed UV laser beam (nitrogen laser λ = 337 nm). Five microliters of each sample (structural subunits) were mixed with 5 μl of sinapinic acid matrix solution (saturated solution in H2O/acetonitrile (50/50; v/v) containing 0.1 % trifluoracetic acid). About 1 μl of this mixture was deposited on the stainless steel sample holder. External mass calibrations were done using the Protein Calibration Standard II programme (Bruker Daltonics), based on the average values of [M + H]+ of trypsinogen, protein A, bovine albumin (BSA), and the average values of [M + 2H]2+ of the same proteins at mass/charge (m/z) 23982, 44613, 66500, 22307, and 33300, respectively.
Carbohydrate Analysis
Isolated structural subunits were analyzed by orcinol–sulfuric test, where 2–4 μl of the purified sample solutions were applied to the thin layer plate and air-dried, taking care to restrict the size of the spots to 2–3 mm in diameter. The plate was sprayed with orcinol/H2SO4 and heated for 20 min at 100 °C.
Isolation of Total RNA from E. verrucosa Hepatopancreas
Fresh samples from adult Eriphia verrucosa hepatopancreas tissue were fixed in a stabilization buffer (0.02 M EDTA, 0.025 M sodium citrate, 700 mg ammonium sulfate, pH 5.2) and frozen at −80 °C. Total RNA was isolated with TriFast reagent (Fermentas) following the protocol of Chomczynski and Mackey (1995). Quality and concentration of the total RNA were determined by 1.2 % formaldehyde gel electrophoresis and a NanoDrop spectrophotometer, respectively. Total RNA isolation was followed by Dnase I digestion (Fermentas).
Reverse Transcription and Polymerase Chain Reaction Amplification of Hemocyanin Coding Sequence
Three-microgram total RNA, 1-μl oligo dT primer or degenerative primer F1 (TCT GCA GGA GCG TTC GAC GCG CA, Fig. 1), and nuclease-free water were mixed up to 12 μl and incubated at 65 °C for 5 min. The following components were added: 1 × 5 reaction buffer (4 μl), Ribolock RNase Inhibitor (20 u/μl), 10 mM dNTP Mix (2 μl), revertAid H minus M-MuLV reverse transcriptase (200 u/μl, 1 μl). The mixture was incubated at 60 °C for 1 h and terminated at 70 °C for 5 min (RevertAid™ H Minus First Strand cDNA Synthesis Kit, Fermentas). cDNA was diluted to 1/10, and 5 μl was subjected to PCR. The following degenerate primers were used to amplify the region encompassing a part of CuA and the complete CuB of arthropod hemocyanin site-degenerate sense “CuAI primer” corresponding to the conserved sequence within the copper A site (5′ GAACTTTTTTTTTGGGTTCATCATCAACTTAC 3′) (Durstewitz and Terwilliger 1997) and a degenerate CuBR4 primer located in the CuB site as antisense (5′ TGT GTT CTC TGA AGA TGT TAT CCA TGT ACT T 3′).

Fractionation of 10 mg of E. verrucosa hemocyanin, chromatographed on a Mono Q HR 16/10 column (GE Healthcare Europe GmbH), equilibrated with 50 mM bicarbonate buffer, pH 9.2, containing 2 M urea. The subunits were eluted using the same buffer and a gradient from 0.0 to 1.0 M NaCl at a flow rate of 1 ml/min. Insert shows electrophoretic patterns of isolated fractions on a 10 % SDS gel (fraction/ peaks 1, 2, 3, and 4)
PCR reactions were conducted in 50 μl containing 1× PCR buffer (Long PCR Enzyme Mix, Ferments), 4 mM MgCl2, 10 pmol primers, 0.2 mМ dNTPs, and 2.5 units (Long PCR Enzyme Mix) on a Verity TM 9902 96-well -termed cycler under the following conditions: Denaturation at 94 °C, 2 min; 35 cycles, each including denaturation at 94 °C, 40 s; annealing at 55 °C, 40 s, and extension at 68 °C, 2 min. The final extension step was at 68 °C for 5 min.
3′RACE
The 3′ end of the sequence coding for E. verrucosa hemocyanin was obtained using the 5′/3′ RACE Kit, 2nd Generation, Roche. Two-microgram total RNA from hepatopancreas was reverse transcribed, the obtained cDNA amplified with PCR anchor primer and the degenerate sense “CuAI primer” (Durstewitz and Terwilliger 1997).
