
Abstract
Agricultural food production and sustainability need intensification to address the current global food supply to meet human demand. The continuous human population increase and other anthropogenic activities threaten food security. Agrochemical inputs have long been used in conventional agricultural systems to boost crop productivity, but they are disadvantageous to a safe environment. Towards developing environmentally friendly agriculture, efforts are being directed in exploring biological resources from soil and plant microbes. The survival of the rhizosphere and endosphere microbiota is influenced by biotic and abiotic factors. Plant microbiota live interdependently with the host plants. Endophytes are regarded as colonizer microbes inhabiting and establishing microbial communities within the plant tissue. Their activities are varied and include fixing atmospheric nitrogen, solubilizing phosphate, synthesis of siderophores, secretion of metabolite-like compounds containing active biocontrol agents in the control of phytopathogens, and induced systemic resistance that stimulates plant response to withstand stress. Exploring beneficial endophyte resources in the formulation of bio-inoculants, such as biofertilizers, as an alternative to agrochemicals (fertilizers and pesticides) in developing environmentally friendly agriculture and for incorporation into crop breeding and disease control program is promising. Therefore, in this review, endosphere microbial ecology, associating environmental factors, and their roles that contribute to their effectiveness in promoting plant growth for maximum agricultural crop productivity were highlighted.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Agriculture for food production needs to be intensified to meet the current trend in the world population. The world aims at ensuring food stability, but due to unpredictable climatic changes, there is a need to find an alternative and fast approach to ensure food sustainability (Aloo et al. 2019). Application of chemical fertilizers and pesticides has caused an improvement in agricultural productivity, thus assuring food security, and agriculture for food production needs intensification through adequate and proper use of agro-inputs (Wolde-Meskel et al. 2018). The use of agrochemicals is characterized by high cost, environmental pollution, toxicity to soil microorganisms, and disease in humans (Campos et al. 2019; Sandargo et al. 2019). Similarly, the use of the agrochemical is surrounded with a lot of disadvantages and detrimental influences on the functional ecosystem where living organisms dwell, and their prolonged use reduces crop quality and quantity, soil fertility, causing eutrophication in the water bodies, increase soil acidity, thus, affecting the microbial composition and their resistance which makes them persist in the environment (Segaran and Sathiavelu 2019). Although the use of agro-allied farm inputs, such as synthetic fertilizers and pesticides by farmers, is aimed at improving crop yields, boosting agricultural production, and averting hunger and famine among the growing human population in the world, a lot of environmental problems are associated with their use, due to poor management and improper use, which sometimes escalate to grievous environmental problems (Sharma and Singhvi 2017). In 1992, the World Health Organization (WHO) had reported over 3 million pesticide poisoning affecting individuals with 220,000 deaths.
There are many facets of agriculture, but it is mainly to supply food to the growing population and abundant raw materials to industries as a source of income and economic growth. Nevertheless, its inefficient and inadequate practices have greatly reduced soil fertility. There is a need to design the best option for boosting agricultural production using affordable and scalable farming approaches through technological progress by exploring veritable microbial resources for maximum crop productivity in a sustainable manner (Igiehon and Babalola 2017). In present-time agricultural practice, plant breeding and technological advancement, as well as the application of agrochemicals, such as pesticides and fertilizers, have progressively improved crop yield, though with major concerns threatening the ecosystem and environment. The use of agrochemicals has contributed greatly to global warming which has resulted in the depletion in oxygenation in the aquatic habitat (Zhang et al. 2018b).
Crop yield and agricultural produce are often influenced by environmental factors causing global warming through changes in the temperatures, rainfall, anthropogenic, and other human activities. Global warming is a phenomenon challenging agricultural production and increasing the risk of food insecurity, damage to crop plants, shortage, and scarcity of food for humans, which can result to hunger, and even death. Other anthropogenic and occupational activities, such as urbanization, have stemmed from the emission of greenhouse gasses as a result of deforestation and poor land use; changes in agriculture, forestry, and land-use contribute to about 25% of global emissions (Paul et al. 2019). The effects of global warming and greenhouse gas emission can be ameliorated by setting agricultural policies that would encourage the use of biological-based (biofertilizers or biopesticides) or non-toxic organic material (compost) in place of agrochemicals (fertilizers and pesticides), as biofertilizers and biopesticides are easily formulated from environmental microbes isolated from plants and soil (Bargaz et al. 2018).
Globally, the problems facing agricultural production vary per region. Poor soil fertility, erratic rainfall, and changes in climatic conditions affect the growth of plants with many effects on crop yield. The influence of environmental factors on living organisms, most notably the minute life, affects the nutrient mobilization and assimilation by plants in developing sustainable agriculture and improving crop productivity (Omomowo and Babalola 2019). The reduction in agricultural crop output is usually due to continuous cropping or over-use of cultivated land that results in nutrient loss, land degradation, the long term use, sole reliance on chemical fertilizers and pesticides, pest and rodent attacks and urbanization involving clearing and removal of the topsoil where nutrients are deposited (Doni et al. 2019; Omomowo and Babalola 2019).
Proper agricultural management and practices are the best way to ensure agrarian sustainability by application of farm inputs that promote soil health and retain nutrients in the soil. Human beings tend to amend the soil by application of synthetic agrochemicals and other agro-allies instead of using biologically synthesized products from soil-inhabiting organisms (Mahanty et al. 2017). This could be due to a lack of awareness and knowledge that a populous unseen microbial world exists naturally in a different environment with a range of activities in promoting plant growth, soil health, root elongation, seed germination, crop yields, drought control, formulation of biofertilizers, biopesticides, etc. (Igiehon and Babalola 2017). Poor land-use by continual application of chemical fertilizers and pesticides reduces soil nutrients, alters soil physicochemical characteristics, and makes plants vulnerable to pests and diseases, thus affecting different microbiota communities in the soil. Since adequate nutrient supply to soil boosts the growth of plants and soil health under proper environmental, proper soil management by conserving nutrients in such a way to enhance soil microbiota would better enhance crop yields and productivity (Odelade and Babalola 2019). The activities of microorganisms found in the soil not only limit pests and diseases, but facilitate catalysis, the breakdown of complex organic matter, and removal of toxic heavy metal pollutants, as well as improving soil fertility and soil health (Salam and Varma 2019).
Beneficial plant-microbe interactions are usually symbiotic, as the beneficial microbes confer many benefits on the plant. The internal tissues or organs of plants harboring a community of microorganisms is known as endophytes, which directly affect the physiological features of plants due to their interaction, without developing any disease symptoms or damaging the host immune responses (Aloo et al. 2019). The peculiar niches occupied by endophytes in a habitat differentiating them from the rhizosphere might likely cause a change in their functions, adaptations, responses, competence, and specialization. Even in the endosphere, not every part is colonized by the same microbes; some are specialized as per the region or location they colonized. The colonizer and survival endophytes type are commonly found colonizing plant reproduction organ (ovules) while the seeds endophytes colonizer are characterized to be competitors which make them adapt to the harsh environment (Sahu et al. 2019). The competence of bacterial endophytes and their growth-promoting efficacy in the seed of rice has been reported (Banik et al. 2019; Zhou et al. 2020).
Despite the variance in the microbial population in ecological niches, the free-living microbes found in the soil and root interface, i.e., rhizosphere, such as PGP rhizobacteria, and the endophytes community could share similar mechanisms of actions, directly or indirectly in plant growth promotion and boosting crop immunity (Orozco-Mosqueda et al. 2018). The association of beneficial microbes with the host plants is more advantageous in a suitable host plant. Microbial interactions benefit plants in many ways as they enhance or promote the growth of plants and improve crop yields, control of phytopathogens, mobilization of soil nutrients through mineralization, and solubilization (Ojuederie and Babalola 2017). The microbial endophytes attributed with plant growth-promoting potentials such as siderophore production for iron chelation, fixing of atmospheric nitrogen, phosphate solubilization, growth hormones (auxins, cytokinin, gibberellin, etc.) stimulation, pathogen suppression, tolerance to environmental stresses, secretion of secondary metabolites, and antimicrobial sources have been well documented (Patel et al. 2018; Ojuederie et al. 2019; Fadiji and Babalola 2020a).
