
Abstract
Regime shifts in stochastic ecosystem models are often preceded by early warning signals such as increased variance and increased autocorrelation in time series. There is considerable theoretical support for early warning signals, but there is a critical lack of field observations to test the efficacy of early warning signals at spatial and temporal scales relevant for ecosystem management. Conditional heteroskedasticity is persistent periods of high and low variance that may be a powerful leading indicator of regime shift. We evaluated conditional heteroskedasticity as an early warning indicator by applying moving window conditional heteroskedasticity tests to time series of chlorophyll-a and fish catches derived from a whole-lake experiment designed to create a regime shift. There was significant conditional heteroskedasticity at least a year prior to the regime shift in the manipulated lake but there was no significant conditional heteroskedasticity in an adjacent reference lake. Conditional heteroskedasticity was an effective leading indicator of regime shift for the ecosystem manipulation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Statistical anomalies in time series, such as increased variance and increased autocorrelation, are early warning indicators of ecosystem regime shifts (Scheffer and others 2009). The statistical properties of early warning indicators are well established by theoretical and simulation studies (for example, Carpenter and Brock 2006; van Nes and Scheffer 2007; Guttal and Jayaprakash 2008). However, there are a variety of problems associated with detecting early warnings of regime shifts in real systems including large noise disturbances, observation errors, confounding trends in external perturbations, small sample sizes, and unknown mechanisms causing regime shifts. These difficulties are typically minimized when simulated data are used to test early warning theory (Carpenter 2003 Scheffer and others 2009; Seekell and others 2011). Consequently, the efficacy of early warning indicators is unresolved because there is a critical lack of field-testing for early warning indicators, particularly at spatial and temporal scales relevant to ecosystem managers.
Conditional heteroskedasticity is persistent error variance in time series models that appears as clustered volatility (Engle 1982). This type of variance is a powerful leading indicator of regime shifts in modeled systems (Seekell and others 2011). Conditional heteroskedasticity is present when variance at one time step is dependent on variance at the previous time step. Periods of high variance follow periods of high variance and periods of low variance follow periods of low variance. Variance in stable ecosystems is typically constant, but increases prior to a regime shift (Carpenter and Brock 2006; Scheffer and others 2009). This pattern suggests conditional heteroskedasticity as an early warning indicator because the portion of a time series near an impending shift will appear as a cluster of high volatility while portions of the time series further away from the regime shift will appear as clusters of low volatility (Seekell and others 2011). Thus, there should be conditional heteroskedasticity in ecological time series prior to a regime shift, but no conditional heteroskedasticity in ecological time series without a regime shift (Seekell and others 2011).
Early warning indicators that are based on interpreting statistical patterns (for example, high vs low variance) may detect an impending regime shift when there is none (Scheffer and others 2009; Seekell and others 2011; Supplemental Material). Such false positives could result in expensive and unnecessary management action. Conditional heteroskedasticity tests are easily associated with probability values (Engle 1982; Engle and others 1985). Probability values minimize false positives by providing cut-offs for evaluating when an early warning signal is meaningful (Seekell and others 2011; Supplemental Material).
We previously documented a whole-ecosystem experimental regime shift where top predators were added to a lake to cause trophic cascades and to shift the ecosystem from dominance by planktivorous fish to dominance by piscivorous fish (Carpenter and others 2011). Trophic cascades are a common type of non-linear ecosystem regime shift, and the strong responses in system components such as phytoplankton biomass provide an opportunity for evaluating new early warning indicators (Pace and others 1999; Carpenter and others 2008; Carpenter and others 2011). Here, we evaluate conditional heteroskedasticity as a leading indicator of ecological regime shift. We use existing data from the previously documented experimental regime shift reported by Carpenter and others (2011), as well as an additional year of data acquired after that report. The purpose of our analysis was to evaluate the practicality of conditional heteroskedasticity as an early indicator using a known regime shift with high frequency data at scales relevant to ecosystem managers. We test if conditional heteroskedasticity provides early warnings well in advance of the regime shift and if these tests minimize false warnings when there is no impending regime shift.