The PCR reaction was carried out with 1 μl cDNA, 12,5 μM PCR anchor primer, 12.5 μM “CuAI primer,” 0.05 mM dNTP mix, 3.5 U high fidelity taq polymerase (Roche), 5.0-μl reaction buffer with 4 mM MgCl2. Ten microliters of PCR product were reamplified using 12.5 μM nested CuB 3 primer (5′ AAGTACATGGATAACATCTTCAGA 3′) and 12.5 μM anchored primer. PCR conditions were the same as described above.
Cloning, Sequencing, and Bioinformatic Analysis
PCR products were subjected to electrophoresis on 1 % agarose gel. Clear bands were cut and purified with the Illustra GFX™ PCR DNA and gel band purification kit, GE Healthcare, and then ligated to TA cloning vector pTZ57R/T (InsTAclone PCR Cloning Kit, Fermentas). The ligation product was used to transform E. coli DH5α. Universal primers M13 were used for sequencing. Sequencing was conducted on an ABI 337 sequencer. Up to 5 to 8 independent clones were sequenced to verify the authenticity. Sequencing data were analyzed by Vector NTI software.
Nucleotide and deduced amino acid sequences were compared with NCBI BLAST search programs. Additional sequences for phylogenetic analyses were retrieved from GenBank and aligned with Clustal W. The alignments of the conserved copper A- and copper B-binding site histidines were verified.
CD Measurements
Circular dichroism (CD) spectra were recorded on a J-720 dichrograph (Jasco, Tokyo, Japan). Round quartz cells with a path length of 10 mm were used in all experiments. CD spectra were recorded in the range between 200 and 250 nm with a band width of 1 nm, a scan speed of 50 nm/min, a time constant of 8.0 s, and accumulation −4. The hemocyanin solutions with A280 = 0.215 were prepared in 10 mM Tris buffer (pH 7.4). The pH stability of the native molecule and structural subunit EvH4 was analyzed using buffers with pH values of the following: 11.5, 11.0, 10.0, 9.50, 9.00, 8.50, 5.72, 5.50, 5.20, 5.00, 4.50, 3.80, 3.50, and 3.00. The cocktail buffer at ionic strength 0.075 was adjusted to the given pH in the range pH 2–12: sodium citrate (pH 3.0–5.0); sodium phosphate (pH 5.0–7.0); Tris/HCl (pH 7.0–9.0); carbonate/bicarbonate (pH 9.0–10.5), and 5 M NaOH (pH 11–12).
Protein solutions were prepared in 20 mM cocktail buffer (sodium phosphate buffer, pH 7.4–5.8, sodium acetate buffer, pH 5.4–3.6, and glycine–HCl buffer, pH 3.4–1.2 in equal volumes). The final mixtures with protein concentrations of 20 μM were incubated for 20 min at room temperature before optical measurements. Each spectrum was the average of four scans, and the results are expressed as mean residue ellipticity (MRE, [Θ]222) in deg cm2 dmol−1.
Results and Discussion
Isolation of Eriphia verrucosa Hemocyanin and its Isoforms
The crab E. verrucosa lives in the Black Sea, Mediterranean Sea, and Eastern Atlantic Ocean at different environments and conditions. The hemolymph was obtained from the yellow crab E. verrucosa, collected from the Black Sea, and after centrifugation at 15,000g, 4 °C for 30 min, the native hemocyanin was purified by gel filtration chromatography on a Superdex column. One main peak was eluted from the column by 50 mM bicarbonate buffer (pH 7.5), and after additional purification of 3 mg of this fraction by HPLC on a Nucleosil 7C18 column, its N-terminal sequence was determined. Determination of 16 amino acids (ADSADVALAQKQHDVN-) from the N-terminus by Edman degradation confirmed the E. verrucosa hemocyanin (EvH) aggregates for the purified fractions.