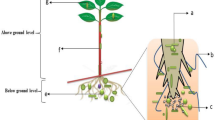
It is not only in the roots, stem, shoot, flowers, and leaves of plants that endophytes can be found, the seeds could as well represent an important source of endophytic microbes, most especially in rice and sunflower plants (Banik et al. 2016). Endophytes abruptly establish a microbial community with the roots and the shoots of the rice, also the sunflower during the growing stage, and this has shown the competence of the rice and sunflower roots in terms of colonization, growth, and survival and transmission to the next generation hosts (Jordaan et al. 2019). The proximity of microbial endophytes in the seeds to the shoot has been reported to be closer than to the roots (Alawiye and Babalola 2019). The adaption of the microbial endophyte community commonly influences their response to the prevailing environmental conditions of the host plant as well as the entire community structure inhabiting the different host plants (Fig. 1).

Plant growth-promoting microbes, effects of environmental factors, and plant nutrition
Different findings and reports have been documented on the activities of isolated and indigenous rhizobacteria and the endophytic community with the host plant (Enebe and Babalola 2018, Omomowo and Babalola 2019). However, little or no information is available on the exploration and harnessing of tiny-big unseen endophytic microbes and their resources in various agricultural applications. It is, therefore, imperative to carry out an overview of the endosphere concept inhabiting the plant roots, seeds, and other parts of the plant by highlighting their significant roles in plant growth promotion for improved agricultural production and sustainability, which is the main focus of this review. The authors believe that more than 40% of microbes in the plant endosphere could possess plant growth-promoting novel genes, and harnessing them would be of great promise in agriculture. Therefore, the understanding of endophytic microbial community structures as contained in this review revealed their possible exploitation potential and applications in developing agriculturally friendly systems in the formulation of biological or microbial-based products (biofertilizers) as an alternative to agrochemicals in enhancing soil fertility for improved crop productivity thus form the principal goal of the study.
The endosphere
The microorganisms inhabiting the internal tissue of plants are referred to as endophytes. If their interactions with the host plant are beneficial, without causing disease symptoms, viz. a viz., their presence does not cause harm to the plants, but instead they help in the control of pathogens and promote plant growth, they can be regarded as plant growth promoters (Hong et al. 2018).
The microbial interactions with the plant, their beneficial roles in promoting plant growth and health are still largely yet to be explored, this has allowed researchers to find out more about endophytes novel attributes, making it an interesting area of study from complex and diverse plant species for various application in medicine, food, and agriculture (Audipudi et al. 2017; De Silva et al. 2019). The use of a culture-independent method in the identification of endophytes using 16S rRNA amplicons sequencing is known. Specifically, the study carried out by Bertani et al. (2016) reported on the isolation of an assumed total number of 318 of endophytes from plant organs confirmed that the isolates displayed plant growth-promoting ability.
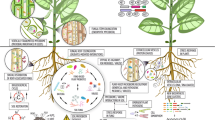
The microbial interactions with the host plants can be mutual, parasitic, symbiotic, or antagonistic. Microorganisms are found inhabiting various ecological niches in plants such as the rhizoplane, rhizosphere, endosphere, phyllosphere, and atmosphere, thus benefiting the host plant (Yang et al. 2018). Recent research findings focusing on the isolation of different microorganisms from various ecological niches have thrown more light on the understanding of microbes, including those inhabiting the interior part of the tissues of the plants (Omomowo and Babalola 2019). The internal tissues of the plant are known to be sterile without contaminants, but they contain beneficial microorganisms that stabilize the internal environment as well as contribute to the physiological functions of the plants (Soldan et al. 2019). More research is now being directed to the understanding of microbial ecology in the endosphere. In the endosphere, the microorganisms inhabiting the region confer benefits to plants rather than being pathogenic. Endosphere represents an endo-compartment within the plant tissues harboring a diverse community of microorganisms living there without developing disease symptoms or negative influence on the host plant. For instance, the microbial communities which include bacteria, archaea, or fungi shall be discussed in detail in subsequent discussion. However, some examples of microbial domains previously isolated from the endosphere compartments of the respective commercial crop are represented (Table 1).
Consequently, the presence of microbial domains within the plant tissues enables them to elicit their biocontrol potential in the control of plant pathogens that affect crop yield (De Silva et al. 2019). Besides, they enable the plant to withstand or tolerate environmental stresses, and to synthesize biologically active metabolites, needed for plant growth. Hence, the plants internally colonized microbes confer benefits, and secretion of metabolites needed for microbial growth in contributing to the general plant growth is referred to as the “endosphere effect.” The group of microbes found in the plants that elicit their plant growth-promoting effect and synthesis of bioactive agents in the control of plant pathogens is classified as PGP endophytes (PGPE) (Soldan et al. 2019).
On the other hand, plants growing on the soil have diverse microbial communities colonizing them. However, the available data on the endosphere community is still small when compared to the success that has been recorded in the study of rhizosphere microbial counterparts. The endosphere is the internal tissue of plants colonized by microorganisms without showing any pathological or severe effects on the host plant, and the microorganisms inhabiting this region are called endophytes (Verma and White 2018; Fadiji and Babalola 2020a).
Endophytes get into the plant via vertical means by seed inoculation and horizontally from the rhizosphere into the plant roots through signals intrusion (Santoyo et al. 2016; Omomowo and Babalola 2019). These two measures enable microbes to establish or colonize the host plant. Endophytes stand as a promising candidate in maintaining host defense mechanisms against foreign pathogens. According to the literature on the importance of rhizosphere microbes (Enebe and Babalola 2018; Alawiye and Babalola 2019; Odelade and Babalola 2019), plant endophytes equally share similar potential in supplying nutrients to plants by nitrogen fixation, phosphate solubilization, protect the plants from foreign pathogens by secretion of essential metabolites compounds, enhance plant tolerance to environmental stresses by induced systemic resistance (ISR), modulate the physiological functions of plants, and synthesize biopesticides in the control of crop pests (Verma and White 2018). Research insights into the herbicidal activities of endophytes are under study.
Endosphere bacteria community
The large community of bacteria colonizing the inner tissue of plants predominantly defends plants against environmental stresses. They elicit strong defense mechanisms in response to the various factors linked with plant growth, survival, and health (Omomowo and Babalola 2019). The microorganisms penetrating through the soil infiltrated into the plant roots with unique biological functions in sustainable agriculture. Recruitment and exploration of soil-endophytes for their ecological and agricultural importance have recently been reported (Fadiji and Babalola 2020b). The endo-compartment is an environment suitable only for microorganisms with colonizing potential benefits the plant. How endophytes colonize the internal tissue of plants in large numbers and elicit defense mechanisms by protecting plants is a question that has not been answered. Understanding mechanisms by which microbe-microbe and plant-microbe interactions would be promising in exploring their agricultural and biotechnological importance. In literature, it has been reported that the ability of microbes to elicit their defense responses in developing a healthy disease-free plant for sustainability depends on their communication pathways, though the actual mechanisms undergone by root endophytes in promoting plant growth have not been clearly defined (Bamisile et al. 2018).
The colonization of the narrow space by endophytes within the plant tissues as well as colonization of endophyte-like pathogens make it suitable in controlling the plant pathogens through the niche exclusion and synthesis of antimicrobials (Fadiji and Babalola 2020a). The greater the number of active microorganisms, the more effective they are in the environment via various microbial activities beneficial to plants through the following processes: Plant growth promotion by the production of plant growth factors, such as growth hormones (e.g., auxin, gibberellin, and cytokinins), suppression of biosynthetic environmental stress-related plant hormones such as ethylene under stress, bio-fixation of atmospheric nitrogen, potassium mineralization, phosphate solubilization and a draft of systemic induced plant resistance (Igiehon and Babalola 2017). The need for plant-microbe interactions is essential in establishing a mutual relationship in supporting plant growth. In an ecosystem, the plants shield and secrete exudates that supply nutrients to the microorganisms while the microorganisms produce growth stimulators and biocontrol agents needed for plant growth and control of phytopathogens for improved crop productivity (Olanrewaju et al. 2019).
Endophytes bacteria are the common microbial domains found associating with plants, stimulating plant growth, and conferring stress tolerance, owing to their firm establishment with the host plant. The allelopathic actions of endophytic bacteria that improve plant growth have been reported (Polyak and Sukcharevich 2019). Studies on bacterial endophytes characterization from different parts of plants, which include roots, seeds, stems, leaves, and flowers have been reported (Tyc et al. 2020; Wolfe and Ballhorn 2020; Xie et al. 2020) (Table 2). The bacteria population is more pronounced in the root zone than the aerial parts of the plant above the ground level i.e., flowers, stems, and leaves. The screening of plant growth-promoting bacterial endophytes isolated from crop plants, which include maize, rice, tomato, and other essential crops, has been well documented (Liu et al. 2020; Zhou et al. 2020).