Methods
Regime Shift Manipulation
Carpenter and others (2011) conducted a food web manipulation on Peter Lake using a second system (Paul Lake) with similar morphometry and chemistry as a reference. Prior to their experiment, Peter Lake was dominated by pumpkinseed sunfish Lepomis gibbosus, a variety of other small species of fish, and few adult (>150 mm) largemouth bass Micropterus salmoides. Paul Lake was dominated by adult largemouth bass with a small population of pumpkinseed. The food webs represent alternative stable structures of similar species composition with the consequence of piscivore dominance in Paul Lake and planktivore dominance in Peter Lake. Prior to the manipulation, Carpenter and others (2011) re-enforced the initial state of planktivore dominance in Peter Lake by adding 1,200 golden shiners Notemigonus crysoleucas on 28 May 2008. Peter Lake was manipulated over four summers (2008–2011) by adding adult largemouth bass to shift the lake to a state of piscivore dominance, similar to the reference system. Because the threshold population of largemouth bass required to fully transition the system to piscivore dominance was unknown, adult largemouth bass were added slowly (12 adult largemouth bass on 7 July 2008, 15 adult largemouth bass on 18 June 2009, and 15 adult largemouth bass on 21 July of 2009) to maximize the potential to test for early warning indicators. In response to the manipulation, the abundance of small fishes declined, zooplankton size structure shifted to larger body-sized forms, and phytoplankton biomass declined (Carpenter and others 2011). Largemouth bass produced a large year class in 2009 and many of these offspring survived the following winter to recruit into the adult largemouth bass population in 2010. This recruitment indicates the transition from planktivore to piscivores dominance in the fish community. Turbulence from this transition cascaded through the lower part of the food web until the latter part of 2010 when the entire transition was complete or nearly so. The lake stabilized in this new condition in 2011 and an additional 32 adult largemouth bass were added on 23 June 2011 to re-enforce the piscivore-dominated state. Conditions in the reference lake did not change during the study. The results of the first 3 years of the manipulation, documented by Carpenter and others (2011), were consistent with the hypothesis that disruption of the food web would lead to increased variance, increased autocorrelation, critical slowing down (that is, slower recovery from perturbations), increased skewness, non-linearity, and shifts to increased low frequency variance of key indicator variables prior to the regime shift. The results of the fourth year of the study were consistent with the manipulation system stabilizing at a new regime including decreased variance and autocorrelation (Supplemental Material).
We applied conditional heteroskedasticity tests to chlorophyll-a time series derived from this experiment because previous theoretical and empirical work suggested chlorophyll-a concentration, a measure of phytoplankton biomass, strongly reflects early warning signals of regime shifts driven by trophic cascades (Carpenter and others 2008, Carpenter and others 2011). Chlorophyll-a was determined daily in the mixed layer of both lakes (for each lake n = 105 in 2008, n = 110 in 2009, n = 110 in 2010, n = 110 in 2011) between mid-May and early September over 4 years. To measure chlorophyll-a, we took 200 ml water samples from a depth of 0.5 m from each lake and filtered the samples onto glass fiber filters. The filters were frozen and chlorophyll-a was subsequently extracted in methanol and measured with a fluorometer according to Holm-Hansen and Riemann (1978).
We also applied conditional heteroskedasticity tests to minnow trap catch time series derived from the experiment. Changes in minnow trap catch time series are driven by both changes in biomass and fish behavior. Previous theoretical and empirical work suggested that these times series display non-linear dynamics and early warning signals of regime shifts driven by trophic cascades (Carpenter and Kitchell 1993; Carpenter and others 2008, 2011). Thirty minnow traps were deployed in the littoral zone of Peter Lake and twenty minnow traps were deployed in the littoral zone of Paul Lake from late May to early September during the four study years. The traps (6 mm mesh with two 25 mm trap openings) were monitored daily (Peter Lake n = 96 in 2008, n = 108 in 2009, n = 110 in 2010, n = 111 in 2011; Paul Lake n = 95 in 2008, n = 108 in 2009, n = 110 in 2010, n = 111 in 2011) and the abundance of each species of fish collected in each trap was recorded. Time series were derived from these data by calculating the average catch per trap in each lake for each day.