After dissociation of the obtained E. verrucosa hemocyanin by dialysis versus 50 mM bicarbonate buffer, pH 9.2 and 2 M urea for 24 h, of the obtained E. verrucosa hemocyanin on a Superdex column, five fractions were eluted on a Mono Q HR column with a linear gradient from 0.1–1.0 M NaCI in 50 mM bicarbonate buffer, pH 9.6 (Fig. 1). The obtained fractions were desalted by gel filtration through a Sephadex G-25 (Pharmacia) column, and electrophoretic analysis confirmed a high-grade separation of four structural subunits (fractions 1–4) with molecular masses of 70–75 kDa (Fig. 1, insert). From the eluted fractions, number 5 is a non-dissociated hemocyanin; therefore, only fractions 1–4 were additionally analyzed. The subunits and molecular weight markers were applied to gel-exclusion chromatography (2 × 150-cm Superdex™ 200 column) and eluted with 0.1 M Tris–HCl, pH 8.2, 5 mM CaCl2, 5 mM MgCl2 at a flow rate of 9 ml/h. The weight-average molecular masses of the subunits were determined to be in the range of about 75 kDa, according to the elution profile of the markers.
To determine more precise the molecular masses of the isolated subunits from E. verrucosa and their N-terminal sequences, more precisely fractions 1, 2, 3, and 4 were additionally purified on a Nucleosil 100 RP-18 column, where only one main peak was eluted from the column (Fig. 2). Then, the masses of the polypeptides in each fraction were determined with more precision by MALDI-TOF. Figure 3 shows the MALDI spectrum of fraction 1 (Fig. 1) for which a molecular mass of 75 098.62 Da was determined (Fig. 3). The masses of the other 3 subunits were found to be in the same region 74,488 Da (EvH2), 75,753 Da (EvH3), and 74,713 Da (EvH4), correlating with the masses from other arthropod hemocyanins (Burmester 1996).

HPLC purification of structural subunits on a Nucleosil 7 C18 column (250 × 10 mm; Machery–Nagel, Düren, Germany) using the following conditions: eluent A, 0.1 % trifluoroacetic acid; eluent B, 80 % acetonitrile in A; gradient program, 15 % B for 5, followed by 15–100 % B in 55 min at a flow rate of 1 ml/min

MALDI spectrum of structural subunit 1 from E. verrucosa Hc isolated on a Mono Q HR 16/10 column, followed by additional purification on a Nucleosil RP C18 column (Fig. 2). The sample was measured by MALDI-TOF Ultraflex II (Bruker Daltonics, Bremen, Germany)
The N-terminal sequences of the purified subunits were determined by Edman degradation, and the obtained results are compared in Fig. 4 with those of other arthropod hemocyanin subunits. A high similarity score was found between N-terminal sequences of hemocyanin subunits from Eriphia verrucosa (EvH1, EvH3) and polypeptide chains from hemocyanins found in other species. Comparison of the N-terminal sequences of EvH1, EvH2, EvH3, and EvH4, with those of C. magister and C. aestuarii of infraorder Brachyura, shows conserved Asp and Ser residues at positions 2 and 3, respectively. Gly5 and Gly6 are also conserved in EvH3, Cp1, Cm3, Cm6, and CaSS2. Predominant Ala, Gln, Lys, Gln, as well as Asp and Ser residues were observed in most of the subunits in positions 10, 11, 12, 13, 15, and 18. Within the infraorder Brachyura, the positions 16 (Val) and 17 (Asp) are conserved with Val and Asp residues, respectively.

The relationship between N-terminal sequences of structural subunits originating from E. verrucosa (Black Sea; EvH1-4), Cancer magister (Cm 1, 2, 3, 4, and 6), and Carcinus aestuarii (CaSS2)
High similarity scores were found comparing the N-terminal sequences of subunit 3 from E. verrucosa (EvH3) with those of C. pagurus (Cp1), C. magister (Cm3 and Cm6), and C. aestuarii (CaSS2). Therefore, these five subunits were considered to be closely related.
Isolation of a Partial Hemocyanin cDNA Sequence (EvH)
Hemocyanin sequences were initially amplified with degenerate crustacean hemocyanin primers based on the conserved regions of copper A and copper B binding sites. Amplification resulted in a 540-bp product that was cloned and sequenced. The sequence was used to design additional primers to further amplify hemocyanin following the 3′ RACE protocol (ROCHE). Using PCR with anchor primer (ROCHE), “CuAI primer” and the nested CuBF3 specific primer (Fig. 5a), a 1.022-bp cDNA fragment was amplified, cloned, and sequenced. The identical overlapping (35 bases) of both fragments, 540 bp and 1.022 bp, allowed their correct assembling. The obtained contig (1523 bp) covering about 2/3 of the putative Hc gene contains ORF of a 1309 bp, 3′UTR region (201 bp), and polyA tail (13 bp) (data not shown).