The exploration of bacterial endophyte resources in the formulation of bio-inoculants and application in agriculture would help sustainably in achieving the desired agriculture productivity. The endophytic bacteria species isolated from various plants which include Bradyrhizobium, Enterobacter, Pseudomonas (rice—Oryza sativa), Pantoea, Pseudomonas, Enterobacter (sugar cane—Saccharum officinarum), Bacillus (millet—Pennisetum glaucum and Citrus spp.), and Curtobacterium (soybean—Glycine max) that promote plant growth, induced plant tolerance to abiotic stress, stimulation of biocontrol agents against plant pathogens in controlling their infestation have been reported (De Silva et al. 2019; Sheirdil et al. 2019; Shen et al. 2019).
In the endosphere, a large population of microorganisms interacts with each other, most notably the bacteria species, and till today, little information has been documented about their isolation. In contrast, some have not been cultured at all. The culturable bacterial endophytes are suspected to be Gram-positive and Gram-negative (Alawiye and Babalola 2019). Studies on the bacterial endophytes from various plants have been successfully carried out by many researchers, which include Pantoea sp., Kosakonia sacchari, Herbaspirillum sp., Enterobacter ludwigii EnVs6, Brevibacillus spp., Enterobacter spp. strain 638, Bacillus megaterium, Ralstonia spp., Pantoea anamatis, Azoarcus spp. BH72, Azoarcus spp. CIB, Azospirillum spp. B510, Burkholderia phytofirmans PsJN, Enterobacter sp. Sa187, E. ludwigii EnVs2, E. ludwigii. LecVs2, Erwinia sp. ErVv1, Pantoea vagans PaVv1, P. vagans PaVv7, P. vagans PaVv9, Gluconacetobacter diazotrophicus Pal5 A, Herbaspirillum seropedicae SmR1, Kibdelosporangium phytohabitans KLBMP 1111(T), Klebsiella pneumoniae 342, Methylobacterium extorquens DSM 1360, Pseudomonas ananatis S6, P. ananatis S7, P. ananatis S8, Pseudomonas fluorescens WH6, Pseudomonas putida W619, P. putida TJI51, Staphyloccous proteamaculans 568, Stenotrophomonas maltophilia R551-3, Staphylococcus epidermidis SE4.6, S. epidermidis SE4.7, Staphylococcus epidermidis SE2.9, S. epidermidis SE4.8, and Variovorax paradoxus S110 from the phylum: Proteobacteria, Alphaproteobacteria, and Firmicutes; class: Betaproteobacteria, Gammaproteobacteria, Actinobacteria, Cyanobacteria, and Bacilli; order: Rhodocyclales, Rhodocyclales, Rhodospirillales, Burkholderiales, Enterobacteriales, Pseudonocardiales, Nostocales, and Bacillales (Liotti et al. 2018; Pinto-Carbó et al. 2018; Eke et al. 2019). The bacterial community in the endosphere thrives well, due to low competition that occurs in the zone with other microbes.
Endosphere fungi community
The fungi form a close mutual (positive) or pathogenic (negative) association with plants. The mycorrhizal association between fungi and bacteria makes their firm interactions with the plant contributing to the physiological functions of the plants (Verzeaux et al. 2017). The mutual coexistence between fungi and the host plant roots enables them to utilize nutrients secreted in the root zone as well as securing the microbes from harsh environmental conditions. In turn, the microbes enhance the plant growth by stimulating plant growth hormones (auxin, cytokinin, gibberellin, etc.), synthesis of metabolites (enzymes and antibiotics), production of siderophores, and stimulating the plant to induce systemic resistance (Odelade and Babalola 2019).
The beneficial fungi domains colonizing the roots, stems, flowers, and leaves of many plants contribute maximally to their growth and improve crop yields. The ability of fungal endophytes secreting antimicrobial compounds confers much resistance on plants against any form of pest or pathogen attack (Segaran and Sathiavelu 2019). Fungal endophyte use as a biocontrol agent is achieved by breaking the intracellular metabolism of most pests at the early stage of their growth, thereby affecting their respiration, reproduction, as well as cell development that lead to their death. Also, the use of entomopathogenic fungi in the control of plant pests is known (Jaber and Ownley 2018). Furthermore, the mycotoxigenic fungi secrete certain metabolites that weaken the attack of pests on plants due to the toxic effect on the pests. Fungi endophytes secrete biologically active compounds that enable the host plant to withstand environmental stresses and improve their growth. They as well produce certain plant growth stimulators, such as growth hormones that confer growth and health benefits to the plants. Some of these hormonal compounds include indo-acetic acid, gibberellic acid, and piperine (Cheng et al. 2019).
Concerning the aforementioned relevance of endophytic fungi, their significant roles in the ecosystem cannot be overemphasized. Endophytic fungi directly or indirectly maintain interaction with the host plant by making nutrients available for plant growth, exhibiting fungicidal effects on the plant pathogens, and their systemic-induced resistance enables plants to tolerate high-salinity environments (Ali et al. 2019). Most endophytes produce enzymes needed in the biodegradation of pollutants in the environment, though not many findings have been documented in this regard. The use of fungal endophytes as a source of bio-inoculants in the formulation biofertilizers is known as this has tremendously contributed to achieving the desired sustainable agriculture through their application in improving crop yield, and they are less toxic to living organisms (Maheshwari et al. 2019). Similarly, fungal endophytes confer natural immunity to the host plant, boosting their adaptations and survival in a stressed or disturbed environment in fighting against phytopathogens (Omomowo and Babalola 2019). The potential application of endophytic fungi is becoming a hub of research interest by many researchers; their invaluable incorporation in sustainable agricultural and industrial biotechnological applications is promising in modern agriculture. The attractive features of these microbes are that they can easily harness to formulate bioinoculants and applications without negative environmental impacts.
Fungal endophytes resources, if adequately harnessed, would make a turnaround in agro-industry in replacing chemical fertilizers as they naturally synthesize essential compounds that promote plant growth (Maheshwari et al. 2019). The growth of fungi in an acidic environment can be a determinant in selecting their genetic traits in improving agricultural production. Fungal endophytes, which include Aspergillus oryzae, Aspergillus awamori, Aspergillus fumigatus, Paecilomyces variotii, Penicillium purpurogenum, Penicillium aurantiogriseum, Alternaria alternate, Piriformospora indica, Fusarium proliferatum, Porostereum spadiceumm that stimulate plant growth promotion and increase plant tolerance against environmental stress have been reported (Ali et al. 2019). The agricultural relevance of fungal endophytes is shown in Table 3.
Endosphere archaeal community
The endosphere archaeal community is the third microbial domain to be studied in this review. The endosphere archaeal community mostly populates the soil, and internal tissues of plants, as many studies have validated this (Moissl-Eichinger et al. 2018; Zhang et al. 2018a; Watanabe et al. 2020), but documentation about archaea is still lacking. The current research into various microbial domains in the internal tissues of the plant continues to receive more attention by researchers since their significant role in an ecosystem is huge.
Naturally, archaea form a close association with the soil and plant root environment as their growth usually influenced by soil and plant types as well as other pathogens (Moissl-Eichinger et al. 2018). The intense research into archaea, as well as their recruitment, would be similar to bacteria and fungi, although their isolation is quite challenging. In this review, we hypothesized that the large biomass of archaea community in the soil might be due to exudates released from the plant roots which encourage them to build a biofilm in the plant root zones, although this is based on speculation because little or no findings have established their colonization, diversity, functions, and symbiotic interactions with the host plant. Some of the archaea examples include Methanobacterium thermoautotrophicum Methanococcus jannaschii, Nitrosopumilus maritimus, Haloferax sulfurifontis, Haloferax prahovense, Halogranum gelatinilyticum, Methanoregula boonei, Methanosphaerula palustris, Nitrosopumilus, Nitrososphaera, and Nitrosocosmicus oleophilus MY3 (Song et al. 2019).