Statistical Analysis
A rolling window Lagrange multiplier test for conditional heteroskedasticity was applied to the time series for each lake (Engle 1982; Engle and others 1985; Seekell and others 2011). Rolling windows are based on calculating the early warning indicator for all observations (n) from n t to n t-wl where t equals time intervals (days in our study) and wl equals window length. The calculation is iterated for each day with the result being a rolling series of conditional heteroskedasticity tests. We used a 50-day window length in this study because this was a good trade-off between statistical power (larger window widths correspond to higher statistical power) and preserving a large number of windows necessary to make meaningful interpretations of changes in indicator values and to precisely delimitate transitions (smaller window widths correspond to a larger number of windows and more precise delimitation of the timing of transitions). The Lagrange multiplier test for conditional heteroskedasticity is calculated by:
-
(1)
Fitting a time series model to the data
-
(2)
Squaring the residuals of the time series model
-
(3)
Regressing the squared residuals on themselves, lagged one time step
-
(4)
If the slope of the regression in step three is greater than 0, multiply the multiple r 2 value from step 3 by the sample size in step 3. If the slope of the regression in step 3 is 0 or less, there is no conditional heteroskedasticity. There is no concept of a negative slope in step 3.
-
(5)
Calculating a probability value by comparing the value obtained in step 4 with a Chi-square distribution with one degree of freedom.
Worked examples of the conditional heteroskedasticity test are provided in Seekell and others (2011). For display, we plot the r 2 value from step 4 instead of the Lagrange multiplier test statistic. Because each window is the same width, a critical value to assess the significance of r 2 values is obtained by dividing the critical value from the Chi-square distribution with one degree of freedom by the sample size of the auxiliary regression. We applied criteria of P less than 0.1 as the critical probability of significance in this study.
Based on prior studies of ecological models, significant conditional heteroskedasticity tests indicate an impending regime shift whereas non-significant conditional heteroskedasticity tests do not indicate an impending regime shift (Seekell and others 2011). We expected no significant conditional heteroskedasticity in Paul Lake during the study. We expected no significant conditional heteroskedasticity in Peter Lake prior to the manipulation and significant conditional heteroskedasticity as trophic cascades created turbulence in the food web as the regime shift proceeded. Based on model analyses, conditional heteroskedasticity is expected to become non-significant quickly after a regime shift (see Seekell and others 2011).
For step 1 of the analysis we applied an auto-regressive lag-4 model \( (y_{t} = b_{0} + b_{1} y_{t - 1} + b_{2} y_{t - 2} + b_{3} y_{t - 3} + b_{4} y_{t - 4} + \varepsilon ) \) to the time series and the conditional heteroskedasticity tests to the residuals of these time series models. We selected a model with four autoregressive terms because in these lakes chlorophyll-a autocorrelation and minnow trap autocorrelation is only significant at 4 or fewer lags and the partial autocorrelation is generally only significant at 2 or fewer lags. Thus in most cases, an autoregressive lag-4 model will over-fit data and such a time series model will contain more lags than necessary. Over fitting the number of autoregressive lags in the time series model will not cause the Lagrange multiplier test to perform more poorly than a correctly specified model and will actually improve performance if an important moving average term or covariate is omitted from the time series model (Lumsdaine and Ng 1999). Under fitting the time series model can adversely affect the performance of the conditional heteroskedasticity test by increasing chance of finding false positives.
Moving window conditional heteroskedasticity tests were robust to a range of window widths, time series models, and choices of threshold probability values for significance based on a sensitivity analysis (see Supplemental Material).
Results
Chlorophyll-a Time Series
Prior to the manipulation (2008), Peter and Paul Lakes had similar chlorophyll-a concentrations (Figure 1). Chlorophyll-a concentrations in Peter Lake were dynamic with substantial oscillations during the manipulation (2009 and 2010), whereas chlorophyll-a concentrations in Paul Lake were less variable (Figure 1). The fourth year of data (2011), not previously reported, was collected using the same methods as the previous 3 years to ensure that Peter Lake had stabilized at the new piscivore-dominated regime. Chlorophyll-a concentrations were low in Peter Lake and similar to Paul Lake during this year. The declining phase of spring blooms was observed in Peter Lake and perhaps in Paul Lake during the first 2 weeks of observations in 2008 and 2011.