a Schematic presentation of primer positions: PCR anchor primer (ROCHE), “CuAI primer”—specific for the CuA binding site region and CuBF3-nested primer; b alignment between partial hemocyanin amino acid sequences, subunits from E. verrucosa (EvH), C. magister CmH 3 (AAW57891.1), 4 (AAW57892.1), 5 (AAW57893.1), 6 (AAA96966.2), Callinectes sapidus (AAF64305.1), and C. aestuarii (CaSS2; P84293.1). Borders of the helics and CuA and CuB regions are underlined. Cubinding histidines are shown in gray color; the positions of N-glycosylated sites are given in boxes while the putative O- glycosylated sites are bolded
The translated amino acid sequence (435 AA in length) showed 70; 83; 84, and 81 % identity with the C. magister hemocyanin subunits 3 (AAW57891.1), 4 (AAW57892.1), 5 (AAW57893.1), and 6 (AAA96966.2), respectively (Fig. 5b). High identity (65 %) was also observed between EvH and CaeSS2 of C. aestuarii (P 84293.1), Callinectes sapidus (AF249297.1) (78 %), and Eriocheir sinensis (AEG64817.1) (74 %).
These values are very close to the homology reported between the subunits of other hemocyanins, and we may suggest that this fragment belongs to EvH4 or EvH5.
There is a vast amount of date supporting the hypothesis that arthropod hemocyanins consist of three domains (Hazes et al. 1993; Decker and Jaenicke 2004; Terwilliger et al. 2006). Crystallographic studies of hemocyanin from Panulirus and Limulus (Volbeda and Hol 1989; Hazes et al. 1993) have shown that arthropod hemocyanins consist of three structural domains. The region between residues 220 and 394 in domain 2 is the most conserved and contains the oxygen-binding CuA and CuB sites. Each copper ion is ligated to three histidine side chains. The six histidines marked by gray color in Fig. 5b are implicated in Cu binding according to their highly conserved character among arthropod hemocyanin subunits (Durstewitz and Terwilliger 1997; Dolashka-Angelova et al. 2005). The Cu binding histidines are located at positions 196, 220, 253, 373, 377, 413, and 423 in EvH and all hemocyanins, studied here (Fig. 5b). The high conservation of the histidine ligands launch the hypothesis that the isolated cDNA is a gene encoding a protein that binds Cu. The CuA helix pair in E. verrucosa subunit extends from residue 214 to residue 230 and from residue 243 to residue 268 indicating that His253 of EvH, CaSS2, CmH5, and CmH6 is involved in oxygen-binding CuA. The histidine residues binding the copper ion at the CuB site are located at positions 373, 377, 413, and 423 (Fig. 5b).
Glycosylation of Structural Subunits of E. verrucosa Hemocyanin
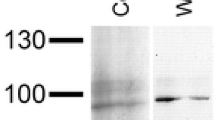
It is known that mollusk and arthropod Hcs are glycosylated, but the glycans represent only about 1–2 % of the protein masses of arthropod Hcs, e.g., for C. aestuarii hemocyanin (CaeSS2), 1.6 % could be determined (Dolashka-Angelova et al. 2001, 2005; Kostadinova et al. 2013). To identify the glycosylated subunits, the orcinol–sulfuric test was applied to four subunits EvH1-4 and native molecule of EvH and compared with the result on glycosylated subunit CaSS2 of C. aestuarii hemocyanin (Dolashka-Angelova et al. 2005). No glycans were identified in subunit 4, but the test demonstrates that subunits 1, 2, and 3 are glycosylated (Fig. 6). Moreover, the spots on positions 1 and 3 are less intense compared to the position 2 indicating a higher sugar content for subunit 2, which, however, is of lower intensity than for position 6, the spot for glycosylated CaSS2.