The archaea community in the endosphere of many plants remains abundant and yet unexplored. Archaea contribute to plant growth promotion owing to their ability to synthesize growth-promoting stimulators, and biocontrol agents in the control of plant pathogens as well as enriching soil nutrients (Odelade and Babalola 2019). Endosphere archaea could serve as a premise to rhizospheric archaea studies because their activities positively influence plant growth physiological fitness. The archaea community in the endosphere can exert beneficial effects on host plant by (a) producing plant hormones (auxin, gibberellins, cytokinins, etc.), (b) enhancing plant nutrition by nitrogen fixation, iron chelation, phosphate solubilization, potassium mineralization, etc., (c) synthesis of antimicrobials as biocontrol agents against phytopathogens, and (d) inducing systemic resistance in the response of the plant to withstand various ecological stresses (Moissl-Eichinger et al. 2018; Odelade and Babalola 2019).
Endophytic archaea participate in the nitrification process that converts ammonia to nitrate in the presence of ammonia monooxygenase, though the mechanism of this is still not well understood. Then, ammonia-oxidizing archaea (AOA) and ammonia-oxidizing bacteria (AOB) could be promising in the biodegradation of complex molecules due to their oxidizing potential. Archaea occupy up to 20% of the world biomass, yet the understanding of the archaea community in the terrestrial habitat is still inadequate, thus limiting their exploration for agricultural use. From literature, little documentation is available on the biotechnological and agricultural importance of archaea. Archaea have the potential to add benefits to plants such as the halo-archaea and methanogenic archaea groups that exhibit similar functions to plant growth-promoting rhizobacteria in producing siderophore, antibiotics, plant hormones (auxin, gibberellins, cytokinins, etc.), solubilization of phosphorus, iron chelation, mineralization of potassium, and nitrogen fixation (Odelade and Babalola 2019).
For the past few decades, the study of archaea has taken a new dimension in understanding their diversity in various ecological functions. Archaea occupy a major habitat in the biosphere, though till now, little information is available on their environmental impacts and functions. Archaea are distributed across various ecospheres within the plants as they are found inhabiting the caulosphere, laminosphere, rhizosphere and endosphere, although they are scarcely found in the phyllosphere. The colonization, distribution, and adaptation of endophytic archaea in a microenvironment are affected by various environmental factors such as energy stress and pathogen competition that have caused a great shift in the microniche. Despite the growth of archaea under different environmental conditions, which has been experimented (Doni et al. 2019), how they interact with plants by eliciting their functional and ecological roles still needs to be researched because it is still unclear.
The isolation of archaea phylum, Crenarchaeota from the roots of leguminous plants has been reported (Yadav et al. 2019), though its further exploration in developing biological products and applications in agriculture is still lacking. Studies on the isolation of AOA, Nitrosocosmicus loeophilus MY3 from the rhizosphere soil of Arabidopsis thaliana as well as other ammonia-oxidizing bacteria showed that their interactions stimulate plant growth and induced systemic resistance against plant pathogens (Jung et al. 2016). The plant-growth-promoting ability and biocontrol potential of archaea Nitrosocosmicus loeophilus MY3 against bacterial pathogens, Pectobacterium carotovorum, and Pseudomonas syringae have been reported (Song et al. 2019). Based on the phylogenetic comparison, archaea are classified as Crenarchaeota mostly hyperthermophilic-sulfur utilizers, Euryarchaeota (methanogens), and halophiles. The type of environmental samples collected determines the archaea activities and gene expression. The phylogenetic trees showing the genetic correspondence of archaea genome depicts its similarity to that of Gram-positive bacteria, thus making antibiotic option easier since archaea are resistant to many antibiotics synthesized by Gram-positive bacteria (Samson et al. 2019).
The characterization of archaea belonging to phyla halo-archaea, family halo-bacteriaceae from the hypersaline environment and mesophilic archaea from swamps, soils, oceans, marshlands, sewage, oceans, the intestinal tract of animals and decomposed organic matter has been reported (Hassani et al. 2018), and their ability to colonize the plant-root region contributes to plant growth for agricultural sustainability. The contribution of archaea to plant growth can be monitored under field experiments. The direct isolation of archaea by convectional culture-dependent methods from environmental samples is tedious, time-consuming, and difficult to cultivate.
Since the culturing of archaea is difficult and complex, the mechanisms of their interactions with plants, and pathogenic nature are not currently known, which may be due to inefficient research, hence the need to acquire knowledge about archaea domains is important. Based on general microbiology knowledge that microorganisms are present everywhere (ubiquitous), found associated with healthy plants, we can say that for this reason archaea interaction with the plant is specie-specific with a positive effect based on their important functions in promoting plant growth, improving crop yields, adaptation to ecological stress, plant diversification due to nutrients absorption at the same time soil health. In the root endosphere, archaea closely interact with other microbes. Therefore, the use of track methods can be employed in determining their phenotype and genotype, as well as physiological functions, which include solubilization of mineral elements, nitrogen fixation, plant hormone, antibiotic syntheses, and production of chelating agents (Hassani et al. 2018).
Many research findings on the identification of PGPM (bacteria and fungi) by both conventional and molecular methods have been documented (Fadiji and Babalola 2020b; Samson et al. 2019). Recently, the use of shotgun metagenomics high throughput sequencing techniques in the analysis of plant-soil microbial community interestingly has enabled in the identification of total microbes present, particularly unidentified archaea with novel genes or traits in harnessing their biological resources in amending soil, formulation of biofertilizers in ensuring agricultural sustainability. The bioremediation of xenobiotic substances such as non-biodegradable pollutants in the soil by soil bacteria has been documented (Ayangbenro et al. 2019). Nevertheless, few reports are available on the use of bacteria in an engineered environment, while no documentation has been reported on archaea. The synergistic actions between archaea with other microbes in the rhizosphere and endosphere enhance the plant’s efficiency in achieving various environmental tasks. The study of the microbial community in the endosphere is not limited to determining only the microbial type or diversity but notably used in identifying novel traits applicable in agriculture, medicine, and industries. To date, the knowledge regarding the archaea community inhabiting the endo-rhizosphere is limited; hence, this limits their full exploration for various agricultural applications. Therefore, employing a metagenomics approach in the study of the archaeal community from environmental samples would provide researchers a fast approach method in detecting their novel genes or traits for specific functions in promoting plant growth, thus improved agricultural productivity sustainably.
Factors influencing microbial diversity in the endosphere
The colonization, abundance, and diversity of microorganisms in the endo-rhizosphere regions are affected by many factors (physical, chemical, and biological). The key attributes of environmental factors influencing the survival of microbial endophytes are represented (Table 4). The physical factors include the temperature, soil pH, light intensity, geographical locations, time, water availability (rainfall), chemical factors including exudates, mineral elements, other organic substances, and the biological factors including pathogens, nematodes, viruses, protozoans, and beneficial plant growth promoter (Ojuederie et al. 2019). All these factors positively or negatively influence the survival of microorganisms in the endosphere. The adaptation strategies of microbes to strive well in an econiche depend on their ability to tolerate or resist environmental stresses (Enebe and Babalola 2018).
The erratic rainfall, abnormal high temperatures leading to the release of toxic gasses (CO2) to the atmosphere due to human activities have resulted in the greenhouse effect causing global warming, and this affects the ecosystem, hence, making it unfriendly for both plants and microorganisms to inhabit. As stated in the previous endosphere bacteria community that rhizosphere microbes form a premise in establishing endosphere community in the plant roots because of the high rhizodeposits and exudation secretion containing nutrients which provide nutrient for the microorganism to stimulate growth factors that enhance plant growth and health. The availability of abundant organic substrates can influence the abundance and performance of diverse endophytic microbes in the root zone, unlike endophytes found in the leaves and stems that are affected by physical factors (Li et al. 2019).
Moreover, soil mineral composition such as iron, phosphorus, and nitrogen affect the microbial population found in the rhizosphere, thus causing a change in the microbial community both outside and in the internal root environment. The nitrogen fixer, as well as phosphate solubilizer microbes, make nutrients more abundant in the soil by enhancing nitrogen, potassium and phosphorus uptake by the resident roots microflora, in response to the improvement in crop yield (Maheshwari et al. 2019; Ramakrishna et al. 2019). The microbial population in the endosphere often stems from the growing plant seeds or rhizoplane or bulk soil and strives under different environmental conditions. Factors, such as plant age, type, soil pH, plant developmental stages, excess or limited organic compounds, and nutrient availability, are known to influence the microbial competence and activities in the root zone (Shymanovich and Faeth 2019). It has been affirmed that the plant growth stage, soil physicochemical parameters, and plant species attest to be the principal factors influencing the rhizosphere and endosphere microbial communities (Ojuederie et al. 2019).