Daily chlorophyll-a measurements (μg l−1) from the mixed layer of the manipulated Peter Lake (red) and reference Paul Lake (black) systems. Vertical dashed blue lines denote the timing of largemouth bass additions to the manipulated Peter Lake. Note the vertical axis scales are different between years for display purposes.
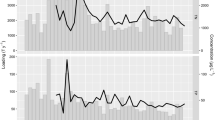
There was no significant conditional heteroskedasticity (P > 0.1) in Paul Lake during the four-summer study based on the rolling window conditional heteroskedasticity analysis (Figure 2). There was also no significant conditional heteroskedasticity in Peter Lake in 2008 during the early phase of the manipulation. There was significant conditional heteroskedasticity (P < 0.1) in Peter Lake for almost all of 2009 and the first half of 2010. Conditional heteroskedasticity became non-significant during the second half of 2010, consistent with results given by Carpenter and others (2011), indicating that the shift in food web structure to piscivore dominance had occurred. There was no significant conditional heteroskedasticity in Peter Lake during the final year (2011) after the regime shift was completed.

Rolling window (window width = 50 days) conditional heteroskedasticity tests for chlorophyll-a time series from the manipulated Peter Lake (red) and the reference Paul Lake (black). The black horizontal line represents the critical value for the conditional heteroskedasticity test. Values above the horizontal line indicate significant (P < 0.1) conditional heteroskedasticity. Values below the horizontal line indicate non-significant (P > 0.1) conditional heteroskedasticity. For display on the log10-scale, values below 0.001 have been plotted as 0.001.
Minnow Trap Catch Time Series
Prior to the manipulation (2008), Peter Lake had high minnow trap catch and Paul Lake had very low minnow trap catch (Figure 3). Minnow trap catch in Peter Lake declined after largemouth bass additions and became more variable during the manipulation (2009 and 2010). Minnow trap catches in Paul Lake were close to zero and were not variable (Figure 3). Some of the oscillations in Peter Lake trap catches, at the beginning of the summers, may be associated with increased near shore activity due to pumpkinseed spawning. Oscillations later in the summer are not consistent with increased activity due to spawning. In the fourth year (2011), minnow abundance in Peter Lake stabilized near the levels observed in Paul Lake with the exception of an increase in near shore activity in Peter Lake due to spawning. This spawning activity was interrupted by a sudden shift to cold weather and hence the near shore activity (and high catches) was only sustained for a brief period of time.

Daily minnow trap catches (catch trap−1 day−1) from the littoral zone of the manipulated Peter Lake (red) and reference Paul Lake (black) systems. Vertical dashed blue lines denote the timing of largemouth bass additions to the manipulated Peter Lake. Note the vertical axis scales are different between years for display purposes.
There was significant conditional heteroskedasticity (P < 0.1) in Peter Lake in 2008 after largemouth bass additions began (Figure 4). There was significant conditional heteroskedasticity (P < 0.1) for most of 2009, although there was not significant conditional heteroskedasticity (P > 0.1) for a period of time at the beginning of the summer when increased trap catch is likely due to spawning activity and not trophic cascades. There was no significant conditional heteroskedasticity (P > 0.1) in 2010 or 2011 with the exception of a brief period in 2011, which was due to the disruption of pumpkinseed spawning when a large and sudden shift in weather drove pumpkinseeds off of their nests. There was no significant conditional heteroskedasticity in Paul Lake in 2008, 2009, or 2011 (Figure 4). There were two significant moving windows at the beginning of 2010, but no significant conditional heteroskedasticity during the remainder of the summer.

Rolling window (window width = 50 days) conditional heteroskedasticity tests for minnow trap catches (catch trap−1 day−1) the manipulated Peter Lake (red) and the reference Paul Lake (black). The black horizontal line represents the critical value for the conditional heteroskedasticity test. Values above the horizontal line indicate significant (P < 0.1) conditional heteroskedasticity. Values below the horizontal line indicate non-significant (P > 0.1) conditional heteroskedasticity. For display on the log10-scale, values below 0.001 have been plotted as 0.001.