Orcinol–sulfuric acid test of (glyco) subunits eluted by HPLC and applied on to a silica-gel plate. The spots on positions: position 1—EvH1; position 2—EvH2; position 3—EvH3; position 4—EvH4; position 5—EvH; position 6—CaSS2
Three consensus sequences for O-glycosylation and one for N-glycosylation were identified in CaeSS2 after trypsin digestion, separation of glycopeptides, and amino acid sequencing (Dolashka-Angelova et al. 2001, 2005). One putative N-linked site was observed at positions 338–340 in CaeSS2 with a consensus sequence Asn-Gly-Ser, typical for N-glycosylation. However, as is shown in Fig. 5b, Asn residues are substituted by Asp at the same position in EvH and CmH 4, 5, and 6 confirming that these subunits can not be N-glycosylated. The absence of N-glycosylation sites (NX(T/S)) in the short part of the primary structure of EvH shows that this subunit is not glycosylated. The putative N-glycosylation sites marked by boxes are located at positions 630–632 in C. magister subunits 3, 4, 5, and 6 (Fig. 5b). One putative N-linkage site was also identified at position 166 in P. elephas α-type subunits 1, 2, and 3 (Meissner et al. 2003), and β-type subunit a, b at position 470 in P. interuptus. Four potential N-glycosylation sites are present in each of the H. americanus pseudohemocyanin (Burmester 1999). No carbohydrate moiety was detected in native E. californicum Hc, and its seven subunits which fits to the observation that chelicerate hemocyanins are synthesized in the cytosol and not by the rough ER as known for crustacean hemocyanins (Voit et al. 2000).
Instead of N-linkage sites, three putative O-linkage sites were identified in EvH which suppose glycosylation at positions 444–446, 478–480, and 547–549 (Fig. 5b). Several other O-glycosylation sites were published for CaSS2 at positions 73–75, 83–85, 198–200, 426–428 (Dolashka-Angelova et al. 2005), but their positions do not correlate with the putative glycosylated sites in EvH.
Stability of E. verrucosa Hemocyanin
There are several reports on the biophysical and structural properties of the function and stability of molluscan and arthropod Hcs (Ali et al. 2000; Coates et al. 2012; Coates and Nairn 2014; Spinozzi et al. 2005).
The conformational stability of the native aggregates of several arthropod Hcs and their isolated structural subunits toward various denaturants (pH and guanidine hydrochloride (Gdn·HCl)) indicate that hydrophilic and polar forces stabilize the quaternary structure (Hristova et al. 1997; Dolashka-Angelova et al. 1999, 2000). The reassociation behavior of artropodan hemocyanin Carcinus maenas was analyzed by ESI-MS (Bruneaux et al. 2009). By returning to a more neutral pH after dissociation of the native C. maenas, a complete reassociation was not observed.
Changes in the secondary structure of native EvH and two structural subunits (glycosylated EvH1 and nonglycosylated EvH4) at different pH values (3.0–11.5) were followed by near UV-CD measurements of [θ]222 in the region between 200 and 250 nm. The far-UV CD spectra of the native protein and two structural subunits in 50 mM Tris/HCl, pH 8.2, are dominated by a negative dichroic band with two minima at 210 and 220 nm (Fig. 7a, b).

a pH stability of structural subunit EvH4 analyzed by circular dichroism (CD). Spectra at pH values 5.72, 5.50, 5.20, 5.00, 4.50, 3.80, 3.50, and 3.00 and b at pH values 11.5, 11.0, 10.0, 9.50, 9.00, and pH 8.50. Spectra were recorded on a J-720 dichrograph (Jasco, Tokyo, Japan) in the range between 200 and 250 nm with a band width of 1 nm, a scan speed of 50 nm/min, a time constant of 8.0 s, and accumulation −4. The hemocyanin solutions with A280 = 0.215 were prepared in different buffers. c Influence of pH on [θ]222 of native CpH and two structural subunits nonglycosylated EvH4 and glycosylated EvH1 at different pH values (3.0–11.5)
Figure 7c shows that the [θ]222 vs pH plots represent denaturation studies of the native molecule and two structural subunits, EvH1 and EvH4, of E. verrucosa hemocyanin by pH.
The curve indicates that the denaturation process of the native molecule in the alkaline region represents dissociation of the native molecule into its structural subunits at pH values over 9.0 followed by their denaturation at higher pH values. However, it may be difficult to distinguish between the effects on the quaternary structure of the oligomeric protein (initial dissociation) and those on the tertiary and secondary structure (i.e., subunit unfolding).
Comparison of the pH dependence of the dichroic curves of the native molecule with those of two structural subunits ([θ]222 vs (pH)) at low and high pH values at 25 °C shows a similar behavior for all 3 proteins. The [θ]222/pH values are more or less independent from pH within the pH range 6.0–9.2 for native EvH, but for both subunits, the constant pH range is shortened to 7.5–9.0 (Fig. 7c). These data confirm that the native EvH is more stable to pH denaturation as compared to its structural subunits EvH1 and EvH4. No influence of glycosylation was observed in EvH1. In the alkaline part (pH 8–11.5), relative changes are too small and non-cooperative (within a wide pH interval), indicating that for alkaline denaturation of EvH, EvH4, and EvH1 as a reversible process cannot be achieved.