The roles of PGPR and endophytes
The biotransformation of organic residue in our environment is subjected to environmental treatment using microorganisms. Plant decomposition due to the activities of microorganisms influences the microbial community. The difference in the microbial community results in changes in the soil composition and characteristics. The ability of PGPR to grow under harsh environmental conditions buttress their potential in subverting the effects of environmental stress on crop yields (Enebe and Babalola 2018; Ojuederie et al. 2019). Under harsh conditions, plant growth and crop productivity are affected by a reduction in the crop yield. One example of soil bacteria, Bacillus spp. has been used in the formulation of biopesticides and promotes plant growth factors. The application of biopesticide formulated from soil microbe Bacillus thuriengensis, Bacillus sphaericus, etc., has been extensively used in agriculture in the control of phytopathogens and pesticides, due to its antimicrobial secondary metabolites produced, as the best alternative to chemical pesticides in disease prevention program (Jiao et al. 2020).
However, since there is a potential of unseen allies with vast importance in soil, exploring their resources can resolve various agricultural challenges through bioproducts production, such as biofertilizers and biopesticides, as an alternative to the use of agrochemicals (Igiehon and Babalola 2017). The growth of a plant in ecological niches can be said to be the function of plant-microbes interactions mediating vital responses allying the plant root as well as the soil environment. The beneficial microorganisms can stimulate plant growth by making nutrients available to plants through nitrogen fixation, phosphate solubilization, and potassium mineralization. All these attributes are cumulated for better crop productivity (Omomowo and Babalola 2019).
Microbes are classified based on their responsiveness to plant growth (Table 5). The microbes involved in direct association with soil or plant root-like Rhizobium partake in plant nutrition by supplying essential soluble nutrients, in-plant growth by phytohormones production, and many studies have reported this (Duan et al. 2019). The use of active microorganisms in the formulation of biofertilizer as sole bio-inoculants in replacement of chemical fertilizers in promoting plant growth and improving crop yields for better agricultural productivity has been reported (Igiehon and Babalola 2017; Rostami and Azhdarpoor 2019).
Some groups of microorganisms can function neutrally but remain in the soil; as such, they could either be classified as beneficial or saprophytic. The neutral saprophytic microbes that are involved in the degradation of organic material in the soil, ensuring nutrients mineralization seem not to harm or benefit plants but transform soil structural microbial dynamics, thereby influencing plant growth, health, and crop productivity (Arora et al. 2019). The free-living bacteria, archaea, and fungi exerting beneficial effects on the plant for improved crop production, contributing to sustainable agriculture are referred to as beneficial plant growth-promoting microorganisms (PGPM). Due to advancements in the taxonomic classification of bacteria, studies have shown that some bacteria stimulating plant growth are referred to as PGP bacteria, most especially the rhizobacteria (Sadeghi et al. 2020). In the present era, interest has been drawn towards PGPR by grouping them into different taxa according to their nomenclature and mode of actions. The use of PGPR in the biofertilization of farm crops has yielded successes by inoculation. Research findings have identified PGPR as classified into various genera, which include Stenotrophomonas, Azotobacter, Azospirillium, Streptomyces, Bacillus, Pseudomonas, Rhizobium, and Serratia. Similarly, some fungi genera such as Trichoderma, Coniothyrium, and Ampelomyces have been reported to be beneficial to the host plant (Sadeghi et al. 2020).
The growth-promoting mechanisms of PGPR encompass multiple mechanisms in enhancing plant protection for higher productivity. The major benefits of PGPR to plant growth can be achieved, firstly; by biofertilization in making soil nutrients readily available to plants; secondly; phytostimulation that involves the synthesis of plant growth hormones (auxin, gibberellin, cytokinins, etc); thirdly; biocontrol of phytopathogens through the synthesis of secondary metabolites possessing antimicrobial properties, and lastly; induction of systemic resistance by stimulating defense responses in plants against environmental stress. Bacteria genera Pseudomonas and Bacillus have been reported as the most abundant PGPR found in the rhizosphere (Xu et al. 2019b).
Conclusion and prospects
In this review, various endophytic microbial domains were studied with more emphasis on their roles in ecosystems. Evidence has shown that bacterial and fungal endophytes colonize above (leaves, stems, flowers, flowers, and seeds) and below (root/soil) of plant zones. Most rhizosphere bacteria and mycorrhiza infiltrating the root tissues can be regarded as root endophytes. Their presence is said to mediate various biological functions and physiological processes in the host plants. The roles of plant endophytes cannot be overemphasized as some of them have been identified and displayed their relevance in agricultural, biotechnological, and pharmaceutical industries. Better still, their significance in agriculture is promising but yet to be fully explored.
The coexistence of various microorganisms in the endosphere could either be beneficial or pathogenic depending on the prevailing environmental factors. Plant age, type, developmental stages, soil pH, soil nutrients, and presence or absence of water/moisture, temperature, and pathogens form major factors influencing the activities of endophytes in the host plants. However, the important roles of endophytes in agricultural crop productivity by secretion of certain metabolites could facilitate their various activities in promoting plant growth through phytohormones synthesis, siderophores production, and secretion of antimicrobial substances, such as antibiotics, ammonia, cyanide, and other volatile compounds, that suppress activities of nematodes, protozoans, and viral, bacterial, and fungal pathogens on plants, thus improving crop productivity.
Many studies have been carried out on plant-root associated microbes, but harnessing their potential use in agriculture is still limited. Findings have equally established that endophytes possessing growth-promoting traits enhance plant growth by various biological activities, thus are promising in sustainable agricultural production. The application of endophytes has been employed in various environmental studies, such as for the removal of recalcitrant metals, decaying of organic materials, mineralization, and solubilization of soil nutrients, as well the synthesis of biocontrol agents against plant pathogens. Also, endophytes exert beneficial effects on plants by enhancing their tolerance to various environmental stresses, such as drought or salinity. Under crop improvement, harnessing endophyte potential can be employed in the formulation of biofertilizers and biopesticides, on application to improve crop yield and reduce dependence on chemical fertilizers and pesticides that pose a serious threat to ecosystems, environment, and human health.
Hence, this review provides information on the roles of hidden tiny-big microorganisms living within the tissues of plants, i.e., “microbial endophytes,” their biologically active properties in the formulation of biofertilizers, biopesticides, bioherbicides, and biostimulants to improve crop production in sustainable agriculture; as such, as an alternative in mitigating the menace posed by the use of agrochemicals to the ecosystem. Consequently, further study on how vital microbial metabolites can be extracted would enhance their potential applications in agriculture. Above all, the study of the endosphere community would revolutionize agriculture if adequately harnessed.