Discussion
Conditional heteroskedasticity was a powerful leading indicator that warned of the incipient regime shift about a year in advance. Conditional heteroskedasticity disappeared from the experimental system after the regime shift, indicating that the system arrived at a new stable state. In the chlorophyll-a time series there were no significant tests in the reference lake or in the manipulated lake in the year prior to and the year after the manipulation. In the minnow trap time series there were only two significant tests after the system had stabilized at the new state, and these significant tests were associated with a disrupted life history process (that is, spawning). There were also only two significant tests in the reference system for the chlorophyll-a and minnow trap time series, combined. These results indicate that conditional heteroskedasticity provided early warning of the regime with minimal false positives.
Time series statistics used as early warning indicators of regime shift are subject to false positives. During the early part of 2008, there was increased chlorophyll-a, and this appears similar to the oscillations observed prior to the regime shift. This pattern also occurred in 2011 and we speculate the dynamics are the consequence of phytoplankton spring blooms. More complete monitoring of these blooms was not possible for this study because difficult or impassable road conditions in the early spring limit access to the study lakes. There were no significant conditional heteroskedasticity tests during these periods. However, such blooms could lead to false positives in other indicators. For instance, the high chlorophyll-a values associated with spring blooms are followed by low values and this could increase variance in a moving window analysis (for example, Carpenter and others 2008, 2011). Such increases in variance are consistent with an impending regime shift. The conditional heteroskedasticity test’s probability values in this analysis aid interpretation by providing a baseline from which to judge the meaningfulness of indicator values.
Early warnings based on the conditional heteroskedasticity tests appeared first in the minnow trap time series. Significant tests were observed in the latter half of 2008 and for most of 2009. Significant conditional heteroskedasticity occurred in 2009 and 2010 for the chlorophyll-a times series. The earlier response associated with minnows reflects the effects of largemouth bass predation on both the abundance and behavior of these fish (Carpenter and others 2011). Upon introduction of piscivores, prey species quickly increase the occupancy of refuges and this shift in behavior contributes to trophic cascade effects (Carpenter and others 2010). The response of chlorophyll-a was more delayed and was the consequence of slower evolving shifts propagating through the food web (Carpenter and others 2011).
A recent review (Scheffer and others 2009) identified significance testing for early warning indicators as an important priority for research on this topic. Conditional heteroskedasticity tests provide a useful method for addressing this priority. An alternative approach to significance testing is to apply trend statistics, such as Kendall’s tau, to early warning indicator values calculated from moving windows (Dakos and others 2008, 2010). A significant upward trend in moving window variance or autocorrelation estimates would be considered the early warning signal using this approach (for example, Dakos and others 2008, 2010). However, moving window indicators are generally highly autocorrelated and trend statistics such as Kendall’s tau are subject to increased false positives under these conditions (Hamed and Rao 1998). Conditional heteroskedasticity as an early warning indicator is based on a sequence of significance tests, each applied to the uncorrelated residuals of a time series filter. Hence, our analysis does not calculate significance values between windows and is not subject to this potential source of error. In other words, like all moving window indicators, the sequence of conditional heteroskedasticity tests is highly autocorrelated. However, the conditional heteroskedasticity significance tests are not based on these highly correlated moving windows. The significance test for trend statistics is based on these highly correlated moving windows.
In conclusion, based on our analysis of data from a whole-lake experiment, conditional heteroskedasticity is a powerful leading indicator of ecological regime shifts that is robust to false positives. These tests have simple (that is, significant vs not significant) interpretations and were successfully applied to a system with natural environmental stochasticity with an unknown amount of observation error. Additional experience in applying the conditional heteroskedasticity approach is needed to further explore the sensitivity and robustness of these tests for a variety of regime shift conditions.
References
Carpenter SR. 2003. Regime shifts in lake ecosystems: pattern and variation. Oldendorf/Luhe: Ecology Institute. p 179p.
Carpenter SR, Brock WA. 2006. Rising variance: a leading indicator of ecological transition. Ecol Lett 9:311–18.