Figure 8a shows the CD spectra of the native molecule of EvH in 50 mM Tris–HCl, buffer, containing 5 mM CaCl2, 5 mM MgCl2, at pH 8.2, recorded in the temperature interval 20–90 °C. The temperature dependence of the ellipticity at 222 nm of the native molecule and the structural subunits EvH1 and EvH4 show that the thermal denaturation of the EvH is also in reversible process (Fig. 8b). The thermostability of EvH and its subunits were compared by their melting temperatures, Tm, the midpoint in the sigmoidal denaturation curves. The melting temperature for the native EvH (Tm = 72 °C) is larger compared to its structural subunits EvH1 (Tm = 60 °C) and EvH4 (Tm = 57 °C) and correlates with thermostability of arthropod hemocyanins C. aestuarii (Tm 72 °C), Callinectes sapidus (Tm∼76 °C), and Maia squinado (Tm∼72 °C) (Dolashka-Angelova et al. 1999, 2000). The larger Tm values found for native EvH compared to its subunits EvH1 and EvH4 is an excellent indication for a stabilization of the protein caused by oligomerization.

a Temperature stability of structural subunit EvH1 and EvH4 of E. verrucosa hemocyanin (A280 = 0.274) analyzed by circular dichroism (CD). Spectra were recorded at 20–90 °C in the range between 200 and 250 nm. b Temperature dependence of the ellipticity [θ]222 at 222 nm of native EvH in 50 mM Tris–HCl, buffer, containing 5 mM CaCl2, 5 mM MgCl2, at pH 8.2 and structural subunits EvH1 and EvH4
Conclusion
In this study, we report for the first time information about the hemocyanin of the marine crab Eriphia verrucosa living in the Black Sea. Hemocyanin was isolated and purified from the hemolymph, and four structural subunits (EvH1, EvH2, EvH3, and EvH4) with molecular masses in the range of least 75–76 kDa were identified. Its isolated partial hemocyanin cDNA (1523 bp) corresponds to the region encompassing part of the conservative CuA, the full CuB binding sites, and the 3′ end of the gene. The alignment shows a high degree of sequence similarity of EvH with those of C. magister, C. aestuarii, C. sapidus, and E. sinensis hemocyanins which suggests a common structure. Three of the EvH subunits (EvH1, EvH2, and EvH3) are glycosylated as confrmed by the orcinol–sulfuric test. Several putative O-linkage sites were found in the identified amino acid sequence of EvH.
In this paper, we present communication for the first time some preliminary results on the pH- and thermostability of the native protein and its structural subunits EvH1 and EvH4, as characterized by CD measurements. We report on the pH values under which reversible structural changes of EvH, EvH1, and EvH4 can exist and strong alkaline as well as acid environment causes changes in quaternary and secondary structure.
References
Ali S, Abbasi A, Stoeva S, Kayed R, Dolashka-Angelova P, Schwarz H, Voelter W (2000) Oxygen transport proteins: III. Structural studies of the scorpion Buthus sindicus hemocyanin, partial primary structure of its subunit Bsin1. Comp Biochem Physiol B 126:361–376
Bruneaux M, Terrier P, Leize E, Mary J, Lallier FH, Zal F (2009) Structural study of Carcinus maenas hemocyanin by native ESI-MS: interaction with L-lactate and divalent cations. Proteins 77(3):589–601
Burmester T (1996) Molecular evolution of the arthropod hemocyanin superfamily. Mol Biol 18:184–195
Burmester T (1999) Identification, molecular cloning and phylogenetic analysis of a nonrespiratory pseudo-hemocyanin of Homarus americanus. J Biol Chem 274:13217–13222
Burmester T (2004) Evolutionary history and diversity of arthropod hemocyanins. Micron 35:121–122
Buzy A, Gagnon J, Lamy J, Thibault P, Forest E, Hudry-Clergeon G (1995) Complete amino acid sequence of the Aa6 subunit of the scorpion Androctonus australis hemocyanin determined by Edman degradation and mass spectrometry. Eur J Biochem 233:93–101