References
Afridi MS, Mahmood T, Salam A, Mukhtar T, Mehmood S, Ali J, Khatoon Z, Bibi M, Javed MT, Sultan T (2019) Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: involvement of ACC deaminase and antioxidant enzymes. Plant Physiol Biochem 139:569–577
Alawiye TT, Babalola OO (2019) Bacterial diversity and community structure in typical plant rhizosphere. Diversity 11:179
Ali S, Khan SA, Hamayun M, Iqbal A, Khan AL, Hussain A, Shah M (2019) Endophytic fungi from Caralluma acutangula can secrete plant growth promoting enzymes. Fresenius Environ Bull 28:2688–2696
Aloo BN, Makumba BA, Mbega ER (2019) The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol Res 219:26–39. https://doi.org/10.1016/j.micres.2018.10.011
Arora P, Wani ZA, Ahmad T, Sultan P, Gupta S, Riyaz-Ul-Hassan S (2019) Community structure, spatial distribution, diversity and functional characterization of culturable endophytic fungi associated with Glycyrrhiza glabra L. Fungal Biol 123:373–383. https://doi.org/10.1016/j.funbio.2019.02.003
Audipudi A, Chakicherla B, Bhore S (2017) Bacterial endophytes as biofertilizers and biocontrol agents for sustainable agriculture. Biotechnol Sustain 1:223–247
Ayangbenro AS, Babalola OO, Aremu OS (2019) Bioflocculant production and heavy metal sorption by metal resistant bacterial isolates from gold mining soil. Chemosphere 231:113–120. https://doi.org/10.1016/j.chemosphere.2019.05.092
Babalola OO, Fadiji AE, Ayangbenro AS (2020) Shotgun metagenomic data of root endophytic microbiome of maize (Zea mays L.). Data Brief 31:105893
Bamisile BS, Dash CK, Akutse KS, Keppanan R, Afolabi OG, Hussain M, Qasim M, Wang L (2018) Prospects of endophytic fungal entomopathogens as biocontrol and plant growth promoting agents: an insight on how artificial inoculation methods affect endophytic colonization of host plants. Microbiol Res 217:34–50. https://doi.org/10.1016/j.micres.2018.08.016
Banik A, Mukhopadhaya SK, Dangar TK (2016) Characterization of N2-fixing plant growth promoting endophytic and epiphytic bacterial community of Indian cultivated and wild rice (Oryza spp.) genotypes. Planta 243:799–812
Banik A, Dash GK, Swain P, Kumar U, Mukhopadhyay SK, Dangar TK (2019) Application of rice (Oryza sativa L.) root endophytic diazotrophic Azotobacter sp. strain Avi2 (MCC 3432) can increase rice yield under green house and field condition. Microbiol Res 219:56–65. https://doi.org/10.1016/j.micres.2018.11.004
Bargaz A, Lyamlouli K, Chtouki M, Zeroual Y, Dhiba D (2018) Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front Microbiol 9:1606
Bertani I, Abbruscato P, Piffanelli P, Subramoni S, Venturi V (2016) Rice bacterial endophytes: isolation of a collection, identification of beneficial strains and microbiome analysis. Environ Microbiol Rep 8:388–398
Bilal L, Asaf S, Hamayun M, Gul H, Iqbal A, Ullah I, Lee I-J, Hussain A (2018) Plant growth promoting endophytic fungi Asprgillus fumigatus TS1 and Fusarium proliferatum BRL1 produce gibberellins and regulates plant endogenous hormones. Symbiosis 76:117–127
Campos EVR, Proença PLF, Oliveira JL, Bakshi M, Abhilash PC, Fraceto LF (2019) Use of botanical insecticides for sustainable agriculture: future perspectives. Ecol Indic 105:483–495. https://doi.org/10.1016/j.ecolind.2018.04.038
Cheng YT, Zhang L, He SY (2019) Plant-microbe interactions facing environmental challenge. Cell Host Microbe 26:183–192. https://doi.org/10.1016/j.chom.2019.07.009
Chowdhury MEK, Bae H (2018) Bacterial endophytes isolated from mountain-cultivated ginseng (Panax ginseng Mayer) have biocontrol potential against ginseng pathogens. Biol Control 126:97–108
Comby M, Lacoste S, Baillieul F, Profizi C, Dupont J (2016) Spatial and temporal variation of cultivable communities of co-occurring endophytes and pathogens in wheat. Front Microbiol 7:403
Cui L, Yang C, Wei L, Li T, Chen X (2020) Isolation and identification of an endophytic bacteria Bacillus velezensis 8-4 exhibiting biocontrol activity against potato scab. Biol Control 141:104156
da Silva LL, Veloso TG, Manhães JH, da Silva CC, de Queiroz MV (2020) The plant organs and rhizosphere determine the common bean mycobiome. Braz J Microbiol 51:765–772
Daungfu O, Youpensuk S, Lumyong S (2019) Endophytic bacteria isolated from Citrus plants for biological control of Citrus canker in lime plants. Tropical Life Sci Res 30:73–88
de Almeida Lopes K, Carpentieri-Pipolo V, Oro T, Stefani Pagliosa E, Degrassi G (2016) Culturable endophytic bacterial communities associated with field-grown soybean. J Appl Microbiol 120:740–755
de Araujo ASF, Miranda ARL, Sousa RS, Mendes LW, Antunes JEL, Oliveira LMS, de Araujo FF, Melo VMM, Figueiredo MVB (2019) Bacterial community associated with rhizosphere of maize and cowpea in a subsequent cultivation. Appl Soil Ecol 143:26–34. https://doi.org/10.1016/j.apsoil.2019.05.019
de Fretes CE, Suryani R, Purwestri YA, Nuringtyas TR, Widianto D (2018) Diversity of endophytic bacteria in sweet sorghum (Sorghum bicolor (L.) Moench) and their potential for promoting plant growth. Ind J Sci Technol 11:200–209
De Silva NI, Brooks S, Lumyong S, Hyde KD (2019) Use of endophytes as biocontrol agents. Fungal Biol Rev 33:133–148. https://doi.org/10.1016/j.fbr.2018.10.001
Doni F, Mispan MS, Suhaimi NSM, Ishak N, Uphoff N (2019) Roles of microbes in supporting sustainable rice production using the system of rice intensification. Appl Microbiol Biotechnol 103:5131–5142
Duan C, Razavi BS, Shen G, Cui Y, Ju W, Li S, Fang L (2019) Deciphering the rhizobium inoculation effect on spatial distribution of phosphatase activity in the rhizosphere of alfalfa under copper stress. Soil Biol Biochem 137:107574. https://doi.org/10.1016/j.soilbio.2019.107574
Eke P, Kumar A, Sahu KP, Wakam LN, Sheoran N, Ashajyothi M, Patel A, Fekam FB (2019) Endophytic bacteria of desert cactus (Euphorbia trigonas Mill) confer drought tolerance and induce growth promotion in tomato (Solanum lycopersicum L.). Microbiol Res 228:126302. https://doi.org/10.1016/j.micres.2019.126302
Enebe MC, Babalola OO (2018) The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: a survival strategy. Appl Microbiol Biotechnol 102:7821–7835
Fadiji AE, Babalola OO (2020a) Elucidating mechanisms of endophytes used in plant protection and other bioactivities with multifunctional prospects. Front Bioeng Biotechnol 8:467
Fadiji AE, Babalola OO (2020b) Metagenomics methods for the study of plant-associated microbial communities: a review. J Microbiol Methods 170:105860
Fors RO, Júnior OJS, Carneiro MAC, Berbara RLL (2020) Selection of arbuscular mycorrhizal fungi for sugarcane in four soils with the presence of dark septate endophytes. Acta Sci Agron 42:42477–42477
Gond SK, Bergen MS, Torres MS, White JF Jr (2015) Endophytic Bacillus spp produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol Res 172:79–87
Gonzaga L, Costa L, Santos T, Araújo E, Queiroz M (2015) Endophytic fungi from the genus Colletotrichum are abundant in the P haseolus vulgaris and have high genetic diversity. J Appl Microbiol 118:485–496
Hassani MA, Durán P, Hacquard S (2018) Microbial interactions within the plant holobiont. Microbiome 6:58
Hong CE, Kim JU, Lee JW, Lee SW, Jo I-H (2018) Diversity of bacterial endophytes in Panax ginseng and their protective effects against pathogens. Biotechnol 8:397
Igiehon NO, Babalola OO (2017) Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Appl Microbiol Biotechnol 101:4871–4881
Jaber LR, Ownley BH (2018) Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol Control 116:36–45. https://doi.org/10.1016/j.biocontrol.2017.01.018
Jiao R, Munir S, He P, Yang H, Wu Y, Wang J, He P, Cai Y, Wang G, He Y (2020) Biocontrol potential of the endophytic Bacillus amyloliquefaciens YN201732 against tobacco powdery mildew and its growth promotion. Biol Control 143:104160