Carpenter SR, Kitchell JF. 1993. Simulation models of the trophic cascade: predictions and evaluations. In: Carpenter SR, Kitchell JF, Eds. The trophic cascade in lakes. New York: Cambridge University Press. p 310–31.
Carpenter SR, Brock WA, Cole JJ, Kitchell JF, Pace ML. 2008. Leading indicators of trophic cascades. Ecol Lett 11:128–38.
Carpenter SR, Cole JJ, Kitchell JF, Pace ML. 2010. Trophic cascades in lakes: lessons and prospects. In: Terbough J, Estes JA, Eds. Trophic cascades. Washington, DC: Island Press. p 59–69.
Carpenter SR, Cole JJ, Pace ML, Batt R, Brock WA, Cline T, Coloso J, Hodgson JR, Kitchell JF, Seekell DA, Smith L, Weidel B. 2011. Early warnings of regime shifts: a whole-ecosystem experiment. Science 332:1079–82.
Dakos V, Scheffer M, van Nes EH, Brovkin V, Petoukhov V, Held H. 2008. Slowing down as an early warning signal for abrupt climate change. Proc Nat Acad Sci USA 105:14308–12.
Dakos V, van Nes EH, Donangelo R, Fort H, Scheffer M. 2010. Spatial correlation as a leading indicator of catastrophic shifts. Theor Ecol 3:163–74.
Engle RF. 1982. Autoregressive conditional heteroscedasticity with estimates of the variance of United Kingdom inflation. Econometrica 50:987–1008.
Engle RF, Hendry DF, Trumble D. 1985. Small-sample properties of ARCH estimators and tests. Can J Econ 18:66–93.
Guttal V, Jayaprakash C. 2008. Changing skewness: an early warning signal of regime shifts in ecosystems. Ecol Lett 11:450–60.
Hamed KH, Rao AR. 1998. A modified Mann-Kendall trend test for autocorrelated data. J Hydrol 204:182–96.
Holm-Hansen O, Riemann B. 1978. Chlorophyll a determination: improvements in methodology. Oikos 30:438–47.
Lumsdaine RL, Ng S. 1999. Testing for ARCH in the presence of a possibly misspecified conditional mean. J Econom 93:257–79.
Pace ML, Cole JJ, Carpenter SR, Kitchell JF. 1999. Trophic cascades revealed in diverse ecosystems. Trends Ecol Evol 14:483–8.
Scheffer M, Bascompte J, Brock WA, Brovkin V, Carpenter SR, Dakos V, Held H, van Nes EH, Rietkerk M, Sugihara G. 2009. Early-warning signals for critical transitions. Nature 461:53–9.
Seekell DA, Carpenter SR, Pace ML. 2011. Conditional heteroskedasticity as a leading indicator of ecological regime shifts. Am Nat 178:442–51.
van Nes EH, Scheffer M. 2007. Slow recovery from perturbations as a generic indicator of a nearby catastrophic shift. Am Nat 169:738–47.
Acknowledgments
This work was supported by the National Science Foundation (DEB 0716869, DEB 0917696, and Graduate Research Fellowship Program) and University of Virginia Department of Environmental Sciences. We thank R. Batt, C. Brosseau, J. Cole, J. Coloso, M. Dougherty, A. Farrell, J. Hodgson, R. Johnson, J. Kitchell, S. Klobucar, J. Kurtzweil, K. Lee, T. Matthys, K. McDonnell, H. Pack, L. Smith, T. Walsworth, B. Weidel, G. Wilkinson, C. Yang, and L. Zinn for technical help in the laboratory and field.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
SRC and MLP designed the experiment; DAS, TJC, SRC, and MLP performed the experiment; DAS performed the statistical analysis; DAS wrote the manuscript with contributions from TJC, SRC, and MLP.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Seekell, D.A., Carpenter, S.R., Cline, T.J. et al. Conditional Heteroskedasticity Forecasts Regime Shift in a Whole-Ecosystem Experiment. Ecosystems 15, 741–747 (2012). https://doi.org/10.1007/s10021-012-9542-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-012-9542-2