Carpenter D, van Holde K (1973) Amino acid composition, amino-terminal analysis, and subunit structure of Cancer magister hemocyanin. Biochemistry 12:2231–2238
Chen HY, Ho SH, Chen TI, Soong K, Chen IM, Cheng JH (2007) Identification of a female-specific hemocyanin in the mud crab, Scylla olivacea (Crustacea: Portunidae). Zool Stud 46:194–202
Chomczynski P, Mackey K (1995) Substitution of chloroform by bromochloropropane in the single-step method of RNA isolation. Anal Biochem 225:163–164
Coаtes J, Nairn J (2014) Diverse immune functions of hemocyanins. Dev Comp Immunol 45:43–55
Coаtes CJ, Whalley T, Nairn J (2012) Phagocytic activity of Limulus polyphemus amebocytes in vitro. J Invertebr Pathol 111(3):205–210
Cong Y, Zhang Q, Woolford D, Schweikardt T, Khant H, Dougherty M, Ludtke SJ, Chiu W, Decker H (2009) Structural mechanism of SDS-induced enzyme activity of scorpion hemocyaninrevealed by electron cryomicroscopy. Structure 17:749–758
Decker H, Jaenicke E (2004) Recent findings on phenoloxidase activity and antimicrobial activity of hemocyanins. Dev Comp Immunol 28:673–687
Di Giamberardino L (1967) Dissociation of Eriphia hemocyanin. Arch Biochem Biophys 118:273–278
Dolashka-Angelova P, Hristova R, Stoeva S, Voelter W (1999) Spectroscopic properties of Carcinus aestuarii hemocyanin and its structural subunits. Spectrochim Acta Part A 55:2927–2934
Dolashka-Angelova P, Stoeva S, Hristova R, Schuetz J, Voelter W (2000) Structural and spectroscopic studies of the native hemocyanin from Maia squinado and its structural subunits. Spectrochim Acta A 56:1985–1999
Dolashka-Angelova P, Beltramini M, Dolashki A, Salvato B, Voelter V (2001) Carbohydrate composition of Carcinus aestuarii hemocyanin. Arch Biochem Biophys 389:153–158
Dolashka-Angelova P, Dolashki A, Savvides SN, Hristova R, Van Beeumen J, Voelter W, Devreese B, Weser U, Di Muro P, Salvato B, Stevanovic S (2005) Structure of hemocyanin subunit CaeSS2 of the crustacean Mediterranean crab Carcinus aestuarii. J Biochem 138:303–312
Durstewitz G, Terwilliger NB (1997) cDNA cloning of a developmentally regulated hemocyanin in the crustacean Cancer magister and phylogenetic analysis of the hemocyanin gene family. Mol Biol Evol 14:266–276
Hagner-Holler S, Schoen A, Erker W, Marden JH, Rupprecht R, Decker H, Burmester T (2004) A respiratory hemocyanin from an insect. Proc Natl Acad Sci 101:871–874
Hazes B, Magnus KA, Bonaventura C, Bonaventura J, Dauter Z, Kalk KH, Holx WG (1993) Crystal structure of deoxygenated Limulus polyphemus subunit II hemocyanin at 2.18 A resolution: clues for a mechanism for allosteric regulation. Protein Sci 2:597–619
Herskovits TT (1988) Recent aspects of the subunit organization and dissociation of hemocyanins. Comp Biochem Physiol B 91:597–611
Hristova R, Dolashka P, Stoeva S, Voelter W, Salvato B, Genov N (1997) Spectroscopic properties and stability of hemocyanins. Spectrochim Acta A 53:471–478
Jaenicke E, Pairet B, Hartmann H, Decker H (2012) Crystallization and preliminary analysis of crystals of the 24-meric hemocyanin of the emperor scorpion (Pandinus imperator). PLoS ONE. doi:10.1371/journal.pone. 0032548
Jekel PA, Bak HJ, Soeter NM, Vereijken JM, Beintema JJ (1988) Panulirus interruptus hemocyanin. The amino acid sequence of subunit b and anomalous behaviour of subunits a and b on polyacrylamide gel electrophoresis in the presence of SDS. Eur J Biochem 178:403–412
Kölsch A, Hörnemann J, Wengenroth C, Hellmann N (2013) Differential regulation of hexameric and dodecameric hemocyanin from A. leptodactylus. Biochim Biophys Acta 1834:1853–1859
Kostadinova E, Dolashka P, Velkova L, Dolashki A, Stevanovic S, Voelter W (2013) Positions of the glycans in molluscan hemocyanin, determined by fluorescence spectroscopy. J Fluoresc 23:753–760