Jordaan E, van der Waals JE, McLaren NW (2019) Effect of irrigation on charcoal rot severity, yield loss and colonization of soybean and sunflower. Crop Prot 122:63–69. https://doi.org/10.1016/j.cropro.2019.04.026
Jung MY, Kim JG, Sinninghe Damsté JS, Rijpstra WIC, Madsen EL, Kim SJ, Hong H, Si OJ, Kerou M, Schleper C (2016) A hydrophobic ammonia-oxidizing archaeon of the Nitrosocosmicus clade isolated from coal tar-contaminated sediment. Environ Microbiol Rep 8:983–992
Kruasuwan W, Thamchaipenet A (2016) Diversity of culturable plant growth-promoting bacterial endophytes associated with sugarcane roots and their effect of growth by co-inoculation of diazotrophs and actinomycetes. J Plant Growth Reg 35:1074–1087
Kuźniar A, Włodarczyk K, Grządziel J, Goraj W, Gałązka A, Wolińska A (2019) Culture-independent analysis of an endophytic core microbiome in two species of wheat: Triticum aestivum L. (cv. ‘Hondia’) and the first report of microbiota in Triticum spelta L. (cv. ‘Rokosz’). Systematic Appl Microbiol 43:126025. https://doi.org/10.1016/j.syapm.2019.126025
Leff JW, Lynch RC, Kane NC, Fierer N (2017) Plant domestication and the assembly of bacterial and fungal communities associated with strains of the common sunflower, Helianthus annuus. New Phytol 214:412–423
Li C, Li H, Yao T, Su M, Ran F, Han B, Li J, Lan X, Zhang Y, Yang X, Gun S (2019) Microbial inoculation influences bacterial community succession and physicochemical characteristics during pig manure composting with corn straw. Bioresour Technol 289:121653. https://doi.org/10.1016/j.biortech.2019.121653
Lin Y, Watts DB, Kloepper JW, Torbert HA (2018) Influence of plant growth-promoting rhizobacteria on corn growth under different fertility sources. Comm Soil Sci Plant Analysis 49:1239–1255
Liotti RG, da Silva Figueiredo MI, da Silva GF, de Mendonça EAF, Soares MA (2018) Diversity of cultivable bacterial endophytes in Paullinia cupana and their potential for plant growth promotion and phytopathogen control. Microbiol Res 207:8–18
Liu Y, Yan H, Zhang X, Zhang R, Li M, Xu T, Yang F, Zheng H, Zhao J (2020) Investigating the endophytic bacterial diversity and community structures in seeds of genetically related maize (Zea mays L.) genotypes. Biotechnol 10:27
Mahanty T, Bhattacharjee S, Goswami M, Bhattacharyya P, Das B, Ghosh A, Tribedi P (2017) Biofertilizers: a potential approach for sustainable agriculture development. Environ Sci Pollut Res 24:3315–3335
Maheshwari R, Bhutani N, Bhardwaj A, Suneja P (2019) Functional diversity of cultivable endophytes from Cicer arietinum and Pisum sativum: bioprospecting their plant growth potential. Biocatal Agric Biotechnol 20:101229. https://doi.org/10.1016/j.bcab.2019.101229
Mareque C, Taulé C, Beracochea M, Battistoni F (2015) Isolation, characterization and plant growth promotion effects of putative bacterial endophytes associated with sweet sorghum (Sorghum bicolor (L) Moench). Ann Microbiol 65:1057–1067
Moissl-Eichinger C, Pausan M, Taffner J, Berg G, Bang C, Schmitz RA (2018) Archaea are interactive components of complex microbiomes. Trends Microbiol 26:70–85
Moronta-Barrios F, Gionechetti F, Pallavicini A, Marys E, Venturi V (2018) Bacterial microbiota of rice roots: 16S-based taxonomic profiling of endophytic and rhizospheric diversity, endophytes isolation and simplified endophytic community. Microorganisms 6:14
Mousa W, Schwan A, Davidson J, Strange P, Liu H, Zhou T, Auzanneau F-I, Raizada M (2015) An endophytic fungus isolated from finger millet (Eleusine coracana) produces anti-fungal natural products. Front Microbiol 6:1157. https://doi.org/10.3389/fmicb.2015.01157
Ndungu SM, Messmer MM, Ziegler D, Gamper HA, Mészáros É, Thuita M, Vanlauwe B, Frossard E, Thonar C (2018) Cowpea (Vigna unguiculata L. Walp) hosts several widespread bradyrhizobial root nodule symbionts across contrasting agro-ecological production areas in Kenya. Agric Ecosyst Environ 261:161–171
Odelade KA, Babalola OO (2019) Bacteria, fungi and archaea domains in rhizospheric soil and their effects in enhancing agricultural productivity. Int J Environ Res Public Health 16:3873
Ofek-Lalzar M, Gur Y, Ben-Moshe S, Sharon O, Kosman E, Mochli E, Sharon A (2016) Diversity of fungal endophytes in recent and ancient wheat ancestors Triticum dicoccoides and Aegilops sharonensis. FEMS Microbiol Ecol 92:152
Ojuederie OB, Babalola OO (2017) Microbial and plant-assisted bioremediation of heavy metal polluted environments: a review. Int J Environ Res Public Health 14:1504
Ojuederie OB, Olanrewaju OS, Babalola OO (2019) Plant growth promoting rhizobacterial mitigation of drought stress in crop plants: implications for sustainable agriculture. Agronomy 9:712
Olanrewaju OS, Ayangbenro AS, Glick BR, Babalola OO (2019) Plant health: feedback effect of root exudates-rhizobiome interactions. Appl Microbiol Biotechnol 103:1155–1166
Omomowo OI, Babalola OO (2019) Bacterial and fungal endophytes: tiny giants with immense beneficial potential for plant growth and sustainable agricultural productivity. Microorganisms 7:481
Orozco-Mosqueda MC, Rocha-Granados MC, Glick BR, Santoyo G (2018) Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol Res 208:25–31. https://doi.org/10.1016/j.micres.2018.01.005
Patel JK, Madaan S, Archana G (2018) Antibiotic producing endophytic Streptomyces spp. colonize above-ground plant parts and promote shoot growth in multiple healthy and pathogen-challenged cereal crops. Microbiol Res 215:36–45
Paul C, Techen A-K, Robinson JS, Helming K (2019) Rebound effects in agricultural land and soil management: review and analytical framework. J Clean Prod 227:1054–1067. https://doi.org/10.1016/j.jclepro.2019.04.115
Pelo S, Mavumengwana V, Green E (2020) Diversity and antimicrobial activity of culturable fungal endophytes in Solanum mauritianum. Int J Environ Res Public Health 17:439
Pinto-Carbó M, Gademann K, Eberl L, Carlier A (2018) Leaf nodule symbiosis: function and transmission of obligate bacterial endophytes. Curr Opinion Plant Biol 44:23–31
Polyak Y, Sukcharevich V (2019) Allelopathic interactions between plants and microorganisms in soil ecosystems. Biol Bull Rev 9:562–574
Potshangbam M, Devi SI, Sahoo D, Strobel GA (2017) Functional characterization of endophytic fungal community associated with Oryza sativa L. and Zea mays L. Front Microbiol 8:325
Ramakrishna W, Yadav R, Li K (2019) Plant growth promoting bacteria in agriculture: two sides of a coin. Appl Soil Ecol 138:10–18. https://doi.org/10.1016/j.apsoil.2019.02.019
Renuka S, Ramanujam B (2016) Fungal endophytes from maize (Zea mays L.): isolation, identification and screening against maize stem borer, Chilo partellus (Swinhoe). J Pure Appl Microbiol 10:523–529
Rojas EC, Jensen B, Jørgensen HJ, Latz MA, Esteban P, Ding Y, Collinge DB (2020) Selection of fungal endophytes with biocontrol potential against Fusarium head blight in wheat. Biol Control 144:104222
Romão-Dumaresq AS, Dourado MN, Fávaro LCL, Mendes R, Ferreira A, Araujo WL (2016) Diversity of cultivated fungi associated with conventional and transgenic sugarcane and the interaction between endophytic Trichoderma virens and the host plant. PLoS One 11:e0158974
Rostami S, Azhdarpoor A (2019) The application of plant growth regulators to improve phytoremediation of contaminated soils: a review. Chemosphere 220:818–827. https://doi.org/10.1016/j.chemosphere.2018.12.203
Russo ML, Pelizza SA, Cabello MN, Stenglein SA, Vianna MF, Scorsetti AC (2016) Endophytic fungi from selected varieties of soybean (Glycine max L. Merr.) and corn (Zea mays L.) grown in an agricultural area of Argentina. Rev Arg de Microbiol 48:154–160
Sadeghi F, Samsampour D, Seyahooei MA, Bagheri A, Soltani J (2020) Fungal endophytes alleviate drought-induced oxidative stress in mandarin (Citrus reticulata L.): toward regulating the ascorbate–glutathione cycle. Sci Hortic 261:108991