Kusche K, Burmester T (2001) Molecular cloning and evolution of lobster hemocyanin. Biochem Biophys Res Commun 282:887–892
Markl J, Gebauer W, Runzler R, Avissar I (1984) Immunological correspondence between arthropod hemocyanin subunits. I. Scorpion (Leiurus, Androctonus) and spider (Eurypelma, Cupiennius) hemocyanin. Hoppe Seylers Z Physiol Chem 365:619–631
Martin A, Depoix F, Stohr M, Meissner U, Hagner-Holler S, Hammouti K (2007) Limulus polyphemus hemocyanin: 10 Å structure, sequence analysis, molecular modelling and rigid-body fitting reveal the interfaces between the eight hexamers. J Mol Biol 366:1332–1350
Marxen JC, Pick C, Kwiatkowski M, Burmester T (2013) Molecular characterization and evolution of haemocyanin from the two freshwater shrimps Caridina multidentata (Stimpson, 1860) and Atyopsis moluccensis (De Haan, 1849). J Comp Physiol B 183:613–624
Meissner U, Stohr M, Kusche K, Burmester T, Stark H, Orlova EV, Markl J (2003) Quaternary structure of the European spiny lobster Palinurus elephas 1x6-mer hemocyanin from cryoEM and amino acid sequence data. J Mol Biol 325:99–109
Mičetic I, Losasso C, Di Muro P, Tognon G, Benedetti P, Beltramini M (2010) Solution structures of 2x6-meric and 4x6-meric hemocyanins of crustaceans Carcinus aestuarii, Squilla mantis and Upogebia pusilla. J Struct Biol 171:1–10
Neuteboom B, Jekel PA, Beintema JJ (1992) Primary structure of hemocyanin subunit c from Panulirus interruptus. Eur J Biochem 206:243–249
Sánchez D, Ganfornina MD, Gutierrez G, Bastani MJ (1998) Molecular characterization and phylogenetic relationship of a protein with potential oxygen-binding capabilities in the grasshopper embryo. A hemocyanin in insects? Mol Biol Evol 15:415–426
Schartau W, Eyerle F, Reisinger P, Geisert H, Storz H, Linzen B (1983) Hemocyanins in spiders, XIX. Complete amino-acid sequence of subunit d from Eurypelma californicum hemocyanin, and comparison to chain e. Hoppe Seyler Z Physiol Chem 364:1383–1409
Spinozzi F, Gatto S, De Filippis V, Carsughi F, Di Muro P, Beltramini M (2005) Contribution of the copper ions in the dinuclear active site to the stability of Carcinus aestuarii hemocyanin. Arch Biochem Biophys 439:42–52
Terwilliger NB, Ryan M, Phillips MR (2006) Crustacean hemocyanin gene family and microarray studies of expression change during eco-physiological stress. Integr Comp Biol 46:991–999
Van Holde KE, Miller KI (1995) Hemocyanins. Adv Protein Chem 47:1–81
Voit R, Feldmaier-Fuchs G, Schweikardt T, Decker H, Burmester T (2000) Complete sequence of the 24-mer hemocyanin of the tarantula Eurypelma californicum. Structure and intramolecular evolution of the subunits. J Biol Chem 275:39339–39344
Volbeda A, Hol WG (1989) Crystal structure of hexameric haemocyanin from Panulirus interruptus refined at 3.2 A resolution. J Mol Biol 209:249–279
Wang W, Xia X, Liu F, Chen X, Yang H, Ning Q (2012) Cloning and characterization of the hemocyanin gene of prawn Macrobrachium nipponense. Turk J Biochem 37:348–355
Acknowledgments
This work was supported by Bulgarian Ministry of Education, projects DHRC /01/6 and “Young researchers” DMU 03/26, Deutsche Forschungsgemeinstchaft (DFG-STE 1819/5-1/2012),Germany, grant no. BG051PO001-3.3.06-0025, financed by the European Social Fund and Operational Programme Human Resources Development (2007–2013) and co-financed by Bulgarian Ministry of Education, Youth and Science.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Dolashki, A., Radkova, M., Todorovska, E. et al. Structure and Characterization of Eriphia verrucosa Hemocyanin. Mar Biotechnol 17, 743–752 (2015). https://doi.org/10.1007/s10126-015-9653-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10126-015-9653-9