Sahu PK, Singh S, Gupta A, Singh UB, Brahmaprakash G, Saxena AK (2019) Antagonistic potential of bacterial endophytes and induction of systemic resistance against collar rot pathogen Sclerotium rolfsii in tomato. Biol Control 137:104014
Salam M, Varma A (2019) Bacterial community structure in soils contaminated with electronic waste pollutants from Delhi NCR, India. Electron J Biotechnol 41:72–80. https://doi.org/10.1016/j.ejbt.2019.07.003
Samson R, Shah M, Yadav R, Sarode P, Rajput V, Dastager SG, Dharne MS, Khairnar K (2019) Metagenomic insights to understand transient influence of Yamuna River on taxonomic and functional aspects of bacterial and archaeal communities of River Ganges. Sci Total Environ 674:288–299. https://doi.org/10.1016/j.scitotenv.2019.04.166
Sandargo B, Chepkirui C, Cheng T, Chaverra-Muñoz L, Thongbai B, Stadler M, Hüttel S (2019) Biological and chemical diversity go hand in hand: Basidiomycota as source of new pharmaceuticals and agrochemicals. Biotechnol Adv 37:107344. https://doi.org/10.1016/j.biotechadv.2019.01.011
Santoyo G, Moreno-Hagelsieb G, del Carmen Orozco-Mosqueda M, Glick BR (2016) Plant growth-promoting bacterial endophytes. Microbiol Res 183:92–99. https://doi.org/10.1016/j.micres.2015.11.008
Segaran G, Sathiavelu M (2019) Fungal endophytes: a potent biocontrol agent and a bioactive metabolites reservoir. Biocatal Agric Biotechnol 21:101284. https://doi.org/10.1016/j.bcab.2019.101284
Shaikh AA, Parmar P, Rajkumar B, Patel D, Desai H, Solanki B (2017) Bioprospecting potential of endophytic bacteria from leaves of Gossypium hirsutum. Int J Curr Microbiol Appl Sci 6:1718–1730
Sharma N, Singhvi R (2017) Effects of chemical fertilizers and pesticides on human health and environment: a review. Int J Agric Environ Biotechnol 10:675–680
Sheirdil RA, Hayat R, Zhang X-X, Abbasi NA, Ali S, Ahmed M, Khattak JZK, Ahmad S (2019) Exploring potential soil bacteria for sustainable wheat (Triticum aestivum L.) production. Sustainability 11:3361
Shen F-T, Yen J-H, Liao C-S, Chen W-C, Chao Y-T (2019) Screening of rice endophytic biofertilizers with fungicide tolerance and plant growth-promoting characteristics. Sustainability 11:1133
Shymanovich T, Faeth SH (2019) Environmental factors affect the distribution of two Epichloë fungal endophyte species inhabiting a common host grove bluegrass (Poa alsodes). Ecol Evol 9:6624–6642
Soldan R, Mapelli F, Crotti E, Schnell S, Daffonchio D, Marasco R, Fusi M, Borin S, Cardinale M (2019) Bacterial endophytes of mangrove propagules elicit early establishment of the natural host and promote growth of cereal crops under salt stress. Microbiol Res 223-225:33–43. https://doi.org/10.1016/j.micres.2019.03.008
Song GC, Im H, Jung J, Lee S, Jung MY, Rhee SK, Ryu CM (2019) Plant growth-promoting archaea trigger induced systemic resistance in Arabidopsis thaliana against Pectobacterium carotovorum and Pseudomonas syringae. Environ Microbiol 21:940–948
Sun B-T, Akutse KS, Xia X-F, Chen J-H, Ai X, Tang Y, Wang Q, Feng B-W, Goettel MS, You M-S (2018) Endophytic effects of Aspergillus oryzae on radish (Raphanus sativus) and its herbivore, Plutella xylostella. Planta 248:705–714
Thompson S, Tan Y, Neate S, Grams R, Shivas R, Lindbeck K, Aitken E (2018) Diaporthe novem isolated from sunflower (Helianthus annuus) and other crop and weed hosts in Australia. Eur J Plant Pathol 152:823–831
Tyc O, Putra R, Gols R, Harvey JA, Garbeva P (2020) The ecological role of bacterial seed endophytes associated with wild cabbage in the United Kingdom. Microbiol Open 9:e00954
Verma S, White J (2018) Indigenous endophytic seed bacteria promote seedling development and defend against fungal disease in browntop millet (Urochloa ramosa L.). J Appl Microbiol 124:764–778
Verzeaux J, Hirel B, Dubois F, Lea PJ, Tétu T (2017) Agricultural practices to improve nitrogen use efficiency through the use of arbuscular mycorrhizae: basic and agronomic aspects. Plant Sci 264:48–56. https://doi.org/10.1016/j.plantsci.2017.08.004
Vieira PDS, Motta CMS, Lima D, Torres JB, Quecine MC, Azevedo JL, Oliveira NT (2011) Endophytic fungi associated with transgenic and non-transgenic cotton. Mycol 2:91–97
Watanabe T, Asakawa S, Hayano K (2020) Long-term submergence of non-methanogenic oxic upland field soils helps to develop the methanogenic archaeal community as revealed by pot and field experiments. Pedosphere 30:62–72
Wolde-Meskel E, van Heerwaarden J, Abdulkadir B, Kassa S, Aliyi I, Degefu T, Wakweya K, Kanampiu F, Giller KE (2018) Additive yield response of chickpea (Cicer arietinum L.) to Rhizobium inoculation and phosphorus fertilizer across smallholder farms in Ethiopia. Agric Ecosyst Environ 261:144–152. https://doi.org/10.1016/j.agee.2018.01.035
Wolfe ER, Ballhorn DJ (2020) Do foliar endophytes matter in litter decomposition? Microorganisms 8:446
Xie S, Vallet M, Sun C, Kunert M, David A, Zhang X, Chen B, Lu X, Boland W, Shao Y (2020) Biocontrol potential of a novel endophytic bacterium from mulberry (Morus) tree. Front Bioeng Biotechnol 7:488
Xu T, Cao L, Zeng J, Franco CM, Yang Y, Hu X, Liu Y, Wang X, Gao Y, Bu Z (2019a) The antifungal action mode of the rice endophyte Streptomyces hygroscopicus OsiSh-2 as a potential biocontrol agent against the rice blast pathogen. Pesticide Biochem Physiol 160:58–69
Xu W, Wang F, Zhang M, Ou T, Wang R, Strobel G, Xiang Z, Zhou Z, Xie J (2019b) Diversity of cultivable endophytic bacteria in mulberry and their potential for antimicrobial and plant growth-promoting activities. Microbiol Res 229:126328. https://doi.org/10.1016/j.micres.2019.126328
Yadav AN (2018) Biodiversity and biotechnological applications of host-specific endophytic fungi for sustainable agriculture and allied sectors. Acta Sci Microbiol 1:01–05
Yadav AN, Gulati S, Sharma D, Singh RN, Rajawat MVS, Kumar R, Dey R, Pal KK, Kaushik R, Saxena AK (2019) Seasonal variations in culturable archaea and their plant growth promoting attributes to predict their role in establishment of vegetation in Rann of Kutch. Biol 74:1031–1043
Yang H, Ye W, Ma J, Zeng D, Rong Z, Xu M, Wang Y, Zheng X (2018) Endophytic fungal communities associated with field-grown soybean roots and seeds in the Huang-Huai region of China. PeerJ 6:e4713
Zhang B, Zhang J, Liu Y, Guo Y, Shi P, Wei G (2018a) Biogeography and ecological processes affecting root-associated bacterial communities in soybean fields across China. Sci Total Environ 27:20–27. https://doi.org/10.1016/j.scitotenv.2018.01.230
Zhang E-P, Tian Y, Li M, Shi M, Jiang Y, Ren R, Zhang S (2018b) Effects of various long-term fertilization regimes on soil microbial functional diversity in tomato rhizosphere soil. Acta Ecol Sin 38:5027–5036
Zhou J, Li P, Meng D, Gu Y, Zheng Z, Yin H, Zhou Q, Li J (2020) Isolation, characterization and inoculation of Cd tolerant rice endophytes and their impacts on rice under Cd contaminated environment. Environ Pollut 260:113990
Zida E, Thio I, Deleuran L, Wulff E, Lund O, Shetty P, Boelt B (2014) Fungal endophytes of sorghum in Burkina Faso: occurrence and distribution. Afr J Microbiol Res 8:3782–3793
Acknowledgments
The Ph.D. stipend of Bartholomew Saanu Adeleke (BSA) is from the National Research Foundation (NRF) of South Africa (UID: 116100). Olubukola Oluranti Babalola (OOB) acknowledges NRF for the grant (UID: 123634) that supports work in her laboratory.
Funding
This study was funded by the National Research Foundation of South Africa (UID: 123634).
Author information
Authors and Affiliations
Contributions
Both authors contributed equally.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Adeleke, B.S., Babalola, O.O. The endosphere microbial communities, a great promise in agriculture. Int Microbiol 24, 1–17 (2021). https://doi.org/10.1007/s10123-020-00140-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10123-020-00140-2